![]()
![]()
![]()
Crossbreeding strategies can be classified into the following three broad categories:
- Breed replacement strategies
- Establishment of stable crossbreeding Systems
- Formation of synthetic populations
In the context of dairy cattle crossbreeding alternatives, there are some practical limitations to the range of possibilities which can be considered. In most cases, attention is limited to two breeds, one a local Zebu population and the other an exotic, usually a Bos taurus breed. The advantages of involving a third breed are seldom substantial. Continuing access to an exotic breed through artificial insemination makes grading up schemes a relatively easy option. The establishment of a synthetic is often operationally easier than the organisation of a rotational breeding structure.
In order to consider any of these breeding strategies it is necessary to balance these questions of operational convenience and cost against the estimated genetic benefits resulting. These in turn depend on the extent of the additive and heterotic effects to be exploited. The scale of these may depend on the environment. The proportions of these effects which are in fact utilised will depend on the fraction of maximum heterozygosity and on the balance of genes between the contributing breeds. The first element in the examination of any particular strategy must therefore be to quantify additive and heterotic effects.
This is probably the most widely practiced of ail crossbreeding Systems. It is in fact a population replacement strategy, replacing more than 90% of the genotype of the local population within four generations.
The breeding structure is shown in Figure 3.1, where E stands for the exotic breed, and L for the local. Figure 3.1 also shows the rate at which gene replacement takes place in the local population, and the extent to which maximum heterozygosity is lost as the generations proceed. After the first two or three generations, the amount of heterosis contributing to performance becomes negligible. This strategy therefore depends almost entirely on the size of the additive difference between the two breeds. It also depends on this additive difference being large relative to any heterosis effects.
In many instances, it is clear that less than 100% replacement of the genes of the local population by exotic Bos taurus genes is desirable. One crossbreeding strategy in these circumstances which has both operational and genetic advantages is to grade up to 50% exotic bulls. The breeding structure, and the way in which heterozygosity is maintained and exotic genes are introduced to the population is shown in Figures 3.2 and 3.3. Bulls from the exotic breed (E) are mated to selected cows of the local population (L1). The F, bulls produced by these matings are then used for general crossing on the main local population (L).
In each generation, 50% of the maximum heterosis effect is maintained. The first crossbred generation in the local population has 25% of exotic genes, but this proportion rapidly approaches a stable value of 50%. The System therefore exploits half of the heterosis effect and half of the additive difference between the local and exotic breeds. The expected performance level is that of the F2 or 50% synthetic (Figure 2.3). Depending on the relative sizes of the heterosis and additive effects, this combination could therefore give close to optimum performance.
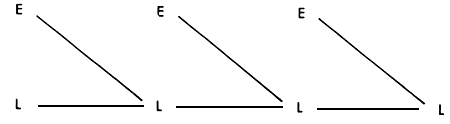
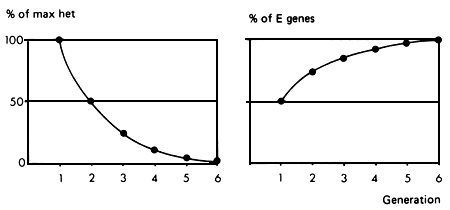
Figure 3.1 Grading up to exotic: breeding structure, rate of retention of heterosis and rate of infusion of exotic genes.
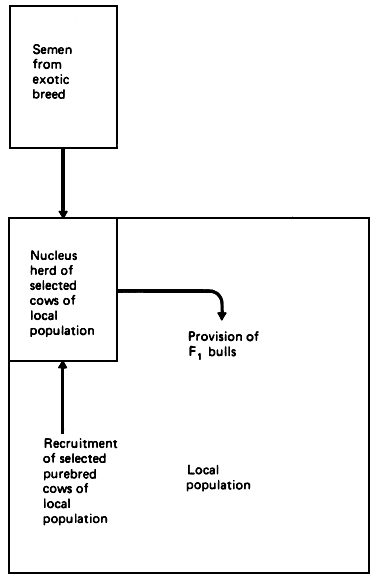
Figure 3.2 Grading up to 50% exotic: breeding structure.
