![]()
![]()
![]()
Biologie de l'Anchois (Engraulis encrasicolus, L.) en Mauritanie
par
I. BA1
1. INTRODUCTION
L'Anchois, Engraulis encrasicolus, est un poisson Téléostéen clupéiforme de la famille des Engraulidés d'une quinzaine de centimètres, caractérisé surtout par l'allongement du museau en rostre au-dessus d'une bouche infère largement fendue. Il est répandu dans tout l'Atlantique oriental, depuis les côtes de la Norvège au nord de Bergen (62°N) jusqu'en Afrique du Sud (23°S). Cette espèce se rencontre aussi dans la Mer Baltique, la Manche, la Mer du Nord.Elle est également répandue dans tout le Bassin méditerranéen y compris la Mer Noire et la Mer d'Azov.
Espèce pélagique formant souvent de grands bancs, Engraulis encrasicolus vit dans des eaux peu profondes et parfois jusqu'à 400 mètres; elle fréquente aussi les estuaires.
Sur les côtes mauritaniennes, l'Anchois ne fait l'objet d'aucune pêche commerçiale. Il y est pourtant rencontré en abondance notamment dans des eaux de températures comprises entre 18 et 20°C (Maigret, 1972). Ainsi, les campagnes de prospection acoustique effectuées dans la région ont en évidence un potentiel exploitable de l'ordre de 20.000 tonnes, localisé au sud immédiat du cap Blanc, le long de la façade ouest du Banc d'Arguin (Anonyme, 1985). Cette espèce n'ayant fait l'objet d'aucune étude particulière dans cette partie de l'Atlantique tropical, il nous a paru intéressant de présenter les principaux traits de sa biologie (reproduction et croissance) en Mauritanie.
2. MATERIELS ET METHODES D'ETUDES
Les données utilisées dans cette étude proviennent des échantillonnages effectués de février 1986 à novembre 1987 au niveau de la flottille pélagique soviétique et au cours des campagnes de prospection acoustique du navire du Centre National de Recherches Océanographiques et des Pêches (CNROP) le N/O N'Diago. Au cours de ces é chantillonnages, les poissons étaient mesurés au 1/2 cm inférieur de longueur à la fourche (LF); et sur chaque individu, deux poids ont été relevés:
le poids total du corps au gramme près;
le poids des gonades au dixième de gramme près.
Le sexe et le stade sexuel ont également été notés. D'autre part, pour la détermination de l'âge, trois à cinq paires d'otolithes par classe de taille ont été prélevées.
3. REPRODUCTION
3.1. METHODES D'ETUDE
3.1.1. Taille à la première mâturité sexuelle
Plusieurs définitions sont données de la taille à la première mâturité sexuelle. Celles le plus souvent admises sont:
la taille du plus petit poisson mûr ou du plus grand individu immature pendant la saison de reproduction;
la longueur à partir de laquelle 50 % des individus d'une population sont sexuellement mûrs lors de la période de ponte. Pour pouvoir comparer avec les observations effectuées dans d'autres études, ces deux définitions ont été retenues.
1 Océanographe biologiste du CNROP, B.P.22, Nouadhibou. Rép. Islam.de Mauritanie
La détermination de la L50 (longueur à partir de laquelle 50 % des poissons sont matures) a été faite en regroupant les individus par sexe et par classe de taille de 1/2 cm LF. Ensuite, le pourcentage des individus matures de chaque classe de taille pendant la principale période de reproduction a été calculé en fixant le seuil de mâturité à partir du stade III qui correspond àla phase de développement des ovaires (Fontana, 1969; Conand, 1977).
3.1.2. Période(s) de reproduction
Pour déterminer avec précision la ou les période(s) de ponte d'une espèce de poisson donnée, il est nécessaire d'employer plusieurs méthodes. Dans le cadre de cette étude, deux types de données ont été utilisées:
les variations mensuelles des stades sexuels;
les évolutions des moyennes mensuelles du rapport gonado-somatique (R.G.S.).
Pour la détermination des stades sexuels, nous avons employé l'échelle de mâturité de FONTANA (1969) basée sur l'étude des sardinelles du Congo. Cette échelle comprend sept stades:
| stades I et II | : période d'inactivité sexuelle |
| stade III | : en cours de maturation |
| stade IV | : pré-ponte |
| stade V | : ponte |
| stade VI | : post-ponte |
| stade VII | : fin de la dernière ponte et involution de l'ovaire. |
Cependant, dans cette étude, nous avons assimilé le stade VII au stade VI car ils sont difficilement différentiables chez l'Anchois.
Le rapport gonado-somatique a été calculé comme suit:
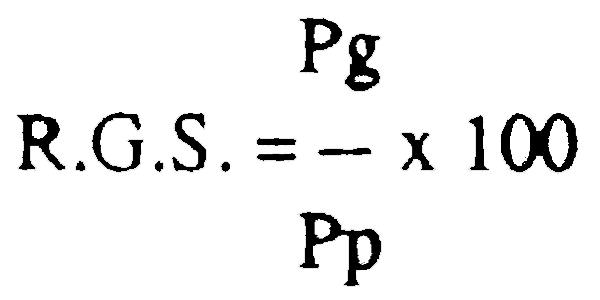
où R.G.S. = rapport gonado-somatique;
Pg = poids des gonades;
Pp = poids total du corps.
4.CROISSANCE
En milieu tropical, la croissance est délicate à étudier car les méthodes usuelles se heurtent à des obstacles majeurs. En effet, si la ponte n'est pas périodiquement interrompue ou du moins fortement ralentie, il est très difficile de décomposer en modes distincts les distributions des fréquences de longueur pour l'étude de la croissance par la méthode des filiations modales. Pour déterminer l'âge de l'Anchois, nous avons donc eu recours aux otolithes, utilisant pour cela la technique des marques supposées journalières préconisée par Panella (1971, 1973).
