![]()
![]()
![]()
There is a natural zoning, biological and geological, in every sizable portion of the hydrosphere. In the marine domain, depth is undoubtedly the leading parameter responsible for zoning. The decisive influence can be only modulated by other parameters. In the continental domain (aquatic or otherwise), latitude, which affects temperature and photoperiodicity, seems to be the fundamental parameter. Is there a dominant parameter in the paralic domain, and if so, what is it?
Until now, salinity has generally been considered as the fundamental ecological parameter of the distribution of the communities in paralic milieux (Kiener, 1978, Remane and Schlieper, 1956; Petit, 1962; Sacchi, 1967; Vatova, 1963), and it is on a salinity scale that the usual classification rests: euryhaline, mesohaline, poikilohaline milieux, etc. A simple observation of the natural surroundings which leads to the considerations mentioned in the previous chapter, shows that salinity (along with the major ionic relations1, etc.) as a fundamental ecological parameter of the paralic domain is a mistaken notion.
It is likely that the main reason is the variability, but also the accessibility of this parameter, the presence in paralic milieux of “abnormal salinities” (with relation to the sea which is considered “normal”) is an inducement to consider this parameter as paramount. It is true, moreover, that for a given basin, the biological zoning can generally correspond with the local geological zoning (Fig. 16); on a regional scale, if the correspondence is less clearcut, it can be ascribed to local differences and other parameters.
Hyperhaline paralic milieux are rare and to date little studies. Therefore few scientists have studied both hyper-and hypohaline milieux: only comparison between the two, together with the revelation of an identical zoning for both could cast doubts on and lead to the discarding of a mistaken idea (Guelorget and Perthuisot, 1982).
Finally, another error accepted by many authors is the chemical stability of seawater (in composition and concentration) in the course of geological time. This would imply that the marine domain is the fundamental original one, from which most aquatic stocks issue, and that those which “adapt” to “abnormal” milieux must “solve problems of osmotic regulation”. Now, there are firm doubts regarding the chemical stability of seawater (perthuisot, 1980), and on the fundamental or original quality attributed to the marine domain.
It is established that the biological zoning organizes itself roughly according to the distance from the open sea, as to the geochemical gradients (whatever their value). In fact, this is only an estimate. In more detail, it is noted that the weaker the exchange of water with the sea the more “typically paralic” the communities become, i.e., they are composed of eurybiontic species, not numerous, with a strong density, etc. Conversely, the nearer the zones where the movements of the water allow a regular renovation of the milieu, the richer in species the communities are and the lower the density becomes (Guelorget et al., 1981, 1982 (a) and (b); Perthuisot, 1983),
The most adequate term to quality the value which organizes the biological zoning of the paralic ecosystems would appear to be “confinement”. This term, used in a both empirical and intuitive way in various disciplines of natural sciences is ambiguous: for example, it is sometimes used to mean “anoxia” (Appendix 3).
This word is used to imply “with relation to the sea”; first, because the lagunar ecosystems are arranged in relation with the communication link with the sea, and second, because of the initial concept that in the smaller, even restricted basins where most of the life-giving elements (mineral salts, trace elements, vitamins, etc.) come from the outside (like oxygen in a nearly closed room); the communities organize themselves, according to the rarity of these elements, rerefaction which they themselves are mostly responsible for. This idea (which will be developed later) is based, for instance, on the amazing capacity of molluscs to fix certain trace elements (Cu, Pb, etc.), within their tissues.
Obviously, in the marine domain, the situation is otherwise, since it can be considered infinite, or at least inexhaustible. if equated to the biomass it contains with the available volumes of water and elements.
Thus, confinement at a given point in a paralic ecosystem would correspond to the lack of “vital elements” of marine origin at that same point. Since this deficiency is essentially linked to the biological activity itself, it is understood that, schematically, confinement is a function of the time it takes water coming from the sea to reach the point in question.
Further, it can be said that the marine domain is the main communication agent between the different fractions of an apparently very fragmentary domain. This is without doubt a determining factor in the dissemination of paralic species, and of the completion of their biological cycles. On the other hand, many marine species, particularly fish, use paralic milieux to feed.
The confinement of a specific zone of a given paralic milieu is the outcome of various factors such as the size of the communication link, the force of the tide, the currentology of the basin, the continental inflows, etc. 1. It is therefore essentially hydrodynamic. In this connection, it must be mentioned that depth can play a fundamental role when it leads to the presence of a deep stable water mass: it then creates a very large local confinement that can bring about the almost total disappearance of the benthos, and the appearance of facies of a euxinic type (Perthuisot, 1975, 1980).
Finally, when the confinement becomes very widespread (in the Far paralic), its leading role in the dynamics of the communities diminishes, and biological logic tends to depend upon other factors.
What is the relationship between confinement and salinity (or concentration)? Climatic and hydrographic data must be examined: it is clear that in any given basin, salinity at a given point depends both on the renewal of the milieu, hence of the confinement, and on the local freshwater supply (by convention, that algebraic value of the difference between evaporation and freshwater supplies is termed “hydrous deficit”). If the hydrous deficit is positive, the basin is hyperhaline, if the hydrous deficit is negative, the basin is hypohaline. In each basin, hyper-or hypohalinity increases with confinement to a given value (not nil) of the hydrous deficit provided the latter is constant for the whole of the basin being considered.
