![]()
![]()
![]()
La pêche est un facteur sélectif qui est susceptible de modifier les populations. De nombreux pêcheurs recherchent une espèce ou un groupe d'espèces particuliers et s'intéressent aux adultes de grande taille par opposition aux juvéniles. Il est bien connu que selon la méthode de pêche employée on prend différentes tailles de poisson dans les mêmes parages: c'est le cas par exemple des pêcheries au chalut et à la palangre de l'Atlantique Nord (ICNAF 1963) et de la pêche du saumon du Pacifique à la senne, au filet maillant et à la traîne au large de la Colombie britannique (Ricker et coll. 1978). Malgré ce potentiel de sélection, on a peu de preuves de modifications génétiques dues à la pêche, notamment chez les populations marines. Lors de ses premiers travaux, Miller (1957) est parvenu à la conclusion que l'on avait peu de preuves qu'il y eût des modifications héritables dues à l'exploitation ou aux introductions chez les poissons d'eau douce. Or, depuis lors, plusieurs travaux ont mis en évidence des modifications génétiques chez des populations de poisson exposées à une forte pression de pêche. La plupart de ces études se fondent sur une modification des caractères du cycle vital dans le temps, caractères qui pourraient subir des modifications non génétiques en réponse aux évolutions de l'environnement.
Cela fait des siècles que le cabillaud de l'Atlantique est exploité dans toutes les eaux côtières de l'Atlantique Nord, mais il n'y a qu'une cinquantaine d'années que l'on procède à l'échantillonnage biologique des captures. La découverte et la fouille d'un bateau de pêche perdu en mer au large des côtes de la Nouvelle-Ecosse vers les années 1750 a fourni une occasion unique de comparer les longueurs des cabillauds, à partir de mesures des cleithrums de la ceinture scapulaire, entre le XVIIIe et le XXe siècles (Kenchington et Kenchington 1993). Les cabillauds pris à l'hameçon moderne au large de la Nouvelle-Ecosse sont beaucoup plus petits que les cabillauds pris dans les années 1750, même si l'on pêche à l'occasion des cabillauds d'une taille maximale similaire (Kenchington et Kenchington 1993).
Au large de la Nouvelle-Ecosse, dans l'Atlantique Nord-Ouest, les captures ont atteint un maximum à plus de 80 000 tonnes en 1968, s'accompagnant d'un net déclin de la biomasse et d'un déclin correspondant de la taille et de l'âge moyens des cabillauds (Beacham 1983a). En outre, la longueur et l'âge médians à maturité ont décliné: l'âge médian à la maturité sexuelle a diminué, passant de > 5 ans dans les années 60 à > 3 ans en 1978 (fig. 1), aussi bien dans le cas des mâles que des femelles (Beacham 1983a). Les cabillauds qui parviendraient à maturité à une taille plus petite ou à un plus jeune âge auraient un avantage sélectif en cas de pression de pêche très intense dans la mesure où les cabillauds venant à maturité à une taille plus grande et à un âge plus avancé seraient capturés avant de la maturité sexuelle (Beacham 1983a). On a pu faire des observations similaires dans le cas du cabillaud Atlantique de l'Ouest-Groenland, où l'âge moyen au début de la maturité a reculé de 9,9 ans en 1917 à 6,4 ans en 1936 dans la pêcherie septentrionale et de 9,3 ans en 1922 à 7,6 ans en 1936 dans la pêcherie méridionale (Hansen 1949).
Dans le cas des stocks arcto-norvégiens de cabillaud de l'Atlantique, la majorité des poissons parvenaient à maturité à 8–10 ans dans les années 30, alors qu'au cours des années 70 la majorité des cabillauds étaient à maturité à 6 ans (Borisov 1979). Ce déclin a été attribué au fait que les cabillauds parvenant tardivement à maturation étaient sélectivement retirés de la population (Borisov 1979). On n'a observé sur une période de 40 ans aucune tendance de la longueur pour l'âge qui puisse laisser supposer que la croissance n'était pas fonction de la densité, encore qu'un déclin de la longueur pour l'âge au cours de la seconde moitié des années 80 ait coïncidé avec une diminution de l'abondance relative du capelan, principale proie des grands cabillauds (Jorgensen 1992). Law et Grey (1989) ont suggéré de réglementer l'effort de pêche portant sur les stocks de 4 ans de la Mer de Barents et du plateau du Spitzberg et que la pêche se concentre sur les poissons frayants, de plus grande taille, de la côte norvégienne. Ce type de pêche entraînerait une sélection pour les individus à maturation tardive et permettrait un accroissement de la production.
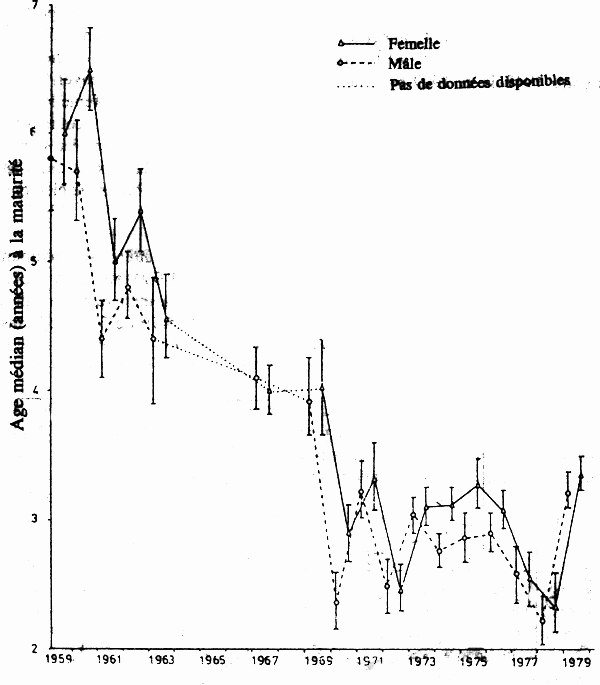
Figure 1.Age médian à la maturité sexuelle pour les cabillauds Atlantique capturés dans la Division des pêches 4W au large de la Nouvelle-Ecosse (Canada) entre 1959 et 1979. Les triangles reliés par un trait plein sont des données concernant les femelles et les cercles reliés par des tirets sont des données concernant les mâles, les pointillés indiquant l'absence de données et les barres verticales les limites de confiance à 95 %. (Données communiquées par Beacham 1983a.)
