![]()
![]()
![]()
par
C.E. Harwood1, M.W. Haines2 & E.R. Williams1
1CSIRO Division of Forestry, P.O. Box 4008 QVT,
Canberra ACT 2600, Australie
2Conservation Commission of the Northern Territory
P.O. Box 496, Palmerston 0831, Australie
RESUME
On a étudié la croissance initiale d'un verger à graines de familles d'Acacia crassicarpa, établi à l'origine comme essai de provenances et de descendances avec 170 familles issues de pollinisation libre de provenances naturelles de Papouasie-Nouvelle-Guinée (PNG) et du Queensland. Quinze mois après la plantation, la hauteur moyenne des familles de PNG était de 5,24 m, la hauteur de fût jusqu'à la première fourche de 2,02 m et le diamètre à hauteur d'homme de 5,10 cm. Les moyennes correspondantes pour les familles du Queensland étaient de 3,33 m, 1,60 m et 2,89 cm. La variation entre provenances locales de PNG est faible: toutes les familles de PNG provenaient d'une zone géographique peu étendue et relativement uniforme que l'on peut considérer comme une seule région de provenance. Il y a en revanche des différences statistiquement significatives entre familles à l'intérieur de provenances locales de PNG. Les résultats obtenus sont en accord avec les études précédentes de provenances de l'espèce. En raison de leurs performances médiocres, on peut s'attendre à voir éliminer toutes les familles du Queensland avant que la floraison ne commence.
INTRODUCTION
Les meilleures provenances naturelles de plusieurs espèces d'acacias tropicaux à croissance rapide se situent dans des zones écartées du nord de l'Australie, de la Papouasie-Nouvelle-Guinée et de l'Indonésie, ce qui rend la récolte de graines de populations naturelles difficile et coûteuse (Gunn et Midgley, 1991). Des peuplements artificiels représentant une base génétique large d'une ou plusieurs provenances naturelles supérieures offrent par conséquent une solution intéressante pour la production de semences de bonne qualité génétique à faible coût. L'éclaircie sélective de ces peuplements pourra permettre une amélioration génétique pour certains caractères économiquement importants tels que vigueur et forme de l'arbre (Zobel et Talbert, 1984). Les semences obtenues pourront être utilisées directement pour des reboisements, ou fournir tout ou partie de la base génétique en vue de programmes d'amélioration ailleurs.
Constatant la demande internationale croissante de semences de haute qualité de certains acacias et eucalyptus tropicaux, le Centre australien de semences forestières de la CSIRO (ATSC) a depuis 1988 établi plus de 30 hectares de peuplements semenciers dans le nord de l'Australie, en collaboration avec le Service forestier du Queensland, la Commission de conservation du Territoire du Nord et la firme Melville Forest Products Ltd. (Harwood et al., 1993). Les essences suivantes y ont été plantées: Acacia aulacocarpa, A. auriculiformis, A. crassicarpa, A. mangium, Eucalyptus pellita, E. urophylla, Casuarina junghuhniana. Dans le cadre de ce programme, une plantation de 2,1 ha d'Acacia crassicarpa a été mise en place dans l'île Melville au nord de Darwin, dans le Territoire du Nord.
Le principal objet de cette plantation était la production commode et relativement peu coûteuse de quantités importantes de semences de bonne qualité génétique à partir d'une large base génétique des meilleures provenances de l'espèce. Cette plantation est désignée ici comme verger à graines de familles, parce que, bien qu'elle soit constituée de descendances de familles non sélectionnées issues de pollinisation libre dans des peuplements naturels, on a noté l'ascendance des familles (cf Snyder, 1972) et que, comme nous le verrons ci-dessous, des éclaircies fortes ont été pratiquées à l'intérieur des familles et que certaines provenances et familles inférieures seront éliminées lors d'éclairicies ultérieures.
Des études d'isozymes ont montré qu A. crassicarpa se reproduit essentiellement par allofécondation dans les peuplements naturels (Moran et al., 1989).