4.1. PREPARATION ET INTERPRETATION DES OTOLITHES
Après extraction, les otolithes sont lavés puis placés sur une lame et inclus dans une résine synthétique puis chauffés. Une fois l'inclusion sèche, l'otolithe est abrasé sur ses deux faces à l'aide de la poudre de carborandum pour bien dégager les stries, puis attaqué à l'acide chlorydrique (2%) pendant 10–20 secondes afin d'augmenter le contraste. Après rinçage abondant à l'eau, l'otolithe est “lu” au microscope optique au grossissement 400. Seules sont comptées les marques qui apparaissent comme l'association d'une bande claire et d'une bande sombre en assimilant le nombre de marques ainsi obtenu à l'âge exprimé en jours.
4.2. EQUATIONS DE CROISSANCE
4.2.1. Croissance linéaire
Le modèle retenu pour décrire la croissance de l'Anchois de Mauritanie est celui de von Bertalanffy (1935). Son expression est:
Lt = L∞ (1-exp (-K (t-to)))
avec Lt = longueur du poisson à l'instant t ;
Loo = longueur asymptotique qui serait atteinte par le poisson à l'âge théorique infini,
K = coefficient de croissance caractérisant la vitesse avec laquelle l'espèce croît vers sa taille asymptotique,
to = âge théorique pour lequel la taille est nulle.
Les trois paramètres (Loo, K, to) de l'équation de von Bertalanffy ont été estimés par la méthode d'Abrasson-Tomlinson (1961) qui repose sur le principe général d'optimisation des estimations par le critère des moindres carrés des écarts entre la courbe estimée et les données de base.
4.2.2. Croissance pondérale
Les lois et courbes de croissance de von Bertalanffy permettent également de décrire la croissance pondérale des poissons dont il est nécessaire de connaître la loi pour estimer les biomasses et les productions. Pour cela, il est nécessaire déterminer la relation qui lie la longueur du poisson à son poids ou relation taille-poids.
Relation taille-poids:
Chez la plupart des poissons, la longueur et le poids peuvent être reliés par une équation du type:
W= a.Lb
où W = poids du poisson,
L = longueur à la fourche,
a = constante,
b = coefficient d'allométrie.
Après transformation logarithmique, cette relation devient:
Log W = Log a + b Log L
La régression utilisée pour déterminer a et b est l'axe majeur réduit connu également sous le nom de droite de Tessier.
Equation de la croissance pondérale:
L'équation de la croissance en poids s'obtient par combinaison de l'équation de la croissance linéaire et de la relation taille-poids. Elle s'écrit:
Wt = Woo (1-exp (-K (t-to)))b
avec W = poids au temps t,
Woo = poids asymptotique correspondant à la longueur asymptotique Loo,
a et b = paramètres de la relation taille-poids,
K et to = paramètres de la croissance linéaire.
5. RESULTATS
5.1. REPRODUCTION
5.1.1. Taille à la première mâturité sexuelle.
Pendant toute la durée de la période de reproduction, les plus petits individus matures capturés mesuraient 4,5 cm chez les femelles et 4 cm chez les mâles, Les plus grands individus immatures rencontrés chez les deux sexes avaient une taille de 10 cm. En revanche, les tailles correspondant au point L50 (longueur à laquelle 50% des individus de la population considérée sont matures) sont de 10,1 cm pour les femelles et de 10,4 cm pour les mâles (Figure 1). Les deux tailles étant très proches l'une de l'autre, il est possible de considérer que 50% des Anchois sont sexuellement mûrs à partir de 10 cm (LF).
Les observations provenant d'études réalisées dans d'autres régions sont indiquées dans le tableau 1. Bien que les résultats de la présente étude (L50) soient proches de ceux obtenus en Oranie (Algérie), la comparaison des différentes observations est difficile. En effet, de nombreux auteurs n'indiquent ni la méthode de détermination retenue, ni le critère de mâturité adopté, ni la taille de référence utilisée. Il apparaît cependant, comme caractère constant, que dans toutes les régions où des observations ont été faites, Engraulis encrasicolus atteint sa première mâturité sexuelle dès sa première année d'existence.
5.1.2. Variations mensuelles des stades sexuels
Les pourcentages mensueles des différents stades de mâturité sexuelle des mâles et des femelles sont présentés sur la figure2. Des individus à tous les stades sont présents toute l'année avec cependant des fréquences variables. Les poissons au stade II (repos) et en cours de maturation (stade III) sont surtout abondants en février, mars, septembre, novembre et décembre. Contrairement aux autres mois, aucun Anchois en reproduction n'a été rencontré en septembre.
La plus forte proportion de poissons en fin d'ovogénèse ou aptes à se reproduire (stades IV, V et VI) est observée d'avril à juillet en 1986 et d'avril à octobre 1987. Cependant, le pourcentage élevé d'individus mûrs (stades III, IV, V) observé en novembre(36%) et décembre(98%) 1986 indique une seconde période possible de forte ponte.
5.1.3. Evolution du rapport gonado-somatique
Les moyennes mensuelles des R.G.S. sont comprises entre 1,65 et 5,12% (Figure 3, Tableau2). Les plus faibles valeurs sont observé es en février, mars et septembre 1986; les plus fortes en mai 1986 (5,12%), avril (3,51%), juin (4,66%) et octobre (4,19%)1987.
Le décalage d'une année à l'autre des pics du rapport gonado-somatique sembleêtre dû à:
la baisse de la température de surface en 1987 qui passe de 19,23°C en avril à 17,89°C en mai,
la présence en octobre 1987 de nombreuses femelles proches ou en cours de ponte (90% des femelles était aux stades IV et V).
