![]()
![]()
![]()
V.M. Timon and R.P. Baber
Food and Agriculture Organization of the United Nations
Via delle Terme di Caracalla, Rome 00100, Italy
RESUME
L'opportunité d'une amélioration génétique des ovins et des caprins en zone tropicale humide d'Afrique de l'Ouest doit tout d'abord être considérée par rapport au niveau de la gestion des troupeaux, à la productivité de ceux-ci et aux problèmes posés par la trypanosomiase. Les facteurs déterminants des modifications génétiques et leur impact furent discutés dans le contexte des difficultés d'organisation et de fonctionnement des programmes conventionnels de sélection dans l'environnement prévalant en Afrique de l'Ouest.
Le rôle éventuel de programmes simples de sélection basés sur un tri dans chaque troupeau fut discuté. Un programme de sélection basé sur le poids vif à six mois fut décrit. Finalement la constitution de services de contrôle des performances et des services d'appui doit être défendue vu le fait que tout programme de sélection doit s'élaborer sur un grand nombre de générations.
SUMMARY
The article assesses the relevance of genetic improvement of sheep and goats in the humid tropics of West Africa in reference to level of management, flock productivity and trypanosomiasis. The major determinants of genetic change are then individually discussed in the context of the difficulties of implementing conventional selection schemes in the environmental conditions of West Africa. The possible role of simple selection schemes based on flock-herd screening is described, outlining a selection scheme based on six-month body weight. Finally, the need for recording, infrastructural and support services is emphasized in that selection must be practised over a number of generations.
INTRODUCTION
The importance of sheep and goats to the livelihood of peoples in the humid tropics of West Africa has been highlighted by many authors and in many fora in recent years. It is fortunate that small ruminant production appears to be increasingly attracting the interest of researchers in a number of countries in West Africa; the Small Ruminant Group at the International Livestock Centre for Africa (ILCA) as well as other international and bilateral agencies have been increasingly active in stimulating and servicing these research programmes. Also on a positive note it has been clearly shown that very considerable improvement in flock productivity at farm level can be achieved by implementing simple well-proven technologies. In north Togo, for example, it has been demonstrated (Van Vlaenderen, 1987) that as a result of improved management, simple flock health control, mineral supplementation and strategic supplementation at the end of the rainy season, ewe productivity was increased from 7.2 kg/lamb/ewe/year to 28.7 kg/lamb/ewe/year; this is a very sizable increase of about 300 percent. On the other hand the literature on the genetic improvement of sheep and/or goats in West Africa is very scant and the information that does exist, especially as regards imported breeds and cross-breeding, is not very encouraging.
Against this background it could be questioned whether any attempt to improve the genetic capacity of the sheep and goat breeds in West Africa should be made. The answer to this question is both affirmative and negative and two unrelated issues have a major bearing on the answer. These are the levels of husbandry and productivity in the target populations and secondly trypanosomiasis. Responding to the first issue, there is little point in attempting to improve any population of sheep and/or goats genetically where the level of management has not, in the first instance, sufficiently exploited the existing genetic potential of the particular breed. This unfortunately is not the situation in many of the traditional village-based flock/herds in much of West Africa. In these situations priority must therefore be given to improving the nutrition, management and health care of the animals before genetic improvement is considered. Unfortunately, many organizations and many countries have ignored this essential priority in livestock development and as a result they embarked on breed or genetic improvement programmes that were doomed to failure from the very beginning.
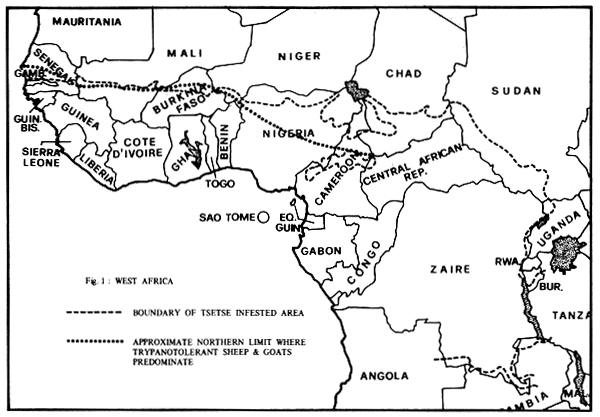
The second overriding consideration when contemplating the genetic improvement of livestock in West Africa is trypanosomiasis. The distribution of the different breeds of sheep and goats in West Africa is almost exclusively determined by the presence or absence of the tsetse fly in the region. The vast majority of sheep and goats in the areas of high tsetse challenge are the dwarf trypanotolerant breeds whereas the larger Sahelian breeds are encountered north of the tsetse belt (see Figure 1).