L'aiglefin a fait l'objet d'une exploitation intensive dans les pêcheries démersales de l'Atlantique Nord-Ouest. Les captures d'aiglefin sur les bancs de Saint-Pierre, au large de Terre-Neuve, ont rapidement progressé jusqu'à atteindre un maximum de 58 000 tonnes en 1955, principalement en raison d'une classe d'âge exceptionnelle en 1949. Cette importante capture a été suivie d'un déclin qui a fait tomber les prises à quelque 6 000 tonnes en 1957, puis à moins de 1 000 tonnes dans les années 70 (Templeman et Bishop 1979a). Les données biologiques recueillies entre 1948–51 et 1969–65 font apparaître un déclin de l'âge moyen à la maturité à 50 % (fig. 2) de 4,6 à 3,3 ans chez les mâles et de 5,9 à 4,3 ans chez les femelles (Templeman et Bishop 1979b). Beacham (1983b) a montré que ce déclin s'est produit au cours d'une période marquée à la fois par des taux de croissance croissants et décroissants, si bien que l'évolution de l'âge à la maturité n'est pas simplement liée aux modifications du taux de croissance dues à l'effet compensatoire d'une biomasse réduite, mais qu'elle est plus vraisemblablement d'origine génétique.
Au large de la Nouvelle-Ecosse, les captures d'aiglefin ont atteint un maximum en 1965 à plus de 50 000 tonnes pour retomber par la suite à moins de 11 000 tonnes en 1967. Au cours de la même période, la longueur médiane à la maturité sexuelle était relativement constante, mais l'âge médian à la maturité a reculé entre 1959–64 et 1975–79 de 3,9 à 2,7 ans pour les mâles et de 4,4 à 3,0 pour les femelles (Beacham 1983c).
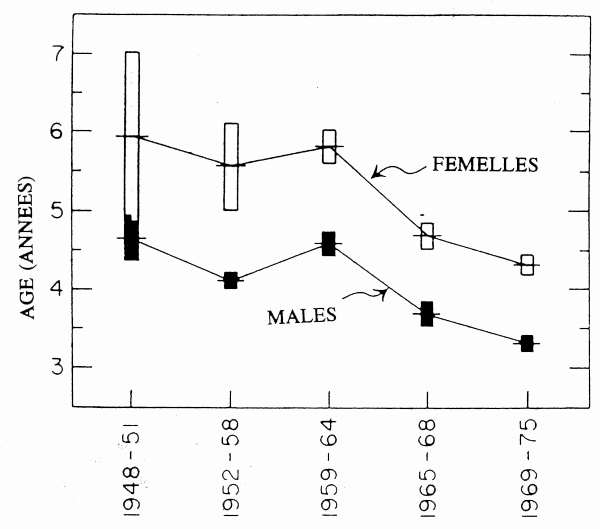
Figure 2. Tendances de l'âge moyen à la maturité sexuelle à 50 % pour les aiglefins mâles et femelles sur les bancs de Saint-Pierre (Canada) entre 1948 et 1975. Les barres verticales représentent les limites de confiance à 95 %. (Données communiquées par Templeman et Bishop 1979b.)
Sur les grands bancs de l'Atlantique Nord-Ouest, les captures de balai (plie canadienne) Hippoglossoides platessoides ont augmenté au cours des années 50 pour atteindre un maximum à environ 90 000 tonnes annuelles dans les années 60 avant de retomber à environ 45 000 tonnes (Pitt 1975). Au cours de la période 1959–72, la taille moyenne pour l'âge a progressivement augmenté alors que l'âge à la maturité sexuelle diminuait (Pitt 1975). La longueur moyenne était liée à l'abondance du stock mais non pas à la température de l'eau, ce qui a amené Pitt (1975) à conclure que l'évolution de lataille pour un âge donné était due dans une grande mesure à une réduction de la densité. La cause de la diminution de l'âge à maturité apparaît moins clairement étant donné qu'on observe des variations régionales de l'âge à maturité; dans certains secteurs, des poissons à la croissance plus rapide parviennent à maturité à un âge plus avancé que des poissons à croissance lente en provenance d'autres secteurs (Pitt 1975). Il est vraisemblable que la pêche retire une plus forte proportion des poissons à croissance plus rapide, entraînant un déclin de l'âge à maturité, non imputable cependant à une augmentation du taux de croissance qui serait sous la dépendance de la densité. Beacham (1983b) a mis en évidence un déclin de la longueur et de l'âge médians à la maturité sexuelle pour la plie canadienne (fig. 3) et la limande à queue jaune Limanda ferruginea dans plusieurs pêcheries de l'Atlantique Nord-Ouest. Ce déclin de la taille et de l'âge à maturité ne se laisse pas facilement expliquer par une augmentation du taux de croissance due à l'effet compensatoire de la biomasse réduite (Beacham 1983b), mais doit vraisemblablement refléter une évolution génétique de la population.
Sur les lieux de pêche de la limande du Japon Limanda espera au large de la Fédération russe, la taille et l'âge auxquels 50 % des femelles parvenaient à maturité ont diminué, passant de 29,9 cm et 8,5 ans à 27,2 cm et 7,2 ans au cours de la période 1961–69 (Tikhonov 1977). Une augmentation de la fécondité des poissons de la même taille a été enregistrée au cours de cette même période.
A l'inverse, en mer du Nord, une augmentation du taux de croissance de la sole Solea solea a pu être mise en relation avec la nourriture rendue disponible par les activités de chalutage plutôt qu'avec une réduction de la densité du stock (de Veen 1976). La taille à maturité est demeurée constante pendant une longue période puis a augmenté, tandis que l'âge à maturité restait sans changement au cours de la même période. De la même manière, Rijinsdrop et coll. (1991) n'ont relevé aucun signe de croissance dépendant de la densité chez les soles et les plies Pleuronectes platessa adultes au cours d'une période de 30 années caractérisée par une exploitation intensive. Chez la plie, l'âge à maturité, mais non pas la taille à maturité, a varié au cours de la période 1958–88 et a pu être mis corrélée positivement avec le taux de croissance des juvéniles. Dans le cas de la sole, il n'y avait aucune relation entre le taux de croissance des juvéniles et l'âge à maturation, et dans un cas comme dans l'autre, qu'il s'agisse des plies ou des soles, il n'y avait pas de tendance cohérente en ce qui concerne le taux de croissance ou la taille et l'âge à maturité (Rijinsdrop 1991).
