![]()
![]()
![]()
32. Quelles modifications peut-on attendre dans la croissance et le rendement des arbres et des forets suite au changement climatique?
33. Quels changements peut-on attendre dans les aires de répartition naturelle des arbres et des communautés végétales sous l'effet du changement climatique?
34. Quel est le risque de voir certaines espèces ou communautés végétales menacées d'extinction par le changement climatique?
35. Comment le changement climatique pourrait-il influer sur la fréquence et l'intensité des feux incontrôlés?
36. Quels effets le changement climatique pourrait-il avoir sur la santé des forets, leur sensibilité aux ravageurs et aux maladies ou leur dépérissement?
Les conséquences de l'accumulation de CO2 sur la croissance et la productivité des arbres et des forêts ne sont pas encore claires. Des études menées en laboratoire sur les taux de croissance et la productivité des plantes cultivées dans des milieux enrichis en CO2 ont mis en évidence une augmentation du taux de photosynthèse, une diminution des besoins en eau des plantes, une augmentation des quantités de carbone piégées et une intensification de l'activité microbienne dans le sol. