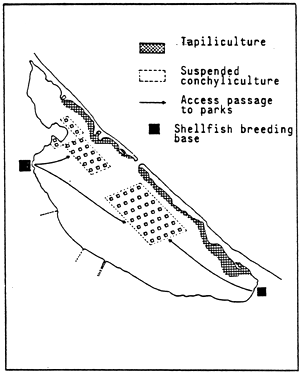
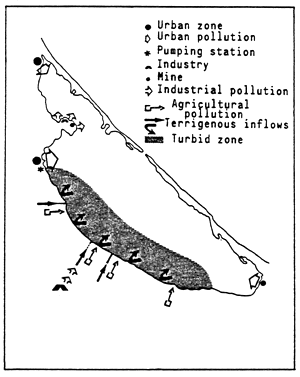
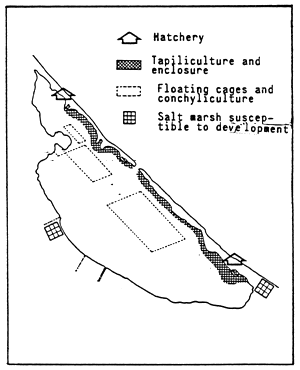
![]()
![]()
![]()
AGUESSE P. (1957). - La classification des eaux poïkilohalines, sa difficulté en Camargue; nouvelle tentative de classification. Vie Milieu. 8, 4, 341–365.
ALIMEN H. (1936). - Étude sur le Stampien du Bassin de Paris.Mém. Soc. Géol.Fr., (NS), XIV, 31, 309 p.
ALLEN G.P., LAURIER D. et THOUVENIN J. (1979). -Étude sédimentologique du delta de la Mahakam. Notes et mémoires CFP, Paris, 15, 156 p.
AMANIEU M. (1967). - Introduction à l'étude écologique des réservoirs à poissons de la région d'Arcachon. Vie Milieu, 18, 2B, 381–446.
(1973). - Écologie et exploitation des étangs et lagunes saumâtres du littoral français. Ann. Soc. roy. Zool. Belgique, 103, 1 79–94.
AMANIEU M. et LASSERE G. (1981). - Niveaux de production des lagunes littorale méditerranéennes et contribution des lagunes à l'enrichissement des pêches démersales. Étud. Rev.C.G.P.M., 58, 81–94.
AMANIEU M., EERRARIS J. et GUELORGET O. (1978). -Organisation et évolution des peuplements des lagunes littorales. Application à la macrofaune benthique des étangs palavasiens. Séminaire d'Ecologie. Laboratoire de Zoologie, ENS, Paris, mai 1978, 251–277.
ANCONA (d') U. (1959). - The classification of brackish waters with reference to the North Adriatic Lagoons. Arch. Oceanogr. Limnol. Venizia, 11, 93–109.
ANDERSIN A.-B., LASSIG J. et SANDLER H. (1976). -Community structures of soft-bottom macrofauna in different parts of the Baltic. in KEEGAN B.F., CEI-DIGH P.O. et BOADEN P.J.S. (ed.): Biology of benthic organisms. Eleventh European Symposium on Marine Biology, Galway, October 1976. Pergamon Press, London.
ARNAUD P. et RAIMBAULT R. (1969). - L'étang de Salses-Leucate. Ses principaux caractères physicochimiques et leurs variations (en 1955–56 et de 1960 à 1968). Rev. Trav. Inst. Péches marit., 33, 4, 335–443.
BARBARO A. et FRANCESCON A. (1976). - I periodi di insediamento dei principali organismi del fouling nelle acque di Venezia. Arch. Oceanogr. Limnol., 18, 195–216.
BARBARE L. et FEYS R. (1965). - Géologie du charbon et des bassins houillers. Masson, Paris, 229 p.
BASSON P.W., BURCHARD J.E., HARDY J.T. et PRICE A.R.G. (1977). - Biotopes of the Western Arabian Gulf. Marine life and environments of Saudi Arabia. ARAMCO, Dharan, 284 p.
BELSER W.L. (1963). - Bioassay of trace substances. In Hill M.N. ed. The Sea. Ideas and observations in the study of the seas. Intersciences publishers, N.Y., London 2, 220–231.
BLANDIN P., BARBAULT R. et LECORDIER C. (1976). -Réflexions sur la notion d'écosystème: le concept de stratégie cénotique. Bull. écol., 7, 4, 391–410.
BOCK K.H. (1971). - Monatskarten der Temperatur des Ostsee. Dargestellf für Verschiedene Tiefenhorizonte, 147 p.
BOJSEN-JENSEN P. (1919). - Valuation of the Limfjord-1. Studies on the fish food in the Limfjord, 1909–1917, its quantity, variation and annual production. Rep. Dan. Biol. Stat., 26, 1–44.
BOUQUEGNEAU J.M. et MARTOJA M. (1982). - La teneur en cuivre et son degré de complexation chez quatre gastéropodes marins. Données sur le cadmium et le zinc. Oceanologica Acta 5, 2, 219–228.
BOYE M., MARMIER F., NESSON C. et TRECOLLE G. (1978). - Les dépôts de la Sebkha Mellala. Rev. Geomorph. dynam. XXVII, 2–3, 49–62.
BURELLI F., ECREMENT F., FRISONI G.F., GUELORGET O., XIMENES M.C. (1979). - Etude des éco-systèmes des Biguglia, Diana et Urbino. Rapport SOMIVAC-GTGREF, 1979, ronéo., 60 p.
BUSSON G. (1972). - Principes, méthodes et résultats d'une étude stratigraphique du Mésozïque saharien. Mém. Mus. Nat. Hist. Nat. Paris C, 26, 441 p.
BUSSON G. et PERTHUISOT J. P. (1977). - Intérèt de la Sebkha el Melah (Sud Tunisien) pout l'interprétation des séries évaporitiques anciennes. - Sediment. Geol, 19, 139–164.
BUROKER N.E., HERSHBERGER W.K. et CHEW K.K. (1979). - Population genetics of the family Ostreidia. II. Inter-specific studies of the genera Crassostrea and Saccostrea . Mar. Biol. 54, 2, 157–169
CARBONNEL P. (1982). - Les Ostracodes, traceurs des variations hydrologiques dans systèmes transitionnels eaux dources - eaux salées. Mém. Soc. Géol. Fr. (NS). 144, 117–128
CHAUVET C. (1979). - Préliminaire à rétude de la biologie et de la dynamique du stock tunisien de Sparus aurata (L.1758), Pisces, Sparidae. Synopsis de la croissance. Groupes 0+, 1+, 2+. Bull. off. natn. Péches, Tunisie, 3,2 ,241–253.
CLOUD P (1968). - Atmospheric and hydrospheric Evolution of the Primitive Earth. In (1970) CLOUD p. (Ed.) Adventures in Earth history, Freeman & C°, San Franciso, 446–457.
COQUE R. (1962). - La Tunisie Présaharienne, Étude géomorphologique . A. Colin, Paris, 476 p.
CORNEE A. (1983). - Sur les bactéries des saumures et des sédiments de marais salants méditerranéens. Importance et rôle sédimentologique. Doc. GRRECO 52 (Nature et genèse des faciès confinès,), 3, 126 p.
DENIZOT M., GUELORGEL O., MASSIEUX M. et PERTHUISOT J.-P. (1981). - Une remarquable construction récifale à Mélobésiées dans une lagune sursalée du SE tunisien (La Bahiret el Biban). Cryptogamie: Algologie, II, 4, 253–266.
DOTY M.S. (1957). - Rocky Intertidal Surfaces. In HEDGPETH J. W. (Ed.): Treatise on Marine Ecology and Paleoecology. Mem. Geol. Soc. America, 67, 1, 535–585.
DZENS-LITOVSKU A.I. (1956). - Le passé géologique et le
temps présent du Kara-Bogaz-Gol. - Dokl. ezhegod.
Chen. Pam. L.Sl. Berga, U.R.S.S., 1–3 (1952–54),
131–169.
(1962). - La Mer Caspienne et le golfe du Kara-Bogaz-Gol.
- lzv. vsesojuvn. geogr. Obshch,, 94, 1,
34–44 (russe; trad. B.R.G.M. № 3747).
FALQUE M., GALLAND A. et TARLET J. (1975). - La planificaton écologique, méthode d'intégratoin des données du milieu naturel en matière d'aménagement de l'espace. Le monteur des Travaux Publies et du Bàtiment, 29 mars 1975, 12–23.
FRANCEDSCON A. et BARBARO A. (1976). - Distrbuzione del fouling lungo i canali Marghera-Mare. Atti. Ist. Veneto Sci., 134, 33–45.
FRSONI G-F., GUELORGEL O., XIMENES M.-C et PERTHUISOTT I.-P. (1983). - Étude écologique de trois lagunes de la plaine orientale corse (Biguglia, Diana, Urbino): expressions biologiques qualitatives et quantitatives du confinement. Journ. Rech. Oceanogr., VIII, 1, 57–80.
FRISONI G.-F., GUELORGET O., PERTHUISOT J.-P., XIMENES M.-C. et MONTI D. (1982). - Méthodologie des études d'impact de l'aquaculture en milieu lagunaire. Rapport IARE présenté àla CEE, novembre 1982, ronéo, 187 p.
FURON R. et SOYER R. (1947). - Catalogue des fossiles tertiaires du Bassin de Paris. Savoir en Histoire Naturelle , XXII, Paul Lechevalier, Paris, Guides techniques du naturaliste, VI 240 p.
GAUDANT J. (1982). - Apport de l'ichthyofaune à la caractérisation des milieux saumâtres cénozoïques. Mém. Soc Geol. Fr. (NS). 144, 139–146.
GAUJOUS D. (1981). - Étude écologique de la macrofaune benthique des mangroves de Guadeloupe. Mémoire d'Ingéniorat S.T.E., Montpellier. 108 p.
GAUJOUS D. et GUELORGET O. (1983).- Étude écologique des lagunes de mangroves de Guadeloupe. Publication en cours.
GOLDBERG E.D. (1963). - The oceans as a chemical system. In Hill M.N. (ed.). The Sea Ideas and observations in the study of the seas. Intersciences Publishers, N.Y.- London, 2,3–20.
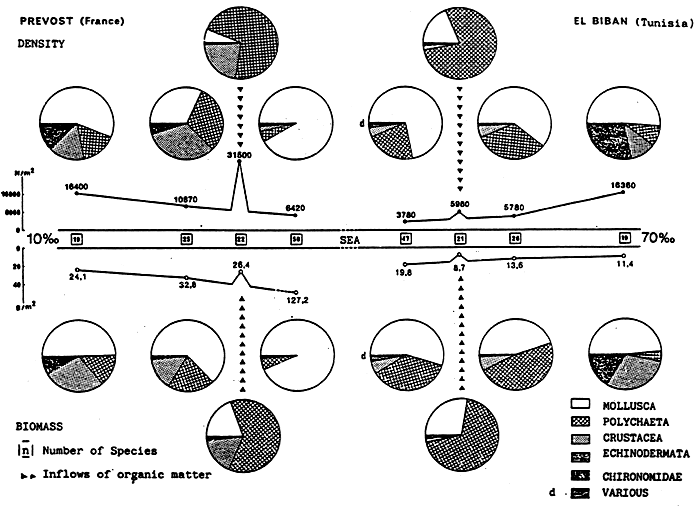
GUELORGETO. et MAYERE C. (1981a). - Croissance, biomasse
et production d'Abra ovata dans une lagune
méditerranéenne, l'Étang du Prévost à Palavas (Hérault,
France). Journ, Rech, Océanogr., VI, Bull. 3 et 4.
(1981 b). - Croissance, biiomasse et production d'Abra
ovata dans l'Etang du Prévost. Rapp. Comm. int. Mer.