Les observations faites sur des populations naturelles de langouste suggèrent que la maturité sexuelle est liée à l'âge et non pas à la taille. Lorsque la densité des reproducteurs de Panulirus longipes cygnus de la côte ouest de l'Australie est faible, la taille au début de la reproduction et la gamme des tailles supérieures des femelles est plus élevée (Chittleborough 1979). Lorsqu'il y a abondance (absence d'exploitation) le taux de croissance est ralentie et la maturité sexuelle survient à une taille plus petite (Chittleborough 1979). Chez l'espèce Jasus lalandii de l'écosystème de Benguela au large de l'Afrique du Sud, les taux de croissance et la taille à la maturité sexuelle sont les plus élevés lorsqu'il y a abondance de nourriture (Byers et Goosen 1987). Une réduction de la taille au début de la production des oeufs et un accroissement de la longueur asymptotique ont été signalés chez la langouste Panulirus marginatus des eaux hawaiiennes après qu'un stock vierge eut été soumis à une forte pression de pêche (Polovina 1989). Ce déclin de la taille au commencement de la production des oeufs chez P. marginatus va à l'encontre de ce que pouvaient laisser prévoir les relations tenant à la densité et pourrait indiquer une évolution génétique de la population.
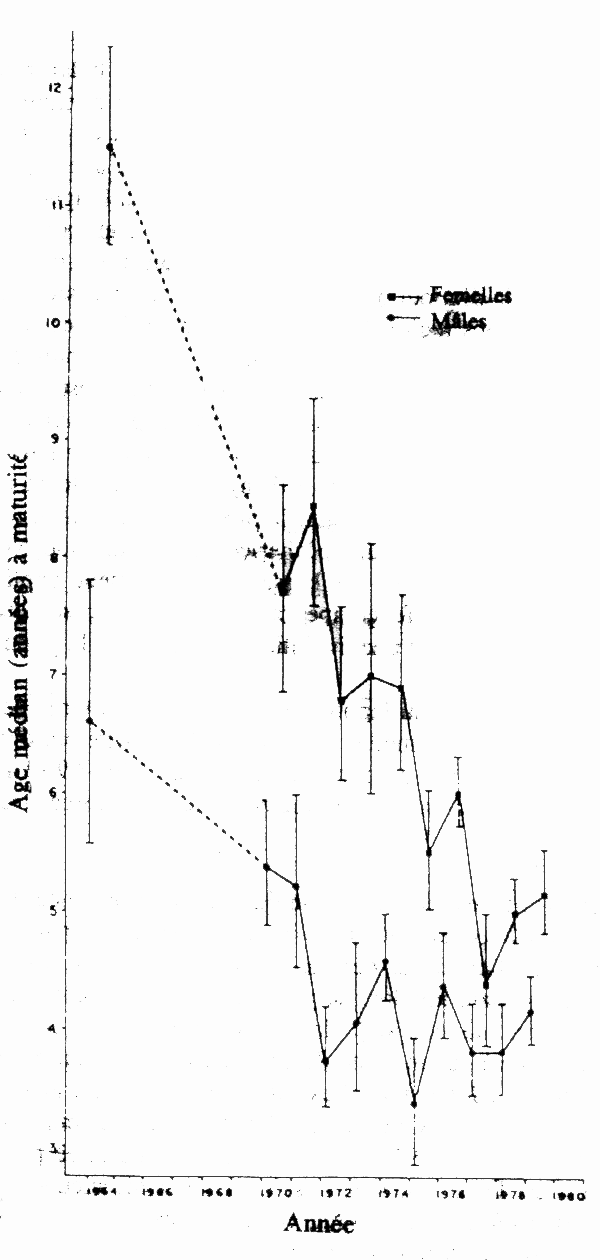
Figure 3. Age médian à maturité sexuelle pour les plies (balais) de l'Atlantique capturées dans la Division de pêche 4W au large de la Nouvelle-Ecosse (Canada) entre 1964 et 1980. Les carrés par un trait plein sont des données concernant les femelles et les cercles reliés par un trait plein des données concernant les mâles; les lignes en pointillés indiquent l'absence de données, tandis que les barres verticales indiquent les limites de confiance à 95 %. (Données communiquées par Beacham 1983c.)
Les Pandalidés sont des crevettes hermaphrodites protéandriques (qui se reproduisent d'abord en tant que mâles) chez lesquels l'âge du début de la reproduction et du changement de sexe qui intervient ensuite est lié à la taille de l'individu (Charnov 1979). Sur les lieux de pêche de la crevette P. borealis dans le Skaggerak une diminution de la taille des femelles s'est produite en période d'exploitation intense (Jensen 1965, 1967). Les captures danoises et suédoises combinées de crevette ont plus que doublé entre 1956 et 1960, tandis que le pourcentage des grosses crevettes, femelles pour la plupart, est passé de 44 % en 1944 à 14 % en 1961. Plus surprenante a été l'apparition d'une classe de femelles de petite taille (tableau 1), à peu près de la même taille que les mâles d'un an. Les femelles de petite taille (< 75 mm) que l'on ne voyait pas en 1953 formaient dès 1961–62 21–30 % de la capture (Jensen 1965). Charnov (1981) a proposé comme explication que l'accroissement de la mortalité par pêche aux dépens des femelles de grande taille a opéré une sélection en faveur des individus qui sont matures en tant que femelles à l'âge du début de la reproduction.