MATERIELS ET METHODES
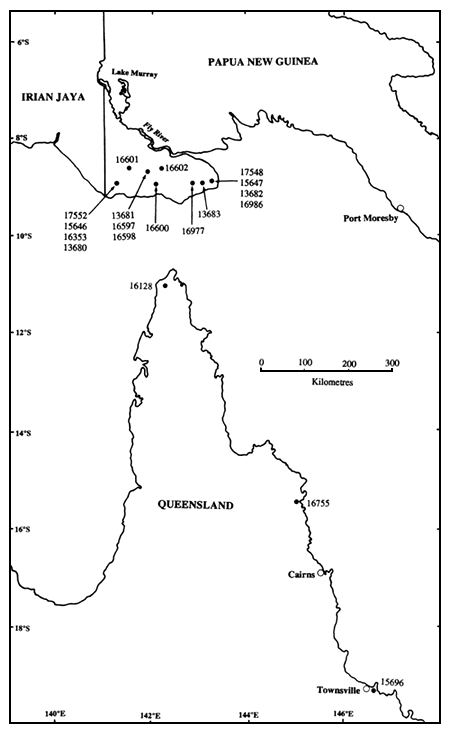
La Figure 1 montre la localisation géographique des lots de semences CSIRO ayant servi à la création du verger à graines. Chaque numéro représente plusieurs lots d'arbres uniques récoltés dans une provenance locale (définie arbitrairement ici comme une surface d'environ 5 × 5 km en vue de discussion ultérieure - voir Turnbull et Griffin, 1986). Les lots de semences d'arbres individuels, désignés ici sous le terme de familles, ont été récoltés sur des arbres éloignés d'au moins 100 mètres les uns des autres afin de limiter la parenté entre familles (les arbres faisaient partie de peuplements d'A. crassicarpa, et n'étaient donc pas des pieds isolés). Le Tableau 1 donne la liste des lots de semences CSIRO, et des familles par lot, inclus dans la plantation. Ces lots de semences couvrent la plus grande partie de l'aire naturelle de l'espèce, à l'exception de peuplements d'Irian Jaya (Gunn et Midgley, 1991). Les trois lots de semences CSIRO du Queensland sont séparés l'un de l'autre de plusieurs centaines de kilomètres. Dans la partie méridionale de la Province Occidentale de Papouasie-Nouvelle-Guinée, l'espèce est largement répartie, et la principale contrainte de la stratégie d'échantillonnage est la difficulté de déplacement dans un pays sans routes. On verra qu'un certain nombre de lots de semences CSIRO de PNG viennent de la même provenance locale (par exemple Oriomo, Gubam-Bimadebun), ou de localités contiguës.
Les plants ont été élevés dans des sachets plastique, contenant 200 cm3 de terre composée d'un mélange de sable, de limon et de tourbe additionné d'un engrais à action lente. Ils ont été inoculés en pépinière avec une souche de Rhizobium appropriée à l'espèce.
Le site de plantation de Yapilika dans l'île Melville (11°34'S, 130°34'E, alt. 20 m) reçoit 1750 mm de précipitations annuelles moyennes avec une saison sèche de 4 mois et une température moyenne annuelle de 24°C. Le sol est un sable fin infertile dérivé de latérite, avec un pH de 5 environ. Le terrain a été labouré avant la plantation, et chaque plant a reçu 229 g d'engrais complet (6:12:8 NPK, plus zinc et cuivre, mis au point pour les reboisements de Pinus caribaea), appliqué manuellement peu après la mise en place.
La plantation, qui comprend 24 plants de chacune des 170 familles, comporte six répétitions complètes de 170 parcelles, chaque famille étant représentée dans chaque répétition par une parcelle linéaire de 4 arbres. Les six répétitions sont contiguës, et les parcelles sont réparties selon un dispositif alpha avec des caractéristiques rang-colonne (Williams et Matheson, 1993), en utilisant le progiciel de création de dispositif “ALPHA + ” élaboré par E.R. Williams. La précision des comparaisons entre différents lots de semences CSIRO a été portée au maximum entre rangs et colonnes en utilisant une option d'ALPHA +. L'espacement est de 3 mètres entre les rangs et 1,5 mètre sur les rangs, ce qui donne une taille de placette de 3 × 6 m. Deux rangs périphériques ont été établis avec des plants en surplus au même espacement tout autour du peuplement pour tenir lieu de bande tampon.
