Virus and virus-like pathogens were discovered because they produce diseases. For instance, in the 1930s citrus growers in Argentina noted that sweet orange trees on sour orange rootstock declined and died. This disease was called tristeza. Two other observations were made: trees on certain rootstocks other than sour orange did not decline and sour orange trees and sweet orange trees on their own roots showed no symptoms. Intensive research into the cause of tristeza disease, which eventually killed 20 million trees in South America, led to the discovery in the 1940s that the tristeza agent is transmissible by aphids and graft inoculation. The agent was eventually identified as a virus in the 1960s. It is now recognized that CTV has probably been present in India and the Far East for several centuries although the virus did not attract attention there as the infected citrus trees showed no symptoms and were tolerant of CTV. In Japan, there are many satsuma trees grafted on trifoliate orange (Poncirus trifoliata), which are infected by CTV but remain symptomless.
These examples show that, for a given virus or virus-like pathogen, not only are there susceptible hosts and host associations, but also tolerant hosts and host associations. In the case of CTV, sweet orange and mandarin species are tolerant of the virus. Sour orange is also tolerant, but when sweet orange or mandarin is grafted on sour orange, the association obtained is susceptible. In fact, most commercial citrus species, except lemon, give a susceptible association when grafted on sour orange. When trifoliate orange and rough lemon are used as rootstocks for sweet orange, mandarin or satsuma, however, the trees are tolerant of CTV. Rough lemon is a tolerant rootstock. Trifoliate orange is more than tolerant, it is resistant to CTV.
Resistant plants are those in which the pathogen is unable to multiply, and such plants do not act as hosts for the agent. Unlike tolerant plants, resistant plants are not only symptomless, they are also free of the agent. In tolerant host plants, the pathogen multiplies without inducing symptoms, but the plants nevertheless carry the agent. Therefore, tolerant plants are sometimes called symptomless carriers. Mandarin is a symptomless carrier of CTV, for instance.
Susceptible host plants are those in which the agent both multiplies and induces symptoms. In tolerant and susceptible plants, the agent is present throughout the host. Such plants are systemic hosts. Furthermore, with certain viruses, such as tobacco necrosis virus (TNV), hypersensitive host plants are known.
When TNV is mechanically inoculated on a leaf, certain leaf cells become infected. The hypersensitive reaction of the plant lies in the production of necrotic rings of dead cells around the points of entry of the virus. These so-called necrotic local lesions prevent the virus from becoming systemically distributed throughout the plant. Such plants are thus free of virus except in the necrotic local lesions. TNV induces necrotic local lesions in several citrus species when it is mechanically inoculated on their leaves.
Tables 5 and 6 list, in alphabetical order, rutaceous and non-rutaceous plants and indicate their susceptibility to or tolerance of the various virus and virus-like pathogens of citrus. Inversely, Tables 7 and 8 list the virus and virus-like diseases of citrus and indicate the symptomatic or symptomless plants. Most citrus species given as symptomless are tolerant. Resistant species are indicated but sometimes it is not known whether species are tolerant or resistant.
The above discussion considered species as susceptible, tolerant or resistant. The situation is, however, more subtle. For instance, there are many sweet orange varieties and many strains of CTV. While Washington navel and Valencia sweet oranges are tolerant of Brazilian strains of CTV, these same strains induce severe symptoms (stem pitting) in Pera sweet orange. Certain strains of CTV from the Far East produce symptoms even on Washington navel and Valencia late sweet orange. Small-fruited acid lime (Kagzi, Key, Mexican, West Indian) seedlings are susceptible to CTV and show CTV-specific symptoms - leaf vein clearing and stem pitting. However, a strain of CTV, discovered recently in Corsica (France), produces no symptoms whatsoever in lime seedlings even though the virus strain multiplies well in the infected lime plants.
These examples show that a given host plant can be tolerant or susceptible, depending on the strain involved and that, vice versa, a given strain can induce symptoms in certain varieties of a species but be symptomless in others. Hence, the distinction between tolerance and susceptibility is not as clear as it would appear to be. According to the strains considered, a given host may show severe, moderate or mild symptoms or even none at all. Moreover, a strain giving a severe reaction in one citrus species may well give a moderate or mild reaction in a different species. When referring to the severity of a strain, therefore, one must indicate the nature of the host in which the severity of the strain is evaluated. For instance, in South Africa there are strains of CTV from sour orange or lemon sources that induce severe stem pitting in acid lime seedlings but no reaction (decline) in sweet orange on sour orange rootstock.
Various strains of an agent can infect a single plant. In Spain, up to 23 different sub-isolates of CTV have been obtained from a single Mexican lime seedling carrying an aphid-transmitted field isolate of the virus. These results suggest that a field plant is rarely infected by a pure strain of an agent, but rather by a population of various strains.
