![]()
![]()
![]()
por
C.E. Harwood1; M.W. Haines2 & E.R. Williams1
1. CSIRO Division of Forestry, PO Box 4008 QVT
Canberra ACT 2600, Australia
2. Conservation Commission of the Northern Territory
PO Box 496, Palmerston 0831, Australia
RESUMEN
El artículo informa sobre el crecimiento precoz de un huerto semillero de brinzales de Acacia crassicarpa, diseñado inicialmente como un ensayo de procedencias y progenies de 170 familias de polinización abierta originarias de procedencias naturales de Papua Nueva Guinea (PNG) y Queensland. Quince meses después de la plantación, la altura media de las familias de PNG era de 5,24 metros, la longitud de tronco hasta la primera bifurcación 2,02 m. y el diámetro normal 5,10 cm. Las medias correspondientes a las familias de Queensland eran 3,33, 1,60 y 2,89. La variación entre procedencias locales de PNG era pequeña: todas las familias de PNG procedían de un área geográfica pequeña y relativamente uniforme que puede considerarse como una sola región de procedencia. Había, sin embargo, diferencias estadísticamente significativas entre familias dentro de las procedencias locales de PNG. Los resultados obtenidos estuvieron de acuerdo con los estudios anteriores de procedencias de la especie. Debido a su mal rendimiento, se prevé que serán eliminadas del huerto todas las familias de Queensland, antes de comenzar la floración.
INTRODUCCION
Las procedencias naturales de mejor comportamiento de varias especies tropicales de acacia de crecimiento rápido corresponden a zonas remotas del norte de Australia, Papua Nueva Guinea (PNG) e Indonesia, lo que hace difícil y costosa la recolección de semilla de las poblaciones naturales (Gunn y Midgley, 1991). Los rodales plantados que representan una extensa base genética de una o más procedencias naturales superiores, proporcionan por tanto una oportunidad atractiva para la producción a bajo costo de semilla de buena calidad genética. El aclareo selectivo de tales rodales puede permitir el mejoramiento genético de caracteres económicos seleccionados como el vigor y la forma del árbol (Zobel y Talbert, 1984). La semilla producida puede emplearse directamente para plantaciones operativas o puede servir como base genética parcial o total para programas de mejoramiento de árboles en otros lugares.
Teniendo en cuenta la creciente demanda internacional de semilla de buena calidad de ciertas acacias y eucaliptos tropicales, el Centro de Semillas de Arboles de Australia (ATSC) de CSIRO ha establecido desde 1988 más de 30 ha. de producción de semilla en el norte de Australia, en colaboración con el Servicio Forestal de Queensland, la Comisión de Conservación del Territorio Norte y la Melville Forest Products Ltd (Harwood et al., 1993). Las especies plantadas incluyen Acacia aulacocarpa, A. auriculiformis, A. crassicarpa, A. mangium, Eucalyptus pellita, E. urophylla y Casuarina junghuhniana. Como parte de este programa se estableció una plantación de 2,1 ha. de Acacia crassicarpa en la Isla de Melville, al norte de Darwin, en el territorio norte de Australia.
La primera finalidad de la plantación fue la producción práctica y relativamente económica de cantidades sustanciales de semilla de buena calidad genética a partir de unas bases genéticas extensas de las mejores procedencias de cada especie. En adelante, se hará mención a la plantación como huerto semillero de brinzales (HSB), porque aunque el rodal está compuesto de progenies de familias no seleccionadas de polinización abierta, procedentes de masas naturales, se han mantenido los pedigrees de la familia (cf. Snyder, 1972), y, como se analiza más adelante, se han realizado claras extensas dentro de las familias y en las claras subsiguientes se van a eliminar algunas procedencias y familias inferiores.
Los estudios con isoenzimas han demostrado que la A. crassicarpa se hibrida, mediante cruzamiento lejano, en las masas naturales (Moran et al., 1989).