La croissance a été évaluée en mai 1993, 15 mois après la plantation. La hauteur totale et la hauteur de fût jusqu'à la première fourche ont été mesurées avec des perches graduées. On considère qu'il y a une fourche lorsqu'une grosse branche a un diamètre supérieur à la moitié de celui de la tige principale. Le diamètre à hauteur d'homme sur écorce était mesuré avec des gabarits gradués en centimètres. Lorsque le plant est fourchu plus bas que la hauteur d'homme (1,30 m), on mesure les diamètres d1 et d2 des deux tiges, et on calcule un diamètre équivalent de tige unique d à l'aide de la formule suivante:
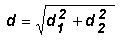
Tableau 1. Lots de semences CSIRO et familles d'Acacia crassicarpa inclus dans le verger à graines, et hauteur moyenne, hauteur de fût et diamètre à hauteur d'homme pour chaque lot de semences à 16 mois
| Lot CSIRO | Localité | Lat. S | Long. E | Alt. (m) | Nb. fam. | h (m) | hf (m) | d (cm) |
| Papouasie-Nouvelle-Guinée | ||||||||
| 17552 | Bensbach | 8°53' | 141°17' | 25 | 16 | 5.19 | 1.99 | 5.13 |
| 15646 | Wemenever | 8°56' | 141°17' | 20 | 8 | 5.38 | 1.96 | 5.26 |
| 16353 | N.E. of Balamuk | 8°49' | 141°20' | 20 | 2 | 5.38 | 1.69 | 5.42 |
| 13680 | Wemenever | 8°51' | 141°26' | 30 | 15 | 5.27 | 1.84 | 5.23 |
| 16601 | Konifok-Serisa | 8°34' | 141°27' | 25 | 2 | 5.15 | 1.75 | 4.96 |
| 13681 | Mata | 8°40' | 141°45' | 30 | 8 | 5.14 | 1.75 | 4.96 |
| 16597 | Gubam-Bimadebun | 8°37' | 141°55' | 25 | 21 | 5.27 | 2.10 | 5.12 |
| 16598 | Gubam-Bimadebun | 8°37' | 141°55' | 25 | 11 | 5.28 | 2.02 | 5.23 |
| 16600 | Mai Kussa | 8°52' | 142°03' | 15 | 3 | 4.97 | 1.98 | 4.81 |
| 16602 | Dimisisi | 8°31' | 142°13' | 50 | 7 | 5.07 | 1.65 | 4.96 |
| 16977 | Wipim | 8°49' | 142°48' | 45 | 18 | 5.24 | 2.18 | 4.96 |
| 13683 | Woroi-Wipim | 8°49' | 143°00' | 20 | 15 | 5.25 | 2.28 | 5.04 |
| 17548 | Oriomo Old Zip | 8°48' | 143°06' | 20 | 4 | 5.29 | 2.36 | 4.85 |
| 15647 | Oriomo | 8°50' | 143°08' | 10 | 2 | 5.10 | 1.87 | 5.17 |
| 13682 | Oriomo | 8°50' | 143°10' | 20 | 11 | 5.30 | 1.87 | 5.16 |
| 16986 | Oriomo DPI | 8°51' | 143°11' | 35 | 7 | 5.31 | 2.17 | 5.11 |
| Moyenne de 150 familles de PNG | 5.24 | 2.02 | 5.10 | |||||
| Queensland | ||||||||
| 15696 | Townsville | 19°23' | 147°01' | 15 | 6 | 2.80 | 1.57 | 1.93 |
| 16128 | Jardine R | 11°02' | 142°22' | 20 | 11 | 3.46 | 1.60 | 3.27 |
| 16755 | Parish of Annan | 15°36' | 145°19' | 80 | 3 | 3.95 | 1.66 | 3.49 |
| Moyenne de 20 familles du Queensland | 3.33 | 1.60 | 2.89 | |||||
Nb. fam. = nombre de familles par lot de semences CSIRO
h = hauteur totale
hf = hauteur de fût
d = diamètre à hauteur d'homme sur écorce
Figure 1. Localisation géographique des lots de semences CSIRO inclus dans le verger à graines
Très peu de plants avaient plus de deux tiges à hauteur d'homme; seuls les deux tiges les plus grosses étaient mesurées dans ce cas. Les moyennes des trois caractères retenus ont été calculées pour l'ensemble des parcelles, et une analyse de variance de ces moyennes a été effectuée en utilisant le progiciel statistique GENSTAT 5.2. Une analyse initiale a été faite en utilisant un modèle linéaire dans lequel les répétitions, les régions (PNG et Queensland), les lots de semences CSIRO à l'intérieur de ces régions, et les familles à l'intérieur des lots de semences, étaient estimés en tant qu'effets fixes. Une autre analyse a ensuite été effectuée sur les familles de la région PNG. Elle excluait tous les lots de semences du Queensland, et utilisait un modèle linéaire analogue mais en omettant le terme de région.