Cross-protection or pre-immunization is based on interference between different strains of a virus in a single plant and refers to the phenomenon in which infection of a plant by a mild strain of a virus protects the plant against a severe strain of that virus. A classical case of cross-protection concerns Pera sweet orange in Brazil. This variety is sensitive to
Brazilian strains of CTV in that it shows severe stem pitting in the orchard after natural aphid transmission of the virus. When a nucellar clone of Pera sweet orange was infected with a mild CTV strain, this preimmunized clone showed only mild CTV stem pitting in the field as it was protected against the severe, aphid-transmitted strains of CTV. Over the last 20 years, the preimmunized Pera sweet orange clone has yielded more than 50 million trees without any loss of cross-protection. Today, efforts to find mild CTV strains for cross-protection are carried out in many CTV-affected countries. While cross-protection with mild strains against severe stem pitting strains has generally met with success, mild strains capable of protecting sweet orange trees on sour orange rootstock against CTV-induced decline have yet to be found.
The above considerations apply not only to viral agents, but also to viroids. A good example is that of the viroid responsible for potato spindle tuber (PSTV). This viroid also affects tomatoes. Various PSTV isolates cause mild, moderate, severe or lethal symptoms in tomato. Assuming the isolate that produces the mildest symptoms to be standard, all other isolates differ in certain nucleotides which are either exchanged, inserted or deleted, with the total number of nucleotides (359) being strictly maintained. All these changes are found in only three regions of the PSTV primary nucleotide sequence. For instance, the deletion of a uridylic residue (-U) in region I and insertion of an adenylic residue (+A) in region III changes a mild isolate into a severe one. A lethal isolate has the following modifications: -G and +A in region I,-U +AA in region II, and +C -C -U in region III. In the case of CEV, similar differences in the nucleotide sequences of mild, moderate and severe isolates probably exist. However, viroids other than CEV are also involved in what is called citrus exocortis disease.
When a series of stable exocortis isolates (judged by symptoms on inoculated citron plants) were recovered from different citrus sources in California (United States of America and Spain and then analysed by a sequential polyacrylamide gel electrophoresis (sPAGE) procedure, some isolates contained CEV but always in association with additional viroids; other isolates did not show the presence of CEV but harboured some of the other viroid RNAs. The effect on P. trifoliata rootstocks of the various viroids listed in Table 3 on p. 12, when singly inoculated into trees on this rootstock, is only partially known, and Table 3 therefore has question marks in the column for P. trifoliata. However, the reactions of P. trifoliata and Troyer citrange rootstocks to the individual viroids are being studied. It is already known that CV-IIa alone induces bark cracking, not bark scaling, on P. trifoliata grafted with sweet orange. This viroid causes the mildest symptoms in citron (see Tables 2 and 3). Recent observations have shown that CV-Ia causes inverse pitting (pinholing) on P. trifoliata and that CV-IIIb causes stem grooving (finger marks) on the same host (see Table 3).
A striking synergism has been observed between CVEV woody gall and citrus yellow vein virus. The symptoms of yellow vein are greatly enhanced in host plants (lime, sweet orange, lemon, rough lemon) also infected with CVEV. This synergism has been used in the identification of virus strains. An unexpected phenomenon of protection by CVEV against CTV has been observed in Japan.
Interferences and interactions among citrus viroids have also been found. Certain citrus viroids (CV-IIa) or viroid combinations (CVIIa + CV-III) may mask or suppress symptom expression of cachexia (CV-IIb). Indeed, there seems to be mutual antagonism between CV-IIa, the viroid producing mild symptoms on citron, and CV-IIb, the cachexia viroid. If CV-IIa prevents CV-IIb from becoming established in Parson's Special mandarin, for instance, there will be no symptom expression of cachexia in the mandarin. This may explain certain irregularities and inconsistencies in symptom expression that have been observed in routine indexing of cachexia on Parson's Special mandarin.
Strain differences have also been established in the case of uncharacterized virus-like agents. For instance, while all isolates of the cristacortis agent induce severe stem pitting on Orlando and Wekiwa tangelos and sour orange, only some isolates cause symptoms on Clementine and a single isolate produces symptoms on lemon.
Bar-Joseph, M., Marcus, R. & Lee, R.F. 1989. The continuous challenge of citrus tristeza virus control. Ann. Rev. Phytopathol., 27: 291-316.
Bové, C., Vogel, R., Albertini, D. & Bové, J.M. 1988. Discovery of a strain of tristeza virus (K) inducing no symptoms in Mexican lime. In Proc. 10th Conf: IOCV, p. 14-16. Riverside, Univ. Calif.