This proposition can be formalized mathematically with a schematic and theoretical example, starting with the following hypotheses:
- The freshwater supplies of the basin (rain, continental supply) consist of pure water, are constant and are diffused instantly over the entire basin where evaporation is also uniform.
- Depth is low and constant.
- All major ions coming from the sea are preservative (this constitutes a condition incompatible with natural processes, but entails only a minimal error).
- The hydrodynamic conditions of the basins remain stable in time.
In these conditions, it is possible to define the loss of freshwater by unity of volume and unity of time (in an algebraic value), taking Δ h as that value directly proportional to the hydrous deficit. If Δ h is positive, it means an excess of evaporation; if negatives, it shows an excess of freshwater supply in the basin. This value is specific in each basin.
Furthermore, one should consider a point p of this ideal paralic basin. At this point, a volume of water Vp contains N major ions of marine origin at a time t. Concentration at that point is:
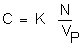
These N elements were all in the sea at the instant of time to, where they occupied a volume Vo of solution: the concentration of sea water is expressed as:
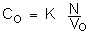
The time lapse T = t - to is a constant of this point p if the hydrodynamic conditions remain stable and can be defined as the time of renewal of the milieu at point p which controls the “confinement rate” at that point.
The volume variation from Vo to V is:
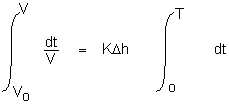
as a result:
Log (V - Vo) = K Δ h T
V = Voe - K Δ h T
and
C = Co K Δ h T
Deciding a priori upon a given concentration C, T = constant. For example, C = Co can be obtained for Δ h = O or for T = O, i.e., for basins whose hydrous deficit is nil, or for zero confinement, i.e., the marine domain itself.
This formula is, of course, over-simple and not a natural reality. However, it shows that granted an equal confinement, two points from a same basin or from different basins in similar climatic conditions, have closely related salinities. Conversely, in very different climates, two points of equal confinement have very different salinities. In all cases, as confinement increases, the difference in salinity (or concentration) from the sea increases, except when the hydrous balance is close to zero: in that case, the salinity remains close to that of the sea, whatever the confinement. It is clear that in nature, the more confined a milieu is - hence enclosed within the continental domain - the less likely it is to belong to an environment with a balanced hydrous value (Fig. 17); except if it is a bathymetric confinement: in the case of stratified basins, the lowest body of water is characterized by a zero hydrous deficit. Lastly, if the confinement is nil, the point in question belongs to the marine domain.
Figure 17 illustrates the previous and present considerations and ideas on the respective roles of confinement and the local hydrous balance in the organization of the biological and geochemical gradients of the paralic domain.
In this diagram, as the parameters used (confinement, hydrous deficit) cannot be given accurately, various paralic basins have been introduced in an approximative but not an arbitrary way. Their juxtaposition is intended to show the apparent concordance in each basin of the biological gradients and the chemical gradients, and also the total obliquity of the geochemical zoning over the biological zoning on the scale of the entire paralic domain since this biological zoning depends largely on confinement alone.
In spite of present inability to quantify either the confinement or the “hydrous deficit” in natural milieux, this concept seems to account for the situation encountered in the field better than an ecological classification of the paralic milieux according to their salinity.
Figure 18 is an attempt to picture the sedimentology of the paralic domain: its only aim is to confirm the total obliquity of the faciological zoning over the biological zoning.
Thus, to each of the three domains which constitute the hydrobiosphere corresponds a fundamental parameter: depth in the marine domain, latitude in the continental domain, confinement in the paralic domain.
As emphasized before, confinement is a complex and abstract value, which cannot be measured in the present state of knowledge. Not having a unit of measurement, a qualitative scale is proposed (comparable for example, to Mohs' hardness scale or Mercalli's scale for the intensity of earthquakes). This attempt cannot rely on either geochemistry or sedimentology: it is therefore necessary to use the peculiar features of the biological zoning, and in this connection, benthos proves convenient: apart from the fact that it is relatively easy to harvest, that it allows an easy direct observation, and in addition to its autochthonous species, it integrates in the short term, minor variations of the milieu, and reflects the latter's global conditions.
In comparing various Mediterranean paralic basins, it is possible to establish 6 degrees of confinement starting from the particular features of the distribution of benthic species on a soft bottom (Fig. 19). This scale, valid for the Mediterranean, concerns only the portion of the paralic domain close to the sea (Near paralic) (Perthuisot and Guelorget, 1982; Guelorget et al., 1982, 1983).
Zone I - This zone falls between degrees 0 and 1. In this zone situated in the immediate vicinity of the communication links with the sea and which is simply a continuation of the marine domain, the macrofauna consists of many strictly thalassic species generally belonging to the fauna of the Well Calibrated Fine Sands (WCFS) biocenosis (pérès and Picard, 1964).