MATERIALES Y METODOS
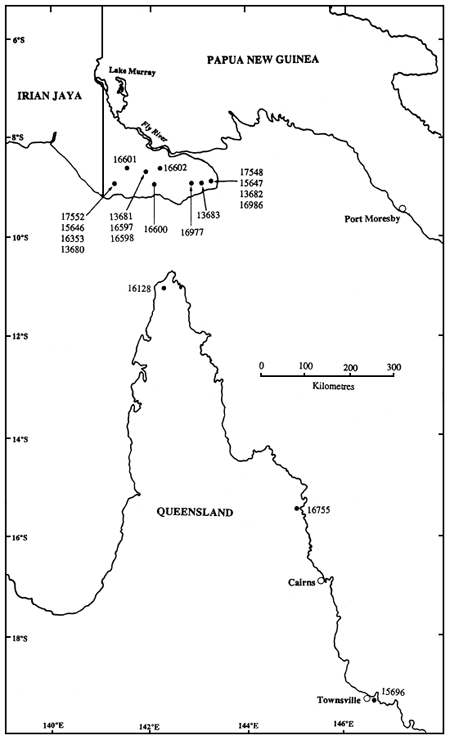
En la Figura 1 se muestran las localidades geográficas de los lotes de semilla de CSIRO que contribuyen al HSB. Cada número de lote de semillas de CSIRO representa varios lotes de semilla de un solo árbol recogidas dentro de una procedencia local (definida aquí arbitrariamente como una superficie hasta de 5 × 5 Km. al objeto de la discusión posterior -véase Turnbull y Griffin, 1986). Se recogieron lotes de semilla de un solo árbol, denominados en adelante familias, a partir de árboles situados entre sí al menos a 100 m. de distancia a fin reducir al mínimo la afinidad entre familias (los árboles estaban en rodales de A. crassicarpa, es decir, que no eran individuos aislados). El Cuadro 1 enumera los lotes de semilla de CSIRO, y las familias por lote, incluidos en la plantación. Los lotes de semilla abarcaban la mayor parte del ámbito natural de la especie, excepto para las existencias de Irian Jaya (Gunn y Midgley, 1991). Los tres lotes de semilla CSIRO procedentes de Queensland están separados entre sí por varios centenares de kilómetros. En la parte meridional de la provincia occidental, PNG, la especie tiene una amplia distribución y la estrategia de muestreo está limitada sobre todo por la logística de los viajes en un país sin carreteras. Como se puede ver, un cierto número de lotes de semilla de CSIRO procedentes de la muestra PNG son o de la misma procedencia local (p. ej. Oriomo, Gubam-Bimadebun), o de localidades geográficas inmediatamente adyacentes.
Se produjeron las plantas en envases de plástico de tipo libro, con un volumen de tierra de 200 cm3, utilizando una mezcla de tierra con arena, limo y turba a la que se incorporó un fertilizante de liberación lenta. Se inocularon en el vivero con una raza de rizhobium apropiada para la especie.
La estación de plantación de Yapilika, en la lsla de Melville (11°34 S, 130°34 E, 20 m.s.n.m.), tiene una precipitación anual media de unos 1.750 mm. con cuatro meses de estación seca y una temperatura media anual de 24°C. El tipo de suelo es una arena fina, poco fértil, procedente de lateritas, con un pH próximo a 5. Se labró el sitio antes de plantarlo y cada planta recibió 229 g. de fertilizante completo (6:12:8 NPK, más zinc y cobre), desarrollado para plantaciones operativas de Pinus caribaea, aplicado a mano poco después de la plantación.
La plantación, que incluye 24 brinzales de cada una de las 170 familias, empleó seis repeticiones completas de 170 parcelas, estando representada cada familia en cada repetición por una parcela lineal de cuatro árboles. Las seis repeticiones estaban contiguas y las parcelas se dispusieron con un diseño alfa con propiedades fila-columna (Williams y Matheson, 1993), utilizando el paquete de diseño “ALPHA +” preparado por E.R. Williams. La precisión de las comparaciones entre los distintos lotes de semilla CSIRO se hizo máxima dentro de filas y columnas utilizando una opción en ALPHA +. El espaciamiento era de 3 metros entre filas y 1,5 m. entre árboles dentro de las filas, con un tamaño de parcela de 3 × 6 m. Alrededor del rodal se plantaron dos hileras perimetrales de material sobrante con el mismo espaciamiento, con fines de amortiguación.