Médi, 27, 4, 137–138.
(1982 a). - Croissance, biomasse et production de
Cerastoderma edule dans une lagune méditerranéenne,
l'Étang du Prévost (Hérault, France). Téthys (à paraître).
(1982 b). - Croissance, biomasse et production de
Scrobicularia plana dans lagune méditerranéenne,
l'Étang du Prévost à Palavas (Hérault, France). Téthys
(àparaître).
GUELORGET O. et MICHEL P. (1976). - Recherches écologiques sur une lagune saumâtre méditerranéenne, l'Étang du Prévost (Hérault) - I Le milieu (95 p.)., II Les peuplements benthiques (122 p.). Thèse de spécialité U.S.T.L. Montpellier.
GUELORGET O. et PERTHUISOT J.-P. (1982). - Structure et évolution des peuplements benthiques en milieu paralique. Comparaison entre un modèle dessalé (l'Étang du Prévost, France) et un modèle sursalé (la Bahiret el Biban, Tunisie). Conséquences biologiques et géologiques. Journ. Rech. Océanogr. VII, 2.3.4., 2–II.
GUELORGET O., FRISONI G.-F et PERTHUISOT J.-P.
(1981). - Les communautés phytoplanctoniques et
benthiques d'un milieu paralique hypersalé: la Bahiret
el Biban (Tunisie). Critères d'analyse du fonctionnement
d'un écosystème lagunarie. Communication présentée
au Symposium international sur les lagunes
côtières (ISCOL) UNESCO, Bordeaux, septembre
1981.
(1982a). - Contribution à l'étude biologíque de la
Bahiret el Biban, lagune du Sud-Est tunisien. Symposium
SGF et ASF, «Transition eaux douces - eaux
salées.» Mém. Soc. Géol. Fr., N.S., 1982, № 144,
p. 173–186.
(1982b). - Structure et fonctionnement d'un écosystème-type
du domaine paralique méditerranéen.
Communication présentée au Congrès de la CIESM,
Cannes, décembre 1982 (à paraître).
(1983a). - La zonation biologique des milieux lagunaires:
définition d'une échelle de confinement dans
le domaine paralique méditerranéen. Journ. Rech.
Océanogr./, VIII 1, 15–36.
(1983b). - Caractérisation écologique des milieux en
domaine paralique, critère de choix des paramètres
utiles. Bull. Écol. (Sous presse).
GUELORGET O., MAYERE C. et AMANIEU M. (1980). - Croissance, biomasse et production de Venerupis decussata et Venerupis aurea dans une lagune méditerranéenne, l'Étang du Prévost à Palavas (Hérault, France). Vie marine 2. 25–38.
GUELORGET O. PERTHUISOT J.-P. et FRISON G.-F. (1983). - Contribution à l'étude écologique du Lac Melah (Algérie). Rapport FAO-GREDOPAR (à paraître).
GUELORGET O., MAZOYER-MAYERE C., PERTHUIISOT J.-P et AMANIUL M. (1982). - La production malacologique d'une lagune méditetrranéenne: l'Etang du Prévost, Communication présentée au Congrès de la CIESM, Cannes, décembre 1982 (à paraître).
HARTOG (Den) C. (1971). - De Naderlandse Ruppiasoorten. Gorteria, S, 7/10, 148–153.
IBRAHM A., GUELORGET O. et PERTHUSOT J.-P. (1982). - Contribution à l'étude hydrologique et sédimentologique de la lagune de Guemsah. Rapport CFP-CREDOPAR, 36 p.
JAUZEIN A. (1982). - Deutérium et oxygène 18 dans les saumures: modélisation et implications géologiques. C.R. Acad. Sc. Paris, II, 294, 663–668.
JONES L.H., JONES N.V. et RADLETT A.J. (1976). -Some effects of salinity on the toxicity of copper to the polychaete Nereis diversicolor. Estuarine and coastal marine Science, 4, 107–111.
JOUVENIAUX A.et PALANCHON A. (1978). - L'étude d'impact en France: éléments de pathologie. Pub. Ministère de l'Environment en du Cadre de Vie. (Service technique de l'Urbanisme). 255 p.
KIENER A. (1978). - Écologie, physiologie et économie des eaux saumâtres. Coll. de Biologie des Milieux Marins, 1, Masson, Paris, 220 p.
KUMARAGURU A.K. et RAMAMOORTHIS K. (1979). -Accumulation of copper in certain bivalves of Vellar Estuary, Porto Novo, S. India, in natural and experimental conditions. Estuarine and Coastal marine Science 9, 467–475.
LAGNY P. (1980). - Les gisements stratiformes associés aux évaporites: position dans le temps et place dans l'espace des bassins sédimentaires évaporitiques. Bull. Centre Rech. Explor. Prod. Elf Aquitaine, 4, 1, 445–478.
LEGENDRE L. et LEGENDRE P. (1979). - Écologie numérique. 2: La structure des données écologiques. Masson, Paris. 254 p.
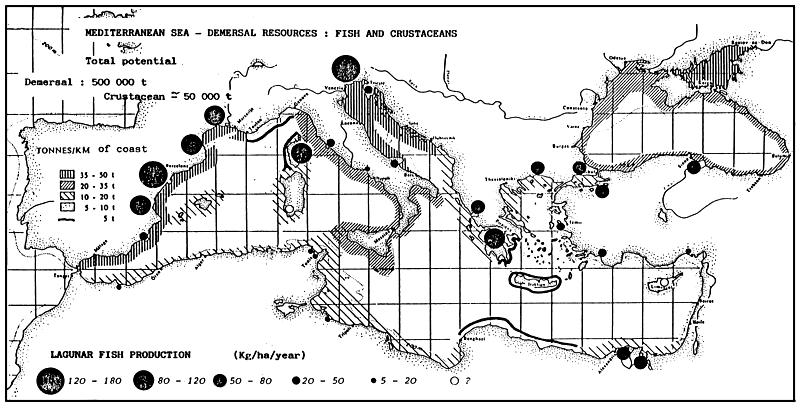
LEVI D. et TROADEC J.-P (1974). - Les ressources halieutiques de la Méditerranée. Étud. Rev. CGPM, 54, 29–52.
LEVY A. (1970). - Contribution à l'étude des milieux margino-littoraux. Influence des facteurs hydrologiqus et
sédimentologiques sur les peuplements microfaunistiques.
Thèse d'État. Université P. et M. Curie, Paris,
multig. 450 p.
(1982). - Sur la survie de certains formainifères dans
les eaux continentales et sur ses conséquences. Mém.
Soc. Géol. Fr. (NS), 144, 161–171.
LUOMA S.N. et BRYAN G.W. (1982). - A statistical study of environmental factors cntrolling concentrations of heavy metals in the burrowing Bivalve Scrobicularia plana and the Polychaete Nereis diversicolor. Estuarine, Coastal and Shelf Science 15, 95–108.
MARAZANOF F. (1972). - Contribution à l'étude écologique des Mollusques des eaux douces et saumâtres de Camargue. Thèse d'État. Fac. Sci. Orléans, 213 p.
MARS P. (1966). - Recherches sur quelques étangs du littoral méditerranéen et sur leurs faunes malacologiques. Vie Milieu, 20, 359 p.
MASSE H. (1968). - Sur la productivité des peuplements marins benthiques. Cah. Biol. mar., IX. 363–372.
MEDHIOUB K. (1979). - La Bahiret el Biban. Étude géochimique et sédimentologique d'une lagune du Sud-Est tunisien. Trav. Lab. Géol. École norm sup., Paris, 13, 150 p.
MEDHIOUB K. et PERTHUISOT J.P. (1977). - Le comportement géochimique des eaux de la Bahiret el Biban
Ses conséquences sur la pécge, Bull. Off. natn. péches
de Tunisie, 1,1, 23–35.
(1981). - The influence of peripheral sabkhas on the
geochemistry and sedimentology of a Tunisian lagoon:
Bahirel el Biban. Sedimentology, 28, 679–688.
MERCIER A. (1973).- Étude écologique de la végétation du complexe lagunaire de Bages-Sigean. Biomasse et production primaire des Macrophytes. Thèse 3e cycle. Université P. et M. Curie, Paris, 105 p.
MORRIS R.C. et DICKEY P.A. (1975). - Modern evaporite deposition in Peru. AAPS Bull. 41, 2461-–2474.
MUUS B.J. (1967). - The fauna of Danish estuaries and lagoons. Distribution and ecology of the dominating species of the shallow beaches of the mesohaline zone. Medd. Komm. Havundersøg., Kbh. (Fisk), N.S., 5(I), 1–316.
NAUMAN C.P. (1954). - Lehrbuch der Geognosie, II, Leipzig.
PERES J.M. et PICARD J. (1964). - Nouveau manuel de bionomie benthique de la Mer Méditerranée. Recl. Trav. St. mar. Endoume. 31 (47): 137 p.
PERTHUISOT J.-P. (1971). - Présence de la magnésite et de
hutite dans la Sebkha el Melah de Zarzis. C.R. Acad.
Sci. Paris, 272, 185–188.
(1975). - La Sebkha el Melah de Zarzis. Genèse et
évolution d'un bassin salin paralique. Trav. Lab.
Géol. École norm. sup., Paris, 9, 252 p.
(1980). - Sites et processus de la formation d'évaporites
dans la nature actuelle. Bull.Centre Rech. Explor.-Prod.
Elf-Aquitaine, 4, 1, 207–233.
(1983). - Introduction générale des marais salants de
Salin-de-Giraud (Sud de la France). Le cadre géographique
et le milieu. Géol. Médit. (à Paraître).
PERTHUISOT J.P. et GUELORGET O. (1982a). - Le domaine
paralique: dynamique biologique et sédimentaire.
Comm. XXIe Congr. Intern. Sédimento. Hamilton
(Canada), Abstracts, p. 26.
(1982b). - Le confinement, paramètre essentiel de la
dynamique biologique du domaine paralique (réunion
GRECO 52, mai 1982). Sciences Géologiques (à paraître).
PERTHUISOT J.P. et JAUZEIN A. (1975). - Sebkhas et
dunes d'argile: l'enclave endoréïque de Pond du Fahs,
Tunisie. Riv. Géogr. Phys. Géol. Dynam. Paris, (2),
WVII, 3, 295–306.
(1978). - Le Khour el Aadid, lagune sursalée de
l'Émirat de Qatar. Rev. Geogr. Phys. Geol. Dynam.
Paris, (2), XX, 4, 347–358.
PERTHUISOT J.P., GUELORGET O., FRISONI G.F. et MAURIN A. (1983). - Expressions hydrochimiques, sédimentologiques et biologiques du gradient de confinement dans un partènement salinier abandoné: le Triangle des Bermudas (Santa Pola, Espagne). Journ. Rech. Océanogr., VIII, 1, 38–56.
PETIT G. (1953). - Introduction à l'étude écologique des
étangs méditerranéens. Vie Milieu, 4(4), 569–604.
(1962). - Quelques considérations sur la biologie des
eaux saumâtres méditerranéennes. Publ. Staz. Zool.
Napoli, 32, 205–218.
PETIT G. et SCHACHTER D. (1959). - Les étangs et les lagunes du littoral méditerranéen et le problème de la classification des eaux saumâtres. Ocean. Limnol. 11, 75–91.
PETIT-MAIRE N. (et coll.) (1982). - Le Shati, lac pleistocène du Fezzan (Libye). Ed. CNRS, Paris, 118 p.
PEYPOUQUET J.P. (1977). - Les ostracodes et la connaissance des paléomilieux profonds. Apllication au Cénozoïque de l'Atlantique oriental. Thèse Sc. Nat. Bordeaux I, 1452, 191 p.
PIELOU E.C. (1966). - The measurement of diversity in different types of biological collections. J. Theoret. Biol., 13, 131–144.