RESULTATS ET DISCUSSION
On a enregistré une croissance initiale rapide d'A. crassicarpa, excédant celle d'A. auriculiformis et A. mangium plantés dans des essais voisins à la même époque. Le taux de survie moyen était de 94,4 pour cent pour l'ensemble de la plantation, et excédait 92 pour cent tous les lots de semences CSIRO. Les moyennes de hauteur totale, hauteur de fût et diamètre à hauteur d'homme sont présentées dans le Tableau 1. Tous les lots de semences de PNG ont poussé beaucoup plus vite que les trois du Queensland (P < 0,001). Un meilleur comportement des provenances de PNG par rapport à celles du Queensland a également été rapporté dans des essais d'espèces et provenances en Thaïlande (Chittachumnonk et Sirilak, 1991) et dans l'île de Haïnan en Chine (Yang et Zeng, 1991).
La variation entre lots de semences de PNG n'était pas significative pour la hauteur totale, mais elle l'était (P < 0,05) pour la hauteur de fût et le diamètre (Tableau 2). Les différences entre lots de semences étaient bien moins importantes que les différences entre régions. On n'a pas décelé de tendance géographique très nette dans la variation entre lots de semences de PNG; on trouve des lots au-dessus et au-dessus de la moyenne dans les parties est, centrale et ouest de la zone de récolte (Tableau 1). Cette constatation est en accord avec les résultats des essais cités ci-dessus, et avec ceux de Sim et Gan (1991) au Sabah, dans lesquels les différences entre provenances locales de PNG sont faibles. Les lots de semences de PNG testés à ce jour proviennent tous d'une zone relativement restreinte, peu accidentée, d'environ 200 × 50 km (Figure 1, Gunn et Midgley, 1991), de sorte que l'absence de variation entre lots de semences n'est pas surprenante. L'aire d'A. crassicarpa en Papouasie-Nouvelle-Guinée et dans les régions voisines d'Irian Jaya peut être considérée comme une seule région de provenance (Turnbull et Griffin, 1986), bien qu'elle ait été subdivisée par Harwood et al. (1993) en régions de provenance plus petites pour la gestion des ressources génétiques des autres espèces d'Acacia et d'Eucalyptus pellita. La principale tendance écologique susceptible d'exercer une pression de sélection entraînant une différenciation des populations dans cette région est la longueur décroissante de la saison sèche et l'accroissement de la pluviométrie et de l'humidité relative du sud au nord (Vercoe et McDonald, 1991).
Le verger à graines a fait l'objet d'une éclaircie sélective après cette évaluation, en maintenant le meilleur sujet dans chaque parcelle linéaire de quatre arbres, choisi visuellement pour sa forme (en donnant la préférence aux sujets à tige simple rectiligne) et sa vigueur (l'arbre montrant la meilleure croissance était conservé s'il avait un net avantage de taille sur l'arbre le mieux formé). Cette éclaircie a réduit la densité du peuplement à environ 550 tiges/ha, et elle avait pour objet de favoriser le développement de cimes larges et denses pour une production maximale de graines. Elle assure aussi que les arbres voisins conservés ne soient pas parents, ce qui réduit la consanguinité dans le verger à graines. Dans l'hypothèse où une deuxième évaluation confirmerait la moins bonne croissance des provenances du Queensland, tous les arbres du Queensland (20 familles) seront éliminés du verger à graines lors d'une deuxième éclaircie, avant que la floraison ne commence. On escompte qu'une production appréciable de semences débutera en 1995. Lors d'une nouvelle éclaircie visant à réduire la densité du peuplement au niveau désiré de 150 tiges/ha en vue d'une production normale de semences, on éliminera en tout ou partie les familles de PNG inférieures. A ce stade il apparaît improbable qu'aucune des “provenances” locales de PNG (comprenant jusqu'à quatre lots de semences) soit éliminée complètement, étant donné que dans toutes on trouve une ou plusieurs “bonnes” familles ayant une hauteur, un diamètre ou une longueur de fût supérieure à la moyenne générale de toutes les familles de PNG. La variation entre familles à l'intérieur des lots de semences est hautement significative pour tous les caractères.