Se evaluó el crecimiento en mayo de 1993, 15 meses después de la plantación. Se midió con jalones largos la altura y la longitud del tronco hasta la primera ramificación importante. Se consideraba que existía ramificación cuando una rama principal tenía más de la mitad del diámetro del tronco principal. Con una forcípula graduada en centímetros se midió el diámetro normal a la altura del pecho (dbn). Cuando las plantas se ramificaban por debajo de la altura del pecho, se medían los diámetros dbh1 y dbh2 de ambas ramas y se calculaba el diámetro de un solo tronco, d, de la forma siguiente:
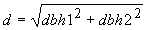
Cuadro 1. Lotes de semilla CSIRO y familias de Acacia crassicarpa incluidos en el huerto semillero y altura media, longitud del tronco y diámetro normal de los lotes de semilla CSIRO a los 16 meses
| Lote de semilla CSIRO | Estación | Lat. oS | Long. oE | Alt. (m) | No. fams | Ht. (m) | L.T. (m) | Dn (cm) | |
| Papua Nueva Guinea | |||||||||
| 17552 | Bensbach | 8 53 | 141 | 17 | 25 | 16 | 5.19 | 1.99 | 5.13 |
| 15646 | Wemenever | 8 56 | 141 | 17 | 20 | 8 | 5.38 | 1.96 | 5.26 |
| 16353 | N.E. of Balamuk | 8 49 | 141 | 20 | 20 | 2 | 5.38 | 1.69 | 5.42 |
| 13680 | Wemenever | 8 51 | 141 | 26 | 30 | 15 | 5.27 | 1.84 | 5.23 |
| 16601 | Konifok-Serisa | 8 34 | 141 | 27 | 25 | 2 | 5.15 | 1.75 | 4.96 |
| 13681 | Mata | 8 40 | 141 | 45 | 30 | 8 | 5.14 | 1.75 | 4.96 |
| 16597 | Gubam-Bimadebun | 8 37 | 141 | 55 | 25 | 21 | 5.27 | 2.10 | 5.12 |
| 16598 | Gubam-Bimadebun | 8 37 | 141 | 55 | 25 | 11 | 5.28 | 2.02 | 5.23 |
| 16600 | Mai Kussa | 8 52 | 142 | 03 | 15 | 3 | 4.97 | 1.98 | 4.81 |
| 16602 | Dimisisi | 8 31 | 142 | 13 | 50 | 7 | 5.07 | 1.65 | 4.96 |
| 16977 | Wipim | 8 49 | 142 | 48 | 45 | 18 | 5.24 | 2.18 | 4.96 |
| 13683 | Woroi-Wipim | 8 49 | 143 | 00 | 20 | 15 | 5.25 | 2.28 | 5.04 |
| 17548 | Oriomo Old Zip | 8 48 | 143 | 06 | 20 | 4 | 5.29 | 2.36 | 4.85 |
| 15647 | Oriomo | 8 50 | 143 | 08 | 10 | 2 | 5.10 | 1.87 | 5.17 |
| 13682 | Oriomo | 8 50 | 143 | 10 | 20 | 11 | 5.30 | 1.87 | 5.16 |
| 16986 | Oriomo DPI | 8 51 | 143 | 11 | 35 | 7 | 5.31 | 2.17 | 5.11 |
| Media de 150 familias PNG | 5.24 | 2.02 | 5.10 | ||||||
| Queensland | |||||||||
| 15696 | Townsville | 19 23 | 147 | 01 | 15 | 6 | 2.80 | 1.57 | 1.93 |
| 16128 | Jardine R | 11 02 | 142 | 22 | 20 | 11 | 3.46 | 1.60 | 3.27 |
| 16755 | Parish of Annan | 15 36 | 145 | 19 | 80 | 3 | 3.95 | 1.66 | 3.49 |
| Media de 20 familias Queensland | 3.33 | 1.60 | 2.89 | ||||||
| no.fams. | = | número de familias por lote de semillas CSIRO |
| Ht. | = | altura |
| L.T. | = | longitud del tronco |
| Dn | = | diámetro normal (a la altura del pecho) |
Figura 1. Estaciones geográficas de los lotes de semilla CSIRO incluidos en el huerto semillero
Muy pocas plantas tenían más de dos troncos a la altura del pecho; en ellas sólo se midieron los dos troncos mayores. Se calcularon las medias por parcela de las tres características y se llevó a cabo un análisis de varianza sobre los datos medios de las parcelas utilizando el paquete estadístico GENSTAT 5.2. Un primer análisis empleó un modelo lineal en el que se estimaron como efectos fijos las repeticiones, las regiones (PNG y Queensland), los lotes de semilla CSIRO dentro de las regiones y las familias dentro de los lotes de semilla CSIRO. Se realizó a continuación un análisis sobre las familias procedentes de la región PNG. Este excluyó todos los lotes de semilla de Queensland y utilizó un modelo lineal similar pero omitiendo el término región.