It can be asked why should trypanotolerance be taken into account when considering genetic improvement of sheep and goat breeds in West Africa? Ironically and sadly, it is not because the genetics of trypanotolerance are understood. It is not known how, or indeed if, the trait is controlled by simple additive or interactive allelic effects; all that is known and then loosely presumed to be genetic, is that trypanotolerance is breed-associated. This association, coupled with the many failures to establish exotic breeds successfully within the tsetse challenge area, provides sufficiently strong evidence to suggest that in the main, efforts toward the genetic improvement of sheep and goat breeds in the humid tropics of West Africa should be concentrated on selection within the local trypanotolerant breeds. Furthermore, such genetic improvement should only be considered for sheep or goat populations that are being fed and managed reasonably well, namely, well above flock/herd maintenance levels.
WITHIN-BREED SELECTION
To effect genetic change through selection the following are required:
Clearly identified, realistic and consistent objectives.
Selection pressure must be exercised and based on good evidence of the animals' genetic merit for the set objectives.
Selection must be practised year after year, generation after generation.
Figure 1: Distribution of trypanotolerant livestock
To optimize genetic gain on a unit of time basis, i.e. per annum, it is essential to ensure that the average age structure of the flock is as young as possible so as to minimize the generation interval. These well-known determinants of selection response are listed so that each of them can be examined more fully in the context of this paper - namely, realizing genetic change within the sheep and goat populations of West Africa.
Breeding objectives
Most animal breeders, and in particular breeders of ruminant animals, identify breed improvement with increased absolute levels of productivity, be that meat or milk. This rationale has emerged from the more advanced countries where input costs in animal production are high, whether these are land, labour, capital or other inputs. Regrettably, these same breeding objectives, based on the premise that ‘big is beautiful’, have been exported to many of the less advanced countries and to West Africa. There are many examples of failed attempts to achieve instant genetic improvement by importing high-producing breeds into unsuited environments. For this reason breeding objectives must be considered in reference to the major constraint(s) in the environment and in the production system in which genetic improvement is sought. In New Zealand, without question one of the best developed sheep industries in the world, breeders saw that their main limitation was the availability of labour and low market prices. They therefore did not pursue high output per animal. Rather, they bred an ‘easy care sheep’, the Perindale, that enabled them to capitalize on large flocks, high stocking and consequently high productivity per hectare, per farm and per unit of labour. This is in marked contrast with European sheep breeding, which of necessity emphasized high productivity per ewe, due to small unit size and high input cost, including labour and capital. This type of assessment should always be undertaken and breeding objectives must be anchored to the major constraint on production efficiency.
In the tropics, sheep and goats must be selected that can perform well under heat stress and cope with the seasonally available feed supply and the variation in feed quality. Animals must not be selected or evaluated under better managed medium to high feeding conditions, sometimes developed on government farms or research stations, when these conditions are not typical of the farming systems for which the animals are being bred. The classic experiment carried out by Falconer (1960), albeit with experimental animals, should be recalled at this point. His experiments showed that selection for growth rate on an ad libitum plane of nutrition increased appetite, whereas selection for the same trait, on a restricted diet, increased efficiency of feed utilization. This clearly highlights the importance of selecting animals from and within the environment in which they must perform, particularly where quality of feed or its seasonal availability are major limiting factors.
When genetic improvement within a harsh or limiting environment, such as in the tropics of West Africa, is considered, high output cannot be considered as the sole criterion of the animals' merit. Brody (1945), comparing a range of animal breeds, and indeed animal species, in terms of productivity per unit of metabolic weight, suggested that there were very small differences between big and small animals in net production efficiency. Taylor et al. (1985) have largely confirmed Brody's thesis in a set of well-controlled experiments with different breeds of cattle. Comparable experimental evidence from sheep or goats is not available but the conclusion is unlikely to be different. In other words, big is not necessarily beautiful, in the context of net production efficiency.
The importance of high yield of a particular product, e.g. milk yield, growth rate or carcass weight for age, stems from production economics in countries or systems of production where input costs are high and often where one part of the production chain is in the hands of the commercially strong section of the industry and the balance of the system is carried out by the state (subsidized feed) or by a weaker sector of the farming community. The economic thrust of this development has led to specialized single-purpose breeds.
