![]()
![]()
![]()
En todas las grandes cuencas hidrográficas de América Latina existen especies de peces que realizan migraciones más o menos extensas con fines reproductivos y tróficos (Welcomme, 1985); Petrere, 1985). Estos desplazamientos son longitudinales a lo largo del canal principal y de los afluentes. En la mayoría de los casos las migraciones ascendentes son realizadas por los peces con fines reproductivos. Existen excepciones para una misma especie dependiendo del lugar de la cuenca donde estas se realicen (Petrere, 1985). En algunos casos, sobre todo en los trechos superiores, las migraciones ascendentes son masivas. En otros, por ejemplo en el río Paraná en la baja Cuenca del Plata, sólo algunos pocos ejemplares realizan extensas migraciones ascendentes, mientras el grueso de los ejemplares permanece en el lugar de marcación o realizan migraciones laterales hacia la llanura de inundación o cauces secundarios (Bonetto et al., 1981). Los desplazamientos pueden alcanzar los 1 500 km (Petrere, 1985) y generalmente exceden los 200 km (Welcomme, 1985). Las velocidades medias de desplazamiento, entre marcación y recaptura, pueden superar los 20 km/día. Estos valores dependen de la especie, época del ciclo hidrológico del río y tramo del mismo en los cuales se realiza la marcación.
Poddubnyi et al. (1981) determinaron velocidades de desplazamiento y ritmos de actividad diaria durante las migraciones reproductivas en el río Paraná Medio. Los peces fueron marcados con cápsulas ultrosónicas y seguidos durante varios días. Se estimó una velocidad media del río en 0.8–1.0 m/s durante los seguimientos. De los cinco ejemplares de Prochilodus platensis marcados que se desplazaron por el sistema de brazos de la llanura de inundación, uno derivó aguas abajo y otro quedó localizado en una zona reducida del cauce principal realizando cortos desplazamientos por dos días. El quinto ejemplar se desplazó aguas arriba siguiendo la pendiente de la ribera izquierda de menor velocidad de corriente y frecuentó a menudo la zona litoral, permaneciendo en reposo en la cercanía de acumulaciones de árboles sumergidos en las pendientes de algunas elevaciones del fondo. Sólo se desplazó durante el día con tres picos de actividad (Fig. 3). La velocidad máxima contra la corriente fue de 1.0 km/h, mantenida por una hora y media. Los ejemplares que se desplazaron por los cauces secundarios y lagunas de la llanura de inundación, se movieron a una velocidad de 0.6 km/h. En los cauces secundarios bordeaban la zona de vegetación acuática y se detenían a menudo para penetrar en ella. En las lagunas permanecieron la mayor parte del tiempo en las zonas de vegetación densa. En experiencias similares realizadas en Salto Grande, un embalse tipo río, los peces se mantenían activos durante el día y la noche, y la máxima velocidad registrada fue de 0.45 km/h. Siguieron una trayectoria muy sinuosa a lo largo de la pendiente del antiguo cauce y en las cercanías de la costa (Poddubnyi et al., 1981).
Un ejemplar de Salminus maxillosus, seguido durante nueve días, se movió en una zona reducida del río. Presentó tres picos de actividad diaria y no se desplazó durante la noche. La velocidad máxima contra la corriente fue de 2 km/h. En el embalse de Salto Grande, Salminus se desplazó aguas arriba con una velocidad máxima de 1.3 km/h. Para Pseudoplatysoma coruscans y Luciopimelodus pati se determinaron velocidades máximas de desplazamiento contra la corriente de 2.6 y 1.0 km/h respectivamente. Al transladarse contra la corriente lo hacen siguiendo la topografía del fondo. La actividad fue de tipo vespertino y nocturno (Poddubnyi et al., 1981).
Los peces tuvieron períodos de actividad que variaron entre 8 y 12 horas con 1 a 3 picos de actividad diaria. Prochilodus y Salminus se orientarían según los gradientes de velocidad de corriente y Pseudoplaysoma y Luciopimelodus según la topografía del fondo. En sus desplazamientos tienden a evitar, por lo general, las zonas de mayores velocidades de corriente. Las velocidades medias diarias en dirección ascendente variaron entre 5 y 17 km/día según la especie (Poddubnyi et al., 1981). Coinciden con los valores medios obtenidos por marca y recaptura (Poddubyni et al., 1981; Welcomme, 1985; Petrere, 1985).