RESULTADOS Y DISCUSION
Se registró un rápido crecimiento precoz del A. crassicarpa, superior a los de la A. auriculiformis y la A. mangium plantados al mismo tiempo en ensayos adyacentes. La supervivencia fue como promedio del 94,4 por ciento para toda la plantación y sobrepasó el 92 por ciento para todos los lotes de semilla CSIRO. Las medias de los lotes de semilla CSIRO en cuanto a altura, longitud de tronco y dn se presentan en el Cuadro 1. Todos los lotes de semilla PNG crecieron mucho más deprisa que cualquiera de los tres procedentes de Queensland (P<.001). También se han registrado en los ensayos de especies y procedencias de Tailandia (Chittachumnonk y Sirilak, 1991) y en la Isla de Hainan, China (Yang y Zang, 1991), mejores rendimientos de las procedencias de PNG, en relación con las procedencias de Queensland.
La variación entre los lotes de semilla PNG no fue significativa en cuanto a la altura pero sí lo fue (P<0,05) para la longitud del tronco y el dn (Cuadro 2). Las diferencias entre lotes de semilla fueron mucho menores que las diferencias entre las dos regiones. No hubo una tendencia geográfica clara en la variación entre los lotes de semilla PNG, existiendo lotes de semilla por encima y por debajo de la media procedentes de las áreas orientales, centrales y occidentales de la zona de recolección (Cuadro 1). Este resultado está de acuerdo con los resultados de los ensayos antes mencionados y los de Sim y Gan (1991) en Sabah, en los cuales fueron pequeñas las diferencias entre las procedencias locales de PNG. Los lotes de semilla PNG ensayados hasta ahora son todos procedentes de un área relativamente pequeña de escaso relieve que tiene unos 200 Km. por 50 Km. de extensión (Figura 1, Gunn y Midgley, 1991), por lo que no es sorprendente la falta de variación entre estos lotes de semilla. El área donde está presente el A. crassicarpa en PNG y el área adyacente de Irian Jaya pueden considerarse como una sola región de procedencia (Turnbull y Griffin, 1986) aunque Hardwood et al. (1993) la han subdividido en regiones más pequeñas de procedencia, para la ordenación de los recursos genéticos de otras especies de acacia y de Eucalyptus pellita. La principal tendencia ambiental que podría esperarse que ejerciese una presión de selección, traducida en la diferenciación de las poblaciones dentro de la región, es la duración decreciente de la estación seca y el aumento de las precipitaciones y de la humedad relativa, de sur a norte (Vercoe y McDonald, 1991).
El HSB se aclaró selectivamente después de la evaluación, manteniendo el mejor árbol en cada parcela lineal de cuatro árboles, elegido a ojo por su forma (rectitud, árboles de un solo tronco) y vigor (se mantiene el árbol que presenta el mejor desarrollo si tiene una ventaja sustancial de tamaño sobre el árbol de mejor forma). Esta clara redujo la densidad del rodal a unos 550 pies por hectárea y estuvo destinada a promover el desarrollo de copas amplias y densas para la máxima producción de semilla. Esta clara garantiza también que los árboles adyacentes que se mantienen no estén emparentados, a fin de reducir la consanguinidad en el HSB. Suponiendo que una segunda evaluación confirme el peor crecimiento de las procedencias de Queensland, todos los árboles de Queensland (20 familias) serán eliminados del HSB en una segunda clara, antes de que comience la floración. Se espera que comience en 1995 una producción sustancial de semilla. Las oportunidades de una nueva clara, para reducir la densidad del rodal hasta un nivel conveniente, de unos 150 pies por hectárea para la producción conveniente de semilla, incluyen la eliminación parcial o total de las familias inferiores de PNG. En esta etapa parece poco probable que se eliminen por completo las procedencias locales de PNG (compuestas hasta de 4 lotes de semilla CSIRO) ya que todas ellas tienen una o más familias “buenas” con valores medios de altura, diámetro y longitud de tronco superiores a la media global de todas las familias PNG. La variación entre familias dentro de los lotes de semillas CSIRO es muy significativa para todos los caracteres.