PLAZIAT J.C. (1982). - Introduction à l'écologie des milieux de transition eau douce-eau salée par l'identification des paléoenvironnements correspondants. Critique de la notion de domaine margino-littoral. Mém. Soc. géol. Fr., NS. 144, 187–206.
POMEROL C. (1973). - Stratìgraphie et Paléogèographie. Ère Cénozoïque (Tertiaire et Quaternaire). Doin, Paris, 270 p.
PROVASOLI L. (1963). - Organic regulation of phytoplancton fertility. In Hill M.N. (ed.). The Sea, Ideas and observations in the study of the seas. Intersciences publishers, N.Y., London, 2. 165–219.
QUEMENEUR J. (1974). - Les gisements de magnésite du Pays Basque: Cadre géologique et sédimentologique. Genèse de la magnésite en milieu sédimentaire. Thèse 3e Cycle, Université, P. et M. Curie, 275 p.
QUIGNARD J.P. et ZAOUALI J. (1980). - Les lagunes périméditerranéennes. Bibliographie ichthyologique annotée. Bull. Off. nam. Péches. Tunisie, IV, 2, 293–360.
RELINI G., BARBARO A. et FRANCESCON A. (1972). -Distribuzione degli organismi del fouling in relazione all'inquinamento urbano di Venezia, Atti. Ist. Veneto. Sci., 130, 433–448.
REMANE A. et SCHLIEPER C. (1956). - Die Biologie des Brackwassers. Die - Binnengewässer, Stuttgart, 22. 348 p.
RIOUALL R. (1981). - Étude quantitative des algues macrophytes de substrat meuble de l'étang du Prévost (Hérault). Naturalia monspeliensa ser. bot., 26, 73–94.
ROUCHY J.M. (1981). - La genèse des évaporites messiniennes de Méditerranée. Thèse d'Etat, Mus. Nat. Hist. Nat. et Univ. P. et M. Curie, 295 p.
ROUTHIER P. (1980). - Où sont les métaux pour l'avenir? Les provinces métallogéniques. Essai de métallogénie globale. Mém. BRGM, Orléans, 105, 320 p.
RUTTNER-KOLISKO A. (1971). - Rotatiorien als Indikatoren für den chemismus von Binnensalzgewässern. Abhandlung des Natrongewässer - Symposiums Tihany-Szeged-Szarvas. 12, 283–298.
SACCHI C.F. (1967). - Rythmes des facteurs physicochimiques du milieu saumâtre et leur emploi comme indice de production in Problèmes de productivité biologique. Masson, Paris, 131–158.
SEGERSTRÅLE S.G. (1957). -Baltic sea. In HEDGPETH
J.W. (Ed). Treatise on Marine Ecology and Palcocecology.Mem.
Geol. Soc.America, 67, 1, 751–800.
(1959). - Brackish waters classification. A historical
survey. Arch.Oceanogr. Limnol., Venezia, 11, 7–13.
THOMAS J.C. (1983). - Peuplements benthiuqes à Cyanophycées des marais salants de Salin-de-Giraud (Sud de la France). Geol.Médit. (à paraître).
TOULEMONT M. (1980). - Les gypes lutétiens du Bassin de Paris. Bull. Cent. Rech. Explor. - Prod. Elf-Aquitaine , 4, 1, 245–279.
VACELET J. (1983). - Les éponges clacifiées et le srécifs anciens. Pour l aScience, ,86 14–22.
VATOVA A. (1963). - Conditions hydrographiques et productivité dans la lagune moyenne de Venise. Rapp. Comm. Int. Mer Médit., 17, 3, 753–755.
VUILLEMIN S. (1965). - Contribution à l'étude écologique du Lac de Tunis. Biologie de Mercierella enigmatica Fauvel. Thèse d'Etat. Université Paris VI. 554 p.
WORMS J. et PASTEUR N. (1982). - Polymorphisme biochimique de la palourde, Venerupis decussata, de l'Étang du Prévost (France). Oceanologica Acta., 5, 4, 395–397.
XIMENES M.C. (1980). - Observations sur les faunes ichthyologiques des étangs corses: Biguglia, Diana et Urbino (inventaire, alevinage, croissance et démographie de certaines espèces). Mém. Ingéniorat, USTL, Montpellier. Ronéo. 98 p.
ZANINETTI L. (1983). - Les foraminifères. Milieu de vie et transport dans le marais salant de Salin-de-Giraud. Comparaison avec les microfaunes marines. Géol. Médit. (à paraître).
ZENKEVITCH L.A. (1957). - Caspian and Aral seas. In HEDGPETH J.W. (Ed.): Treatise on Marine Ecology and Paleoecology. Mem. Geol. Soc. America, 67, 1, 891–916.
GFCM/CGPM. (1980). - GFCM Statistical Bulletin: nominal catches. Bulletin statistique du CGPM : captures nominales. 1968–78. GFCM stat. Bull./Stat. Bull. CGPM, 3, 124 p.
Taxonomic resources of the phytoplankton in various Mediterranean paralic basins.
| Basin | Taxonomic resources | Prevailing confinement zones |
| Lagoon of Diana | 41 | II (III) |
| Mar Chica | 29 | II |
| Lagoon of Mauguio | 13 | IV (V) |
| Lake Melah | 10 | IV - V |
| Bermuda Triangle | 8 | IV - V (VI) |
| Lagoon of Biguglia | 27 | V - VI |
| (freshwater influences) |
From FRISONI's data, personal communication
Number of species and diversity of the ichthological populations of the lagoons of the eastern plain of Corsica. From FRISONI et al., 1983.
| BIGUGLA | DIANA | URBINO | ||||||||
| Mouth | Centre of lagoon | South | North | North | South | |||||
| Sedentary | 78 % | 82.0 % | 94 % | |||||||
| Migrant | 21 % | 10.4 % | 4 % | |||||||
| Occasionals (% of the captures*)In numbers | 1 % | 7.6 % | 2 % | |||||||
| M/S * Ratio (numbers) (1981) | 0.31 | 0.07 | 0.09 | 0.15 | 0.03 | 0.02 | ||||
| Number of species*(Migrant + sedentary) (numbers) | 16 | 11 | 23 | 20 | 20 | 18 | ||||
| ESH (numbers) | Maximum | 0.659 (Spring) | 0.716 (Summer) | 0.558 (Summer) | ||||||
| Minimum | 0.463 (Winter) | 0.304 (Winter) | 0.342 (Winter) | |||||||
“Sedentary” species : percentages of captures
(Over one year)
| BIGUGLIA | DIANA | URBINO | |||||
| Mouth | Centre | North | South | Mouth | North | South | |
| Year | 81 | 81 | 81 (79) | 81(79) | 81 | 81 | 81 |
| Atherina | 26 | 61 | 83(87.8) | 68(71.3) | 38 | 65 | 72 |
| Aphanius | 36 | 20 | 13(2) | 17(23) | 59 | 29 | 25 |
| Syngnathus | 1 | 18 | 2(3.6) | 9(2.3) | 1 | 4 | 1.5 |
| Mottled Goby | 38 | 1 | 2(6.6) | 6(13.2) | 2 | 2 | 1.5 |
| weight | |||||||
| Atherina | 58 | 81.5 | 89 | 87 | 51.5 | 75 | 75 |
| Aphanius | 24 | 15.0 | 9 | 12 | 47.5 | 20 | 23 |
| Syngnathus | 0.5 | 2.5 | 1 | 0.5 | 0.5 | 2.5 | 1.5 |
| Mottled Goby | 18 | 1 | 1 | 1 | 0.5 | 2.5 | 0.5 |
Quantity of Molluscs in the Etang du Prévost
N = Density in number of individuals per m2
B = Biomass in grammes of decalcified dry weight per m2
| Fine sands | Muddy sands | Lagunar muds | ||||||||
| St. X | St. 15 et 16 | North. emis. S t. 7 | West. bas. | East. bas. | ||||||
| Molluscs | N | B | N | B | N | B | N | B | N | B |
| 9052 | 401.3 | 3083 | 33.8 | 1083 | 15.7 | 4031 | 8.4 | 919 | 8.3 | |
| Total fauna | 9716 | 415.4 | 4010 | 44.9 | 6282 | 22.6 | 7331 | 16.9 | 2851 | 14.9 |
Average values of density and biomass, annual production of the main Pelecypod Molluscs at different stations in the Etagne du Prevost
| Station X | Station 16 | Station 15 | Station 7 | |||||||||||||
| N | B | P | P/B | N | B | P | P/B | N | B | P | P/B | N | B | P | P/B | |
| Venerupis decussata | 641 | 56,7 | 221,7 | 3,9 | 427 | 8,6 | 30,4 | 3,5 | 139 | 0,7 | 2,2 | 3,1 | ||||
| Venerupls aurea | 2705 | 54,6 | 190,9 | 3,5 | 1546 | 39,8 | 65,4 | 1,6 | 1301 | 13,5 | 45,3 | 3,3 | ||||
| Scrobicularia plana | 4901 | 122,9 | 451,9 | 3,7 | 1952 | 6,3 | 34,3 | 5,4 | 1162 | 5,1 | 16,2 | 3,2 | 322 | 9,6 | 29,1 | 3,0 |
| Cerastoderma glaucum | 189 | 7,4 | 24,2 | 3,3 | 68 | 1,3 | 0,1 | 0,1 | 234 | 2,6 | 8,4 | 3,2 | 428 | 9,8 | 43,9 | 4,5 |
| Abra ovata | 312 | 0,5 | 1,0 | 1,9 | 561 | 1,6 | 4,3 | 2,7 | 506 | 1,4 | 5,3 | 3,8 | ||||
| TOTAL | 8436 | 241,6 | 888,7 | 4305 | 56,5 | 131,2 | 3397 | 23,5 | 76,4 | 1256 | 20,8 | 78,3 | ||||
| Station 12 | Station 11 | Station 4 | Station 3 | |||||||||||||
| N | B | P | P/B | N | B | P | P/B | N | B | P | P/B | N | B | P | P/B | |
| Venerupis decussata | 11 | 0,2 | 0,6 | 3,0 | ||||||||||||
| Veverupis aurea | 30 | 0,5 | 1,4 | 2,8 | ||||||||||||
| Scrobicularia plana | 22 | 1,1 | 4,4 | 4,0 | ||||||||||||
| Cerastoderma glaucum | 2401 | 11,0 | 26,8 | 2,4 | 6071 | 8,0 | 38,2 | 4,8 | 613 | 9,4 | 22,3 | 2,4 | 260 | 4,4 | 10,9 | 2,5 |
| Abra ovata | 503 | 1,2 | 6,4 | 5,3 | 1220 | 0,8 | 2,5 | 3,0 | 492 | 2,6 | 6,1 | 2,3 | 199 | 0,7 | 2,8 | 4,1 |
| TOTAL | 2904 | 12,2 | 33,2 | 7291 | 8,8 | 40,7 | 1168 | 13,8 | 34,8 | 459 | 5,1 | 13,7 | ||||
N : Number of individuals per m2
B : Weight of dry organic matter in g/m2
P : Production of dry organic matter in g/m2/year.