Calavan, E.C. & Weathers, L.G. 1961. Evidence for strain differences and stunting with exocortis virus. In Proc. 2nd Conf: IOCV, p. 26-33. Gainesville, Univ. Fla. Press.
Calavan, E.C., Christiansen, D.W. & Weathers, L.G. 1965. Variability of cachexia reactions among varieties of rootstocks and within clonal propagations of citrus. In Proc. 3rd Conf IOCV, p. 78-85. Gainesville, Univ. Fla. Press.
Costa, A.S. & Muller, G.W. 1980. Tristeza control by cross protection: a US-Brazil cooperativesuccess. Plant Dis., 64: 538-541.
Costa, A.S., Grant, T.J. & Moreira, S. 1954. Behaviour of various citrus rootstockscion combinations following inoculation with mild and severe strains of tristeza virus. Proc. Fla Hort. Soc., 67: 26-30.
Duran-Vila, N., Pina, J.A. & Navarro, L. 1991. Exclusion and/or uneven distribution of viroids in four citrus hosts. In Proc. 11th Conf: IOCV, p. 219-223. Riverside, Univ. Calif.
Duran-Vila, N., Roistacher, C.N., Rivera-Bustamante, R. & Semancik, J.S. 1988. A definition of citrus viroid groups and their relationship to the exocortis disease. J. Gen. Virol., 69: 3069-3080.
Duran-Vila, N., Pina, J.A., Ballester, J.F., Juarez, J., Roistacher, C.N., Rivera-Bustamante, R. & Semancik, J.S. 1988. The citrus exocortis disease: a complex of viroid-RNAs. In Proc. 10th Conf IOCV, p. 152-164. Riverside, Univ. Calif.
Frolich, E.F., Calavan, E.C., Carpenter, J.B., Christiansen, D.W. & Roistacher, C.N. 1965. Difference in response of citron selections to exocortis virus infection. In Proc. 3rd Conf: IOCV, p. 113- 118. Gainesville, Univ. Fla. Press.
Grant, T.J. 1959. Tristeza virus strains. In J.M. Wallace, ed. Citrus virus diseases. p. 45-55. Berkeley, Div. Agric. Sci., Univ. Calif.
Grant, T.J. & Higgins, R.P. 1957. Occurrence of mixture of tristeza virus strains in citrus. Phytopathol., 47: 272-276.
Hughes, W.A. & Lister, C.A. 1949. Lime disease in the Gold Coast. Nature, 164: 880.
Kitajima, E.W., Silva, D.M., Oliveira, A.R., Muller, G.W. & Costa, A.S. 1964. Thread-like particles associated with tristeza disease of citrus. Nature, 201: 1011-1012.
Koizumi, M. & Sasaki, A. 1980. Protection phenomena against tristeza in trees preinoculated with vein enation virus. In Proc. 8th Conf IOCV, p. 48-50. Riverside, Univ. Calif.
Koizumi, M., Kano, T., Ieki, H. & Mae, H. 1988. China laurestine: a symptomless carrier of satsuma dwarf virus which accelerates natural transmission in the fields. In Proc. 10th Conf IOCV, p. 348352. Riverside, Univ. Calif.
McClean, A.P.D. 1974. The tristeza virus complex. In Proc. 6th Conf. IOCV, p. 59-66. Berkeley, Div. Agric. Sci., Univ. Calif.
Meneghini, M. 1946. Sobre a natureza e transmisiblade da doença "tristeza" dos citrus. O Biológico, 12: 285-287.
Moreno, P., Guerri, J. & Ballester Olmos, J.F. 1991. Segregation of citrus tristeza virus strains evidenced by double stranded RNA analysis. In Proc. 11th Conf: IOCV, p. 20-24. Riverside, Univ. Calif.
Muller, G.W. & Costa, A.S. 1968. Further evidence on protective interference in citrus tristeza. In Proc. 4th Conf: IOCV, p. 71-82. Gainesville, Univ. Fla. Press.
Muller, G.W. & Costa, A.S. 1977. Tristeza control in Brazil by preimmunization with mild strain. Abstr. Int. Citrus Congr., Orlando, Florida.
Muller, G.W., Rodriguez, O. & Costa, A.S. 1968. A tristeza virus complex severe to sweet orange varieties. In Proc. 4th Conf: IOCV, p. 64-71. Gainesville, Univ. Fla. Press.
Muller, G.W., Costa, A.S., Castro, J.L. & Guirado, N. 1988. Results from preimmunization tests to control the Capao bonito strain of tristeza. In Proc. 10th Conf: IOCV, p. 82-85. Riverside, Univ. Calif.
Pina, J.A., Duran-Vila, N. & Navarro, L. 1991. Interference of citrus viroids with cachexia symptoms on Parson's Special mandarin. In Proc. 11th Conf: IOCV, p. 206-208. Riverside, Univ. Calif.