| Période | ||||
|---|---|---|---|---|
| Sexe | 1949–53 | 1954–57 | 1958–60 | 1961–62 |
| longueur moyenne (mm) | ||||
| Mâles et femelles en transition | 74 | 69 | 68 | 67 |
| 96 | 87 | 83 | 82 | |
| pourcentage | ||||
| Mâles et femelles en transition | 65 | 81 | 85 | 83 |
| < 74 mm | 0 | 2 | 3 | 4 |
| > 75 mm | 35 | 17 | 12 | 14 |
Chez les espèces caractérisées par une période de frai étendue et les schémas saisonniers de comportement et de mouvement que cela suppose, la pêche peut provoquer une mortalité plus importante à l'une des composantes de la ressource. S'agissant de pêcheries saisonnières et non réglementées, les opérateurs commerciaux peuvent être tentés de faire un maximum de prises dès l'ouverture de la campagne, ce qui peut soumettre à une pression accrue les individus revenus précocement sur les bancs et favoriser les retardataires. Le moment de la première arrivée, puis du frai du hareng de l'Atlantique C. harengus sur les côtes norvégiennes a évolué sur une période de 60 ans (Devold 1963). Vers le début du siècle, les harengs commençaient à revenir sur les frayères en septembre-octobre, mais cette date de retour s'est trouvée progressement retardée, si bien que dans les années 50 les harengs n'apparaissaient plus sur les frayères que vers le mois de janvier (Devold 1963). Pour Mathisen (1989), cette évolution s'expliquerait par les effets de la pêche qui a sélectivement prélevé les sous-groupes de hareng qui se présentaient les premiers, si bien que, avec le temps, ces premiers arrivés contribuent moins à la pêche et sont progressivement remplacés par des sous-groupes dont le retour a été plus tardif.
Cette interprétation des observations de Devold est à tempérer par les évolutions non génétiques qui ont pu se produire au cours de la période. Les changements de l'époque du frai observés chez d'autres groupes de hareng de l'Atlantique ont été expliqués par des modifications des conditions du milieu. Le hareng est une espèce phénotypiquement plastique que l'on a subdivisée en un grand nombre de groupes taxonomiques sur la base de caractères morphométriques et de périodes et de lieux de frai (Parrish et Saville 1965). On ne sait pas si l'époque du frai chez le hareng est héréditaire, mais tout semble indiquer l'intervention d'une forte composante environnementale. Dans la mer Baltique, les harengs fraient entre le printemps et l'automne (Aneer 1985), les moins féconds frayant au printemps et les plus féconds à l'automne (Anokhina 1971). La disparition de harengs qui fraient en automne a été expliquée par une amélioration des conditions de nutrition due à l'eutrophication, les adultes ayant suffisamment de réserves alimentaires pour frayer au printemps (Aneer 1985). Dans l'Atlantique Nord-Ouest, on a constaté que les harengs nés en automne et au printemps, identifiés par les caractéristiques de leurs otolithes, frayaient durant la saison opposée (Messieh 1972).
Etendant à une autre espèce cette hypothèse reposant sur la sélection par le frai, Mathisen (1989) a suggéré que le déclin de l'anchois péruvien Engraulis ringens était accéléré par les effets disruptifs d'une pêche sélective pratiquée aux dépens des “meilleures unités reproductrices”, ne laissant, pour la reconstitution des stocks, que des groupes marginaux.
Dans les pêcheries de saumon du Pacifique en Colombie britannique, la plupart des poissons capturés sont des adultes matures qui reviennent vers les rivières pour y frayer. A cet égard, les pêcheries ont moins de chances de mettre en évidence l'effet d'épuisement par la pêche constaté dans les pêcheries au chalut dans le cas des téléostéens marins. Une étude approfondie portant sur cinq espèces de saumon du Pacifique a permis de conclure qu'au moins trois espèces présentaient une diminution de la taille moyenne pour l'âge après des années de pêche sélective à l'égard de la taille (Ricker 1981). Les pêcheurs au filet maillant et à la traîne de saumon argenté Oncorynchus kisutch et de saumon rose O. gorbuscha ont tendance à prendre les poissons les plus gros et ce sont ces espèces qui ont présenté les plus fortes réductions des tailles moyennes (Ricker 1981). On a observé entre 1951 et 1975 (Ricker 1980) une double diminution de la taille (fig. 4) et de l'âge moyens du saumon royal O. tshawytscha. Pour Ricker (1981), cette diminution de la taille et de l'âge serait due à la pêche à la traîne qui, en capturant à la fois les individus qui sont et qui ne sont pas en voie de maturation, opère une sélection défavorable aux individus à maturation tardive. Toutefois, les données sont complexes étant donné que les méthodes de pêche et les grandeurs de maille ont changé au cours de la période d'échantillonnage et que dans le cas d'O. nerka on a assisté au cours de la période à une augmentation du taux de croissance liée au refroidissement océanique (Ricker 1981).
Chez le saumon argenté O. kisutch, les mâles arrivent à maturation après six mois dans l'océan, pour devenir des madeleineaux, ou se transforment en bécards après 18 mois. Ces deux cycles biologiques différents sont le résultat d'une sélection disruptive (naturelle) favorisant les petits mâles fuyants et les grands mâles agressifs (Gross 1985). Or, la pêche a sélectivement exploité les poissons de grande taille (voir fig. 5), et conduit à un accroissement relatif de madeleineaux (Gross 1991). Toutefois, Gross (1991) a montré que des changements intervenus lors d'autres stades du cycle biologique peuvent influencer le rapport entre madeleineaux et bécards. Le déblaiement des ruisseaux, par exemple, peut réduire les abris disponibles pour les madeleineaux, favorisant ainsi la présence de bécards sur les aires de frai. D'autres phénomènes écologiques comme l'eutrophisation peuvent accélérer les taux de croissance des alevins et conduire à une augmentation de la proportion de ces derniers qui arrivera à maturité sous forme de madeleineaux (Gross 1991).
Le saumon de l'Atlantique S. salar présente d'importantes variations de l'âge moyen au premier frai selon les rivières qu'il fréquente. On pense que ces variations résultent de l'adaptation et qu'elles sont entretenues par la sélection naturelle (Schaffer et Elson 1975). La pression de pêche qui s'exerce sur les adultes à leur retour a éliminé les poissons les plus gros et les plus âgés de certains parages, d'où l'âge précoce des premiers poissons qui reviennent vers les rivières (Schaffer et Elson 1975).