It is doubtful whether this specialized single-purpose breed logic should be applied to sheep or goat breeding in the difficult environments of West Africa. Indeed it may well be questioned in the advanced countries at present as food mountains grow, as market bouyancy and prices are under pressure and as product to energy, feed and labour cost ratios continue to decline. The basic question at issue is the biological efficiency of multipurpose breeding to produce meat, milk, fibre (skins) and where relevant, draught animal power, as compared with the specialized production of one product, the other capacities of the animal being largely ignored. Clearly in the absence of factual data, it is not possible to reach a conclusion on this question but it is important to focus attention on the need for a much more comprehensive evaluation of breeding objectives when sheep and goat breeding strategies for West Africa are being determined, and particularly when considering difficult environments, where economic and social values are very different from those in the more industrialized countries.
The real choice of breeding objectives and selection criteria must be based on an in-depth study of the economic and other forces that influence the farmers' use of sheep and/or goats for food, for profit or capital investment. It can be argued that animals should be ranked and selected on an overall productivity index that aggregates all the outputs from the animal relative to the inputs. Peacock (1985) and a number of earlier authors have proposed and discussed such indices of productivity. While these indices have a role and usefulness in summarizing flock or herd productivity levels, they are not very useful as a basis for selection. Ideally selection must be based on an index (I) which weights each of the component traits (Xi) with a weighting (bi) that takes into account the genetic variances (α2ι) and co-variances (αμ) of the traits and their relative economic values (ai). Such an index, based on well-established phenotypic, genetic and economic parameters for the target population, does provide the optimum basis of selection in any population.
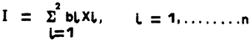
However, the possibility to operate selection programmes in sheep and/or goat flocks in West Africa on such an optimal index is limited as the data necessary to estimate the parameters do not exist. On the other hand, to aggregate the component traits into an arbitrary, although logical, productivity index, such as proposed by Peacock (1985), is equally inappropriate for genetic selection. This is because the variance of a product index, being the product of the variances of its components, will inevitably be very large and consequently the accuracy of selection will be limited.
Instead, it is suggested that simple selection criteria that identify one or two easily measured major components of productivity and production efficiency should be identified. Immediately we are limited to traits such as body size, growth rate, some measures of reproduction, milk or fibre yield. In meat-producing breeds a convenient measure of growth, e.g. a standardized weaning weight, 180-day weight or yearling weight, can be chosen. If milk production is important then obviously selection must be on some standardized measure of lactation output. It is assumed that the animals are performing and being measured in the natural environment for which they are being bred. This will automatically ensure adaptation and arguably net production efficiency. In other words, it is suggested that simple measures of performance for growth, milk or fibre production are sufficient. Given that the most appropriate selection criteria can be agreed on, then how to rank and select animals on their genetic merit must be determined.
Determining genetic merit
The basis on which to determine the genetic merit of an animal and to make accurate selection decisions has long since been established. Equally the relative accuracies of different forms of selection, be they pedigree selection, performance testing, progeny or other forms of indexed family or combined selection, are well established. As already pointed out, the absence of well-documented genetic parameter estimates makes it difficult in most situations to develop reliable selection indices for sheep or goat breeding in West Africa. However, this is not the only point to be considered. In the majority of countries in the region, the infrastructure and support services necessary to collect the records on which accurate genetic rankings can be made simply do not exist. Allied to this is an even greater problem, i.e. the very high level of phenotypic variation usually observed in these populations (Sands and McDowell, 1978). Coefficients of variation can range from 30–60 percent. This unusually big variation exacerbates the problem of estimating the genetic merit of an individual or group of individuals. An ILCA study (Peters, 1985) has shown that very large numbers of animals are required when comparing goat breeds in the tropics; for example a group size of 700–1000 animals is required to detect (one-tailed test, β = 80 percent) a difference of 5 percent if, as is often the case, the coefficient of variation lies between 35 to 40 percent. In these medium to poor nutritional environments, conventional genetic ranking procedures do not work effectively because of this large variation. McDowell (1983), analysing cattle records from a number of such environments, concluded that progeny testing, the standard orthodox method of sire evaluation, simply does not work in poor environments.
What can be done in these situations? Ironically the lack of a breed infrastructure and this very large variation may be the key to future progress - progress through population screening.