Rodriguez, O., Moreira, S., Salibe, A.A. & Sobrinho, J.T. 1972. Further study of the tolerance to citrus tristeza virus of citrus varieties suitable for rootstock in Brazil. In Proc. 5th Conf: IOCV, p. 121-124. Gainesville, Univ. Fla. Press.
Roistacher, C.N., Dodds, J.A. & Bash, J.A. 1988. Cross protection against citrus tristeza seedling yellows and stem pitting viruses by protective isolates developed in greenhouse plants. In Proc. 10th Conf: IOCV p. 91-100. Riverside, Univ. Calif.
Rossetti, V., Fassa, T.G. & Nakadaira, J.T. 1965. Reaction of citrus varieties to the stem pitting of Pera orange. In Proc. 3rd Conf: IOCV, p. 46-48. Gainesville, Univ. Fla. Press.
Salibe, A.A. 1965. Occurrence of stem pitting in citrus types in Brazil. In Proc. 3rd Conf: IOCV, p. 40-45. Gainesville, Univ. Fla. Press.
Sanger, H.L. 1984. Minimal infectious agents: the viroids. p. 281 -33. In B.W. Mahy & J.R. Pattison, eds. The microbe. Part I: Viruses. Cambridge, Cambridge University Press.
Semancik, J.S. 1988. Citrus exocortis disease - 1976 to 1986. In Proc. 10th Conf: IOCV, p. 136-150. Riverside, Univ. Calif.
Semancik, J.S., Gumpf D.J. & Bash, J.A. 1991. Interactions among group 11 citrus viroids: a potential for protection from cachexia disease. In Proc. 11th Conf: IOCV, p. 189-195. Riverside, Univ. Calif.
Speroni, H.A. & Frezzi, M. 1939. Podredumbre de la raicilla del naranjo. Rev. Corpor. Frutic. Argentina, 5: 25-31.
Vogel, R. & Bové, J.M. 1976. Evidence for the existence of strains of the cristacortis pathogen. In Proc. 7th Conf IOCV, p. 101-104. Riverside, Univ. Calif.
Vogel, R. & Bové, J.M. 1991. Strains of cristacortis characterized by specific host range susceptibility. In Abstracts' brochure, Proc. 11th Conf: IOCV. Riverside, Univ. Calif.
Chapter 3: Transmission and spread of virus and virus-like citrus pathogens
Polyembryony of citrus seeds
Bud
propagation
Mechanical
transmission
Insect and mite transmission
Other
means of transmission
Bibliography
Most virus and virus-like citrus pathogens are not seed-borne
The seeds of many varieties of the genera Citrus Fortunella and Poncirus, used as scions and rootstocks, have two types of embryo: a single, sexual (zygotic) embryo; and several non-sexual, nucellar embryos. The origin of these embryos is as follows. In a mature ovule of a citrus flower, the embryo sac containing the female gamete or egg cell (n chromosomes) is surrounded by the nucellus; at the apex of the nucellus is an opening, the micropyle. On pollination, when the pollen tube has reached the embryo sac by way of the micropyle, the nucleus of a male gamete or sperm eel] (n chromosomes) passes into the embryo sac where it fuses with the egg cell nucleus to yield the fertilized egg (2n chromosomes), thus accomplishing fecundation. The embryo resulting from the development of the fertilized egg is the sexual, zygotic embryo.
The extra embryos derive from somatic cells (2n chromosomes) of the nucellus. These nucellar embryos grow into the embryo sac and lie alongside the sexual embryo. Although there may be exceptions, pollination is usually necessary for the formation of nucellar embryos. Since nucellar embryos develop asexually by ordinary mitotic division of 2n chromosome cells of the nucellus, no male cells contribute to their formation and no reduction division (meiosis) occurs in the seed-parent cells which produce them. Nucellar seedlings, therefore, not only inherit from the seed parent alone but they are actually identical with it in genetic constitution, except for possible differences brought about by somatic (bud) variation. The growth of the nucellar embryos into seedling plants thus represents a true-to-type vegetative propagation of the seed parent or mother tree.
The zygotic embryo must compete for space and nutrients with the nucellar embryos. The result of this competition depends on the number of nucellar embryos, their time of starting, their location and the relative genetic vigour of the two kinds of embryo. In many varieties, a seed produces only one or more nucellar seedlings and no zygotic seedlings, indicating that the sexual embryo was crowded out or was simply too weak to survive. In such cases, all or nearly all of the seedlings produced will be from nucellar embryos and the seed parent will "breed true" with respect to all or nearly all of its offspring.