Chez les espèces ayant une longue saison de frai, l'irrégularité de la pression de pêche au cours de la saison peut entraîner une mortalité sélective. C'est ainsi que chez le saumon argenté Oncorhynchus tsawytscha du fleuve Columbia la saison de frai dure d'avril à août avec un pic en juin-juillet lorsque la pêcherie a commencé à fonctionner au siècle dernier (Thompson 1951). La pêcherie était surtout exploitée l'été, saison favorable à la fois à cause du temps et de l'abondance du poisson. Une pression de pêche très forte au cours de la saison de pointe a réduit les captures, si bien qu'en 1938 l'effectif total capturé avait diminué et que les pics de la saison du frai se situaient au début du mois de mai et au mois d'août (fig. 6), peu de poissons étant capturés au cours de l'ancienne période de pointe de juin-juillet (Thompson 1951). Mathisen (1989) a interprété ces observations comme une sélection disruptive, avec capture sélective des reproducteurs précoces et tardifs, lesquels n'ont pas remplacé les reproducteurs de la période de pointe intermédiaire.
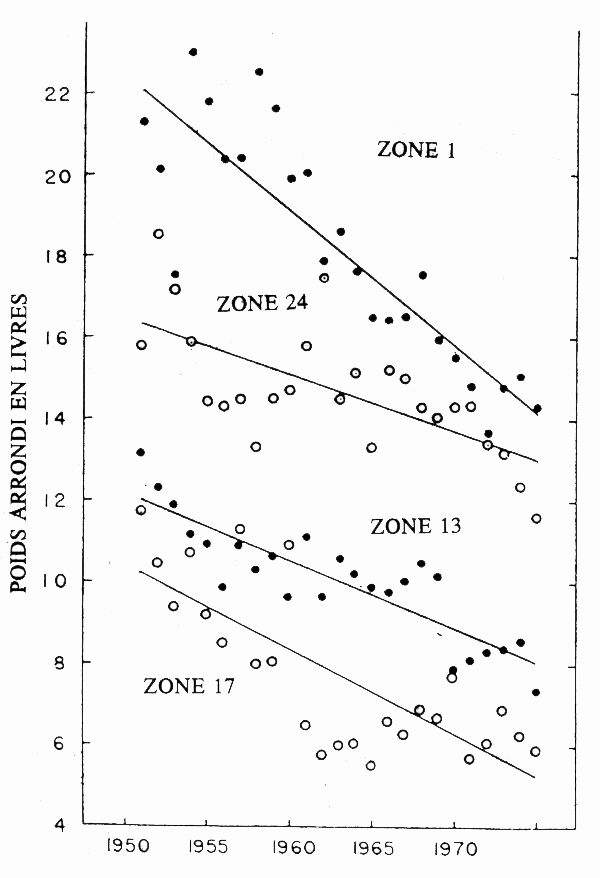
Figure 4. Poids médians des saumons argentés pris à la traîne au large de la Colombie britannique (Canada). (Données communiquées par Ricker 1981.)
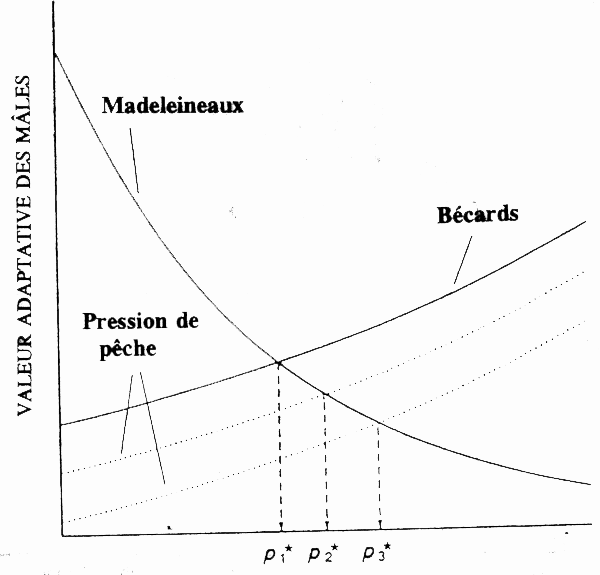
Figure 5. Effets de la pêche sélective à l'égard de la taille sur les madeleineaux et bécards des saumons argentés mâles. Le prélèvement sécards mâles accroît le pourcentage des madeleineaux dans la pêcherie; p1* pêche nulle, p2* et p3* niveaux de pêche croissants (d'après Gross 1991).
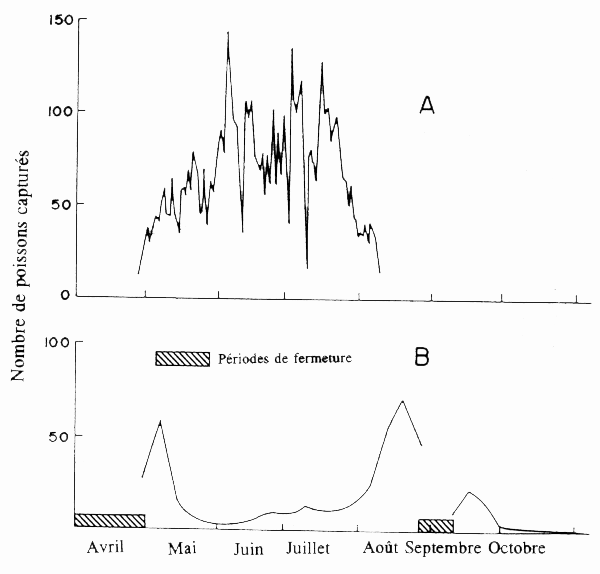
Figure 6. Evolutions saisonnières des captures de saumon argenté dans le fleuve Columbia. A. Captures journalières au filet maillant en 1876 et B. Captures en 1938. (Données communiquées par Mathieson 1089.)
Ricker (1982) a décrit une sélection disruptive pour la taille chez le saumon rouge Oncorhynchus nerka. L'opération de pêche au filet maillant a capturé des poissons appartenant à la gamme des tailles moyennes, favorisant ainsi la survie des petits poissons océaniques de trois ans et des gros poissons océaniques de quatre ans. De ce fait, les poissons de trois ans sont devenus plus petits et les poissons de quatre ans plus gros, la différence entre les deux augmentant d'environ 500 g (Ricker 1982).