Zone II - This zone falls between degrees 1 and 2: from degree 1, the penetration into the paralic domain can be considered effective. The benthic macrofauna is characterized by the loss of the sensitive strictly marine species, the most “stenohaline”. Molluscs can still be found: Mactra corallina, M. glauca, Tellina tenuis, Donax semistriatus, D. trunculus, Acantnocarida echinata, Dosinia exoleta, the Polychaeta: Audouinia tentaculata, Magelona papillocornis, Owenia fusiformis, Phyllodoce mucosa, Pectinaria koreni, the crustaceans: Portunus latipes, and the echinoderms: Asterina gibbosa, Holothuria polii, Paracentrotus lividus. Lastly, the presence should be mentioned of Branchiostoma lanceolatum in the sandy facies devoid of “organic pollution”.
The phanerogamic macroflora loses the Posidonia oceanica; and small Phanerogams predominate such as Cymodocea nodosa, Zostera noltii and Caulerpa prolifera.
The limit of this zone (degree 2) is marked by the total disappearance of echinoderms.
Zone III - This zone falls between degrees 2 and 3: it is dominated by “mixed species” 1 Venerupis decussata, V. aurea, Scobicularia plana, Corbula gibba, Loripes lacteus, Gastrana fragilis, Akera bullata, Nephthys hombergii, Armandia cirrosa, Glycera convoluta, Upogebia littoralis) as far as the benthic macrofauna is concerned.
Zone IV - Zone falling between degrees 3 and 4: at degree 3, all thalassic fauna disappears, and the species found are strictly paralic: Abra ovata, Cerastoderma glaucum, Hydrobia acuta, Nereis diversicolor, Gammarus insensibilis, G. aequicauda, Corophium insidiosum).
The macroflora is characterized by the appearance of Ruppia sprialis.
Zone V - Zone falling between degrees 4 and 5: in this zone, the phytoplanktonic production (pinnated diatoma, Cyanophyceae) is at its maximum. The surface of the sediment is in the process of being colonized by the Cyanophyceae. Thus the sedimentation is essentially organic, and the interstitial milieu highly reducing.
The community consists essentially of a vagile fauna with detritivorous crustaceans (Sphaeroma hookeri, S. rugicauda, Corophium insidiosum, Idothea balthica), browsing gastropods (Hydrobia acuta, Pirenella conica), Polychaetes such as Nereis diversicolor, and larvae of Chironomidae in the uppermost stratum of the sediment (a few millimetres). In this zone, and most particularly in very hypohaline milieux, elements of freshwater fauna appear (Tricoptera, Oligochaeta, Odonata). Further, in hyperhaline milieux elements of evaporitic fauna (Artemia salina) can be found.
The plant communities - if any - are dominated either by Potamogeton pectinatus, or by the Characeae. They can also be mixed.
Zone VI - Zone falling between degrees 5 and 6. It represents the passage into Far paralic, either freshwater or evaporitic, and is usually characterized by an almost total colonization of the substratum by Cyanobacteria which form algal fields or stromatolithic structures. The passage into freshwater is marked by the appearance of strictly freshwater species and the persistence of a few vagile paralic species (Sphaeroma hookeri, Microdeutopus gryllotalpa, Gammarus insensibilis). The sub-evaporitic milieu is characterized by the absence of benthic macrofauna, except browsers which momentarily visit that zone for trophic reasons (browsing of the algal fields).
The far edges of this zone are the outer limit of the Near paralic. Forminifera disappear, which coincides, around the evaporitic pole, to a considerable extent with the disappearance of stromatolithic structures.
Beyond this, tin the Far paralic, the role of confinement seems to decrease, as far as present knowledge indicates.
According to the confinement scale proposed, the organization of a peri-Mediterranean type lagoon can be illustrated (Fig. 19). This graph shows the zoning of the milieu according to its confinement. With regard to this standard diagram, the lagoons in question can either spread over the entire scale, or occupy only a portion of it.
Bahiret el Biban, a hyperhaline lagoon, covers the entire confinement scale; however, the largest portion is situated in zones II and III, thus showing low confinement of sizable and regular exchanges with the sea (wide pass, strong tidal currents) (Guelorget et al., 1983).
This basin which has a complex history, is little confined at present, because it communicates extensively with the sea. Only a few small marginal areas can suddenly become confined, thus leading to the development of algal fields (Frisoni et al., 1982).
Almost completely situated in zones IV and V, this lagoon - of deltaic formation - hyperhaline in summer, reaches the cyanobacterial zone in its northern part (Frisoni et al., 1982). Only a narrow strip along the lido indicates some influence of the marine domain.
These lagoons, close to that of Logarou and belonging to the same deltaic formation, have the same pattern of confinement, but with a negative salinity gradient (Frisoni et al., 1982).
These two lagoons are very similar. The marine influence prevails. However, the hydrological originality of Diana has permitted the establishment of a little-confined zone (II) around the sides furthest from the communicating link with the sea. Due to the great depth (9–10 m) in relation to the surface area (500–700 ha), the central zones of these lagoons, little renewed, are submitted to a bathymetric confinement linked to a “hydrological remoteness” from the sea, and above all, to an organic enrichment; the latter entails the appearance of a specific Corbula gibba facies (Burelli et al., 1979; Frisoni et al., 1983).
Like the Bahiret el Biban - but with a prevalence of zones IV and V - this lagoon covers the whole of the confinement scale, but towards the freshwater pole (Frisoni et al., 1983).