Nucellar polyembryony is used in citriculture for two purposes: the production of homogeneous, ready-to-graft rootstock seedlings and the production of scion seedlings from which nucellar clones can be developed. Both the rootstock seedlings and the scion seedlings will not only be true to type, they will also be free of virus and virus-like agents. Indeed, while several non-rutaceous viruses are known to be transmitted through the seed embryos (seed-borne viruses), transmission of virus and virus-like agents of citrus through embryos of citrus seeds (seed transmission) is of minor importance. Seed transmission of citrus crinkly leaf virus (CCLV) and the related CVV has been observed but is rare. Observations indicating psorosis (concave gum) transmission by seed of Carrizo and Troyer citranges and Poncirus trifoliata have been reported. SDV is transmitted through the seed of bush bean, but not citrus. Seed transmissions of exocortis have not been confirmed. None of the other virus and virus-like agents of citrus seems to be seed-transmitted.
Phloem-restricted prokaryotes, in particular, are not transmitted from an infected mother tree to seedling progeny plants via the seeds. This absence of seed transmission does not imply, however, that the seeds as a whole are free of the pathogens. CVV was detected at a high level in the seed-coats of Volkamer lemon seeds. It is also well known that homogenates of seeds from stubborn-infected trees are good sources for the isolation and culture of Spiroplasma citri, the causal agent of stubborn. It has been shown that, in this case, the embryo is free of the spiroplasma but the seed-coats, or, more precisely, their phloem tissue where the spiroplasma is located, are not. Seeds from stubborn- or greening-affected trees often show various degrees of abortion, probably related to the presence of the pathogens in the seed-coats.
A grafted citrus tree is composed of two elements: a rootstock and a scion. The rootstock is practically always derived from a seedling which is, in essence, free of virus and virus-like pathogens. Hence, when grafted trees are found to be infected by a virus or virus-like pathogen, the infection often comes from the scion bud that was grafted on the rootstock seedling, not from the seedling itself. Indeed, the propagation of buds taken from trees infected by virus and virus-like pathogens is one of the main means by which these agents have been, and still are being, spread. In this form of dissemination, humans are the vector! Spreading graft-transmissible agents of citrus in this way can easily be prevented by taking buds only from trees known to be free of virus and virus-like agents.
However, under certain circumstances, the use of budwood free of such agents is not enough. A tree obtained from a healthy rootstock seedling and a "clean" bud can become infected by a given pathogen as a result of mechanical and/or insect vector transmission. Hence, three classes of virus and virus-like pathogens of citrus can be considered: i) agents that are mechanically transmitted (see Table 9); ii) agents that are transmitted by insect and mite vectors (see Table 10); and iii) agents that are transmitted neither mechanically nor by insect or mite vectors.
Under orchard conditions, mechanical transmission occurs when the pathogens (cachexia and exocortis viroids) are carried unintentionally from an infected tree to a healthy one by way of budding knives, pruning scissors or hedging or topping machines. In the laboratory, mechanical inoculation can be carried out in several ways. In the stem-slash procedure, slashing the stem of an infected plant with a knife or scalpel will contaminate the blade, which is then used to slash the stem of a healthy plant.
TABLE 9 Mechanically viruses and viroids of citrus
| Disease or agent | Type of agent | Mechanically transmitted to: |
| Algerian navel orange virus | Virus | Sweet orange, Chenopodium quinoa |
| Cachexia | Viroid | Etrog citron, cucumber |
| Dweet tangor agent | Virus | Etrog citron |
| Exocortis | Viroid | Etrog citron, gynura, cucumber, chrysanthemum, tomato |
| Infectious variegation-Crinkly leaf | Ilarvirus | Lemon, cowpea. cucumber, Gomphrena globosa, Crotalaria spectabilis |
| Leaf rugose | Ilarvirus | As for Infectious variegation plus Chenopodium quinoa, Phaseolus vulgaris, Petunia hybrida, Nicotiana tabacum |
| Psorosis (Calif) | Virus | Etrog citron, Nicotiana benthamiana, Nicotiana megalosiphon, C quinoa, Capiscum annuum and frutescens |
| Psorosis P 203 (Psorosis A) | Virus | Etrog citron C quinoa |
| Psorosis B | Virus | Etrog citron C quinoa, Gomphrena globosa |
| Ringspot | Virus | Etrog citron, C quinoa (not system c), G globosa, Crotalaria spectabilis, Petunia hybrida, Zinnia elegans cowpea, bean |
| Satsuma dwarf | Virus | Sesame, cowpea, bean, petunia, tobacco, Chenopodium spp, Physalis floridana, G. globosa |
| Tatterleaf-Citrange stunt | Virus | Etrog citron rough lemon seedling, Valencia late sweet orange on Poncirus trifoliata, cowpea, bean, C quinua (systemic) |
| Tobacco necrosis virus citrus isolate | Virus | Lemon, sweet orange sour orange Orlando tangelo, Etrog citron, acid lime cowpea, cucumber, Xanthni n.c. tobacco |
| Tristeza | Closterovirus | Etrog citron, sweet orange |
Notes:
1 Phloem-restricted prokaryotes are not mechanically
transmissible.