De nombreuses pêcheries lacustres utilisent des filets maillants qui prélèvent sélectivement les gros poissons lourds, ce qui entraîne une mortalité non aléatoire sous le rapport de la taille (Hamly 1975). Au Canada, les corégones sont abondants et le corégone de lac Coregonus clupeaformis a été exploité dans plusieurs lacs. Si l'on compare les populations exploitées et non exploitées de corégone lacustre, on constate les effets compensatoires caractéristiques de la pêche avec augmentation du taux de croissance due à l'exploitation (Healey 1975). Le taux de croissance chez les populations fortement exploitées était similaire au taux de croissance le plus rapide chez les populations non exploitées (Healey 1975). Toutefois, chez les populations fortement exploitées, les poissons parvenaient à maturité à un âge plus jeune et à une taille plus petite que les poissons appartenant aux populations non exploitées.
Dans le Petit Lac de l'Esclave, en Alberta, les ressources en poisson ont été fortement exploitées. Dès les années 20, la pêche avait eu raison de la truite de lac Salvelinus namaycush, tandis que les autres espèces devenaient moins abondantes (Handford et coll. 1977). Entre 1941 et 1975, des échantillons de C. clupeaformis prélevés sur des captures commerciales et des captures réalisées dans le cadre de la recherche ont été analysés. Après un accroissement initial du poids et de la longueur moyens on observait une diminution de la longueur et du poids à un âge donné. Dès les années 70, les poissons d'un âge donné pesaient moins de la moitié que les poissons du même âge dans les années 40, alors que la longueur moyenne pour un âge donné était environ la même dans les années 70 que dans les années 40 (Handford et coll. 1977). Des observations similaires ont pu être faites dans d'autres lacs (Handford et coll. 1977). En outre, les captures réalisées dans le Petit Lac de l'Esclave ont fait apparaître un accroissement global de l'âge moyen, résultat auquel on pouvait s'attendre aux niveaux d'exploitation modérés, la sélection favorisant des taux de croissance plus lents et annulant les effets contraires de la compensation dépendant de la densité (Handford et coll. 1977). Dans le cas des Percidés, on a observé dans les pêcheries lacustres des schémas analogues avec accroissement du taux de croissance et abaissement de l'âge au premier frai (Spangler et coll. 1977).
Les populations de poissons du Lac George (Ouganda) ont été étudiées entre 1967 et 1972 lorsque le Cichlidé Tilapia nilotica représentait environ 80 % des captures. C'est la pêche qui a été la source majeure de mortalité des adultes de cette espèce dont la taille moyenne est passée d'environ 0,9 kg en 1950 à 0,4 kg en 1970. Au cours de la même période, la taille à maturité a diminué, de 20–29 cm à 18–24 cm (Gwahaba 1973).
Dans le Lac Femund, en Norvège, le lavaret polymorphe présente trois morphologies. Une exploitation de pêche au filet maillant pélagique, créée dans les années 80, a opéré une sélection aux dépens de la forme pélagique, laquelle a diminué par rapport aux deux autres formes et a fait apparaître une diminution du pourcentage des gros poissons et une réduction de la taille pour un âge donné (Sandlund et Naesje 1989).
Dans les années 80, il s'est créé au large des côtes de Nouvelle-Zélande une nouvelle exploitation de l'hoplosthète orange et au cours des dix premières années d'exploitation la biomasse a été réduite de 60–70 %. La diversité génétique, mesurée par électrophorèse sur gel, a fait apparaître une diminution significative sur trois bancs de pêche géographiquement isolés (tableau 2) entre 1982–83 et 1988 (Smith et coll. 1991). La récente découverte de nouvelles frayères fournit une nouvelle occasion de vérifier les effets génétiques de l'exploitation sur les stocks vierges. Les premiers résultats fournis par l'analyse des allo-enzymes font apparaître des niveaux de diversité génétique similaires aux niveaux rapportés en 1982–83, avant le début de l'exploitation (Smith, non publié). Dans une situation analogue, concernant cette fois la langouste Panulirus marginatus, aucune modification génétique n'avait été décelée sur une période de neuf ans consécutive à l'exploitation intensive d'un stock vierge (Seeb et coll. 1990).
| Banc de pêche | Année d'échantillonnage | Hétérozygotie moyenne | Probabilité |
|---|---|---|---|
| Chatham Rise | 1982 | 0,20 | <0,02 |
| (Océan Pacifique) | 1988 | 0,14 | |
| Côte est | 1983 | 0,25 | <0,02 |
| (Océan Pacifique) | 1988 | 0,17 | |
| Challenger Plateau | 1983 | 0,22 | <0,01 |
| (Mer de Tasmanie) | 1988 | 0,15 |
Chez les populations de saumon rouge qui fraient dans des lacs isolés de la péninsule du Kamtchatka, les juvéniles passent d'un à trois ans dans les lacs pour gagner ensuite la mer pour une nouvelle période d'un à quatre ans avant de revenir sous forme d'adultes sexuellement matures. Les mâles arrivent à maturité à deux tailles différentes: comme mâles de grande taille au bout de deux ans et plus en mer, et comme madeleineaux ou “kayiriki” au bout d'un an en mer (Altukhov 1990). En outre, un certain nombre de mâles résiduels des lacs atteignent la maturité sous forme de “nains” (Altukhov 1990). Ce dimorphisme sexuel est semblable à celui signalé chez le saumon argenté O. kisutch (Gross 1985). Les saumons rouges mâles de grande taille ont l'avantage sur les mâles de petite taille sur les frayères, alors que les mâles de petite taille reprennent l'avantage dans les années de basses eaux et de frayères peu profondes où les mâles de grande taille ne peuvent pas pénétrer (Altukhov 1990).