The whole stretch of water is located in zones IV and V. This high confinement is due to the hydrological isolation (long and narrow channel) on the one hand, and the high bathymetry (6 m) with regard to the surface area (800 ha) on the other. The conjunction of these two characteristics causes a confinement which is much more pronounced than in lagoons of similar size and morphology (Diana, Urbino).
This lagoon offers a particular originality with the presence of algal fields (zone IV) in the vicinity of the communicating link with the sea: those regions contiguous to the embanked channel are isolated from marine currents and undergo a high confinement (Guelorget et al., 1983).
This entire lagunar system with few communicating links with the sea (2 graus) is almost entirely situated in zones IV and V, except for the regions of the graus which allow the implementation of thalassic communities.
The prevailing influence of the catchment area leads to the appearance of freshwater zone VI at the periphery of the complex.
This former salt production condenser (Perthuisot at al.; 1983) is an annex of hypohaline paralic marsh. More confined and saltier than the latter, the entire basin is situated at the upper end of the confinement scale (II, IV, VI).
With regard to the biological characterization of benthic macroflora and macrofauna, the biological zoning of these characteristics is sufficiently clear and stable as to define a confinement scale. This is not the case for the other links of the trophic chain which, composed as they are of more mobile species reacting more strongly to the minimum variation in the milieu, present a less obvious, more fluctuating zoning pattern.
Although there are, in most of the paralic basins studied pronounced and significant gradients concerning the phytoplanktonic biomass and the chlorophyll production, it seems that the phytoplanktonic communities are relatively homogeneous in each basin. This is probably due to the fact that the major part of each basin spreads over a limited number of confinement zones (usually one or two). Thus only the comparison of a considerable number of paralic basins can trace the outline of a specific zoning of paralic phytoplankton.
(a) Taxonomic resources
Finally, it should be noted that an increase in confinement results in a decrease of the taxonomic resources; however, the basins strongly influenced by continental inflows gain in freshwater taxa (Table 1).
(b) Specific composition of the phytoplanktonic communities
The zones directly under reviving marine influence (I, II, III), are characterized by the presence of typically neritic plankton essentially composed of centric Diatoma - up to more than 25% on relative average - (Skeletonema costatum, Melosira sp., Coscinodiscus sp., Chaetoceros sp., Rhizosolenia sp.,
Thalassiosira sp.) along with pinnated Diatoma (Nitzschia closterium, Thalassiotrix frauenfeldi) and with some Peridinieae (Ceratium furca) and various nannoplanktonic forms (Chlorophyceae, Cryptophyceae, Cyanophyceae).
In the typically paralic zones (IV, V) centric Diatoma are very rare; pinnated Diatoma, although always present, are rarely plentiful (Navicula sp., Gyrosigma sp., Amphora sp., Striatella sp.). The community is generally dominated by Peridinieae and the nannoplankton groups.
Among the Peridinieae can be noted the frequent and abundant presence of Exuviella compressa and Prorocentrum scultellum which can represent up to 99% of the community (Lake Melah). These species can also be observed all through the annual cycle in the same basin (Etang de Biguglia (Frisoni, personal communication)1.
In the most confined zones of the Near paralic (zone VI), the phytoplanktonic communities are dominated by species which are usually periphytic or benthic. In this connection it should be noted that in those usually very shallow milieux, the micro-phytobenthos easily regains a state of suspension, that sampletaking can perturb the milieu and affect the element of the phytobenthos all the more as the individuals are affected by lagunar “dwarfism”; lastly, in hyperhaline milieux, the density of the water or the brines practically prevents the existence of free microphytobenthos. The species that would have composed it are in suspension in the milieu.
Beyond zone VI, in the Far paralic, the communities differ according to whether the freshwater pole or the evaporitic pole is considered. In the nearly fresh waters, the milieu is invested by typically freshwater species. In the evaporitic brines, the community consists essentially of Dunaliella salina (or viridis) Chlorophyceae, with salina being the red form giving the colour so characteristic of active salt crystallizers, and viridis the green form of the moderately salty condensers; to these can also be added the cyanobacterian structures (various blue algae and S and Fe bacteria) (Cornee, 1983).
Thus, as far as phytoplankton is concerned, a zoning similar to and superimposable on that which is presented by the macrobenthos and which defines the confinement scale (see Table 1) has been outlined.
As with the phytoplankton, the study of various paralic basins shows a common ichthyofaunistic pattern according to the confinement.
The following species are noted:
- Sedentary species (paralic). These are few, present throughout the year, small and shortlived. Their entire biological cycle is completed in a paralic milieu. They have a very high breeding potential and are present in large numbers. As far as Mediterranean basins are concerned, Atherina boyeri, Aphanius fasciatus, Potamoschistus marmoratus, Syngnathus abaster should be mentioned.
- Migrant species, often larger than the sedentary ones, with a longer life-span and whose biological cycle comprises a necessary reproductive phase at sea (or of ill-defined type). Within this group are to be distinguished:
The regularly migratory species (“mixed” species) whose fry and juveniles (0+) enter a paralic milieu to spend a compulsory (trophic) period of their biological cycle (Appendix 3): These include mullet, some Sparidae (Sparus aurata, Diplodus sargus, Pagellus mormyratus), sea bass (Dicentrarchus labrax), sole (Solea vulgaris), plaice (Pleuronectes platessa), eels (Anguilla anguilla). Some adult or juvenile (1+) individuals of these species are liable to return occasionally to the lagoon to feed.