Cuadro 2. Análisis resumido de los cuadros de varianza en cuanto a la altura, longitud de tronco y Dn (los lotes de semillas de Queensland están excluidos del análisis)
| (a) Altura | ||||||
| gl | sc | mc | F | P | ||
| Estrato de repeticiones Repeticiones. Estrato de unidades | 5 | 6.9426 | 1.3885 | 5.61 | ||
| Lotes de semillas CSIRO | 15 | 5.7081 | 0.3805 | 1.54 | 0.086 | |
| Familias dentro de los lotes de semilla | 134 | 184.1608 | 0.5036 | 2.03 | <.001 | |
| Residual | 744 | (1) | 184.1608 | 0.2475 | ||
| Total | 898 | (1) | 264.2462 | |||
| (b) Longitud | ||||||
| gl | sc | mc | F | P | ||
| Estrato de repeticiones Repeticiones. Estrato de unidades | 5 | 10.432 | 2.086 | 1.88 | ||
| Lotes de semillas CSIRO | 15 | 29.542 | 1.969 | 1.78 | 0.034 | |
| Familias dentro de los lotes de semilla | 134 | 206.534 | 1.541 | 1.39 | 0.005 | |
| Residual | 744 | (1) | 825.473 | 1.110 | ||
| Total | 898 | (1) | 1.071.941 | |||
| (a) Diámetro (Dn) | ||||||
| gl | sc | mc | F | P | ||
| Estrato de repeticiones Repeticiones. Estrato de unidades | 5 | 14.3765 | 2.875 | 6.45 | ||
| Lotes de semillas CSIRO | 15 | 12.7875 | 0.8535 | 1.91 | 0.019 | |
| Familias dentro de los lotes de semilla | 134 | 107.0839 | 0.799 | 1.79 | <0.001 | |
| Residual | 744 | (1) | 331.8379 | 0.446 | ||
| Total | 898 | (1) | 465.7881 | |||
| gl | = | grados de libertad |
| sc | = | suma de cuadrados |
| mc | = | media de cuadrados |
| F | = | relación F |
| P | = | probabilidad de que la relación F se produzca al azar |
(1) = una parcela sin árboles - esto es, falta valor
El diseño y gestión de este HSB aclara algunos aspectos generales de la programación de HSB de primera generación de especies cuya información en cuanto a procedencias es limitada (Nanson, 1972). El análisis de la información disponible antes de la plantación había indicado que el rendimiento de las procedencias de Queensland era bastante inferior al de las procedencias de PNG. La inclusión de un pequeño número (20 de un total de 170) de familias de tres procedencias de Queensland muy separadas, ha permitido confirmar esta suposición para el ambiente de la isla de Melville, sin comprometer el papel primordial del rodal como un HSB de las mejores procedencias. Las familias de Queensland pueden eliminarse totalmente sin crear grandes vacíos físicos en el HSB. No se previeron grandes diferencias de crecimiento y fenología de floración entre las procedencias locales de A. crassicarpa dentro de PNG, de modo que se consideró viable un huerto semillero que representase todas las procedencias de PNG. Se incluyeron todas las familias disponibles de PNG para tener una representación máxima del material genético de PNG, en lugar de limitar las familias utilizadas por “procedencia” local a un número tanto arbitario como 10 ó 20. Los recursos disponibles no permitieron la plantación de rodales independientes que representasen subdivisiones del material genético de PNG, lo que habría tenido interés para la manipulación de ancestros comunes en posteriores generaciones. Sin embargo, sería posible reconstruir sublíneas independientes genéticamente mediante propagación vegetativa de árboles sobresalientes individuales en el HSB, utilizando las técnicas de propagación desarrolladas para la A. auriculiformis y A. mangium (Carron y Aken, 1992). Unos análisis estadísticos de modelo mixto, en los que se calcularon las filas y columnas como efectos al azar, empleando la opción REML de GENSTAT 5.2 (Williams y Matheson, 1993), redujeron sustancialmente la media cuadrática residual en relación con el modelo de bloque completo al azar sobre el que se informa aquí. Tales análisis se van a presentar en una publicación posterior.