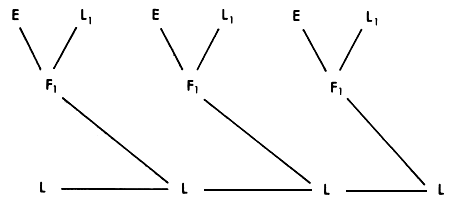
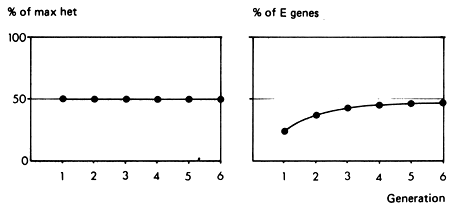
Figure 3.3 Grading up to 50% exotic: breeding structure, rate of retention of heterosis and rate of infusion of exotic genes.
The operational advantages of the scheme are as follows. In many local populations, it is difficult to organise any kind of breeding intervention other than the provision of bulls. On the other hand, it may well be possible to organise an effective nucleus herd of selected females from the local population. These females could then be mated with semen imported from the chosen exotic, and the resulting F1 bulls could be used either in natural service or artificial insemination to breed the local cow population. In addition to achieving a planned exploitation of both heterosis and additive differences, therefore, the structure would also permit exploitation of any selection gains achieved in assembling the nucleus herd. It would also permit continuous exploitation of any genetic improvement in the exotic population.
The overall advantages are that the desired intermediate genotype is rapidly achieved; a significant proportion of maximum heterozygosity is retained; no organised activity is required in the commercial population, other than the acceptance of F1 males; if the selection of B1 females as dams of F1 males is effectively done, then the benefits of improved local genes are recirculated to the population; and finally, the continuing benefits of any selection program in the exotic population are carried through rapidly, and continuously, into the local population.
If a higher degree of exotic genes is required in the crossbred population, but still short of 100%, one possibility is to grade up to three-quarter exotic bulls. This structure is illustrated in Figures 3.4 and 3.5. As in the preceding structure, exotic bulls are first mated to selected females of the local population to produce an F1 generation. These are backcrossed to the exotic to produce three-quarter bred males, which are then used on the local population. This procedure could be repeated in each generation, thus exploiting the opportunity of using whatever genetic improvement was taking place in the exotic population. Alternatively, a synthetic population could be formed by mating the backcross bulls to females of the same genetic constitution, and this nucleus synthetic group could be a continuing source of three-quarter bred bulls for use in the local population.
This breeding System brings the local population up to three-quarters exotic genes within about four generations. On the other hand, the proportion of maximum heterosis, which begins at 75% in the first generation, drops to a level of 37.5%. The expected performance level is that of a synthetic population with 3/4 exotic genes.
As can be seen from the different situations shown in Figure 2.4, the relative performance of populations obtained by grading up to 50% or 75% exotic bulls will depend on the relative sizes of the additive and heterosis effects. For example, the data summarised elsewhere in this report suggest that the additive difference between Bos taurus and Bos indicus breeds is of the order of 100% (of mid-parent mean), while the heterosis effect is about 25%. With these values, the 3/4 exotic population will have a performance about 13% higher than the 50% exotic. If additive and heterosis effects are each equal to 50% of mid-parent mean, then the advantage is only 4%. If heterosis is high, say 50% of mid-parent mean, while the additive difference is low, say 20%, then the 3/4 exotic has an expected performance 1% below that of the 50% exotic population.
This is in some ways the simplest crossbreeding System of ail. Bulls of the exotic breed are used on females of the local population to produce F1 offspring which form the commercial population. The two pure breeds are used in each generation to repeat this process. The main disadvantage in a cattle context is that a large population of the local genotype would need to be maintained in pure form as a source of these crossbred offspring. The system is widely used in pigs and in poultry where, because of the high reproductive rate, a small nucleus of purebred females can provide all the replacements for the commercial population.