The occasionally or accidentally migratory species (usually thalassic) are sometimes found by chance in a paralic milieu. Among the more frequent are: Labridae, Gobiidae, garfish (Belone belone), anchovy (Engraulis encrasicolus), red mullet, Rajidae (Myliobatis aquila), Caeculum caeca.
Hereafter all individuals belonging to sedentary species will be referred to as “sedentary”, and the fry and juveniles (0+) of the regularly migratory species as “migrant”. The term “occasional” will be applied to all the others.
The study of the three largest lagoons of the eastern plain of Corsica (Bigugila, Diana and Urbino) (Frisoni et al., 1983) outlines the relative connection of the ichthyofaunistic communities to the confinement scale, both from a qualitative, and a quantitative point of view. The diagram thus proposed can be applied to all the paralic basins having been similarly studied (Frisoni et al., 1982; Guelorget et al., 1983; Guignard and Zaouli, 1981).
In the Corsican lagoons, migrant and sedentary species constitute more than 90% of the stock collected.
(a) Number of species and diversity
In the three lagoons, the same annual evolution of the number of species can be observed with one peak in spring and one in autumn, each corresponding to the propitious periods for the “lagoon-ward movement”, of the migrant fry and juveniles.
Taking these seasonal variations into consideration, the overall number of species in Biguglia is much lower than that in Diana and Urbino. Moreover, whereas the last two are very homogeneous (Table 2), at Biguglia a decrease in the number of species can be observed between the centre of the lagoon and the grau: this again corresponds to migrant movement and can be shown by the study of the proportion of migrant stock to sedentary stock(M/S). At Urbino, the ratio is very low because of the poor communication with the sea. At Diana, the opening of the basin allows a better stocking with fry. However, as the edges are generally only slightly confined, the migrants spread themselves evenly throughout the basin. At Biguglia (a much more confined lagoon), the migrants, although numerous, are mainly localized in the vicinity of the grau and seldom penetrate the rest of the lagoon. These considerations can also apply to the “occasionals ” which are more numerous at Diana than in the two other lagoons (see Table 2).
The diversity of the resident species, measured by the equitability of the Shannon coefficient (ESH) also shows (1981) a cycle with a spring maximum at Biguglia, and a summer one at Diana Urbino. The minimum diversity occurs in winter, a period when stocking with young fish is low: it therefore characterizes the sedentary populations.
At Biguglia, the rather high minimum diversity can be explained by the presence of a large range of the sedentary ichthyic populations. On the other hand, the relatively low maximum diversity shows that the influence of the migrants upon the resident populations remains moderate (except in the vicinity of the grau), and affects only a small number of species: the migrants do not greatly favour highly confined zones.
At Diana and Urbino, the very low minimum diversities are in keeping with the high homogeneity of both the basins. At Diana, the maximum diversity is high because stocking with young fish is easy, and moreover, as the basin is on the whole little confined, the migrants spread throughout the basin. On the contrary at Urbino there is a low maximum diversity because of the small number of migrant arrivals related to the uncertainly of communication with the sea.
(b) Sedentary species
Table 3 gives the average annual percentages of the captures of the various sedentary species in various zones of the lagoons studied, and the corresponding weights.
Considering only Aphanius and Atherina, it is noted that the former predominates in the vicinity of the graus in the low confinement zones, while Atherina predominates in the more confined zones. This is also the case at Diana whose northern basin, although near the mouth, is on the whole more confined than the southern basin.
The mottled goby seems to behave like the Aphanius as far as its distribution is concerned, except at Urbino where it is evenly distribution, but low in numbers.
The syngnathid, very dependent on the plant communities, is seldom seen in the zones near the mouths.
(c) Migrant species
They are 20 species of fry and juvenile migrants listed in the three lagoons. They can be divided into three groups:
| Mullets (5 species) - present in the three lagoons | ||
| Sparidae | - | Diplodus (5 species) Puntazzo (1 species) Boops (1 species) Lithognathhus (1 species) |
| Other species - | ||
| Sea-base (Dicentrarchus labrax) Soles (Solea vulgaris and Solea sp. at Urbino) Eel (Anguilla anguilla) Garfish (Belone belone) Anchovy (Engraulis encrasicolus) Red mullet (Mullus barbatus) | ||
At Biguglia, the fry belonged mostly to eurybiontic species (garfish, eels, sea-bass). On the other hand, there were very few sparidae (only 3 species), and no soles, red mullets or anchovies. At Diana and Urbino, the migrant species are more diversified and include more specifically marine species. This is also the case for the “occasionals”.
The specific composition of the migrant populations varies according to the season: if the mullets are present all the year round, due to successive arrivals of the different specifies, the Sparidae and the sea-bass appear mostly in the spring, and the other species in late summer and in autumn.