POPULATION SCREENING
In most breed improvement programmes, within-breed selection is limited to a rather small section of the population - the pedigree or stud breeders. This limits selection pressure, the most important determinant of genetic advance. In sheep and goat breeds in West Africa a pedigree breed structure does not exist and in that sense selection can be considered at the level of the national herd. For the following reasons this may offer an opportunity to change a population genetically that has hitherto been ignored:
Most of the sheep and goat populations in the region have not been subjected to controlled man-directed selection.
The large observed variation in these populations may in part be genetic, caused by major genes segregating at low frequencies.
Lastly and not unimportantly it should not be forgotten that the major improvement in animal breeding, particularly as affecting growth, body composition and conformation, made by livestock breeders in the last century, was achieved using very simple selection procedures. Breeders such as Robert Bakewell and the Colling brothers made dramatic genetic progress by ‘screening’ local animal populations for better animals as they subjectively assessed them. As a result they developed the modern European breeds of livestock known today. They exploited the big variation in the then unselected livestock populations with which they worked. Bakewell is quoted as saying “Breed the best to the best and hope for the best”. Since Bakewell's time, the genetics of animal production have become more fully understood and the problem can be approached with much greater scientific resolve and understanding. Two hypotheses, to explain and hence attempt to capitalize on this unusually large variation that exists in many of the livestock populations in the developing countries, can be considered:
The genetic variation in these unselected populations is larger than normal to the extent that the total variation is unusually large.
Major genes with significant effects on production traits are segregating in these populations at low gene frequency.
Evidence to support the first hypothesis partially does exist in that reported estimates of the genetic parameters for sheep and goat populations in the developing countries are very similar, if not higher, to those reported for animal populations in the more advanced countries. There is no hard evidence to support the second hypothesis in that it has not been tested, but there is increasing evidence that the screening of national populations may identify major genes. In recent years at least five examples have been identified in sheep breeds (Australia, United Kingdom, Ireland, Iceland and Indonesia) and currently there are indications of a major gene controlling milk protein in a French goat breed. In any event the screening of national populations for ‘exceptional animals’ makes good sense. At worst it is a means of exercising maximum selection pressure within a population and of course it allows the possibility of identifying animals that are carriers of important major genes.
In the difficult environmental conditions which predominate in West Africa, it can be questioned - how can national populations be screened in the absence of national recording schemes? The only solution is to follow the same approach as the pioneer breeders of the last century. The initial screening should be done subjectively - in other words the breeders are asked to identify ‘exceptional’ animals, if any, in their herds or flocks. These ‘so-called exceptional animals’ should then be recorded on the farm and compared with a random sample of contemporaries. If their performance approaches or exceeds twice the average level of the herd or flock, then those animals should be acquired, purchased or leased, for more controlled evaluation and breeding on a central test farm or research station. In this way an open nucleus herd or flock can be established to act as a source of improver stock for the genetic improvement of the target population.
Screening target
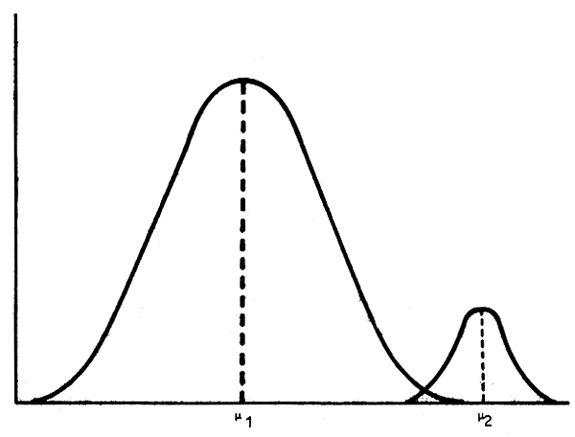
It is deliberately suggested that the selection target is pitched very high, at or near twice the flock average. Assuming a normal distribution for the trait in question (see Figure 2) and that animals are identified at the very upper end of the distribution, namely three standard deviations (α) above the mean (μ), it is easy to calculate the resulting selection differentials in terms of the variance or coefficient of variation (cv). If a selection differential (i = μ2 - μ1) is obtained that is equal to or greater than three standard deviations above the mean (i = ≥ 30 = 3μ1 . cv) then it can be calculated that ‘exceptional animals’ may be defined as being 100 to 200 percent above average as the cv ranges from 30 to 70 percent.
If on the other hand, a major gene or block of linked genes is segregating in the herd at low frequencies then individual animals outside this range may be found. Piper and Hanrahan (pers. comm.) in a simulation study of the possible effects of the Booroola gene in the Australian Merino and in a specially selected (national screening) sheep flock in Ireland, have shown that coefficients of variation and repeatabilities in these flocks can be very high (60 – 70 percent) as a result of the expression of the major gene.