Some heavy metal concentrations measured in the tissues (whole animal) of marine and/or paralic species (in ppm of the dry weight)
| AG | Cd | Co | Cu | Zn | Pb | |
| Monodonta crassa (1) | 1,1 | 33 | 30 | |||
| Littorina littorea (1) | 1,6 | 45 | 50 | |||
| Thais lapillus (1) | 28,5 | 198 | 535 | |||
| Murex brandaris (1) | 32,2 | 270 | 285 | |||
| Anadara granosa (2) | 56 | |||||
| Meretrix casta (2) | 44 | |||||
| Crassostrea madrasensis (2) | 99 | |||||
| Ostrea edulis (3) | 1,8 | 90 | 710 | 0,1 | ||
| Mytilus galloprovincialis (3) | 0,5 | 2 | 36 | 0,2 | ||
| Scrobicularia plana (4) | 122 | 32,4 | 55 | 372 | ||
| Nereis diversicolor (4) | 8 | 3 | 25 | 947 | ||
| Sea water (ppm) (5) | 0,0003 | 0,0001 | 0,0005 | 0,0003 | 0,01 | 0,00003 |
1 From BOUQUEGNEAU and MARTOJA (1982)
2 From KUMARAGURU and RAMAMOORTHI (1979)
3 From BURELLI et. al., (1979)
4 From LUOMA and BRYAN (1982)
5 From GOLDBERG (1963)
Comparison of the number of species in the Caspian fauna and the Mediterranean fauna
| Group | Mediterranean Sea | Caspian Sea | ||
| Number of species | % of total fauna | Number of species | % of total fauna | |
| Echinodermata | 101 | 1,7 | 0 | 0 |
| Protozoa | 138 | 2,3 | 3 | 0,7 |
| Polychaeta | 433 | 7,2 | 5 | 1,2 |
| Pelecypoda | 366 | 6,1 | 16 | 4,0 |
| Gastropoda | 937 | 15,6 | 26 | 6,5 |
| Superior Crustaceans | 620 | 10,3 | 90 | 22,5 |
| Fish | 529 | 8,7 | 71 | 19,0 |
Monthly variations of the total lagunar algal (macrophyte) biomass (expressed in tonnes of drained wet weight). Etang du Prévost. From RIOVALL, 1976.
| WEST BASIN | CENTRAL BASIN | EAST BASIN | OVERALL LAGOON | |
| 25 April 1975 | 900 | 600 | 3700 | 5200 |
| 20 June 1975 | 1700 | 1800 | 3700 | 7200 |
| 23 July 1975 | 600 | 1800 | 1600 | 4000 |
| 25 August 1975 | - | - | - | - |
| 10 September 1975 | - | - | - | - |
| 28 October 1975 | 300 | 300 | 6300 | 6900 |
| 26 November 1975 | 400 | 200 | 2600 | 3200 |
| 7 January 1976 | 700 | 300 | 2900 | 3900 |
| 16 February 1976 | 600 | 200 | 1700 | 2500 |
| 18 March 1976 | 1200 | 800 | 3700 | 5700 |
Monthly variations of the lagunar algal biomass (tonnes) for the four principal types. Etang du preévost From RIOVALL, 1976.
| ULVA | ENTEROMORPHA | GRACILARIA | STICTYOSIPHON | |
| 25 April 1975 | 3900 | - | 100 | 1000 |
| 20 June 1975 | 6800 | - | 200 | - |
| 23 July 1975 | 3800 | - | - | - |
| 25 August 1975 | - | - | - | - |
| 10 September 1975 | - | - | - | - |
| 28 October 1975 | 200 | 6250 | 150 | - |
| 26 November 1975 | 350 | 2700 | 100 | - |
| 7 January 1976 | 1000 | 2200 | 150 | 100 |
| 16 February 1976 | 1000 | 1200 | 150 | 50 |
| 18 March 1976 | 4600 | 800 | 50 | 100 |
Present-day stratigraphic classification of the Paleogene formations in the Paris Basin (POMEROL, 1973), and equivalences with the classification of FURON and SOYER 1947).
| Pomerol (1973) | Furon et Soyer (1947) | ||
Layers | Sub-layers | Layers | |
| AQUITANIAN | CHATTAIAN | ||
STAMPIAN s.l. | STAMPIAN s.s. | STAMPIAN | |
| SANNOISIAN | SANNOISIAN | ||
| BARTONIAN | LUDIAN | LUDIAN | |
| MARINESIAN | BARTONIAN | ||
| ADVERSIAN | |||
LUTETIAN | LUTETIAN | ||
YPRESIAN | CUISIAN | CUISIAN | |
| SPARNACIAN | SPARNACIAN | ||
THANETIAN | THANETIAN | ||
| DANO-MONTIAN | MONTIAN | ||
Composition of total thalassoid fauna for each of the Paleogene layers of the Paris Basin From FURON and SOYER's data, 1947.
The continental forms (Limnaeidae, Helicidae, Pupidae, etc …) were not counted.
| MONT. | THAN. | SPAR. | CUIS. | LUTE. | BART. | LUDI. | SANN. | STAM. | CHAT. | |
| Foraminifera | 4 | 1 | 3 | 8 | 10 | 1 | ||||
| Coelenterata | 10 | 2 | 30 | 23 | ||||||
| Echinodermata | 10 | 72 | 2 | 2 | 1 | |||||
| Bryozoa | 1 | 23 | 130 | 12 | 9 | |||||
| Brachiopoda | 2 | 2 | 18 | 2 | ||||||
| Pelecypoda | 21 | 137 | 14 | 340 | 550 | 380 | 44 | 2 | 118 | |
| Gasteropoda ( )Hydrobiidae | 45 | 229 (7) | 56 (11) | 555 (10) | 1136 (19) | 779 (21) | 59 (12) | 18 (9) | 185 (7) | 11 (8) |
| Pteropoda | 3 | |||||||||
| Cephalopoda | 2 | 3 | 5 | 12 | 5 | |||||
| Anthropoda | 6 | 5 | 5 | 2 | ||||||
| Anphinora | 3 | 7 | 1 | |||||||
| Scaphopoda | 7 | 21 | 10 | 1 | ||||||
| Dominance of Zones | 0-I | IV-V | V-(VI) | (II)-III | 0-I | 0-I(II) | IV-V | V-VI | (II)-III | VI |
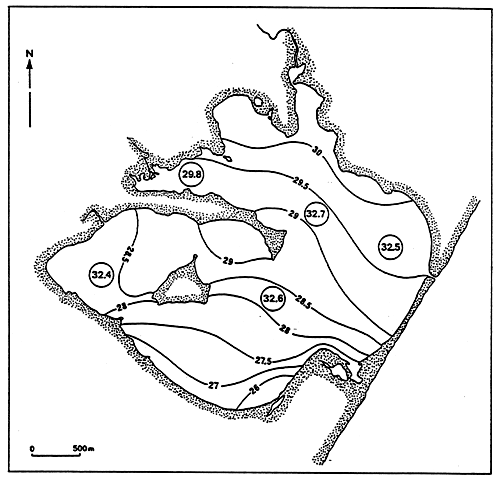
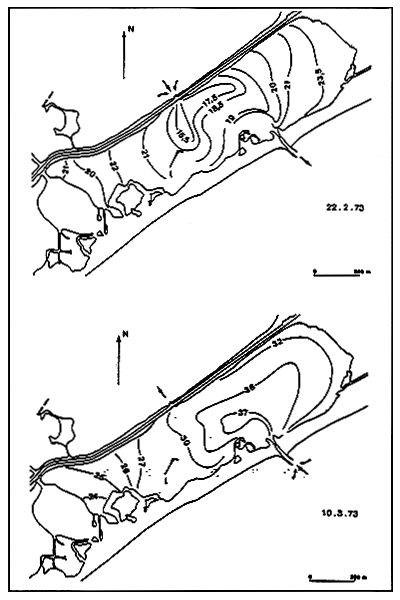
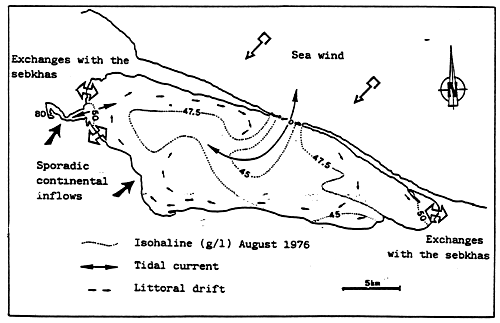
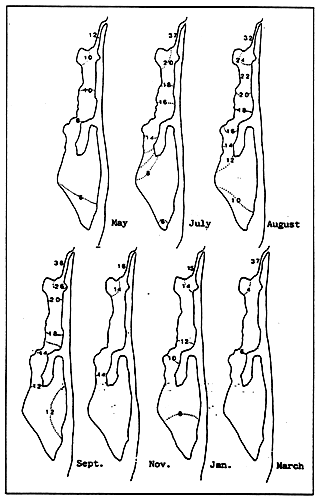
Fig. 4 - Salinity levels (°/00) in the Etang de Biguglia, from May 1978 to March 1979.
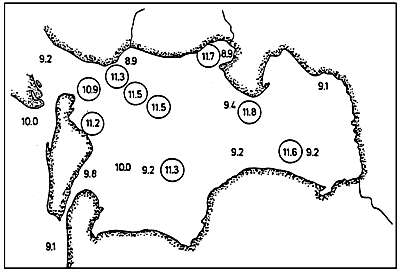
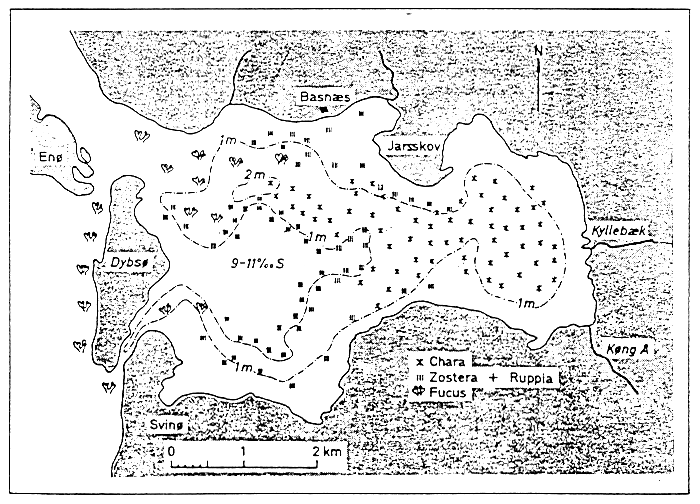
Fig. 6 - Macrophytobenthic associations in the Dybsø Fjord. From MUUS (1967).
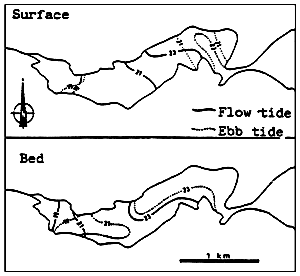
Fig. 7 - Salinity levels (in %0) in the Kysing Fjord. From MUUS (1967).
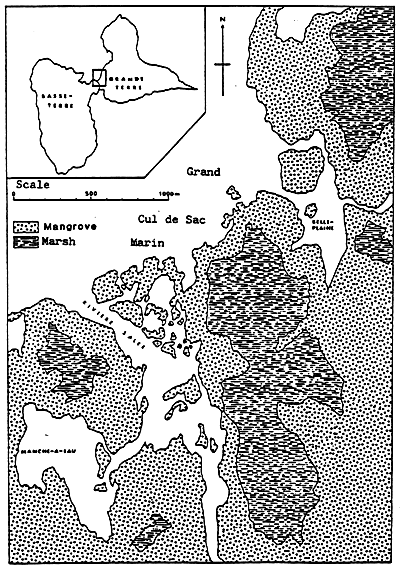
Fig. 8 - Mangrove around the Grand Cul de Sac Marin (Guadeloupe).