The distribution of the different species is not uniform. The mullets are concentrated in the grau regions (Diana and Urbino) where they often pass through in shoals. The other species are more evenly distributed. Moreover, the stocking with young fish is very irregular: at Diana, for example, in May 1978, 16 % mullets, 59 % Sparidae and 25 % other species were counted: in May 1979, 97 % mullets, 2 % Sparidae, 1 % other species. Obviously, the state of the grau influences the stocking with fry: at Urbino, it remained practically nil for several years while the grau was closed. Its recent opening and maintenance have allowed a better stocking, and a proportion of migrants more in keeping with the position of this lagoons on the confinement scale.
(d) Diets and coefficients of condition
The species of the paralic milieux are essentially carnivorous or omnivorous, and very few microphagous. Therefore, the fish consume directly very little phytoplankton.
As far as the sedentary species Atherina boyeri is concerned, the highest average weights and sizes of the captured individuals are to be found at Biguglia, taking the year as a whole. Again at Biguglia is noted the highest monthly average coefficient of condition. A. boyeri reaches the maximum coefficient of condition in the spring, i.e., during the reproduction period.
The coefficient of condition is given by the formula:
Kc = W.L.-3 where W is the weight in grammes and L the length in centimetres.
With regards to mullet (Liza saliens), a regularly migrant species, a similar evolution is found according to seasons; but the individuals reach their maximum coefficient of condition towards the end of summer, before migrating towards the sea. This coefficient is given higher at Biguglia than the two other basins.
Thus, the Biguglia lagoon seems to be more favourable for the growth of individuals, especially sedentary ones, than the other lagoons. This is likely to be related to the heavy phytoplanktonic biomasses which characterize this highly confined lagoon, and which result in the presence of great numbers of primary consumers.
(e) Conclusions
Unlike the relatively fixed benthic communities which settle in a stable zones, according to the confinement range of each basin, the ichthyological populations are much more mobile. Therefore, their organization system is of a more global type, and depends upon the totality if the characteristics of each basin and particularly on the relative surface areas occupied by the various zones of confinement, on the type of communication with the sea, etc. Thus, the qualitative and quantitative characteristics of the ichthyological fauna of the paralic milieux account for the originality of each of them, and for the place each occupies on the confinement scale.
Certain data suggest, however, that within the ichthyological population biological gradients exist, which are identical from one basin to another (the Atherina/ Aphanius population, for example), which further studies may perhaps specify more accurately.
Biological study of the peri-Mediterranean lagoons shows that the quantitative variations of the phytoplanktonic and benthic communities (macrofauna of invertebrates) differ according to confinement.
The chlorophyll biomass (expressed in mg of chlorphyll a per m3- fluorimetric analysis) increases regularly from the marine waters (values close to 1) up to zone V. Thus, the lagoons of Diana, Thau, el Biban and Nador, have a biomass of between 1 and 4; the lagoon of Biguglia reaches values of about 20; the lagoon of Mauguio (subject to urban and agricultural pollutions) has biomass values of about 1 000.
Beyond this zone V, the shape of the curve will depend on which pole (freshwater or evaporitic) is approached. In a freshwater milieu, either the persistence of a high biomass (eutrophic milieu), or a decrease of this biomass (oligotrophic milieu) is noted. In a sub-evaporitic milieu, in all cases a drastic decrease can be observed in the phytoplanktonic biomass. In this extreme zone, the primary link is hardly represented except by the microphytobenthos (algal field).
The biomass of benthic of benthic macrofauna (expressed in grammes of dry decalcified weight per m2) shows a similar curve. However, this curve shifts in relation to the preceding one: the position of the maximum moves into zone III (Fig. 29). However, as with the primary link, this diagram can show vertical fluctuations (maximum values) according to seasonal, interannual variations of the enrichment of the milieu by organic pollution.
The following values should be kept in mind:
- Around 1 g in one I, about 10 g in zone II, about 50 g in zone III subject to few organic deposits (el Biban), reaching as much as 500 g in highly enriched zones (Prévost).
Beyond zone III, the curve shows a decrease of the values as far as zone VI where, as with the primary link, two possibilities arise:
- increase of biomass through contamination of the freshwater fauna when in hypohaline aquatic milieu;
- disappearance of benthic macrofauna when moving towards the evaporitic pole, which is the limit of the passage into the terrestrial domain.
Thus, at the level of zone IV, the decrease of benthic biomass and the increase of phytoplanktonic biomass are demonstrated by a crossing of the two curves.
Insofar as the majority of the peri-Mediterranean lagoons are located within zones III, IV and V, this phenomenon is shown by a quantitative antagonism in the milieu between benthic macrofauna and phytoplankton. Thus, at the Bahiret el Biban, the confinement gradient results in phytoplnktonic biomass increases, and a benthic biomass decrease (Guelorget et al., 1981). This is also the case at Biguglia, for instance, where the inverse evolution of these two gradients is obvious (Frisoni el al., 1983). On the contrary, Diana and Urbino, not highly confined and less rich in chlorphyll than Biguglia, possess a greater weight of macrofaunistic communities. The single point made by the crossing of the two curves is likely to correspond to a characteristic (as yet not explained) of the organization and the working of the paralic domain. However, considering this single point, it is obvious that the very high primary biomass (essentially phytoplankton and microphytobenthos) shows a large excess with respect to what is consumed by the low biomass benthic populations where the filter-feeders have disappeared and only browsers and detritivorous species remain. Thus the major part of the chlorophyll biomass is unutilized. Besides, within these zone situated at the hydrological limits, the masses of water move extremely slowly, thus creating conditions which contribute to the accumulation of organic matter. The highly “abnormal” salinity of these zones contributes to the stratification of the water, the generalization of the reducing milieux, and the diagenetic preservation of this organic matter. Zones V and VI therefore create sediments liable to become “mother-rocks” for hydrocarbons.