5.1.4. Période de reproduction
Le suivi du rapport gonado-somatique associé aux variations mensuelles des différents stades de mâturité sexuelle ont permis de situer la principale période de ponte de l'Anchois de'avril à octobre. Cependant, la reproduction ne cesse pas totalement puisque des individus mûrs s'observent toute l'année. En effet, chez cette espèce, tous les individus de grande taille (LF > 10 cm) se reproduisent quelle que soit la saison (Tableau 4). Une reproduction étalée sur toute l'année a aussi été observée au Sénégal par Conand (1970) avec un maximum d'oeufs en janvier et novembre et par Marchal (1966) en Côte d'Ivoire avec deux pics: le premier de janvier à mars et le second de septembre à octobre.
5.1.5. Facteurs hydrologiques et reproduction
En zone tropicale, la reproduction est souvent considérée comme continue du fait de la faible amplitude des variations de l'environnement au cours d'un cycle annuel. Cependant, l'existence le long des côtes mauritaniennes de phénomènes d'upwelling fait apparaître des variations saisonnières bien marquées. L'on sait également que le processus de maturation et la ponte sont soumis à l'influence de nombreux facteurs extérieurs qui peuvent accélérer, déclencher ou retarder l'ensemble des phénomènes liés à la reproduction. Parmi ces facteurs, la température et la salinité sont les paramètres généralement observés en raison de leur facilité de mesure. Toutefois, en ce qui concerne l'Anchois, E. encrasicolus, il a été établi que le rôle de la salinité était très négligeable (Furnestin et Furnestin, 1959; Sobral, 1975 et Rê et al., 1983). Selon ces auteurs, il est possible de trouver des rapports entre la salinité et la reproduction de cette espèce uniquement dans la mesure où l'évolution de ce paramètre est liée à une élévation de la température. Nous nous limiterons donc à examiner les relations existant entre la température et la reproduction de l'Anchois.
| Régions | Taille (cm) | Age (an) | Méthodes | Auteurs |
|---|---|---|---|---|
| Méditerranée | 12–13 | 1 an | - | Fage, 1911 |
| Golfe de Gascogne | 11–12 | 1 an | - | Furnestin, 1945 |
| Méditerranée Occidentale (Costa Brava) | 11.8 | 1 an | - | Bas et Morales, 1954 |
| Oranie (Algérie) | 11.0 | 1 an | - | Arrignon, 1966 |
| Côtes Basques Espagnoles | 8.2 | lère année | plus petit individu mature | Navaz et Lozano Cabo, 1966 |
| Adriatique centrale | ♂8.6 ♀ 9.7 | lére année | plus petit individu mature | Sinovcic, 1978 |
| ♀ 4.5 ♂ 4.0 | 3 mois | plus petit individu mature | ||
| Mauritanie | Présente étude | |||
| 10.0 | 8 mois | Lo o |
| 1986 | 1987 | ||||
|---|---|---|---|---|---|
| Mois | N | R.G.S. | N | R.G.S. | |
| F | 130 | 1.73 ± 0.91 | 69 | 1.65 ± 1.28 | |
| M | 43 | 2.12 ± 0.55 | 227 | 2.01 ± 1.63 | |
| A | 143 | 3.98 ± 1.33 | 217 | 3.51 ± 2.77 | |
| M | 112 | 5.12 ± 1.46 | 153 | 3.15 ± 2.51 | |