2 Serologically related to infectious
variegation-crinkly leaf virus.
This technique has been found to be most effective in the mechanical transmission of exocortis and cachexia viroids as well as tristeza virus to citron plants. To prevent such transmission occurring in the field or laboratory, budding knives and other such tools can easily be disinfected with domestic bleach diluted to give a 1 percent sodium hypochlorite solution.
Herbaceous plants can be mechanically infected by rubbing sap or extracts from infected plants on the leaves of healthy plants in the presence of carborundum powder. the presence of this fine abrasive results in mild mechanical damage to the cell wall, a condition that allows the pathogen to enter the cell (leaf-abrasion method). Depending on the plant and/or the pathogen in question, the inoculated leaves will show either chlorotic or necrotic local lesions and the infection will either become systemic or remain local.
TABLE 10 Insect and mite vectors of virus and virus-like disease agents of citrus
Mechanically inoculated plants may be tolerant of the inoculated agent and remain symptomless, such as citron infected with cachexia viroid. Table 9 lists the citrus pathogens that have been mechanically transmitted and some of the mechanically infected plants are also indicated. Note that phloemrestricted prokaryotes have not been mechanically transmitted. In the case of exocortis and cachexia viroids, the efficiency of mechanical transmission varies according to the donor and receptor hosts.
Several virus and virus-like citrus pathogens are transmitted by insect vectors (see Table 10). Healthy field trees in a given region will become infected by one or other of these pathogens if the agent and the vector occur together in the region. For instance, CTV was not present in Argentina in the 1900s but the tropical citrus aphid, Toxoptera citricida, was. Around the 1920s, citrus budlings infected with CTV were introduced, probably from South Africa. From then on, the spread of CTV occurred swiftly as T. citricida turned out to be a very efficient vector of CTV, with one viruliferous aphid being enough for transmission to occur. After invading Argentina and Brazil, where CTV killed 20 million trees between 1930 and 1950, the virus is now widespread in the northern parts of South America. In Venezuela it has destroyed almost 500 000 trees and it has recently been found in Costa Rica, Honduras, Panama, El Salvador, Nicaragua, Belize and Guatemala. F. citricida has been isolated in Panama. Costa Rica, Salvador, Nicaragua and Martinique in the French West Indies.
Aphid species other than T. citricida are also vectors of CTV. One of the second most important CTV vectors is Aphis gossypii, which can transmit destructive isolates of CTV at relatively high rates of efficiency. In Israel, CTV strain VT was transmitted by A. gossypii at a much higher efficiency rate (40 percent) than other isolates (5 percent). In Spain, A. gossypii transmitted the common CTV isolate T-300 at a higher efficiency rate 190 percent) than the seedling yellows isolate T-387 (60 percent). These results underline the fact that certain virus strains are transmitted at higher rates than others by a given vector.
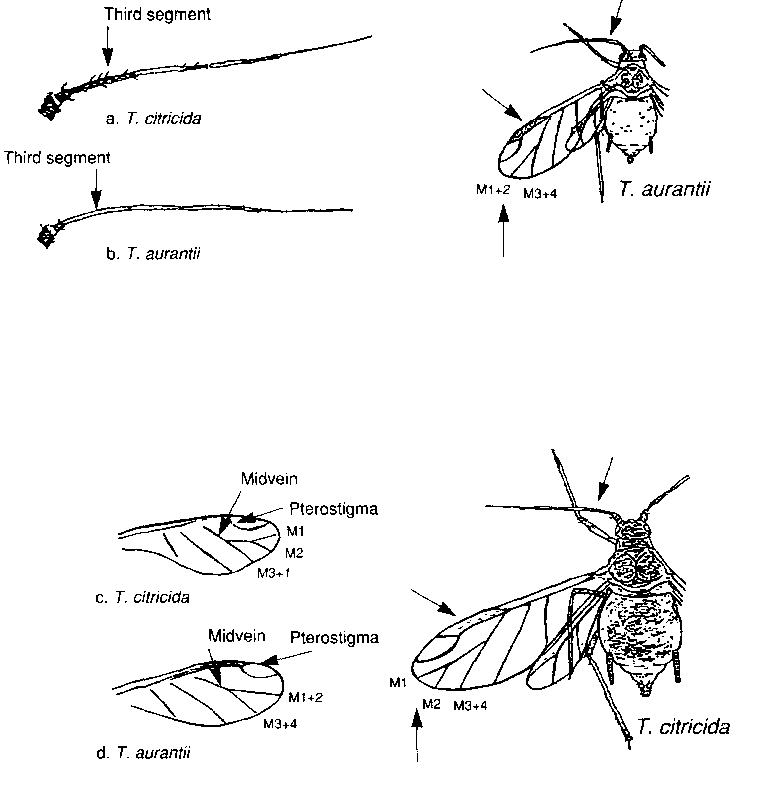
Other CTV aphid vectors are: Aphis citricola (syn. A. spiraecola), Toxoptera aurantii and My-us persicae. A. citricola is a less efficient vector than A. gossypii but it is frequently abundant on citrus and may be more important in spreading the virus in the field than laboratory transmission tests indicate. In Spain, A. citricola is the most abundant aphid species in citrus areas and is believed to be responsible for the natural spread of CTV in the field. In the Mediterranean and the Near East, including Pakistan, T. citricida has not been reported but A. gossypii and some of the other aphid vectors are pandemic from Morocco to Pakistan. They are responsible for the spread of CTV in Mediterranean countries, such as Spain and Israel, where this virus has been introduced. They could become the major CTV vectors in other countries of the Mediterranean area and the Near East if CTV were introduced. In other words, even in the absence of T. citricida, other vectors of CTV are present in the Mediterranean and Near East regions. Hence, efforts have to be made to keep CTV out of countries where it is not yet present. Both T. citricida and T. aurantii are black aphids. The drawings on p. 36 show how one can easily distinguish between them on the basis of the venation of their front wings and the pigmentation of their antennae.