The evaluation of benthic macrofauna production could only be approached with respect to pelecypods which have been the subject of a particular study in the Etang du Prévost (Guelorget and Michel, 1976; Guelorget et al., 1982).
This restriction to one zoological group is due to the impossibility of assessing the production of all the benthic species present in a lagoon, even though there are fewer species present than in the marine domain. It is impossible, with a reasonably workable sampling system, to collect enough individuals of each species to be able to assess their production. A great number of pelecypod, which are largely dominant in the Near paralic, appear in every sample of sediment and they offer reliable, easily measurable biometrical criteria, which is not the case for other groups, especially Polychaeta. It should also be remembered that pelecypod populations integrate the various factors of the milieu on a more or less long-term basis, owing to their relative immobility dependent on their connection with the bottom; they achieve this much better than the more mobile Polychaeta and crustaceans 1.
The study of the production curves (Table 4) is based on the study of five species which alone constitute almost the whole of the benthic macrofauna (Venerupis decussata, Scrobicularia plana, Abra ovata, Venerupis aurea, Cerastoderma glaucum) (Guelorget et al., 1980; Guelorget and Mayere, 1981 a and b, 1983 a and b).
The samples of sediment, taken monthly with a suction pump over an area of 0.5 m2, are sifted through a 1 mm grid. The measurements of the pelecypods (maximum antero-posterior diameter) are taken with callipers to the nearest millimetre. The biomass is measured in dry weight after decalcification. Production is expressed in g/m2/year (Table 5).
Within the malacological fauna, three groups of species can be distinguished:
- “Thalassic” species only found in the sea and in the vicinity of the grau which, because of their density and their biomass, represent only a minor fraction of the total macrofauna. They are not taken into consideration in the present study.
- “Paralic” species (Abra ovata and Cerastoderma glaucum whose distribution follows the general rule of the increase in density and the decrease in the size in conjunction with the rising gradient of confinement.
- “Mixed” species (Venerupis aurea, V. decussata, Scrobicularia plana) present both in the marine milieu and in the lagunar milieu. Here can be noted a decrease in density and size as the confinement increases, but they have a high or very high biomass, considering the size of the individuals.
After having compared the result obtained by various methods, preference was given to the Bojsen-Jensen method (1919) adapted by Masse (1968) to short time-intervals (a month). This simple method allows the comparison of different milieux and takes migratory phenomena into consideration.
The profiles of annual production (Fig. 30) show the extent of the exchange zone of the mouth from where the total production descends from 890 g/m2/year down to 75 g/m2/year within only 200 m (Station 15). Moving away from this zone, annual output stabilizes or decreases more slowly, for the paralic species begin to take over from the mixed species. Finally, towards the lagunar confines (Station 11), whereas the production of mixed species is practically nil, that of paralic species increases distinctly, but here the prononced “dwarfism” of the individuals reduces the effect of the increase in density, and the yearly production remains moderate (40 g/m2).
This general pattern is valid when considering the seasonal variations in output: particularly the vivified zone of the mouth remains the main producer all year-round (Fig. 31). Winter and spring correspond to the maximum outputs (respectively 60 and 54 g/m2/year on average), for both the mixed species in the vivified zone and the paralic species in the lagunar confines. Production decreases considerably in summer and autumn (respectively 19 and 13 g/m2/year on average) because of the dystrophic crises (“malaigues”) which affect mainly the paralic populations which are less soil-dwelling than the mixed species. In autumn, production is minimal, for it is the period of juvenile recruitment for all the pelecypod species, which yield only very little organic matter despite their high density.
The total annual output of the Etang du Prévost represents 171 000 kg of decalcified dry organic matter, which corresponds to a “marketable” quantity of around 1 700 t.
Further, the cartography of the annual overall production shows the economic advantages of the medium confinement zones (Zones II and III, Guelorget et al., 1982). Here, the malacological communities consist mainly of young individuals undergoing a phase of exponential weight increase, explained no doubt by the local concentration of larvae and the flow of nutriment originating in both the marine domain and the lagoon itself (Fig. 32).
Thus, the paralic domain shows capacities of production which are considerably superior to those of the marine milieu. Another fundamental characteristic is the small number of species constituting this production, which is moreover limited to a specific zone in the confinement area (moderate confinement zone where mixed species flourish).