By way of example the method and results of a screening experiment carried out in Ireland are cited (Timon, 1964). Using a simple postcard questionnaire, livestock extension officers in the country were asked to inform the Agricultural Research Institute if they knew of any farmers in their area that had (or had had) individual ewes that displayed exceptional lambing performance. Exceptional lambing performance was defined as:
Ewes that produced four or more lambs at one lambing and previously gave birth to two or more lambs.
Ewes that produced three successive sets of triplets.
These standards were set on the rationale outlined in Figure 2 based on normal distribution theory. It is also worth pointing out that triplet births in the national flock at that time were very infrequent and the average litter size was less than 1.3 lambs/ewe.
Figure 2: Population screening - Genetic rationale
GENETIC ADVANCE: ∆G = h2i Lim (i = h2 = 3 : 1)
SELECTION INTENSITY - SCOPE
| MAJOR GENES | μ2 - μ1 | → 3α | ||
| → 3μ1 - cv | ||||
| VARIATION | if | cv = 30% |  μ2 - μ1 > + 0.9 μ2 - μ1 > + 0.9 | μ1 |
| if | cv = 50% | μ2 - μ1 > + 1.5 | μ1 | |
| if | cv = 70% | μ2 - μ1 > + 2.1 | μ1 |
More than 300 pertinent questionnaires were returned and just over 100 animals were purchased, based on verbal statements from the farmer; in a few cases it was possible to verify the results. Ewes having 4, 5, 6 and one with 7 lambs were identified. Litter size in the purchased ewes of variable and unknown ages averaged more than 2 lambs/ewe; now several generations and 23 years later, it averages 2.3 lambs/ewe. This nucleus flock has made a very significant contribution to the genetic improvement of sheep breeding in Ireland. Now incorporated into a synthetic improver strain, known as the Belclare Improver, it shows all the characteristics of having a Booroola-like gene affecting ovulation rate; at the extreme, one ewe in this breed has recently displayed and repeated an ovulation rate of 17 ova while four daughters from this ewe have had an ovulation rate of 6 ova at 8 months of age. The Javanese Thin Tailed sheep (JTT) is also showing clear evidence of the existence of a major gene controlling ovulation rate (Bradford, Subandruja and Iriquez, 1986). This latter example is cited as evidence that major genes may and have been found in small ruminant populations living in harsh environmental conditions.
A SUGGESTED BREEDING PROGRAMME
While there is little available documentation on the success or otherwise of ongoing genetic improvement programmes in West Africa, the recent interest in the definition and quantification of village-level production systems provides a basis for the determination of objectives and consequently a breeding programme (Mathewman, 1977; Capps, Bonniwell and Bennett, 1979; Adeoye, 1985; Sumberg and Mack, 1985). The results obtained in these studies and those reported by Mason (1980), Ademosun et al. (1983), Berger (1983), Bradford (1983) and Dettmers (1983) show that the reproductive potential of local sheep and goats is relatively high with over 1.7 to 2.0 offspring/dam/per year being recorded for sheep and goats.
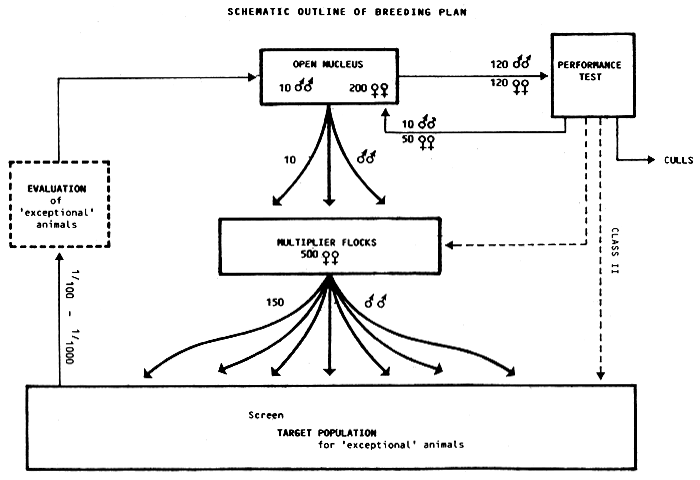
In consideration of this and the different points raised previously, a simple breeding programme based on bodyweight at 6 months of age and relying on mass selection is outlined by way of example in Figure 3. The target ewe population in this programme is 15 000 ewes with an average flock size of approximately 50 ewes. The breeding objective is to increase lamb growth rate and body size. The breeding programme involves five separate phases initiated sequentially as follows:
Screening of the target population for ‘exceptional’ ewes and rams.