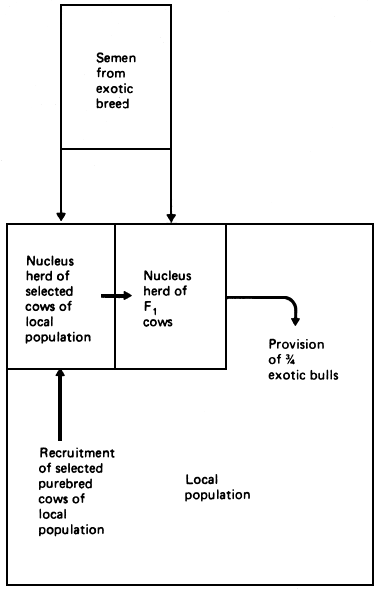
Figure 3.4 Grading up to 75% exotic: breeding structure.
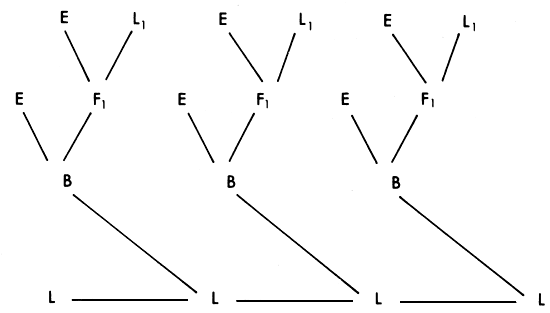
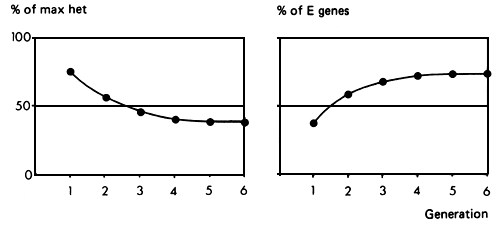
Figure 3.5 Grading up to 75% exotic: breeding structure, rate of retention of heterosis and rate of infusion of exotic genes.
The breeding structure and its consequences for maintenance of heterozygosity and proportions of genes from the two sources is illustrated in Figure 3.6. The biggest advantage of this System is that it maintains the full extent of heterozygosity, and thus of heterosis in ail generations. If heterosis is very important relative to the size of the additive difference, then the System could be worth considering. Despite the fact that the production of F1 offspring is usually the first step in any exotic crossing program in a Zebu population, the continued use of the System beyond the first generation is operationally difficult, and is unlikely to be justified by the relative sizes of the additive and heterosis effects.
This system (also known as crisscross or reciprocal backcross), which appears to be more frequently advocated than practiced, has some operational advantages. The mating pattern is illustrated in Figure 3.7.
The system gives 100% of achievable heterozygosity in the first generation, 50% in the second, 75% in the third, and gradually settles down to a consistent 67% in succeeding generations. The percent of genes from the two parent strains fluctuates, and for any one of the strains, alternates from 33% to 67% in alternate generations. This is no disadvantage in cases where the two types of crosses are very similar. However, in crosses between European and tropical cattle, these two types might perform quite differently. The System has the advantage that, since purebred males are used in each generation, very small pure bred nuclei are needed to service a quite large commercial population. This advantage is enhanced if artificial insemination is used, and the system should therefore have some potential in cattle populations.
The expected performance level fluctuates between two points (Figure 2.4). The difference between these two points amounts to 1/3 of the additive effect. In the case of Bos taurus by Bos indicus crosses, it could therefore be of the order of 33% of the mid-parent mean, which is rather bigger than the maximum heterosis effect. The average expected performance level is better than that of the F2 or 50% synthetic by an amount equal to 17% of the heterosis effect. With an additive effect of 100% (of mid-parent mean) and a heterosis effect of 25%, then the average performance value of a two breed rotation would be 3% better than that of a synthetic.
The principles of operation are similar to those in a two breed rotation, and the mating pattern is shown in Figure 3.8. Each of the three breeds is used in turn in pure form as a sire breed. We assume in this case that there is one local population, and that two exotic breeds (E1 and E2) are used. Each breed is used in pure form as a sire breed. The rotation begins with the use of the first exotic breed on the local population, to produce an F1 population. These are mated to the second exotic breed, to produce a population which has 75% exotic genes. These are then mated to bulls of the local breed, to produce a population with 37.5% exotic genes. This completes one cycle of the rotation.