| J | 197 | 4.59 ± 2.16 | 101 | 4.66 ± 1.36 | |
| J | 29 | 3.21 ± 0.69 | 197 | 2.94 ± 1.37 | |
| A | - | - | 80 | 3.14 ± 1.29 | |
| S | 134 | 2.58 ± 1.12 | - | - | |
| O | - | - | 79 | 4.19 ± 1.54 | |
| N | 88 | 3.02 ± 1.96 | 80 | 2.66 ± 1.01 | |
| D | 104 | 3.32 ± 2.48 | - | - | |
| Stades | J | F | M | A | M | J | J | A | S | O | N | D | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ≤ 10 CM L.F. | II | 113 | 101 | 110 | 35 | 15 | 3 | 1 | 43 | - | 63 | 27 | |
| III | 42 | 65 | 56 | 33 | 24 | 5 | - | 91 | 2 | 44 | 15 | ||
| IV | 7 | 5 | 24 | 23 | 21 | 3 | 1 | - | 9 | 32 | 35 | ||
| V | 1 | 3 | 8 | 14 | 28 | 6 | - | - | 1 | 10 | 10 | ||
| VI | - | - | - | 7 | - | 3 | - | - | 4 | 5 | |||
| > 10 CM L.F. | II | - | - | - | - | - | - | - | - | - | - | ||
| III | 5 | 20 | 21 | 20 | 13 | 50 | 3 | - | 1 | 1 | |||
| IV | 19 | 40 | 92 | 46 | 45 | 57 | 30 | - | 24 | 5 | 6 | ||
| V | 11 | 21 | 33 | 58 | 72 | 63 | 31 | - | 39 | 6 | 3 | ||
| VI | 4 | 15 | 16 | 29 | 80 | 36 | 14 | - | 4 | 6 | 5 |
| Nb.marq. | Age | L.observ. | L.calc. |
|---|---|---|---|
| journal. | (an) | (cm) | (cm) |
| 97 | 0.27 | 4.0 | 4.2 |
| 103 | 0.28 | 4.5 | 4.5 |
| 108 | 0.30 | 5.0 | 4.7 |
| 122 | 0.33 | 5.5 | 5.4 |
| 137 | 0.37 | 6.0 | 6.1 |
| 151 | 0.41 | 6.5 | 6.8 |
| 161 | 0.44 | 7.0 | 7.2 |
| 170 | 0.46 | 7.5 | 7.6 |
| 177 | 0.48 | 8.0 | 7.8 |
| 186 | 0.51 | 8.5 | 8.2 |
| 203 | 0.55 | 9.0 | 8.9 |
| 212 | 0.58 | 9.5 | 9.2 |
| 234 | 0.64 | 10.0 | 10.0 |
| 259 | 0.71 | 10.5 | 10.8 |
| 266 | 0.73 | 11.0 | 11.1 |
| 281 | 0.77 | 11.5 | 11.5 |
| 298 | 0.82 | 12.0 | 12.0 |
| 317 | 0.87 | 12.5 | 12.56 |
| 335 | 0.92 | 13.0 | 13.0 |
| 347 | 0.95 | 13.5 | 13.3 |
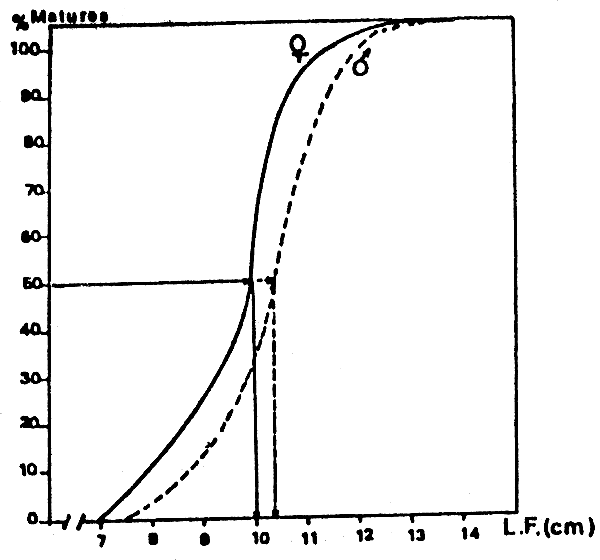
Figure 1 Taille à la première sexuelle d'Engraulis encrasicolus
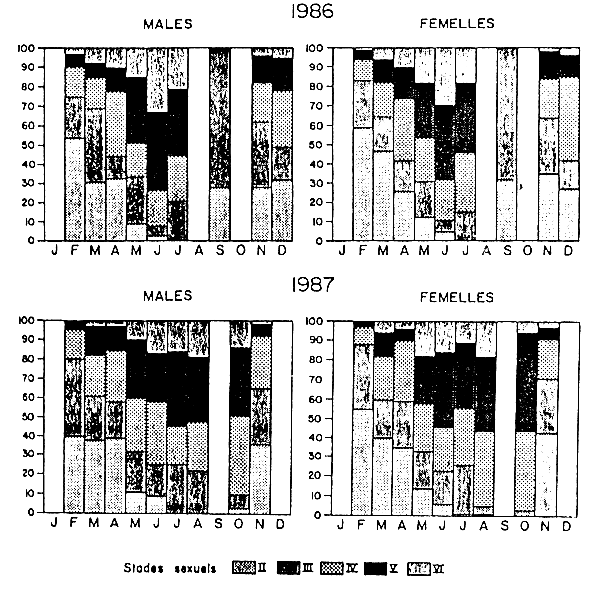
Figure 2 Evolution mensuelle des stades de maturité sexuelle d'Engraulis encrasicolus
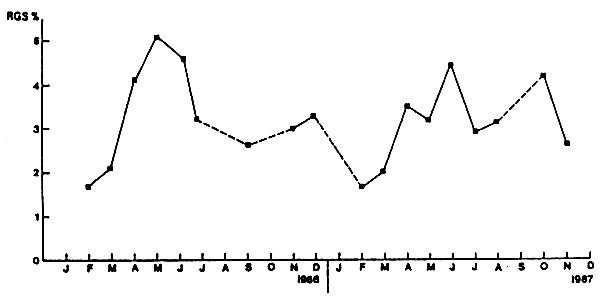
Figure 3 Evolution mensuelle du rapport gonado-somatique moyen des femelles d'Engraulis encrasicolus
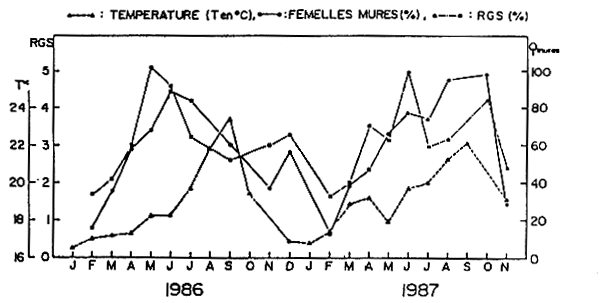
Figure 4 Evolution des R.G.S. moyens et du pourcentage des femelles d'Engraulis encrasicolus mures en fonction de la température de surface.
La figure 4 montre que l'augmentation du pourcentage des femelles mûres et du R.G.S. est parallèle à l'accroissement de la température de surface. Cependant, l'intensité maximale de ponte ne correspond pas au maximum thermique, mais a lieu lors du réchauffement progressif des eaux (transition saison froide-saison chaude) et quelques fois pendant la transition saison chaude-saison froide (novembre-décembre).
La prédominance des saisons de transition sur la reproduction a été également notée par Domanevsky (1968) et par Domain (1979) chez la plupart des espèces pélagiques et démersales de la région sénégalo-mauritanienne.