Other insect vectors of citrus viruses are indicated in Table 10. Note that all insect vectors of citrus viruses are aphids, except in the case of leprosis, where mites are responsible for transmission.
In the case of the phloem-restricted prokaryotes, the insect vectors are leafhoppers and/or psyllids. the two psyllid vectors of the bacterium-like organism (BLO) associated with greening are restricted to rutaceous hosts, both cultivated and wild. In contrast, the leafhopper vectors of S. citri (Figs 34 and 35) are polyphagous and live primarily on non-rutaceous hosts, visiting citrus only occasionally and under certain conditions. This explains why natural spread of the greening BLO is much more efficient than that of the stubborn spiroplasma, except under special conditions. For instance, when young citrus trees are grown in the presence of the weed Salsola kali (Fig. 174), high infection rates of citrus with S. citri can be observed, as in the Syrian Arab Republic. This weed is the major host plant of the two leafhopper vectors of the spiroplasma. Thus, the presence of S. kali favours high populations of the leafhoppers which, in turn, favour the spread of the spiroplasma, in particular to young citrus trees nearby. In Oman and the United Arab Emirates, witches' broom disease of lime, an MLO disease, is spreading very fast. The putative insect vector, probably a leafhopper or a psyllid, has not yet been identified but monoclonal antibodies (MAs) and DNA probes for the detection of the MLO have now become available and enable identification of the vector(s). Indeed, in May 1991 in Oman, and in February 1993 in the United Arab Emirates, the leafhopper Hishimonus phycitis was found in great numbers on arid lime trees and it might be the vector of the witches' broom MLO. This is the first time that this leafhopper has been found in the Arabian peninsula although it is a well-known leafhopper in India where it transmits the MLO of eggplant little leaf disease.
A situation similar to that in Argentina prior to the introduction of CTV exists today in Brazil for greening. An abundant population of Diaphorina citri, the Asian psyllid vector of the greening BLO (Fig. 37), thrives on citrus throughout the State of Sao Paulo, and probably elsewhere. Introduction of the greening BLO would probably result in an epidemic as bad as that of tristeza in the 1930s.
Aphids, leafhoppers and psyllids, the major insect vectors of virus and virus-like disease agents of citrus, are members of the Homoptera. All members of the Homoptera feed in the same way, by means of sucking mouthparts, which consist of two pairs of stylets- mandibles and maxillae - sliding in the grooved labium. The mandibular stylets lie outside the maxillary stylets. Grooves on the inner faces of the maxillary stylets match in such a way as to form two canals, down one of which saliva is injected into the plant while a mixture of sap and saliva passes up the other. Thus, such insects are ideally equipped for pathogen transmission. The relationships between these homopterous vectors and the pathogens are of three major types (see Table 11). the phloem-restricted prokaryotic agents (spiroplasmas, MLOs and BLOB) are persistent in the vectors in which they multiply. Once infected, the vectors remain infected and can transmit the pathogen throughout their lifetime. The leafhopper and psyllid vectors acquire the pathogen when feeding on infected plants. These insects suck up the rich sap in the sieve tubes of their specialized mouth parts. If the sieve tubes contain pathogens, such as spiroplasmas, MLOs or BLOB, they are also sucked up, and subsequently enter the gut, cross the intestinal membrane, reach the blood (haemolymph) of the insect, multiply and invade tissues and organs, eventually reaching the salivary glands.