The consequences of this breeding pattern for the maintenance of maximum heterozygosity, and for the percent of exotic genes in the population is also shown in Figure 3.8. In practice, little heterosis is expected when genes of the two exotic breeds are brought together. In contrast, substantial heterosis is expected whenever exotic genes of either are brought together with genes of the local population. The illustration therefore distinguishes between the expected heterosis effect if all breed combinations gave equal heterosis (_) and the more relevant expected heterosis arising only from exotic by local heterozygosity (…). The former situation has some relevance in, for example, three-breed rotational crossing of pigs or beef cattle as practiced in North America. It can be seen that a high proportion of maximum heterozygosity is maintained. However, if only E × L heterozygosity is of value, the proportion of maximum heterosis expected in each generation then fluctuates very widely from one generation to the next. In Bos taurus-Bos indicus crosses, where heterosis is important, this is an undesirable feature.
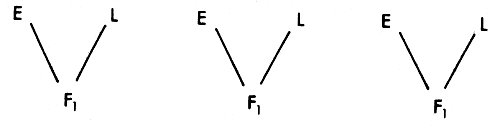
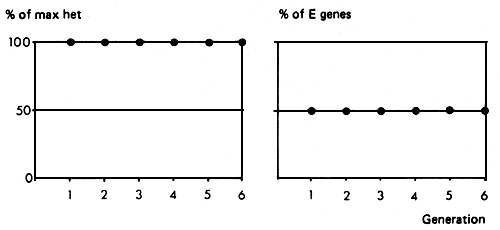
Figure 3.6 Continuous F1 production: breeding structure, rate of retention of heterosis and rate of infusion of exotic genes.
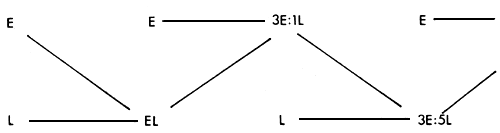
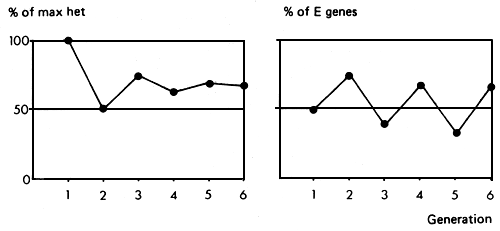
Figure 3.7 Two breed rotation: breeding structure, rate of retention of heterosis and pattern of infusion of exotic genes.
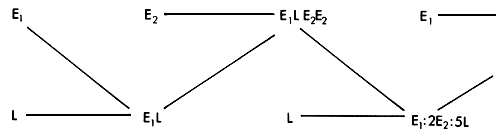
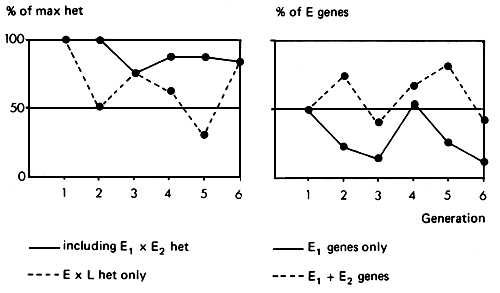
Figure 3.8 Three breed rotation: breeding structure, rate of retention of heterosis and pattern of infusion of exotic genes.
The breed composition as measured by the percent of genes from, say, the E1 strain fluctuates considerably, and eventually settles down to a pattern with repeats every three generations, with proportions of 4/7, 2/7 and 1/7 of E1 genes in each of the three generations (_). It is probably of more interest to follow the proportion of total exotic genes (E1 + E2). This proportion is maintained at a higher level, but also fluctuates widely from one generation to the next (…).
The expected performance of the different generations in such a three breed rotation can be followed, given the expected size of the additive and heterosis effects. The great variability in performance expected over the generations is a disadvantage. The System also has the disadvantage of operational complexity, requiring records of the breed identity of the each cow, as well as the provision of pure bred bulls of ail three breeds. In practice, there could also be difficulties in obtaining acceptance for the use of bulls of the local population once a crossbreeding program has been established.