The study of the soft-bottom benthic settlement of several Mediterranean lagunar milieux gave an insight into the biological organization of the paralic domain. Is the qualitative and quantitative biological zoning, established according to confinement and which applies to other biological compartments such as phytoplankton and ichthyofauna, also valid for benthic communities on hard substrate? The question is all the more important as hard substrates are rare or absent in the natural paralic milieax - especially in the lagunar milieux - and most of the lagoon-bred species are fixed species of considerable economic importance (oysters, mussels). Because of this lack of hard substrates in the paralic milieux, the authors followed the establishment and the qualitative and quantitative evolution of the benthic communities on bare artificial surfaces, immersed in the lagoons of Diana and Urbino, and the Etang du prévost : a general organization system of the fixed communities appears both with regard to the specific composition and to the characteristic values of the macrobenthofauna.
This is an exact parallel with the zoning calculated for the benthic populations of the sediment.
Zone I includes most of the thalassic species of the infracoastal communities of the Mediterranean rocky coastlines (Peres and Picard, 1964).
In zone II, the taxonomic diversity is considerably reduced and comprises only just over fifty thalassic and mixed species. Among these, the more commonly found are: Hydroides elegans, Serpula vermicularis, Bowerbankia imbricata, Bugula stolonifera, Anomia ephippium, Modiolus barbatus, Botryllus schlosseri (see Appendix 2). The plant communities are dominated by Bangia fuscopurpurea, Ceramium rubrum, Padina pavona, Cystoseira (various species), Acetabularia mediterranea, Codium vermilara, Halimeda tuns. In the vicinity of the boundary between zones II and III reef structures with Neogoniolithon notarisii may appear (Denisot et al., 1981).
In zone III, mainly the mixed suspension-feeding species develop: Mytilus galloprovincialis, Ostrea edulis, Avicula hirudo (Bahiret el Bou Grara), Ciona intestinalis, Styella plicata, Phalusia mamillata, Bugula neritina, Membranipora membranacea, Balanus eburneus, represent most of the fixed fauna. In the flora, various species of Enteromorpha and Ulva lactuca appear.
Zones IV and V are characterized by a small number of paralic species, and the community is dominated by the Cirripedia Balanus amphitrite amphitrite (associated with B. eburneus), the pelecypod Brachydontes marioni and the Polychaeta Mercierella enigmatica (Vuillemin, 1985), which is liable to produce reef-type structures, as for example in the northern lagoon of Tunis. The flora is almost exclusively composed of Chlorophyceae Enteromorpha gr intestinalis and Ulva lactuca locally associated with Gracilaria verrucosa.
All fixed macrofaune disappears in zone VI to give way to a cyanophytic covering (Lyngbya confervoides, Callotrix eruginea, C. scopulorum, Anabaena sp., Oscillartoria nigro-viridis).
These were analysed from data collected in the Etang du Prévost (Fig. 33) and were confirmed by studies on other Mediterranean lagoons, the Venice lagoon (Barbaro and Francescon, 1976; Francescon and Barbara, 1976) and lagoons of eastern Corsica.
From the communicating link with the sea towards the confines of the lagoon, these can be observed;
- a significant decrease in number of species
- a considerable increase in density
- a decrease in biomass, despite the increase of density, with regard to the small size of the individuals (lagunar dwarfism).
This pattern, identical to that of soft-bottom communities, remains the same whatever time the colonization of the immersed areas takes. However, it is obvious that both biomass and density increase with time at a given point.
As with the soft-bottom communities, the paucity of species, counter-balanced by a high density, generates an incomparably greated biomass than in the marine domain, especially in zone III where mixed species thrive. In this zone, after a year's immersion, at the Etang du Prévost can be collected a biomass of 11 000 g/m2 (in dry weight after decalcification), 96% of which consists of Mytilus galloprovincialis 1.
This shows how well-founded the empirical methods are, based on an ancestral knowledge of the milieu. They alllowed the development of conchyliculture in the paralic domain, whereas random trials, often carried out for political reasons without any previous knowledge of the host milieu, have never led to profitable production because they were situated in zones of inadequate confinement. The economic interest of the zoning of the paralic milieux can be understood in terms of confinement, since it allows the definition of optimal zones of conchylicultural - or even aquacultural - activity. If is even possible to envisage, through development work on the passes, the modification of a given basin with regard to the confinement scale.
Because confinement appeared to be the fundamental parameter of the organization of the paralic domain, the authors thought it necessary to propose an explanation and define its main levels. On the basis of the qualitative biological zoning of the benthic macrofauna, they have established a scale of confinement on which are expressed the various qualitative and quantitative parameters which describe the biological organization.
The arrangement proposed for the dynamics and the biological structure of the ecosystems of the Mediterranean paralic domain appears to apply to each of them, taking into consideration its morphological and hydrological particularities. Thus the different zones defined can vary as to their surface area and localization, but in all cases the zoning is respected.
The study of other biotic (meiofauna, microfauna) and abiotic (Appendix 3) parameters will permit to refine, and indeed perhaps enlarge towards the Far paralic, the zoning which has been established for the Near paralic Mediterranean domain.
However, the knowledge of extra-Mediterranean milieux such as the Caribbean mangrove swamps (Belle Plaine and Menche à Eau; Guelorget and Gaujous, 1983), the Red Sea lagoons (Guemsah; Ibrahim et al., 1982) and the Persian Gulf lagoons (Khour el Hadid lagoon; Perthuisot and Jauzein, 1978), and bibliographical data confirm the general nature of the proposed model (Appendix 6).
![]()
![]()
![]()