Sur une période de 35 ans, entre 1935 et 1976, les effectifs de saumon rouge retournant au Lac Dalneye, sur la péninsule du Kamtchatka, ont marqué une baisse spectaculaire (Altukhov 1990), tandis que la proportion des madeleineaux s'accroissait parmi les mâles de retour et que le pourcentage des mâles nains résidents augmentait considérablement parmi la population (tableau 3). Des récentes analyses d'allo-enzymes, portant sur des populations de saumon rouge dans trois lacs du Kamtchatka (tableau 4) ont montré que l'hétérozygotie mesurée à deux loci enzymatiques est plus grande chez les mâles de petite taille (madeleineaux) que chez les mâles de grande taille (Altukhov 1993). Les fréquences d'allèles sont similaires chez les deux groupes, mais les mâles de petite taille se caractérisent par un excès significatif d'hétérozygotie au locus de la phosphoglucomutase (PGM), tandis que les mâles de grande taille présentent un excès significatif d'homozygotes dans les deux lacs. Chez les femelles, l'hétérozygotie est intermédiaire entre les mâles de petite et de grande taille (Altukhov 1990). Pour Altukhov (1990), ces changements concernant l'hétérozygotie s'expliqueraient par les effets sélectifs de la pêche. Il se peut que la pêche ait favorisé la survie des mâles de petite taille, comme ont pu le faire aussi des changements du milieu aquatique d'eau douce; toutefois on ne dispose pas d'antécédents historiques concernant les allo-enzymes pour les deux catégories de tailles chez les mâles. Dans trois lacs du Kamtchatka, les niveaux d'hétérozygotie vis-à-vis la PGM s'accroissent avec le pourcentage de mâles de petite taille présents dans le lac (Altukhov 1990).
Kirpichnikov et al. (1990) ont montré qu'au cours des stades de l'alevinage les individus à croissance lente sont moins hétérozygotes que les individus à croissance rapide. Si ces individus à croissance rapide parviennent précocement à maturité sous la forme de nains ou de madeleineaux – et chez les salmonidés une croissance rapide va de pair avec une maturité précoce - cela rendrait compte des observations d'Altukhov (1990). L'hétérozygotie globale de la population s'est accrue en raison des effets indirects de la pêche, favorable à la survie des mâles de petite taille.
O. mossambicus est très couramment utilisé en aquaculture tropicale en étang. Il s'agit d'un incubateur buccal, facile à élever en bassin. Pour mesurer l'impact de la pêche sélective, deux populations ont été établies puis, au bout de 39 mois, pêchées à deux mois d'intervalle (Silliman 1975). Environ 10 % des poissons ont été retirés: l'une des populations a fait l'objet d'une pêche sélective qui ne prélevait que les poissons ne pouvant pas passer à travers une grille placée dans le bassin. La seconde population, population témoin, a été pêchée de manière non sélective, avec prélèvement d'un nombre égal de poissons, mais appartenant à toutes les classes de taille. Au bout de 63 mois, la pression de pêche a été portée à 20 % et poursuivie pendant 12 mois. Pour vérifier l'impact génétique de la pêche, des groupes de 46 poissons à maturité ont été retirés des populations sélectionnées et non sélectionnées au bout de 77 mois et maintenus en élevage pendant une nouvelle période. La croissance était mesurée au bout de 55–56, 118–119 et 150–151 jours après le début de la seconde expérience. Aussi bien chez les populations sélectionnées que non sélectionnées les mâles croissaient plus vite que les femelles mais, au sein de la population non sélectionnée, les mâles croissaient plus rapidement que les mâles de la population sélectionnée (Silliman 1975). Ainsi, trois générations ont suffi pour que la pêche sélective entraîne un abaissement du taux de croissance chez les mâles. Il est vraisemblable que les femelles étaient moins affectées par la sélection opérée en fonction de la taille en raison de leur taux de croissance plus faible que celui des mâles.
| Périodes | ||||
|---|---|---|---|---|
| Saumon | 1935–46 | 1947–56 | 1957–65 | 1966–76 |
| Nombre moyen de reproducteurs pénétrant dans le lac (en dizaines de milliers) | 63 | 10 | 6 | 1,6 |
| Pourcentage de madeleineaux parmi les mâles migrateurs | 0,2 | 0,6 | 4,3 | 37,5 |
| Pourcentage de nains parmi la population mâle | 26 | 49 | 74 | 89 |
| Lac | Poisson | Hétérozygosité | |
|---|---|---|---|
| LDH | PGM | ||
| L. Nachikinskoye | mâles de petite taille | 0,43 | 0,55* |
| mâles de grande taille | 0,39 | 0,35* | |
| L. Blizhneye | mâles de petite taille | 0,19 | 0,56* |
| mâles de grande taille | 0,03 | 0,36* | |
| L. Dalneye | mâles anadromes | 0,21 | 0,31 |
| mâles nains | 0,16 | 0,62* | |
* déséquilibre génétique significatif.
Les différences observées dans les populations de guppys sont liées à la prédation: certains prédateurs (Cichlidés) font leurs proies de guppys de grande taille parvenus à maturité sexuelle, tandis que d'autres prédateurs (fondules) font leurs proies de petits guppys immatures. Une population de guppys a été transférée d'une rivière comportant un prédateur Cichlidé dans un affluent où il n'y avait pas de guppys mais des prédateurs fondules. Sur une période de 11 ans, la population transférée a produit une descendance de plus grande taille et des femelles de plus grande taille (fig. 7) qui se sont reproduites pour la première fois à une taille plus grande que les femelles de la population témoin (Reznick et coll. 1990). En élevant des groupes de descendants provenant des deux populations dans les mêmes conditions de laboratoire, on a montré que les différences constatées au cours de l'existence avaient une base génétique (Reznick et coll. 1990).
Une expérience de laboratoire a été conduite avec la puce d'eau D. magna pour mesurer l'impact de deux régimes de récolte opposés, l'un retirant les individus de petite taille et l'autre les individus de grande taille (Edley et Law 1988). A la première récolte, 50 % du stock a été retiré après quoi l'on en a retiré 40 % tous les huit jours. Les rendements diminuaient avec les deux régimes de récolte, mais ils étaient les plus élevés dans le cas des populations sur lesquelles on prélevait les individus de grande taille. Lorsqu'on retire les individus de petite taille, la taille moyenne pour un âge donné augmente ainsi que la taille au début de la reproduction, tandis que lorsque l'on retire les individus de grande taille c'est l'inverse qui se produit, avec diminution de la taille pour un âge donné et diminution de la taille au début de la reproduction (Edley et Law 1988).