Tableau 2. Tableaux résumés d'analyse de variance pour la hauteur totale, la hauteur de fût et le diamètre (les lots de semences du Queensland sont exclus de l'analyse)
| (a) Hauteur totale | ||||||
| dl | sc | mc | F | P | ||
| Répétitions | 5 | 6.9426 | 1.3885 | 5.61 | ||
| Répétitions unités | ||||||
| Lots de semences CSIRO | 15 | 5.7081 | 0.3805 | 1.54 | 0.086 | |
| Familles dans les lots | 134 | 184.1608 | 0.5036 | 2.03 | <.001 | |
| Résiduelle | 744 | (1) | 184.1608 | 0.2475 | ||
| Total | 898 | (1) | 264.2462 | |||
| (b) Hauteur de fût | ||||||
| dl | sc | mc | F | P | ||
| Répétitions | 5 | 10.432 | 2.086 | 1.88 | ||
| Répétitions unités | ||||||
| Lots de semences CSIRO | 15 | 29.542 | 1.969 | 1.78 | 0.034 | |
| Familles dans les lots | 134 | 206.534 | 1.541 | 1.39 | 0.005 | |
| Résiduelle | 744 | (1) | 825.473 | 1.110 | ||
| Total | 898 | (1) | 1071.941 | |||
| (c) Diamètre (1,30m) | ||||||
| dl | sc | mc | F | P | ||
| Répétitions | 5 | 14.3765 | 2.875 | 6.45 | ||
| Répétitions unités | ||||||
| Lots de semences CSIRO | 15 | 12.7875 | 0.8535 | 1.91 | 0.019 | |
| Familles dans les lots | 134 | 107.0839 | 0.799 | 1.79 | <0.001 | |
| Résiduelle | 744 | (1) | 331.8379 | 0.446 | ||
| Total | 898 | (1) | 465.7881 | |||
dl = degré de liberté
sc = somme des carrés
mc = moyenne des carrés
F = F calculé
P = probabilité que ce ratio F se produise par hasard
(1) = une parcelle sans arbre → valeur manquante
Le dispositif et la conduite de ce verger à graines de familles illustrent divers points d'ordre général concernant la planification de vergers à graines de première génération d'essences sur les provenances desquelles on dispose de peu d'information (Nanson, 1972). L'examen de l'information disponible avant la plantation avait indiqué que le comportement des provenances du Queensland était nettement moins bon que celui des provenances de PNG. L'inclusion d'un petit nombre de familles (20 sur un total de 170) de trois provenances du Queensland distantes l'une de l'autre a permis de confirmer cette présomption pour l'île Melville sans compromettre le rôle essentiel du peuplement comme verger à graines des meilleures provenances: les familles du Queensland peuvent être totalement éliminées sans créer de larges trouées dans le verger. On ne s'attendait pas à voir de grandes différences dans la croissance et la phénologie de la floraison entre les “provenances” locales d'A. crassicarpa de PNG, c'est pourquoi on a considéré comme faisable un verger à graines représentant toutes les provenances de PNG. Toutes les familles de PNG disponibles ont été incluses pour avoir un maximum de représentation de matériel génétique de PNG, plutôt que de limiter le nombre de familles utilisées par “provenance” locale à un nombre arbitraire tel que 10 ou 20. Les ressources disponibles ne permettaient pas la plantation de peuplements séparés représentant des subdivisions du matériel génétique de PNG, qui auraient été utiles pour l'exploitation de la coascendance dans les générations suivantes. Cependant, il serait possible de reconstituer des lignées secondaires génétiquement distinctes par multiplication végétative de sujets d'élite du verger à graines, en utilisant les techniques de propagation mises au point pour A. auriculiformis et A. mangium (Carron et Aken, 1992). Des analyses statistiques de modèles mixtes dans lesquels les rangs et les colonnes ont été estimés comme effets aléatoires en utilisant l'option REML de GENSTAT 5.2 (Williams et Matheson, 1993) ont nettement réduit le carré moyen résiduel relatif au modèle de bloc aléatoire complet considéré ici; elles seront présentées dans une publication ultérieure.