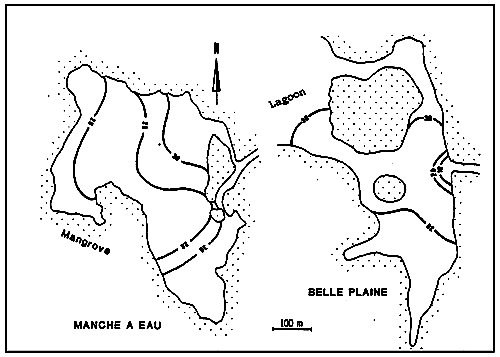
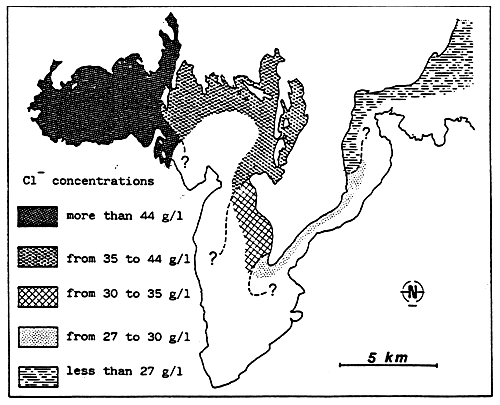
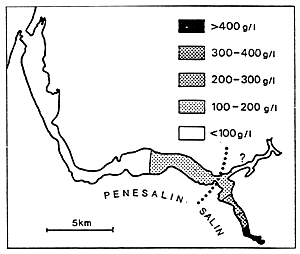
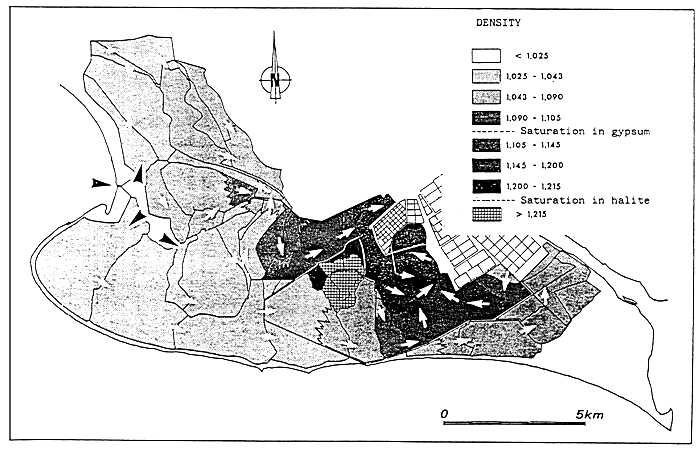
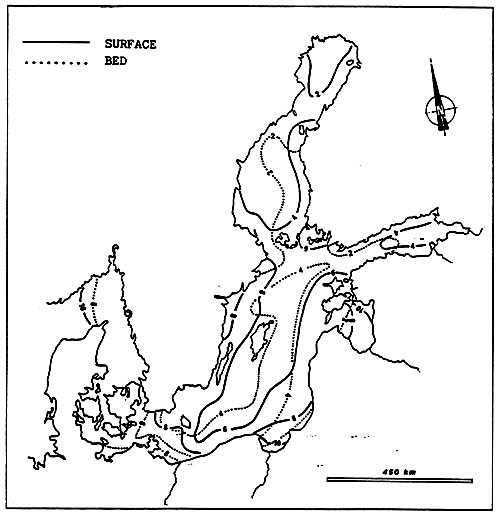
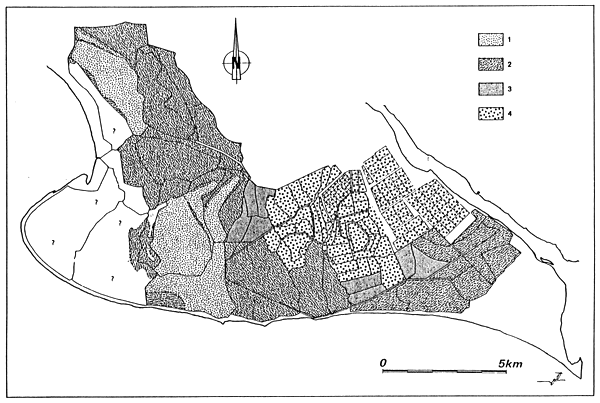
Fig. 14 - Plant associations in the salt marsh of Salin-de-Giraud (Camargue)
1. Communities of Ruppia spiralis and Enteromorpha gr. intestinalis
2. Algar fields of Lyngbya and Microcoleus
3. Algar fields of Microcoleus, Oscillatoria, Phormidium and Spirulina
4. Phytoplanktonic communities of Dunaliella salina and microphytobenthic
communities of Oscillatoria, Phormidium and Spirulina.
From THOMAS (1983) and PERTHUISOT (1983).
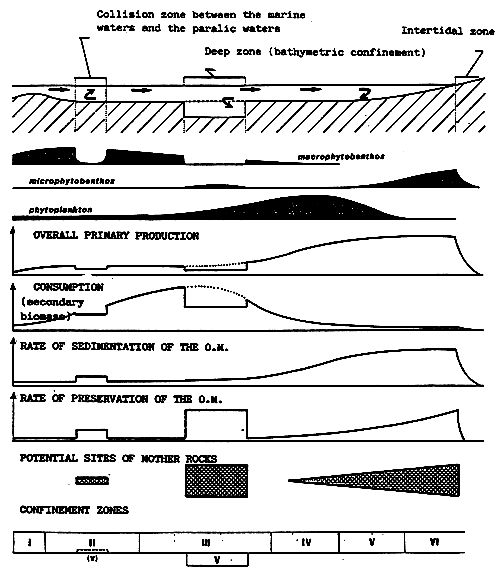
N.B. Local deposits of organic matter (contact between marine waters and lagunar waters, hydraulic umbilicus, zones of intense biosedimentation, inflow of polluting effluents) influence the benthic communities by transforming their qualitative and quantitative features: increase of detritivorous species to the detriment of suspension-feeders, appearance of species indicative of “organic pollution”.
 | 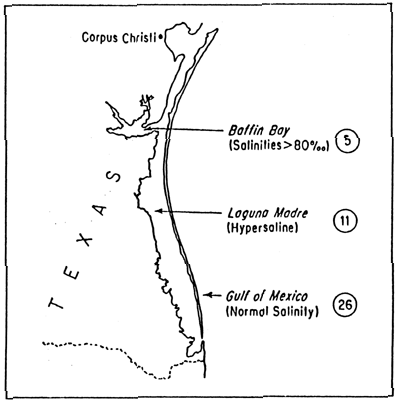 |
| (b) The coast of southern Texas showing, in circles, the difference in relative diversity of the fossilizable faunas at three localities. (Founded on data in Kornicker and Odum, 1958.) | |
| (Reproduction of two nearly consecutive figures, In AGER, D.V., Principles of Paleoecology, McGraw Hill Ed. (1963) | |
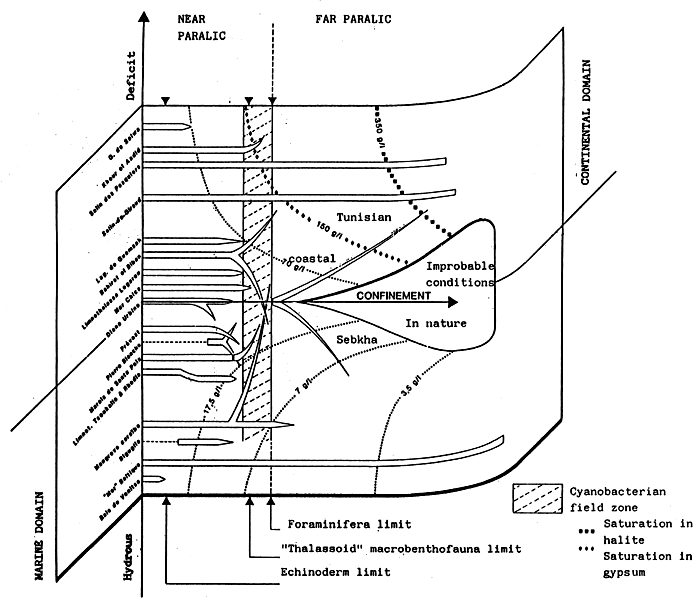
- In the above, the isohaline values of the totality of the totality of the paralic domain are diagrammatically represented as the branches of hyperbolae, Indeed, determining a saline concentration involves determining a Δ h, T couple (hydrous deficit, confinement) so that Δ h, T = constant.
For example, the isohaline corresponding to the normal concentration (36 g/l) is represented on the diagram by the two axes, one for a zero hydrous deficit the other for a zero confinement.
One understand that the higher in absolute value the hydrous deficit is, the less necessary it is that confinement should be great in order to reach a specific concentration, conversely the smaller the hydrous deficit is (in absolute value), the greater the confinement has to be in order to reach any given salinity.
- In the simplest cases (salt marshes) where the hydrous deficit over the whole of the basin can be considered as uniform, the former is represented by a horizontal segment (or a thin rectangle), along which the geochemical an biological zoning of the basin can be read. The figure becomes more complex as the tributaries appear. They are generally situated around the continental or distal ends of paralic basins, when the contact with the sea is only occasional, when the seasonal climatic variations and the local hydrography entail considerable variations of the hydrous situation (e.g., Tunisian coastal sebkhas), or when the water stratification -to which depth contributes-removes a confined body of water from atmospheric influence. This is why it is impossible to maintain a rectangular diagrammatical representation: Therefore, the drawing is covered with branching lines in an attempt to picture the particular conditions of the distal zones.
- The various basins have been arranged on the diagram in the best way possible according to out work and that of other authors.
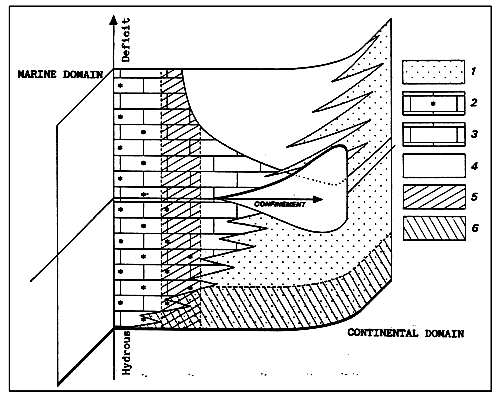
1. Terrigenous detritals.
2. Biodetrital limestone.
3. Biochemical limestone.
4. Evaporties.
5. Autochthonous organic matter (oil ?).
6. Detrital organic matter (coal ?).
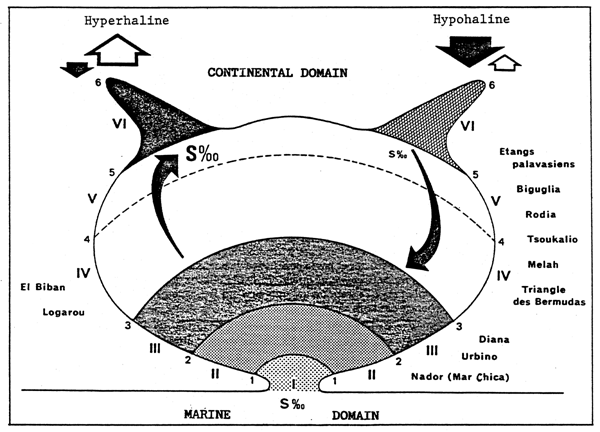
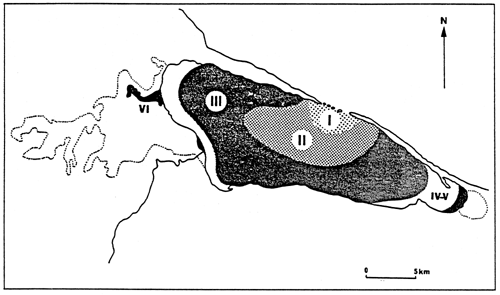
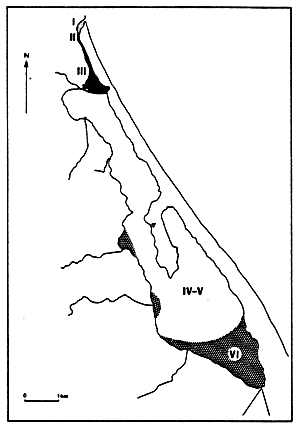
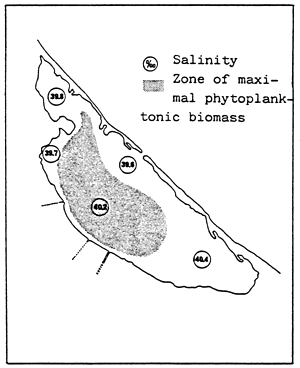
Fig. 20- Diagrammatic map of confinement zones in the Bahiret el Biban. From GUELORGET et al., 1982.