Delfino et al. (1986) observaron que los peces se aproximan a la represa de Salto Grande (ver punto 6.2) siguiendo las zonas de menor velocidad del agua.
Prácticamente nada se conoce sobre las velocidades “de punta” (Webb, 1975) de los peces en América Latina y menos de su dependencia del tamaño de los peces, ni su morfología funcional con respecto a la natación o la temperatura del agua. La experiencia actual nos permite actualmente dar sólo un límite inferior a las velocidades de punta. Para Salminus y Prochilodus, de más de 30 cm de largo total, serían superiores a 1.3 m/s a 11°C (Delfino et al., 1986). Para los géneros Salminus, Leporinus, Prochilodus y Triurobrycon, de más de 30 cm de largo total, superarían los 2.3 m/s a temperaturas superiores a 20°C (Godoy, 1985; Castello, 1982).
Godoy (1985) cita que en el Salto de Urubupungá sobre el río Paraná y antes de las obras de Ilha Solteira, Prochilodus scrofa, Salminus maxillosus, Leporinus copelandii y Pimelodus clarias que fueron marcados, fueron recuperados por encima de los saltos de 18 m de desnivel total y con velocidad del agua de 13.8 m/s. En una experiencia similar en Cachoeira Dorada sobre el río Paranaíba, Prochilodus scrofa y Triurobrycon lundii fueron recuperados en los rápidos (“cachoeira”) y por encima de ella. El desnivel total es de 12 m y las velocidades del agua variaron entre 9 y 12 m/s. También el río Paranaíba, en el canal Sao Simao, Salminus y Prochilodus marcados superaron los 8 km de canal donde el agua circulaba con velocidad de 15 m/s, y en el río Grande individuos de las mismas especies consiguieron superar corrientes de 15 m/s en un canal. En este último caso los peces fueron capturados en diversos puntos de las márgenes del canal. En Cochoeira de Emas sobre el río Mogi Guassu Prochilodus, Salminus, Leporinus y Pimelodus entre otros lograban superar corrientes de entre 6 y 12 m/s de velocidad. Vencían el obstaculo de entrada al primer escalón-tanque de la escalera dando saltos de más de 1.80 m de altura. En el río Parado, en la represa de Itaipava, Godoy y su equipo observaron a Prochilodus y Salminus venciendo velocidades de 20 m/s y dando saltos de 4 m de altura y 5 m de extensión (Godoy, 1985).
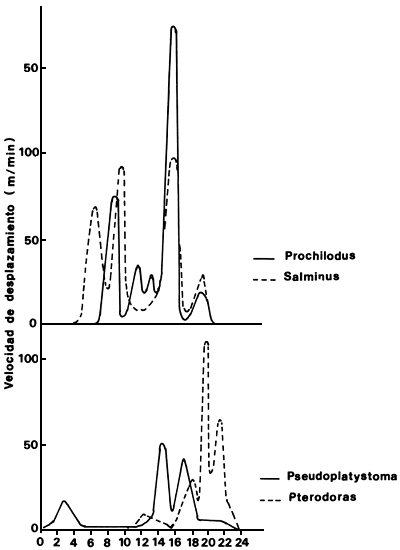
Figura 3. Ritmos diarios de actividad y distribuciones de velocidad para Salminus maxillosus, Prochilodus platensis, Pseudoplatystoma corsucans y Pterodoras granulosus (Poddubnyi et al., 1981)
Las velocidades mencionadas por Godoy (1985) parecen ser un poco elevadas para ser adjudicadas a los peces (Wardle, 1975). Las velocidades de corriente citadas corresponden a velocidades máximas medidas en los puntos de mayor velocidad (Godoy, com. pers.). Es de esperar que los ejemplares de algunas especies se desplacen cerca de las rocas o de las paredes de los canales y que los de otras lo hagan siguiendo los relieves del fondo. En ambos casos las velocidades del agua serían apreciablemente menores (Godoy, com. pers.).