RECONOCIMIENTOS
El programa de producción de semillas del CASA ha sido posible mediante la financiación de la Agencia Australiana de Asistencia Internacional al Desarrollo. El apoyo brindado a las operaciones del CASA por la Dirección de Recursos Forestales de la FAO ha contribuido a reunir los recursos genéticos utilizados. La Melville Forests Products Ltd puso tierras y equipos a disposición de estos ensayos. Se agradece la contribución realizada por el personal de la Comisión de Conservación del Territorio Norte, en particular de Beau Robertson, en todas las etapas del proyecto. Los lotes de semillas de PNG empleados en la plantación se recogieron en colaboración con el Departamento de Bosques de Papua Nueva Guinea, que ha establecido también localmente áreas de producción de semillas de la especie. Peter Dart (Departamento de Agricultura de la Universidad de Queensland) proporcionó el inóculo de Rhizobium. Agradecemos a Ken Eldridge sus valiosos comentarios sobre el primer borrador de este documento.
Referencias
Carron, L.T. & Aken, K.M. eds. (1992). Breeding technologies for tropical acacias. ACIAR Proceedings No 37
Chittachumnonk, P. & Sirilak, S. (1991). Performance of Acacia species in Thailand. pp 153–158 in Turnbull, J.W. ed. Advances in Tropical Acacia Research. ACIAR Proceedings No. 35.
Gunn, B.V. & Midgley, S.J. (1991). Exploring and accessing the genetic resources of four selected tropical acacias. pp. 57–63 in Turnbull, J.W. ed. Advances in Tropical Acacia Research. ACIAR Proceedings No. 35.
Harwood, C.E., Applegate, G.B., Robson, K.J. & Haines, M.W. (1993). Establishment and management of seed production areas of tropical tree species in northern Australia. Paper presented to ASEAN-Canada Forest Tree Seed Centre international conference on genetic conservation and production of tropical tree seed, Chiang Mai, June 14–16 1993. (In press)
Moran, G.F., Muona, O. & Bell, J.C. Breeding systems and genetic diversity in Acacia auriculiformis and A. crassicarpa. Biotropica 21, 250–256.
Nanson. (1972). The provenance seedling seed orchard. Silvae Genetica 21, 243–249.
Sim, B.L. & Gan, E. (1991). Performance of Acacia species on four sites of Sabah Forest Industries. pp. 159–165 in Turnbull, J.W. ed. Advances in Tropical Acacia Research. ACIAR Proceedings No. 35.
Snyder, E.B. (1972). Glossary for forest tree improvement workers. Southern Forest Experiment Station, United States Department of Agriculture.
Turnbull, J.W. & Griffin, A.R. (1986). The concept of provenance and its relationship to infraspecific classification in forest trees. pp. 157–189 in B.T. Styles ed. Infraspecific Classification of Wild and Cultivated Plants. Clarendon Press, Oxford.
Vercoe, T.K. & McDonald, M.W. (1991). E. pellita F. Muell. and Acacia seed collection in New Guinea, September–October 1990. Recursos Genéticos Forestales. Información no 19 - FAO, págs. 38–42.
Williams, E.R. and Matheson, A.C. (1993). Design and analysis of forestry field trials. ACIAR/CSIRO, Australia. (en prensa).
Yang Minquan & Zeng Yutian. (1991). Results from a four-year-old tropical Acacia species/provenance trial on Hainan Island, China. pp. 170–172 in Turnbull, J.W. ed. Advances in Tropical Acacia Research. ACIAR Proceedings No. 35.
Zobel, B.J. & Talbert, J.T. (1984). Applied Forest Tree Improvement. John Wiley and Sons, New York.
Recursos Genéticos Forestales Información No. 21. FAO, Roma (1994)
Recibido en junio de 1993
![]()
![]()
![]()