N.B. The shading scale used here is the same as in figures 19 and 21–27
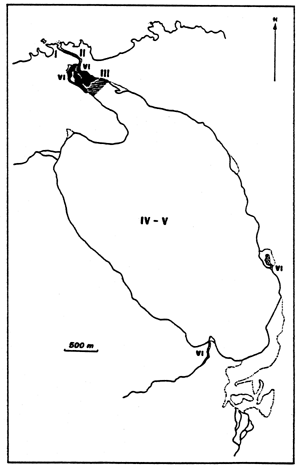
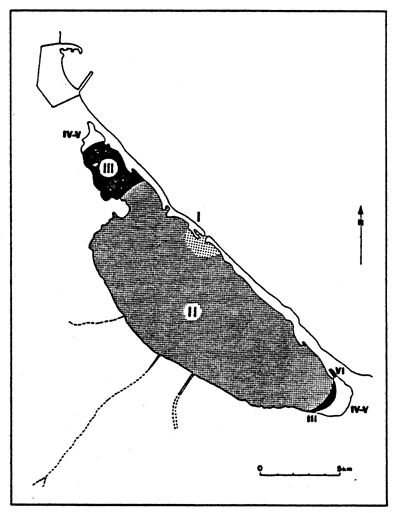 | |
| Fig. 21 - Diagrammatic map of confinement zones in the Nador lagoon (Mar Chica or Sebkha bou Areg). From GUELORGET et al., 1983. |
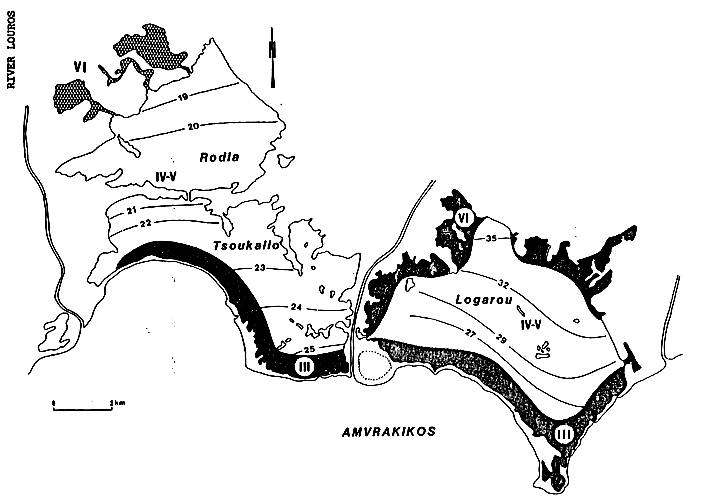
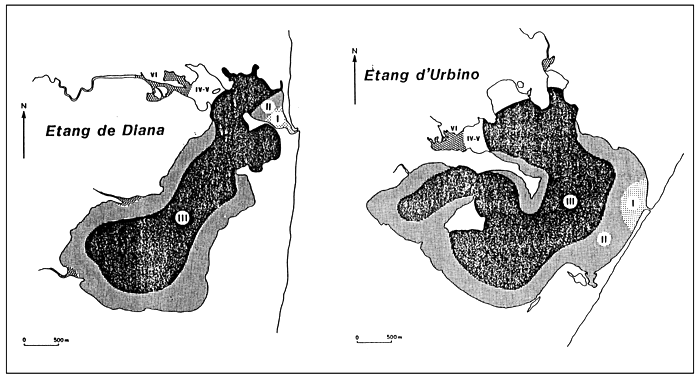
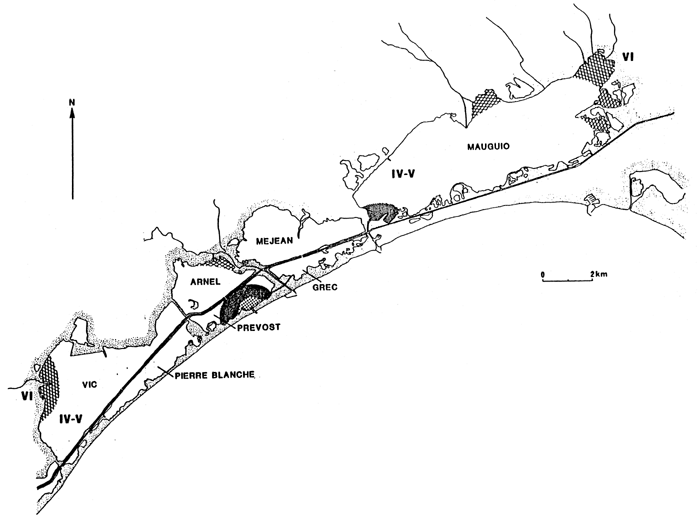
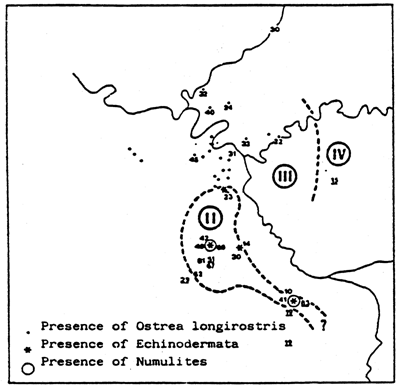
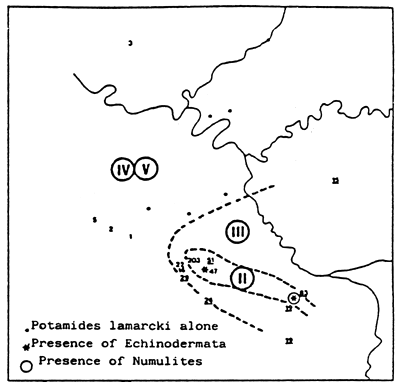
Fig. 26 - Biological zoning in the Palavas lagoons (France) From GUELORGET et al., 1983.
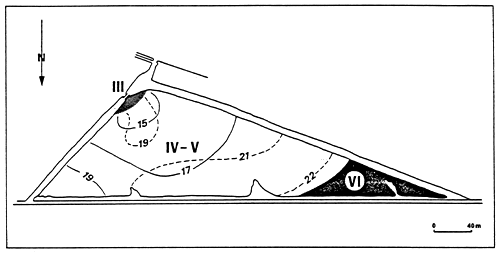
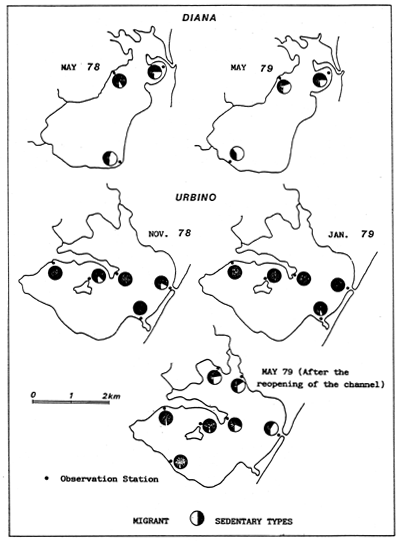
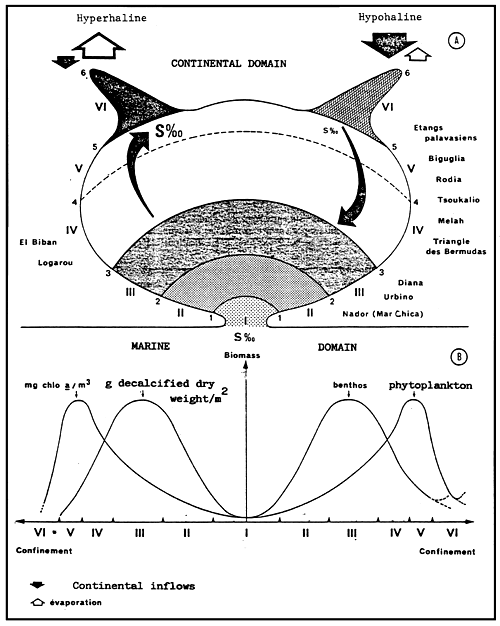 | ||
| Fig. 29 - | A. | A diagrammatic representation of the biological zoning (roman figures) defining the scale of confinement (in Arabic figures) in the model of the Mediterranean paralic ecosystem. |
| B. | Variations showing the phytoplanktonic and benthic biomass in relation tot he confinement scale. The single points of the diagram (maximum curve crossings) remain fixed in relation to the biological zoning. The corresponding values of the biomass can vary according to the overall productivity of each basin. | |
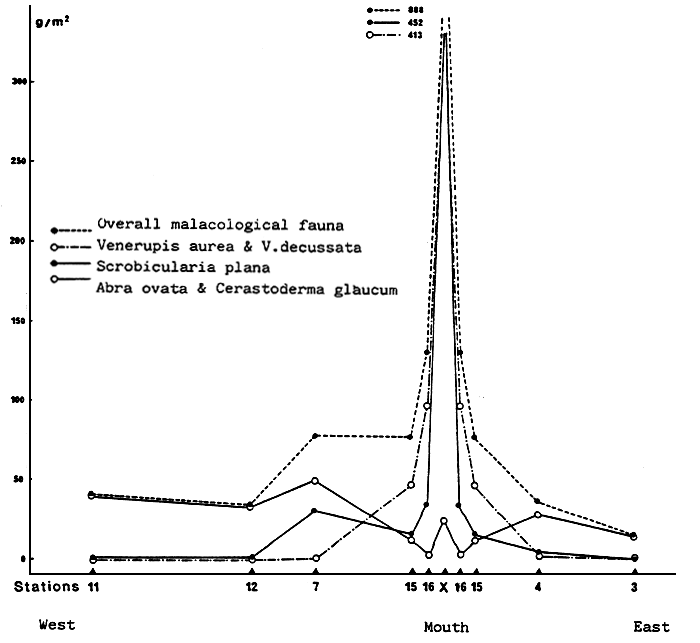
Fig. 30 - Annual malacological production of the Etang du Prévost. From GUELOHGET et al., 1982.