The expected performance level, as before, depends on the scale of the additive and heterosis effects. In all cases, the average performance level will be somewhat below that of a comparable two breed rotation (by 17% of the heterosis effect).
Synthetic populations are most effected by the loss of any epistasis. If epistatic effects can be ignored, then the two breed synthetic has the same expected performance as a population produced by grading up to F1 bulls. In the latter System, a proportion of any epistatic effects present in the exotic breed will be preserved, while in the synthetic these will be largely dissipated. The importance of this consideration, of course, depends on the kind of epistatic effects postulated, and on the size of their contribution to performance.
The formation of synthetic populations has considerable advantages in terms of operational simplicity over rotational crossbreeding Systems, each of which requires the maintainence of separate nucleus populations. The simplest synthetic is one derived from two parent strains. These are crossed to produce an F1 generation, in which inter-se mating of selected individuals produces an F2 generation, in which selection and mating again takes place. The process is repeated in subsequent generations. The pattern is illustrated in Figure 3.9.
The system gives 100% of possible heterozygosity in the F1 generation, and this declines to 50% in the F2 and subsequent generations. The percent of genes from the E strain is exactly 50% in the first generation, and should also be 50% in subsequent generations, though individuals will vary in the proportions of E genes which they carry, while the concurrent selection exercised may simultaneously raise or lower the average proportion of E genes.
More complex synthetics may be put together in a variety of ways. In the context of exotic and local crossbreeding in the tropics, perhaps the most relevant System would be one involving two exotic breeds and one local, to produce a synthetic with 25% local genes, 25% genes from one of the exotics (E1), and the remaining 50% from the second exotic (E2). Such a System is illustrated in Figure 3.10.
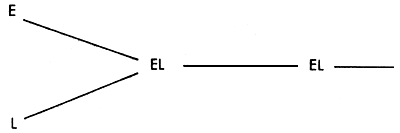
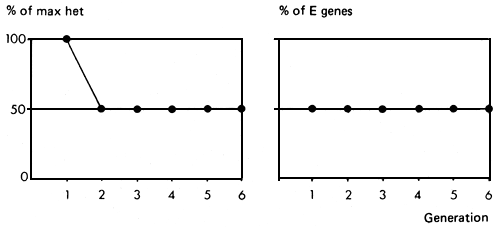
Figure 3.9 Two breed synthetic: breeding structure, rate of retention of heterosis and rate of infusion of exotic genes.
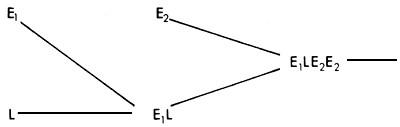
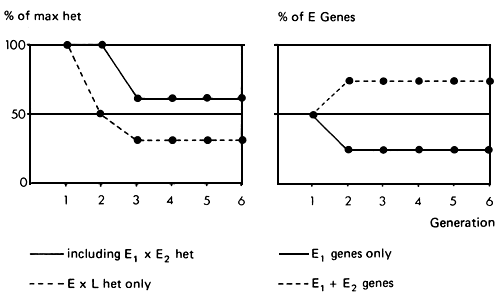
Figure 3.10 Three breed synthetic: breeding structure, rate of retention of heterosis and rate of infusion of exotic genes.
In the first generation, an F1 is produced by E1 and L. These are mated to bulls of the E2 breed, to produce a second generation, which is the foundation for the synthetic population.
As in the three breed rotation, the main interest may lie in maintenance of E × L heterosis. It can be seen (…) that this drops in the third generation to 37.5%, and is maintained at that level in subsequent generations. The proportion of heterozygosity between all three breeds is maintained at a higher level (_). The proportions of genes from the two exotic breeds are maintained at 25% and 50%, giving a 75% exotic gene complement in the synthetic. The expected performance level can again be calculated as a function of the expected heterosis and additive effects, but will in ail cases be slightly below the average performance of a comparable three breed rotation. The latter, however, has the disadvantage of great variability from generation to generation. As in the two breed synthetic, so in the three breed synthetic any loss of epistasis could be of somewhat more consequence than in a comparable rotational cross.
![]()
![]()
![]()