5.1.6. Conclusion
En Mauritanie, la reproduction de l'Anchois a lieu toute l'année, mais du fait de l'existence de saisons hydrologiques assez marquées, certaines périodes sont plus favorables que d'autres. Ainsi, chez cette espèce, la transition saison froide-saison chaude est la plus favorable et induirait le déclenchement de la ponte. La température semble être le facteur essentiel dans le déclenchement de la ponte soit par stimulation des mécanismes physiologiques, soit par enrichissement trophique du milieu. En effet, des études effectuées en Méditerranée et dans l'Océan Atlantique (Furnestin et Furnestin, 1959; Demir, 1965; Aldebert et Tournier, 1971, 1977; Arbault et Lacroix, 1972; Sobral, 1975; Chavance, 1980; Rê et al., 1983) montrent que pour l'Anchois E. encrasicolus, l'optimum thermique pour la reproduction se situe entre 14 et 24°C. En Mauritanie, pendant toute la période d'étude, les températures étaient comprises entre 16,5 et 24° C. C'est vraisemblablement ces facteurs thermiques favorables qui sont à l'origine de la ponte étalée sur toute l'année, la nourriture ne constituant probablement pas, dans cette région à très forte productivité, un facteur limitant.
5.2. CROISSANCE
5.2.1. Croissance linéaire
Le dénombrement des marques journalières sur les otolithes a permis d'obtenir pour chaque classe de taille un couple de données “nombre de marques-longueur” qui, en supposant qu'une marque se fasse chaque jour, correspond à un couple âge-longueur. Les paramètres de l'équation de von Bertalanffy ainsi obtenues sont:
Loo = 21,80 cm,
K = 1,056(annuel),
to = 0,0675 (annuel).
Ce qui permet d'écrire l'équation de la croissance linéaire en fonction du temps:
Lt = 21,8 (1 -exp(- 1,056 (t - 0,0675)))
Limites de validité : 4 à 13.5 cm L.F.
Les couples âge-longueur sont donnés dans le tableau 4 et la courbe de croissance linéaire correspondante dans la figure 5.
5.2.2. Croissance pondérale
La relation taille-poids:
La relation taille-poids obtenue à partir d' un effectif de 90 individus est la suivante:
W = 1,47. 10-3 LF.3,75 (r = 0,96).
Limites de validité ente 6,5 et 13 cm LF.
Cette relation est du type allométrie majorante, le coefficient b étant significativement supérieur à 3. La courbe correspondante, tracée en coordonnées logarithmiques est présentée à la figure 6.
Estimation de la croissance pondérale:
L'équation de la croissance en poids en fonction de l'âge est:
Wt = 153,6(1-exp(-1,056(t-0,0675))) 3,75
Le graphique correspondant est présenté à la figure 7.
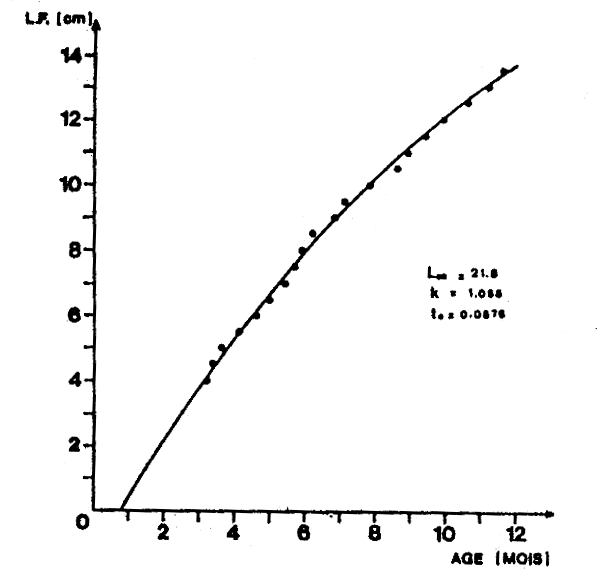
Figure 5 Croissance en longueur d'Engraulis encrasicolus
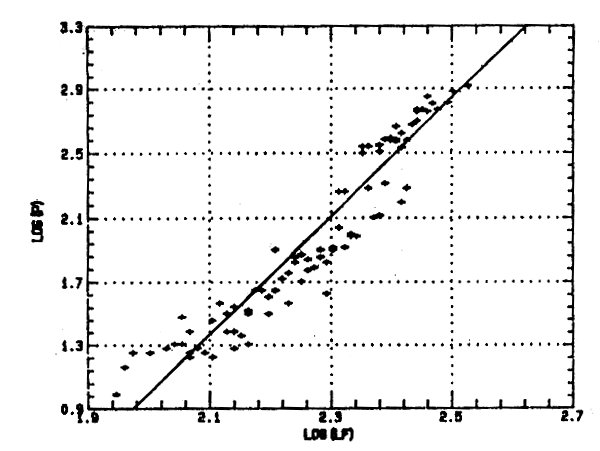
Figure 6 Relation taille-poids d'Engraulis encrasicolus
5.3. DISCUSSION
En Mauritanie, Engraulis encrasicolus a une croissance très rapide. Sa longévité apparente est environ d'un an. Ces résultats diffèrent de tous ceux précédemment obtenus sur cette espèce aussi bien en Mer Méditerranée que dans le Nord Atlantique où la longévité de cette espèce est estimée au moins à trois ans (Tableau 5). Par ailleurs, la comparaison avec les études réalisées par différents auteurs montre, qu'en Mauritanie, E. encrasicolus a une croissance plus élevée pendant la première année; ce qui pourrait s'expliquer par un environnement plus favorable.
Faute de pouvoir contrôler l'âge lu sur les otolithes par une autre méthode et ne disposant d'aucune donnée de croissance sur Engraulis encrasicolus effectuée dans des conditions similaires, une comparaison avec une espèce voisine, Engraulis capensis, a été effectuée. Cette espèce a été choisie parce qu'elle présente beaucoup de caractères communs avec Engraulis encrasicolus:
les tailles maximales observées dans les pêches ne dépassent pas 13–14 cm,
la période de reproduction dure six mois: d'octobre à avril (King, 1977),
le milieu dans lequel vit cette espèce est très similaire à celui d' Engraulis encrasicolus, la région de Walvis Bay (23°S) étant soumise, comme celle du cap Blanc (21°N) à un upwelling quasi permanent.