Controlled evaluation of the ‘exceptional’ animals.
Establishment of an open nucleus flock.
The organization of a performance test of ram and ewe lambs.
The identification of a number of ram multiplier flocks and a ram distribution programme.
Screening: The first stage in the screening process will involve visiting all the flocks in the target population and in discussion with the flock owners visually identify the largest ewes of good conformation; the particular standard to follow will be determined by the animals available and the farmers' willingness to sell or lease the animals. A selection differential greater than 1/100 should be realized and if possible, a screening rate of 1/1000 should be attempted. Over a period of two to three years a total of 200 ewes should be selected. These animals should then be weighed and if feasible, some measures of body size (body length, heart girth) should be recorded to establish screening standards relative to a group of control (n > 20) animals. If possible observations on the growth of the progeny of these ewes should also be considered.
Ram lambs: As the flocks are being screened for elite ewes a limited number (10–15) of exceptional rams should be identified. These may be ram lambs or adult rams; again standards will have to be established subjectively and in the case of ram lambs, consideration of the birth type and effect of season of birth will be necessary. If possible 50–100 rams should be identified and placed on a standard feeding regime for a period of 100 days after which a final selection of 10–15 rams is made. This selection of the foundation rams can also be phased over two to three years.
The nucleus flock: This eventually will comprise 200 ewes and 10 sire lines and may be established on one farm or on a number of farms, say 4 farms of 50 ewes each. It is not necessary to locate the nucleus flock on a central/government farm although this would be the ideal if it is well managed. Essentially the management of the 200 ewes must be as standardized as possible. All matings of nucleus ewes must be pre-planned and fully controlled in groups of 20 ewes per ram. If various flocks are involved rather than one central test farm, then individual animals will need to be ranked on their performance relative to the flock average.
A simple index (I) on the jth animal in flock i would be calculated as follows:
Iij = (Wij - Wi.) + b(Wi. - W..)
Where Iij = index of the jth animal in the ith flock.
Wij = weight of the jth animal in the ith flock.
Wi. = mean of the ith flock.
W.. = overall mean of all flocks recorded.
b = the genetic component of the between flock variance.
In practice the second part of the index may be ignored without much loss of accuracy particularly in areas where there have been village-run flocks with little or no control of ram/buck movements.
Environmental factors: It is well established that birth type, sex and age of dam have significant effects on lamb and kid growth rate; season of birth has a particularly marked effect on animal growth in West Africa. While the effects of these non-genetic factors on six-month weight are diminished somewhat some adjustment of the data will be necessary; adjustment to a standard 180-day weight is also necessary. Ideally all of these adjustments should be made in a complete statistical analysis (least squares or Best Linear Unbiased Prediction - BLUP) of the data. Where this is not possible individual animals should be ranked on a within sub-group contemporary comparison basis.
Figure 3: Schematic outline of breeding plan
Performance testing: Once the initial screening is completed further genetic improvement will come from the selection of ram lambs and ewe lambs on performance test. It is very important that this performance testing is carried out accurately. Ideally, all lambs should be weaned at 90 days and transferred to a central performance test station for a 90-day test period during which all animals should be treated uniformly. In reality, all the lambs will not reach performance test age at the same time as inevitably there will be a considerable range in lambing date; however, every effort should be made to keep this to a minimum. As necessary lambs should be managed and ranked in age-contemporary groups.
Selection intensity: The 200 ewe nucleus flock is expected to produce 240 lambs/yr; consequently a selection differential of 1/12, i.e. 10/120 will be applicable as regards ram lamb selection; selection of ewe lambs (50/120) of necessity will be less intense.
Multiplier flocks: When matings of the nucleus ewes with the selected 10 ram lambs have been completed, these rams should then be used again, this time in the “multiplier” flocks. These multiplier flocks must ensure controlled mating and ideally should have their own selection programme, initially based on some level of screening and a follow-up selection based on on-farm performance recording of ewe lambs. However, if this is not feasible they can simply be used as propagation farms without any recording. The 500 ewes in the 10 multiplier flocks will on average produce 300 ram lambs. The best 150–200 of these should be made available for distribution to the village breeders.