Bonetto (1980) al referirse a la escala de Cachoeira de Emas menciona haber observado a Salminus maxillosus, Salmins hilarii y Prochilodus platensis salvar los obstáculos saltándolos y a Leporinus coperlandii nadando por la lámina de agua.
Al pie de la represa Los Quiroga sobre el río Dulce se observó a Salminus y Prochilodus dar saltos de más de 2 m de altura y 3 a 4 m de extensión en sus intentos por superar los disipadores de flujo al pie del vertedero (Quirós, obs. pers.).
Mientras no se disponga de información más precisa sobre la capacidad natatoria de los peces en América Latina se podrían utilizar, para aquellos peces que de desplazan en la comumna de agua, las fórmulas empíricas obtenidas por Zhou (1982) a partir de mediciones de tiempo de contracción del músculo blanco o anaeróbico en seis especies de peces (Wardle, 1975). La velocidad máxima de natación es afectada por la longitud del pez y por la temperatura del agua (Beach, 1984). Con dicha información se puede predecir la máxima capacidad natatoria de los peces (Fig. 4) y diseñar así las velocidades de atracción a las entradas, el agua auxiliar y las intensidades de flujo dentro de las instalaciones para el pasaje de peces (Beach, 1984). El tiempo que un pez puede mantener el máximo del velocidad (“resistencia”) también está determinado por la longitud del cuerpo y la temperatura y es controlado por la cantidad de glicógeno acumulado en el músculo blanco. La resistencia de un pez a su velocidad máxima se puede predecir a partir de otra fórmula empírica que toma en cuenta la cantidad de energía liberada en la glicólisis anaeróbica. La Fig. 5 relaciona el tiempo que un pez puede mantener su velocidad máxima con la longitud del pez y a varias temperaturas del agua (Beach, 1984). Los tiempos de resistencia obtenidos en la Fig. 5 deberán considerarse los máximos absolutos hasta llegar al agotamiento completo. No parece probable que un pez llegue a este límite. Sin embargo, la tendencia de los peces a salvar obstaculos en sus migraciones reproductivas debe llevar a altas demandas sobre sus reservas de energía y podrían usar parte de sus reservas de glicógeno anaeróbico (Beach, 1984).
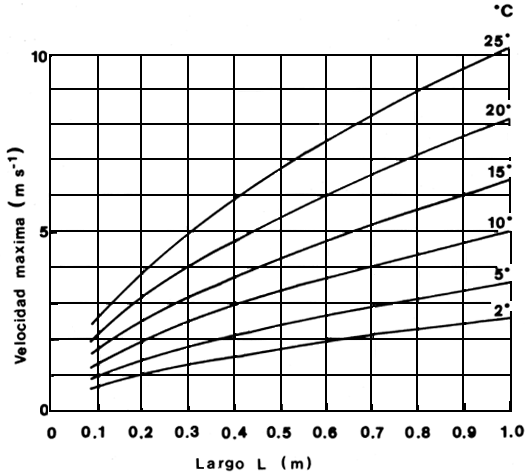
Figura 4. Relación entre velocidades máximas de natación y largo total de los peces para un rango de temperatura de 2°C a 25°C (Beach, 1985)
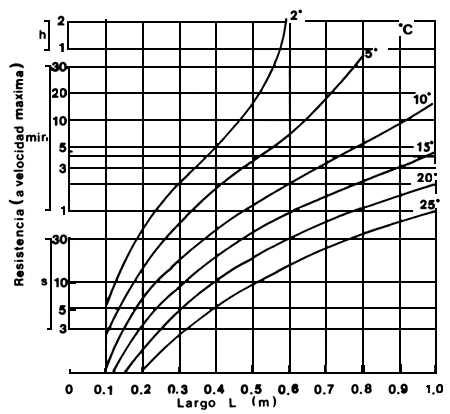
Figura 5. Relación entre la resistencia de un pez en su velocidad máxima y el largo total, para un rango de temperatura de 2°C a 25°C (Beach, 1985)
![]()
![]()
![]()