Prosch (1986) estime que cette espèce, bien que l'on continue à l'appeler Engraulis capensis, est très vraisemblablement la même qu'Engraulis encrasicolus ou tout au moins très apparentée à elle.
Melo (1984) et Prosch (1986), utilisant le comptage des marques journalières sur les otolithes obtiennent respectivement:
| Melo: L00=13,5 | ; | K=1,49; | t0=0,00033 |
| Prosch: L00=14,0 | ; | K=1,58; | t0=0,0012 |
pour des tailles comprises entre 5 et 14 cm.
L'âge maximum observé est de trois ans pour un poisson de 13,2 cm de long. Si au départ L00 n'est pas fixée, sa valeur est alors de 22 cm comme celle obtenue en Mauritanie sur E. encrasicolus. Melo (1984) et Prosch (1986) jugent une telle valeur“ biologiquement impossible” et fixent Loo entre 13,5 et 14 cm correspondant à la taille maximale observée pour cette espèce. Rappelons toutefois que Loo est paramètre mathématique et non biologique.
Le tableau 6 présente les résultats des rétrocalculs âge-longueur à partir des équations établies sur les anchois de l'Afrique du Sud et de Mauritanie. L'on remarquera que le deux espèces ont la même croissance jusqu' à 10 cm, qui correspond à la taille de première mâturité pour les deux espèces2. En revanche, dans les grandes tailles, la différence est grande. Cette différence pourrait être due à une surestimation de la croissance, dans le cas de l'Anchois de Mauritanie, Après la premièe mâturité où les arrêts de croissance ne seraient pas pris en compte.
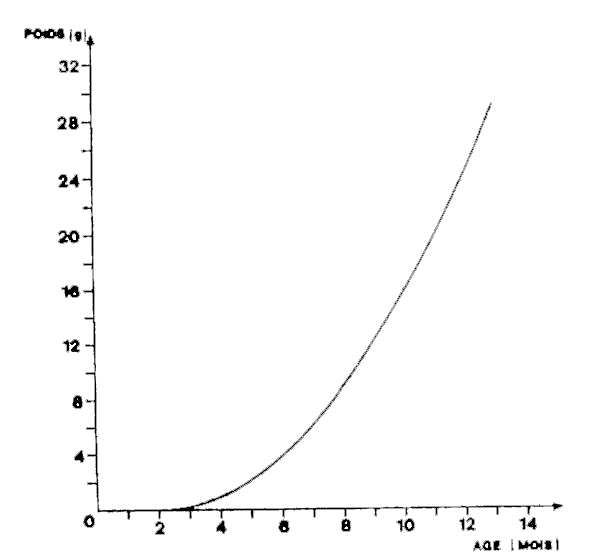
Figure 7 Croissance pondérale d'Engraulis encrasicolus
| Régions | Auteurs | Longueur (cm) | Méthodes | |||
|---|---|---|---|---|---|---|
| 1 an | 2 ans | 3 ans | 4 ans | |||
| Méditérranée | Fage, 1920 | 9.5 | 15.0 | 18 | Scalimétrie | |
| Golfe de Gascogne | Furnestin, 1945 | 12 | 14.0 | 15.8 | Scalimétrie | |
| Oranie (Algérie) | Arrignon, 1966 | 10–12 | 13–15.9 | >16 | Fréquences de taille | |
| Golfe de Gascogne | Guerrault et Avrilla, 1974 | 9.4 | 15.0 | 18.0 | Otolithométrie | |
| Castellon | Suau, 1974 | 10.0 | 14.5 | 17.5 | Fréquences de taille | |
| Golfe de Gascogne | Cort et al., 1976 | 10.5 | 14.4 | 17.2 | Fréquences de taille | |
| Golfe de Cadiz | Rodriguez-Roda, 1977 | 10.6 | 13.8 | 16.2 | Scalimétrie | |
| Golfe de Gascogne | Junquera, 1986 | 11.7 | 15.1 | 17.0 | 18.5 | Otolithométrie |
| Golfe de Gascogne | Uriarte et Astudillo, 1987 | 10.8 | 16.3 | 17.4 | 18.7 | Otholithométrie |
| Mauritanie | Présente étude | 13.5 | - | - | Otolithométrie | |
| Age | Afrique du Sud (taille en cm) | Mauritanie (taille en cm) |
|---|---|---|
| 3 mois | 4.0 | 4.2 |
| 5 mois | 6.5 | 6.5 |
| 8 mois | 10.0 | 10.0 |
| 1 an | 11.1 | 13.5 |
| 2 ans | 12.5 | -- |
| 3 ans | 13.2 | -- |
Ayant vu que la croissance des immatures d'Engraulis capensis était identique à celle d'Engraulis encrasicolus, nous admettrons qu'il en est de même pour les individus matures. En effet, Brothers et al. (1974) et Le Guen (1976) avaient déjà montré que les marques journalières ne sont déposées régulièrement que chez les individus immatures, alors que chez les adultes des arrêts de croissance se produisent notamment au moment de la reproduction. Pour cette même raison, Le Guen (1976) indique que l'âge d'un poisson adulte (par exemple Pseudotolithus elongatus) pouvait être minoré de 30 % . Dans le cas de l'Anchois de Mauritanie, cette sous-estimation peut aussi provenir de la difficulté de lecture des marques localisées près du nucléus et à la périphéric de l'otolithe, qui, dans la plupart des cas, sont difficilement identifiables.