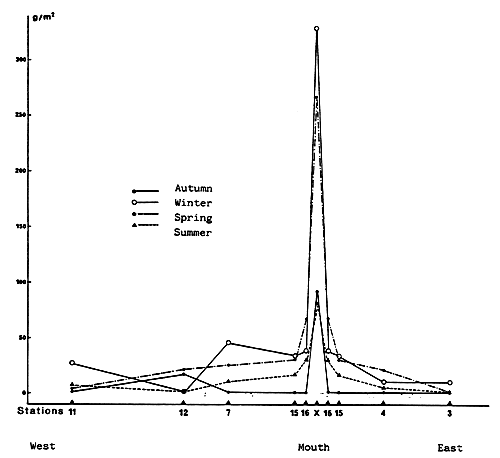
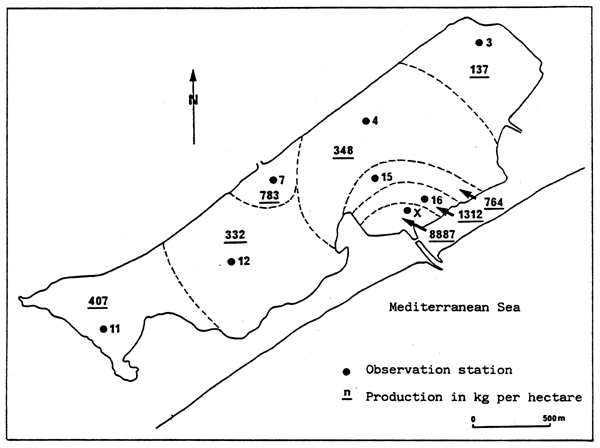
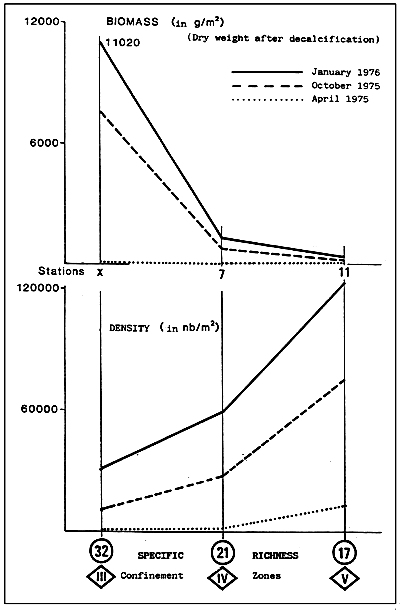
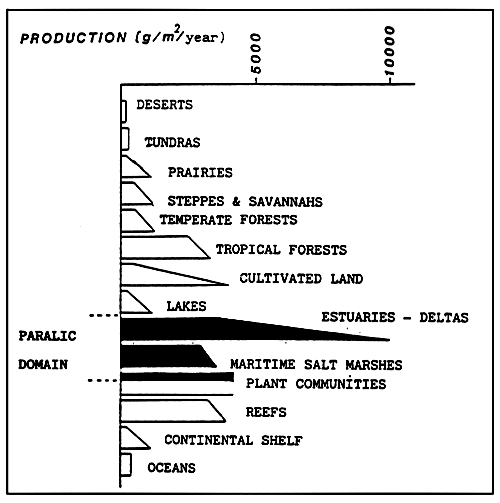
Fig. 34 - Comparative production of some terrestrial and aquatic systems.
(Adapted from BASSON et al., 1977, and ALLEN et al., 1979)
Dotted line: Presence of higher magnesian calcite (note that this is found in zones which are not greatly confined in relation with the sea and in the marginal backwaters where Pirenella conica are found in profusion on a cyanobacterian substrate). The aragonite maximum seems to be in relation with the contact between the confined waters which have travelled along the northern fringe of the basin, and “fresh” waters coming from the Bay of Amvrakikos. From FRISONI et al., 1982.
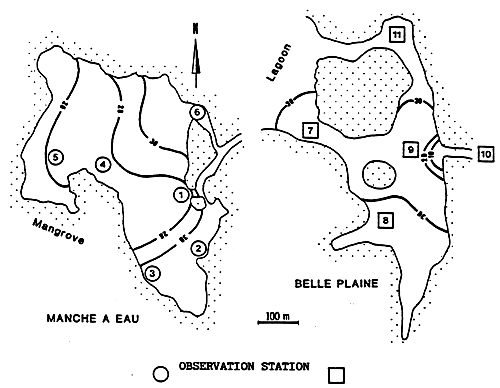
Fig.38 - Situation of observation stations in the lagoons of Bells Plaine and Manche à Eau.
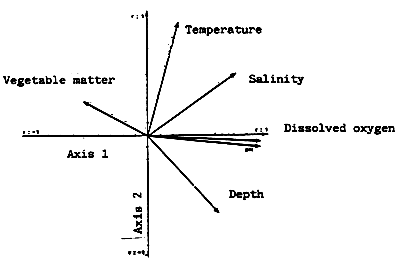
Fig. 39 - Abiotic parameters. Correlations between variables and principal axes.
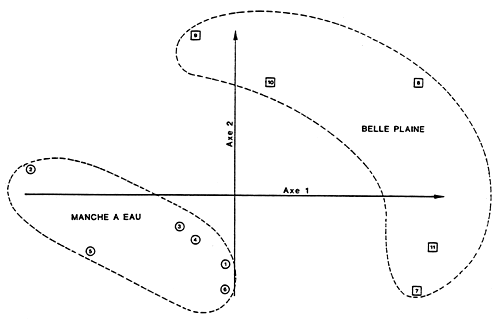
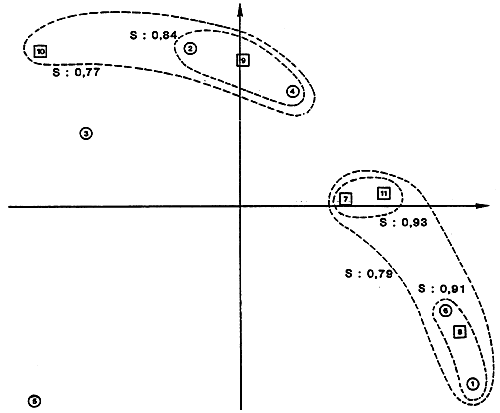
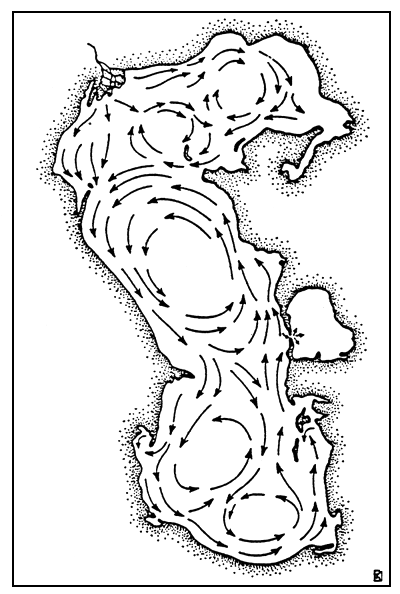 | |
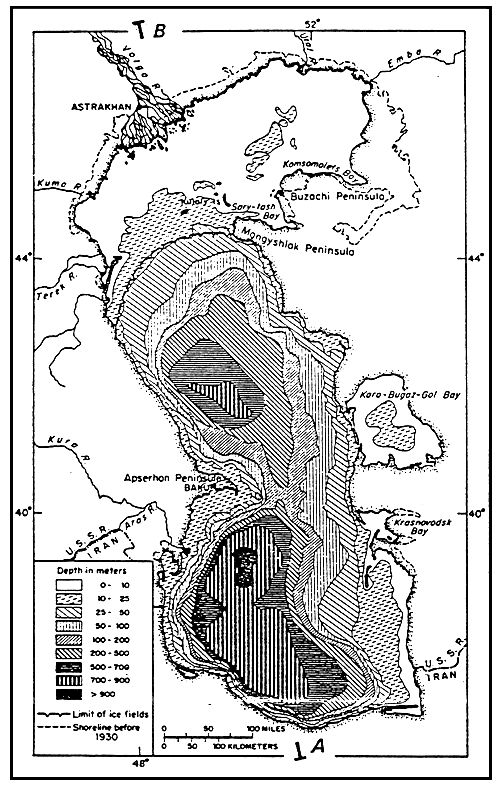
| Fig. 44 - Salinity Levels in the Caspian Sea. From DZENS-LITOVSKIJ, 1962, (modified). | 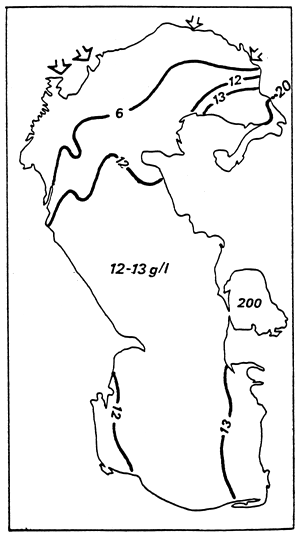 |
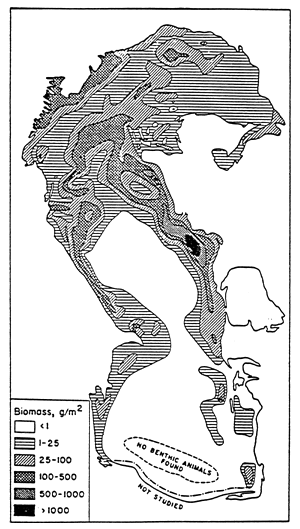
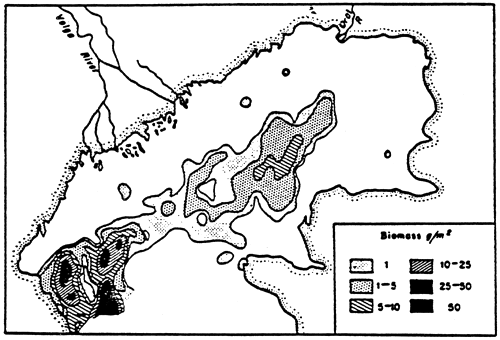
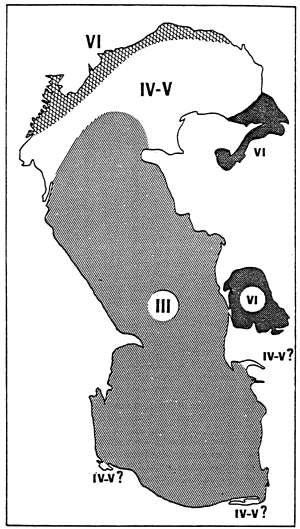
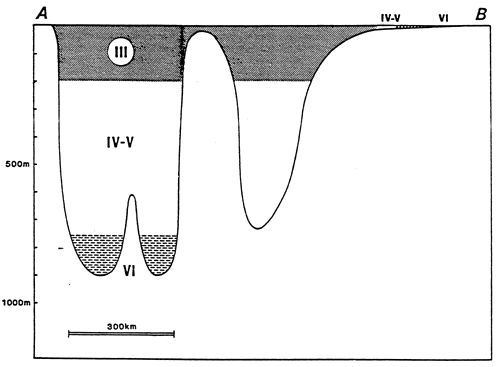
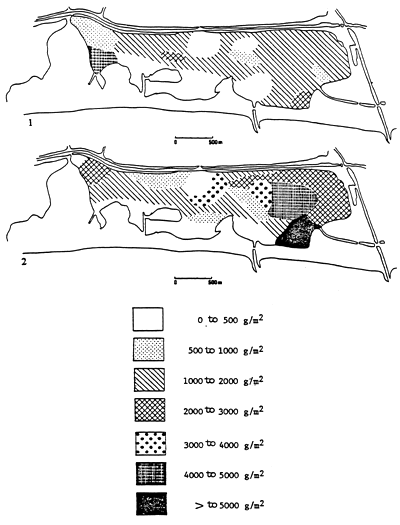
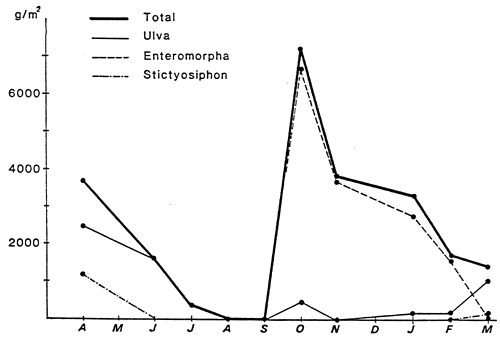
Fig. 50 - Monthly variations of total specific (Ulva, Enteromorpha, Stictyosiphon) algal biomass in the central zone of the Etang du prévost (drained wet weight in g/m2). From RIOUALL, 1976.
Note the increase in the intertidal zone on surfaces exposed to the waves, and the distance of the supratidal zone from the mean tide level. On the contrary, on sheltered surfaces, zonation is much tighter. On exposed surfaces the confinement gradient is reduced by wave lapping, and spray.