Annual genetic gain: The rate of genetic gain (AG) inherent in the proposed scheme will be determined by the heritability (h2) of 180-day weight and the selection intensity (i) as follows: AG = h2i.
In the absence of firm estimates of the heritability of 180-day weight in the particular population it is not possible to make specific predictions of genetic gain. However, some generalized estimates can be made as follows. Given that the initial screening is thorough and that the heritability of body size is circa h2 = 0.10, the initial screening should effect a genetic gain (AG) in the nucleus flock of the order of 3.96 × 0.10 = 0.396 standardized units. Assuming a standard deviation of 180-day weight of 2.5 kg, the genetic merit of the nucleus flock should be approximately 1.0 kg above the population average. Subsequent annual genetic gain will result from the performance test selection, principally the ram lamb selection (i = 1/12); this should increment at a rate of 0.17 - 0.20 standardized units or less than 0.5 kg per annum. The transmittal of these genetic gains from the nucleus stock to the breeders will have a time lag inherent in the production of the multiplier rams. However as proposed above the concurrent use of the selected rams in the multiplier flocks will help to keep this time lag to a minimum.
CROSS-BREEDING
The role of cross-breeding or upgrading has not been considered so far in this paper as it is suggested that where there is a high trypanosomiasis challenge only trypanotolerant breeds should be considered. However, in the northern areas as well as in areas where the risk of trypanosomiasis has decreased due to the decline of bush coverage or other tsetse control measures, cross-breeding may have a role.
Whereas cross-bred animals naturally occur at the border of the area affected by trypanosomiasis there is little published data on their productivity or their resistance to trypanosomiasis. Amegee (1984), in a trial comparing the Vogan (a natural cross-bred between sheep of the Sahel type and the Djallonké) and the Djallonké found that male Vogan lambs had a far higher growth rate than male Djallonké lambs. The weight of Vogan lambs at 8.5 months was 44 kg as compared to 24 kg for Djallonké lambs at 9.5 months of age whereas mortality rate was slightly higher for Vogan lambs, viz. 25 percent vs 20 percent. This study would suggest that cross-breeding may have a role to play in the genetic improvement of sheep and goats in the humid tropics of West Africa. Clearly, further experimentation is required before any such conclusions can be reached. These breed comparison trials should preferably take place at the village level to avoid any difference in the level of trypanosomiasis challenge and to ensure that the level of feeding and management is appropriate to the target population.
It is also very possible that significant genetic differences exist between the different strains of Djallonké sheep in the countries of West Africa. For this reason it is strongly suggested that an inter-strain comparison trial be established in the region; this, in turn, could form the basis of a cross-breeding programme to exploit any heterosis that may exist in crosses between the local trypanotolerant breeds.
IMPLEMENTATION OF BREEDING PROGRAMMES
Whereas good estimates of the genetic and environmental factors necessary to rank animals effectively are desirable but lacking in many countries, an even greater limitation is the lack of organization and support services. A first and obvious requirement is the castration of culled or unproven ram lambs and bucks. This is a must. The second requirement is the setting-up of an appropriate, but simple, recording scheme and an operating structure that will implement the programme year after year over a considerable period. Politically this requires a clear and committed policy by the government over a long period of time. Given that the technical direction and methods are correct, it needs sustained actions to implement the desired change. The initiative to set this development train in motion should in the first instance come from the research workers, the trained geneticists, in the country. National breed improvement programmes in most of Europe or the United States over the past 30 years have originated from the initiative and commitment of research workers, either in state, semi-state or university employment. Fortunately, a number of the countries in West Africa have very well-trained graduates and postgraduates in genetics and animal breeding. Only a small proportion of them attempt to impart their knowledge and research experience at farm level. This trained talent must be harnessed and put to work if the potential developments in small ruminant breeding are to be realized. The research worker dedicated to improve a particular breed or breeding system can be a powerhouse of change.
However, research commitment on its own is not enough. Any sizable improvement programme, on a regional or national basis, requires organization and support services; organization to get the improvement of the selected animals down to the target producers in the quickest possible time span. Here is where the effective use of artificial insemination (AI), if it is practical, would be very helpful. Where it can be used it would avoid the pyramidal, pedigree breed-dominated breed structure, that has bedevilled and slowed down the speed of gene flow through the population in so many countries. In most situations where AI is not practical, then the shape of the pyramid must be flattened and changed to optimize the rate of gene flow down to the producers. Of course all of this requires an organizational structure. This may be operated by the government, on a voluntary cooperative basis or in some combination of both. The structure does not matter as long as it works. Equally important, it must have the capability to organize and in many cases carry out the animal recording necessary in the programme, whether the recording is very simple or quite elaborate. Sufficient attention has not been given to this aspect of genetic improvement in many countries, nor has sufficient attention been given to the data analysis component of many breed improvement schemes.