5.4. CONCLUSION
L'étude de la croissance de l'Anchois de Mauritanie à l'aide des marques supposées journalières sur les otolithes a conduit à des résultats douteux chez les poissons matures. Faute de pouvoir comparer ces résultats avec ceux obtenus par une autre méthode et à défaut de pouvoir vérifier directement la périodicité des marques, nous considérons, dans une première approche, que les paramètres de croissance obtenus par Prosch (1986) sur Engraulis capensis sont les mêmes que ceux de l'Anchois de Mauritanie soit:
Loo = 14 cm; K = 1,58 (annuel) et to = 0,0012 (annuel).
L'équation de la croissance linéaire s'écrira donc:
Lt = 14 (1-exp (-1,58(t-0,0012))).
De la même façon, la croissance en poids en fonction de l'âge sera:
Wt = 29,19 (1-exp(1,58 (t-0,0012)))3,75
Toutefois, ces chiffres sont à prendre avec beaucoup de circonspection et doivent être considérés uniquement comme une première approximation qui doit être précisée par la suite par des études plus suivies.
6. CONCLUSION GENERALE
L'étude du cycle sexuel par les méthodes des stades de mâturité et du rapport gonado-somatique a permis de situer la principale période de reproduction de l'Anchois d'avril à octobre. Toutefois, celle-ci ne cesse pas totalement et tous les grands individus (LF > 10 cm) sont susceptibles de pondre quelque soit la saison. Cependant, du fait de l'existence de saisons hydrologiques plus ou moins marquées, certaines périodes semblent plus favorables que d'autres. Ainsi, en Mauritanie, la transition saison froide-saison chaude serait la plus favorable au déclenchement de la ponte de l'Anchois. La reproduction de cette espèce serait donc sous la dépendance principale de l'élévation de la température, mais un autre facteur pourrait jouer un rôle non négligeable, il s'agit de la richesse trophique du milieu.
Chez l'Anchois de Mauritanie, du fait de la longue période de ponte et de la présence d'individus en reproduction tout au long de l'année, les techniques d'étude ne permettent pas de mettre en évidence les valeurs modales sur les histogrammes de fréquence de longueur. Nous avons donc eu recours aux otolithes pour déterminer la croissance de cette espèce, utilisant pour cela la technique des marques supposées journalières. Toutefois, les résultats de cette méthode ont été jugés douteux chez les individus mûrs. En conséquence, la croissance obtenue sur une espèce voisine, Engraulis capensis (l'Anchois d'Afrique du Sud), a été retenue pour décrire celle de l'Anchois de Mauritanie. La connaissance de ce paramètre biologique est indispensable pour une bonne analyse de la dynamique des populations et les résultats provisoires obtenus devront être confirmés.
7. REFERENCES BIBLIOGRAPHIQUES
Abranson N.J. et Tomlinson P.K., 1961 - Fitting a von Bertalanffy growth curve by least squares, including tables of polynomials. Calif. Dept. Fish and Game, Fish. Bull., 116:1–69.
Aldebert Y. et Tournier H., 1971 - La reproduction de la sardine et de l'anchois dans le golfe du Lion. Rev.Trav. Inst Pêches marit., 35(1): 57–75.
Anonyme, 1985 - L'Anchois (Engraulis encrasicolus). In : Rapport du Groupe de travail CNROP/FAO/ORSTOM, Nouadhibou, Mauritanie, 16–27 septembre 1985.
Arbault S. et Lacroix N., 1971 - Aires de ponte de la sardine et de l'anchois dans le Golfe de Gascogne et sur le plateau Celtique. Résultats de 6 années d'études. Rev. Trav. Inst. Pêches marit., 35 (1): 35–56.
Arbault S. et Lacroix N., 1972 - Reproduction de la sardine, de l'anchois et du sprat dans le Golfe de Gascogne en 1971. Comparaison des zones test en 1970 et 1971. Cons. Inter. Expl. Mer. C.M. 1972/J : 5.
Arrignon J., 1966 - L'anchois (Engraulis encrasicolus) des côtes d'Oranie. Rev. Trav. Inst. Pêches marit. 30 (4).
Bas C. et Morales E., 1954 - Algunos datos para el estudio de la biologia de la anchoa Engraulis encrasicolus L. de la Costa Brava. P. Inst. Biol. Apl. XVI: 53–69; 1954.
Bertalanffy L. von, 1938 - A quantitative theory of organic growth (inquires on growth Laws II). Human biol. 10 (2): 181–213.
Brothers E.B., Mathews C.P .et Lasker R., 1976 - Daily growth increments in otoliths from larval and adult fishes. Fishery Bulletin, 74 (1): 1 8.
Chavance P., 1980 - Analyse quantitative de l'ichtyoplancton. Production des aires de ponte de Sardina pilchardus (Walbaum, 1792) et Engraulis encrasicolus (Linnaeus, 1758). Relation avec les pêches (Est Golfe du Lion). Thèse de 3e cycle, Univ. d'Aix Marseille II, 91 p.
Conand C., 1977 - Contribution à l'étude du cycle sexuel et de la fécondité de la sardinelle ronde, Sardinella aurita: pêche sardinière dakaroise en 1975 et premier semestre 1976. Cah. ORSTOM. Sér. Océanogr.: XV (4): 301– 312.
Conand F., 1970 - Distribution et abondance des larves de quelques familles et espèces de poissons des côtes sénégambiennes. Doc. Scient. Prov. Cent. Rech. Océanogr. Dakar Thiaroye, n° 26:52 p.
Cort J. L., Cendrero o. et Iribar X., 1977 - La anchoa, Engraulis encrasicolus(L) del Cantabrico : Resultados de las campanas de 1974, 1975 y 1976. Bul. Inst. Espan. Océanogr. 220(1).34p.