REMERCIEMENTS
Le programme de production de semences de l'ATSC a été rendu possible par un financement du Service australien d'assistance au développement international. L'appui fourni par la Division des ressources forestières de la FAO pour les opérations de l'ATSC a contribué au rassemblement des ressources génétiques utilisées. La société Melville Forest Products Ltd a mis à disposition un terrain et des équipements pour les essais. Nous remercions les membres de la Conservation Commission du Territoire du Nord, en particulier Beau Robertson, pour leur contribution à toutes les étapes du projet. Les lots de semences de PNG utilisés pour la plantation ont été récoltés en collaboration avec le Département des forêts de Papouasie-Nouvelle-Guinée, qui a également établi localement des peuplements semenciers de l'espèce. Peter Dart, du Département d'agriculture de l'Université du Queensland, a fourni l'inoculum de Rhizobium. Nous remercions enfin Ken Eldridge pour ses utiles commentaires sur une première rédaction de cet article.
Références
Carron, L.T. & K.M. Aken, eds. (1992). Breeding technologies for tropical acacias. ACIAR Proceedings No 371992
Chittachumnok, P. & S. Sirilak (1991). Performance of Acacia species in Thailand. pp 153–158 in Turnbull, J.W. ed. Advances in Tropical Acacia Research. ACIAR Proceedings No. 35.
Gunn, B.V. & S.J. Midgley (1991). Exploring and accessing the genetic resources of four selected tropical acacias. pp. 57–63 in Turnbull, J.W. ed. Advances in Tropical Acacia Research. ACIAR Proceedings No. 35.
Harwood, C.E., G.B. Applegate, K.J. Robson & M.W. Haines (1993). Establishment and management of seed production areas of tropical tree species in northern Australia. Paper presented to ASEAN-Canada Forest Tree Seed Centre international conference on genetic conservation and production of tropical tree seed, Chiang Mai, June 14–16 1993. (in press)
Moran, G.F., O. Muona & J.C. Bell. Breeding systems and genetic diversity in Acacia auriculiformis and A. crassicarpa. Biotropica 21, 250–256.
Nanson (1972). The provenance seedling seed orchard. Silvae Genetica 21, 243–249.
Sim, B.L. & E. Gan (1991). Performance of Acacia species on four sites of Sabah Forest Industries. pp. 159–165 in Turnbull, J.W. ed. Advances in Tropical Acacia Research. ACIAR Proceedings No. 35.
Snyder, E.B. (1972). Glossary for forest tree improvement workers. Southern Forest Experiment Station, United States Department of Agriculture.
Turnbull, J.W. & A.R. Griffin (1986). The concept of provenance and its relationship to infraspecific classification in forest trees. pp. 157–189 in B.T. Styles ed. Infraspecific Classification of Wild and Cultivated Plants. Clarendon Press, Oxford.
Vercoe, T.K. & M.W. McDonald (1991). E. pellita F. Muell. and Acacia seed collection in New Guinea, September–October 1990. FAO Forest Genetic Resources Information 19, 38–42.
Williams, E.R. & A.C. Matheson (1993). Design and analysis of forestry field trials. ACIAR/CSIRO, Australia. (in press).
Yang Minquan & Zeng Yutian (1991). Results from a four-year-old tropical Acacia species/provenance trial on Hainan Island, China. pp. 170–172 in Turnbull, J.W. ed. Advances in Tropical Acacia Research. ACIAR Proceedings No. 35.
Zobel, B.J. & J.T. Talbert (1984). Applied Forest Tree Improvement. John Wiley and Sons, New York.
Informations sur les Resources Génétiques Forestières No 21. FAO, Rome (1994)
![]()
![]()
![]()