REFERENCES
Ademosun, A.A., Benyi, K., Chiboka, O. and Munyabuntu, C.M. (1983). Review of hair sheep studies in southwestern Nigeria. In Fitzhugh, H.A. and Bradford, G.E., eds. Hair sheep of Western Africa and the Americas. Boulder, Col., Westview Press, pp. 219–226.
Adeoye, S.A.O. (1985). Reproductive performance of West African dwarf goats in southwestern Nigeria. In Wilson, R.T. and Bourzat, D., eds. Small ruminants in African Agriculture. ILCA, Addis Ababa.
Amegee, Y. (1984). Quelques aspects des potentialités zootechniques des ovins et caprins au Togo. Un compte rendu des journées techniques de reflexion sur l'élevage et la protection sanitaire des petits ruminants au Togo. FAO, Rome.
Berger, Y.M. (1983). Djallonké hair sheep in Ivory Coast. In Fitzhugh, H.A. and Bradford, G.E., eds. Hair sheep of Western Africa and the Americas. Boulder, Col., Westview Press, pp. 227–240.
Bradford, G.E. (1983). A note on characteristics of hair sheep in Senegal. In Fitzhugh, H.A. and Bradford, G.E., eds. Hair sheep of Western Africa and the Americas. Boulder, Col., Westview Press, pp. 241–243.
Bradford, G.E., Subandruja and Iriquez, L.C. (1986). Breeding strategies for small ruminants in integrated crop-livestock production systems. In Proc. IDRC/SR-CRSP Symp. on Small Ruminant Prod. Systems in South and Southeast Asia. IDRC, Ottawa, Ontario, Canada.
Brody, S. (1945). Bioenergetics and growth, New York, Reinhold.
Capps, J.D., Bonniwell, M.A. and Bennett, P. (1979). Development of sheep and goat production. Ghana, survey of a village sheep population in the closed forest zone. FAO, Rome.
Dettmers, A. (1983). Performance of hair sheep in Nigeria. In Fitzhugh, H.E. and Bradford, G.E., eds. Hair sheep of Western Africa and the Americas. Boulder, Col., Westview Press, pp. 201–218.
Falconer, D.E. (1960). Selection of mice for growth on high and low planes of nutrition. Genet. Res., 1: 91–113.
Mason, I.L. (1980). Prolific tropical sheep. Animal Production and Health Paper No. 17. FAO, Rome.
Mathewman, R.W. (1977). A survey of small livestock production at the village level in the derived savannah and lowland forest zones of south west Nigeria. Study No. 24. Department of Agriculture and Horticulture. University of Reading. Reading, U.K.
McDowell, R.E. (1983). Dairy cattle breeding in the tropics. Cornell International Agricultural Mimeo. Cornell University, Ithaca, New York.
Peacock, C.P. (1985). Measures for assessing the productivity of sheep and goats in Africa. In Wilson, R.T. and Bourzat, D. Small ruminants in African agriculture. ILCA, Addis Ababa.
Peters, K.J. (1985). Principles of evaluating goat populations in tropical and sub-tropical environments. Proc. 36th Ann. Meeting of EAAP. Kallithen, Greece.
Sands, M. and McDowell, R.E. (1978). The potential of the goat for milk production in the tropics. Cornell International Agricultural Mimeo. Cornell University, Ithaca, New York.
Sumberg, J.E., and Mack, S.D. (1985). Village production of West African Dwarf goats and sheep in Nigeria. Trop. Anim. Health and Prod., 17: 135–140.
Taylor, St. C.S., Moore, A.J., Thiesen, R.B. and Baby, C.M. (1985). Efficiency of feed utilization in traditional and sex controlled systems of beef production. Anim. Prod., 40: 401–439.
Timon, V.M. (1964). Selection for high fertility ewes. AFT Research Report. Animal Production. An Foras Taluntais, Dublin.
Van Vlaenderen, G. (1987). Togo. A study of village level sheep and goat development. In Sheep and goat meat production in the humid tropics of West Africa. Animal Production and Health Paper. FAO, Rome. (In press).
![]()
![]()
![]()