Demir N., 1965 - Synopsis of biological data on anchory, Engraulis encrasicolus(Linnaeus) 1758 (Mediteranean and adjacent Seas). F.A.O. Fish. Synops. 26 (1): 42 p.
Domain F., 1979 - Note sur les périodes de reproduction de quelques espèces démersales du plateau continental sénégambien. Centre Rech. Océanogr. Dakar Thiaroye, Doc. Scient., 68: 111–126.
Domanevsky L.N., 1968 - Spawning pecularities of fish from the North Western Coast of Africa. Cons. Intern. Expl. Mer, C.M 1968/G 3: 8 p.
Fage L., 1911 - Recherches sur la biologie de l'anchois, Engraulis encrasicolus (Linné): races-âge-migration. An. Inst. Océanogr. Paris, 2(4): 140 p.
Fage L., 1920 - Engraulidae, Clupeidae. Rep. Danish Oceanogr. Exped. 1908–1910 to the Mediterranean and Adjacent Seas. 2(A9): 140 p.
Fontana A., 1969 - Etude de la mâturité sexuelle des sardinelles, Sardinella eba (Val.) et Sardinella aurita (C. et V.) de la région de Pointe Noire. Cah. ORSTOM. Sér. Océanogr. 7(2) :101–114.
Furnestin J., 1945 - Note préliminaire sur l'Anchois (Engraulis encrasicolus) du Golfe de Gascogne. Rev. Trav. Off. Sci. Tech. Pêches marit., 13 (1–4): 197–209.
Furnestin J. et Furnestin M.L., 1959 - La reproduction de la sardine et de l'anchois des côtes Atlantiques du Maroc (saisons et aires de ponte). Rev. Trav. Inst. Pêches marit, 23 (1): 79–102.
Guerault D. et Avrilla J.L., 1974 - L'anchois du Golfe de Gascogne : taille, âge, croissance. Cons. Intern. Explor. Mer, C.M. 1974/J : 17.
Junquera S., 1986 - Pêche de l'Anchois (Engraulis encrasicolus) dans le Golfe de Gascogne et sur le littoral atlantique de Galice depuis 1920. Variations quantitatives. Rev. Trav. Inst. Pêches marit, 48 (3 et 4): 133–142.
King D.P.F., 1977 - Distribution and relative abundance of eggs of the South West African pilchard Sardinops ocellata and anchovy Engraulis capensis, 1971–1972. Fish. Bull. S. Afr. 9: 23–31.
Le Clus F., 1979 - Fecundity and maturity of Anchovy Engraulis capensis off South West Africa. Fish. Bull. S. Afr. 11: 26–38.
Le Guen J. C., 1971 - Dynamique des populations de Pseudotolithus (Fonticulus) elongatus (Bowd, 1825). Poissons Sciaenidae. Cah. ORSTOM. Sér. Océanogr., 9 (1): 3–84.
Le Guen J. C., 1976 - Utilisation des otolithes pour la lecture de l'âge de Sciaenidés tropicaux. Marques saisonnières et journalières. Cah. ORSTOM. Sér. Océanogr., 4 : 331–338.
Maigret J., 1972 - Campagne expérimentale de pêche des sardinelles et autres espèces pélagiques (juillet 1970 octobre 1971). I. Observations concernant l'océanographie et la biologie des espèces. Rep. Islam. Mauritanie; Minist. des Pêches et Mar. March, Laborat. Pêches; Nouadhibou. (S.C.E.T.) International, 148 p.
Marchal E., 1966 - Oeufs, larves et post larves de l'Anchois du Golfe de Guinée, Anchoviella guineensis (Blache et Rossignol). Doc. Sci. Prov. ORSTOM. Abidjan; 5 :15 p.
Melo Y.C., 1984 - Age studies on Anchovy Engraulis capensis (Gilchrist) off South West Africa. S. Afr. J. mar. Sci. 2: 19–31.
Navaz J. M. et Lozano Cabo F., 1966 - Données sur la taille, l'âge et la croissance de l'Anchois (Engraulis encrasicolus (L.), de la côte basque espagnole. Cons. Inter. Explor. Mer. C.M. 1966/J:6.
Panella G., 1971 - Fish otoliths : daily growth layers and periodical patterns. Science, 173 :1124–1127
Panella G., 1974 - Otolith growth patterns : and aid in age détermination in temperate and tropical fishes. In : The ageing of Fish : 28–39. Bagenal (edit) Uniwin Brothers Ltd. England.
Prosch R.M., 1986 - Early growth in length of the anchovy Engraulis capensis (Gilchrist) off South Africa. S. Afr. J. mar.Sci. 4 : 181–191.
Rê P., Farinha A. et Meneses I., 1983 - Anchovy spawning in portuguese estuaries (Engraulis encrasicolus, Pisces : Engraulidae). Cybium, 1983, 7(1) : 29–38.
Rodriguez Roda J., 1977 - El boquerón, Engraulis encrasicolus (L.), del Golfo de Càdiz. Inv. Pesq. 41 (2): 523–542.
Sinovcic G., 1978 - On the ecology of anchovy, Engraulis encrasicolus (L.)., in the Central Adriatic. Acta Adriatica. 19 (2).
Sobral M.C., 1975 - Contribution à l'étude du plancton et de la reproduction de la sardine et de l'anchois dans la côte portuguaise. Cons. Intern. Explor. Mer. C.M. 1975/J: 11
Suau P., 1979 - Biologia del boqueròn, Engraulis encrasicolus de las costas de Castellon (E de Espana). Inv. Pesq. 43 (3): 601–610.
Uriarte A. et Astudillo A., 1987 - The anchovy in the Bay of Biscay : new data and analysis of the fishery: 1974–1987. Intern. Conc. Explor. Seas. C.M. 1987 /H :20
![]()
![]()
![]()