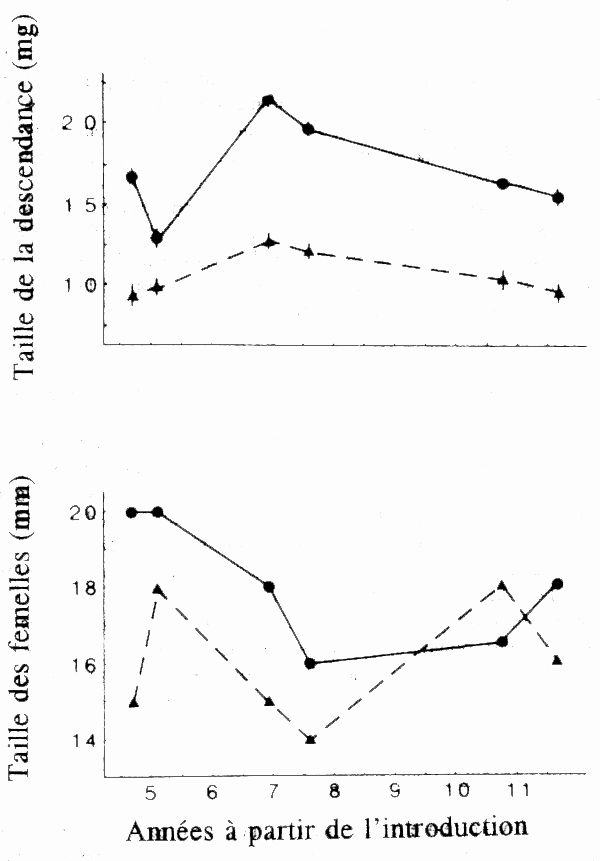
Figure 7. Taille de la descendance et taille moyenne des femelles parvenues à maturité sexuelle dans les échantillons transféré et témoin de guppys sur une période de 11 ans dans la rivière Aripo à Trinidad. Les cercles reliés par un trait plein représentent le site d'introduction tandis que les triangles reliés par un trait en pointillés représentent le site témoin le principal prédateur faisait sa proie prinicipale des guppys de grande taille, tandis qu'au site de transfert le principal prédatuer faisait sa proie principale de guppys de petite taille. (Données empruntées à Reznick et coll. 1990.)
Menshutkin et coll. (1989) ont mis au point un modèle de simulation des opérations de pêche du saumon rouge Oncorhyncus nerka au Kamtchatka. Lorsque la croissance était sous la dépendance d'un seul gène à deux allèles, la population modèle devenait instable avec perte d'un allèle, ce qui corroborait les observations montrant que la croissance est sous dépendance polygénique. Lorsque la croissance était polygénique et multiallélique, le pourcentage des allèles à “croissance rapide” (synonyme de maturité précoce) augmentait dans la population sélectionnée et le pourcentage des poissons non migratoires augmentait. Les observations relatives à la pêcherie où l'on a observé un accroissement du pourcentage des madeleineaux et des poissons demeurant dans le lac (Altukhov 1990) pouvaient laisser prévoir ces résultats. Favro et ses collaborateurs (1979, 1982) ont fait appel à un modèle de simulation pour estimer l'ampleur des effets génétiques sur une pêcherie de truites soumise à des limites de taille minimale et où la croissance était sous la dépendance d'un petit nombre de gènes majeurs. Les résultats ont montré que la taille moyenne et l'effectif total diminuaient pour des niveaux modérés de pression de pêche et recoupaient des observations dont avait fait l'objet une pêcherie de truite Salmo trutta dans le Michigan. Si l'on étendait le modèle de manière à sélectionner les poissons d'une gamme de tailles spécifique, moyennant laquelle une limite de taille minimum et maximum était fixée dans l'exploitation, le modèle montrait que cette double limite de taille entraînait une diminution comparable du nombre des poissons de grande taille que les limites de taille minimale conventionnelles (Favro et coll. 1980).
Law et Edley (1989) ont fait appel à un modèle spécifique basé sur l'âge pour décrire l'exploitation qui exerce une force sélective sur les variations génétiques des caractères du cycle vital. La pêche sélective entraîne dans l'existence des individus des modifications qui, à leur tour, déterminent des changements évolutifs de la production. En principe, le responsable de la pêcherie peut opter pour le schéma de pêche qui opérera une sélection qui donnera un cycle vital optimum, fournissant la production la plus élevée possible, appelé par les auteurs “stratégie de récolte optimale stable évolutive” (Law et Edley 1989). En appliquant ce modèle au cabillaud arcto-norvégien Gadus morhua, qui a présenté un déclin de l'âge à la maturité, un accroissement de la production pourrait être obtenu en opérant une sélection des poissons qui viennent tardivement à maturité. Dans la pratique, il faudrait pour cela limiter la capture des immatures des bancs septentrionaux et faire porter l'effort sur les pêcheries de frai (Law et Edley 1989).
Une approche de gestion expérimentale à grande échelle destinée à tester l'héritabilité du taux de croissance et les effets d'une pêche sélective à l'égard de la taille, concernant une population naturelle de saumon rose Oncorhynchus gorbuscha, a été proposée par McAllister et Peterman (1992) dans le but d'accroître la biomasse capturée. Cette espèce, comme c'est le cas d'autres salmonidés de la côte occidentale, a présenté une réduction de la taille moyenne au cours des vingt dernières années (voir section g) ci-dessus et Ricker et coll. 1978) et l'on ne sait pas très bien si ces changements sont dus au prélèvement sélectif des poissons de plus grande taille, à l'épuisement sélectif des stocks composés de poissons de grande taille ou à des modifications des conditions océanographiques (McAllister et Peterman 1992). Une récolte sélective, contrôlée, de poissons de petite taille permettrait de vérifier l'héritabilité du taux de croissance et, à la condition que ce taux de croissance soit hérité, déboucher sur un accroissement de la taille moyenne dans la pêcherie. Des simulations du type Monte Carlo ont montré que des dispositifs par blocs comportant un petit nombre de répliques et de brèves durées permettaient de fonder statistiquement la détermination de l'importance de la pêche sélective vis-à-vis de la taille. Prolongeant ce dispositif expérimental, McAllister et Peterman (1992) ont fait appel à une analyse de la décision pour démontrer que l'approche expérimentale de la gestion des stocks de saumon rose avait toutes chances de permettre des captures de plus grande valeur que le type de gestion actuellement pratiqué.
![]()
![]()
![]()