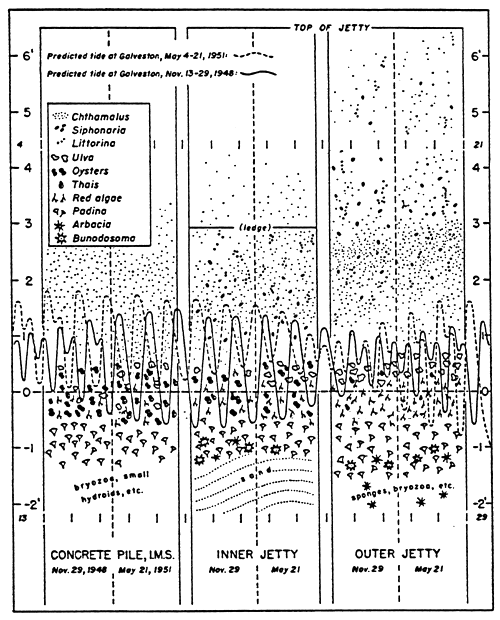
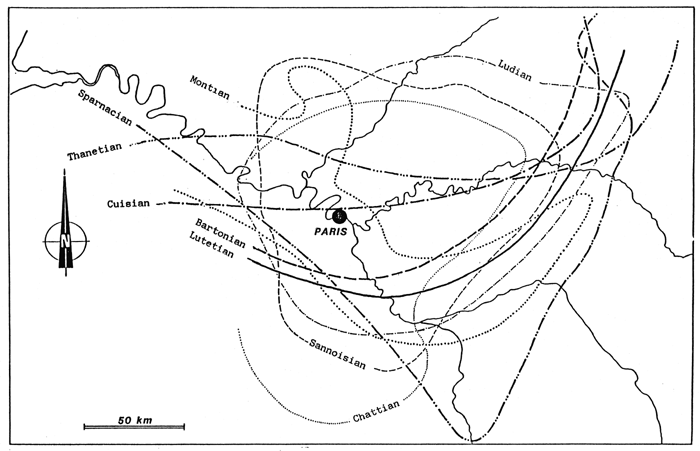
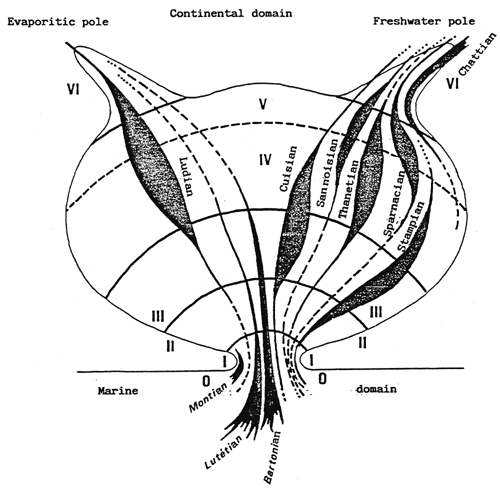
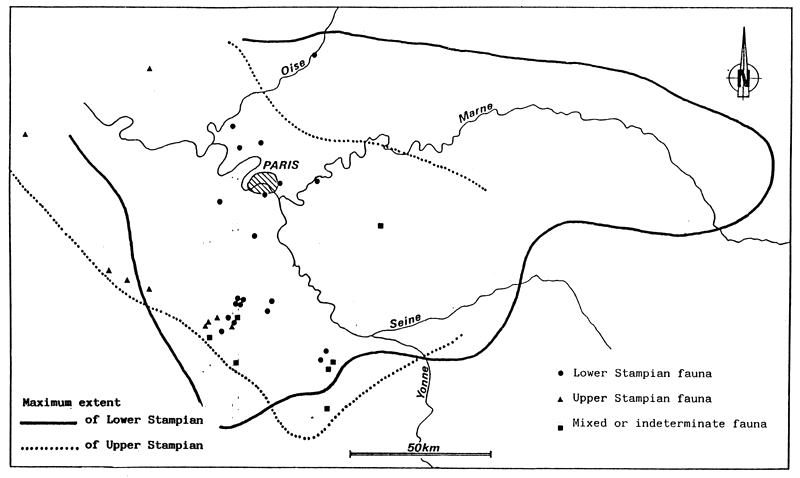
Fig. 54 - Situation of principal Stampian fossil deposits in the Paris Basin. From ALIMEN (1936).
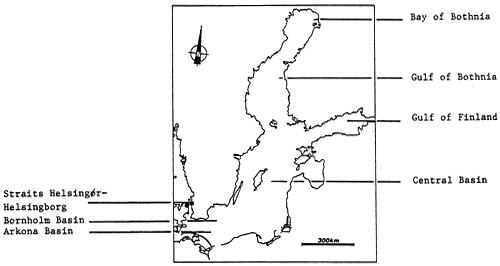
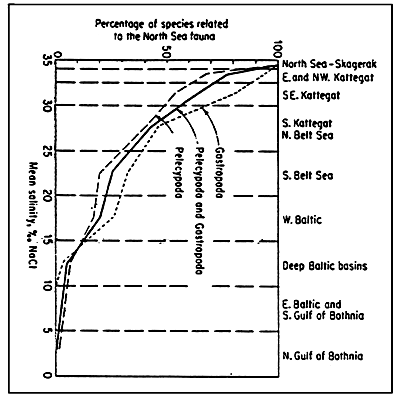
Fig. 59 - The Baltic Sea and its principal basins.
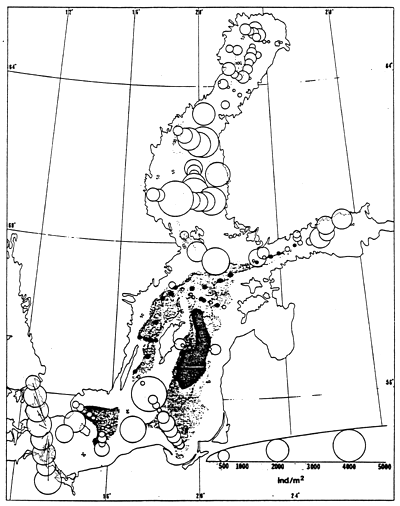
Black dots indicate stations devoid of macrofauna; grey areas indicate regions where the dissolved oxygen level is below 2 ppm; black areas indicate regions where H2S is present.
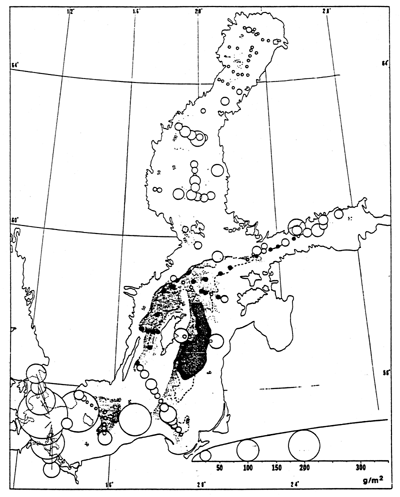
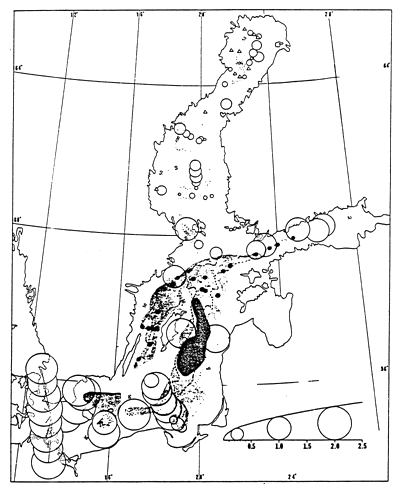
Same symbols as for fig. 60. Triangles represent stations where only one species was found.
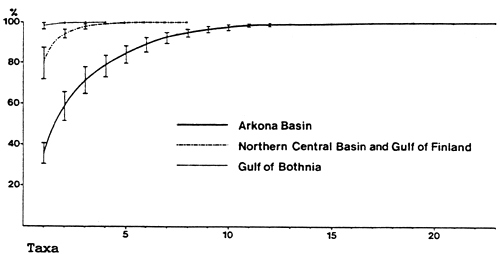
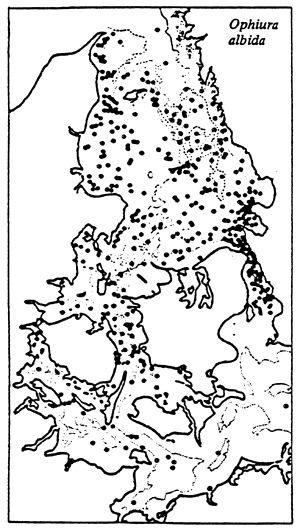 | 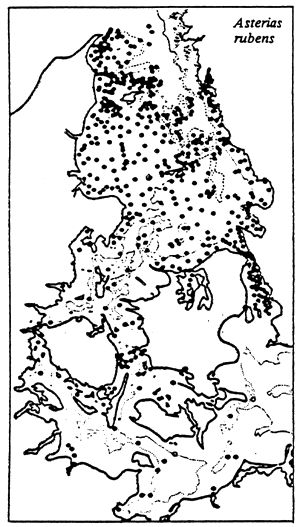 |
| Note the “Channel effect” opposite the Øresund and the other Danish straits where the water flow ensures a sufficient stream of “vital elements” to allow good development of Echinodermata. This “channel effect” was suggested by A. JAUZEIN; it corroborates the authors' hypothesis as the ways in which confinement acts. | |
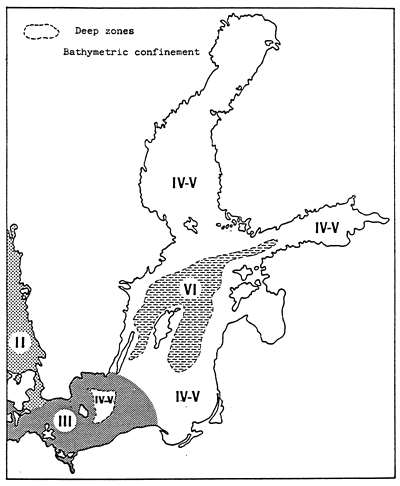
Fig. 65 - Map of confinement zones in the Baltic Sea.
 |  |
 | |
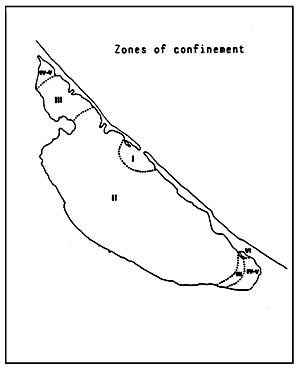 | 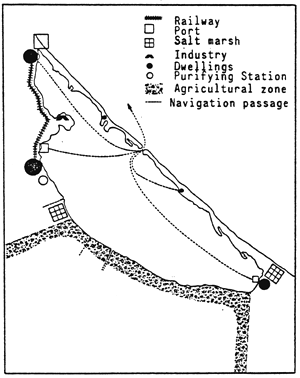 |
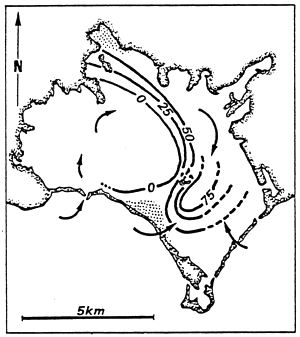
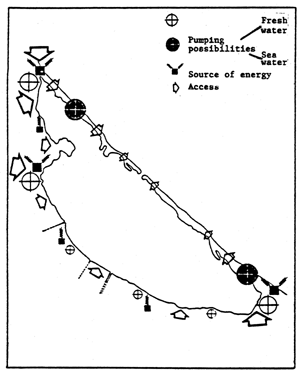
| Fig. 69 - Confinement in the Nador lagoon. | |
 | |
![]()
![]()
![]()