Only when the pathogen is established in the salivary glands can the insect transmit it to the healthy plants on which it feeds. Indeed, the saliva formed in the salivary glands is required to lubricate the stylets in the search for phloem and sieve tubes. When the saliva contains pathogens, these may be introduced into the phloem with the saliva. Thus, they circulate all the way from the gut to the salivary glands, passing through the whole body of the insect. This is why these pathogens are sometimes called "circulative" agents; they are also termed "propagative" because they multiply in the insect (see Table 11). The time required for the insect to acquire the pathogen while feeding on the infected plants - acquisition feeding- is the acquisition period. This period is fairly long: 24 to 48 hours for Neoaliturus haematoceps feeding on S. citri-infected periwinkle plants (Catharanthus roseus syn. Vinca rosea). The time required for the agent to multiply within the insect and arrive in the salivary glands is the incubation period. This is the period required for the insect to become viruliferous, i.e. able to transmit the disease (eight to 12 days in the case of IV. haematoceps). The retention period is the time during which the insect remains viruliferous. With the phloem restricted pathogens of citrus, the retention period is the remaining lifetime of the insects.
CTV behaves as a semi-persistent agent in relation to its aphid vectors. Citrus mosaic virus of India is a non-persistent or stylet-borne agent. CVEV, a luteovirus, is a persistent agent. The virus circulates in the insect vector but, unlike the spiroplasmas, MLOs and BLOB, there is no multiplication of the virus in the insect. The incubation period is long, since the circulative virus has to reach the salivary glands of the insect before it can be transmitted. The retention period is about two or three weeks. Table 11 lists some of the characteristic relationships of persistent, semi-persistent and non-persistent agents with their insect vectors. The term "stylet-borne" indicates that the pathogen adheres to the stylets after they have been introduced into and then removed from the infected plant. Stylet-borne pathogens can be transmitted almost immediately to a healthy plant, an action that is accomplished in a minute or so. In serial transfers, usually only the first plant is infected, after which the insect loses its infectivity; hence, the pathogen is termed non-persistent. It does not multiply or circulate in the insect. Semi-persistent agents resemble circulative ones in that they are not generally acquired during brief acquisition probes. The probability of transmission increases with longer acquisition-access feeding periods of 12 to 24 hours or more' suggesting that these pathogens are somehow able to accumulate in the vector's anterior alimentary canal and resist being quickly flushed through with clean sap from healthy plants. The pathogen is not present in the insect's haemolymph. Furthermore, experimental injection of the agent directly into the haemolymph does not render the recipient insect viruliferous. With persistent circulative agents. such injections render the insect inoculative, but only after an adequate incubation period. For instance, when S. citri is injected into the leafhopper vectors Neoaliturus haematoceps or N. tenellus, the minimum incubation period is eight days.
TABLE 11 Relationships between citrus disease agents and homopterous insect vectors
It is often desirable to transmit citrus pathogens to herbaceous, non-rutaceous hosts. This can be done with certain pathogens by mechanical transmission (see Table 9). In the case of S. citri, infected leafhopper vectors are able to transmit the spiroplasma to a whole range of herbaceous, non-rutaceous host plants. This is because the leafhoppers are not restricted to rutaceous hosts. However, the psyllid vectors of the greening BLO are strictly restricted to rutaceous hosts, and attempts to vector-transmit the BLO to herbaceous plants have failed. In the case of the witches' broom MLO, the natural vector of the agent is not known, and insect transmission with known leafhopper species has not yet been achieved. Fortunately, the use of the parasitic plant dodder, Cuscuta campestris, has made it possible to transmit the following agents to periwinkle seedlings (Fig. 219): S. citri (Fig. 175), the witches' broom MLO (Fig. 142) and several serotypes of the greening BLO (Fig. 220). However, when using dodder, one must remember that certain viruses pass through its seed.
No pathogens of citrus are known to be naturally transmitted through nematodes or pollen. Pollen from cristacortis graft-infected Orlando tangelo seedling trees, when placed under a strip of bark on the cambial surface of the wood of healthy Orlando tangelo seedlings, was able to transmit the cristacortis agent. indeed, stem pitting and psorosis young leaf symptoms typical of cristacortis developed on five of the six pollen-inoculated seedlings. However, no natural infection of citrus by the cristacortis agent has ever been observed and, therefore, pollen is not a natural means of cristacortis dissemination. Some viruses, such as psorosis, can be maintained in callus tissues grown from infected citrus shoot internodes. Virus transmission can be achieved by placing callus pieces under the bark of healthy plants.
{kind=link}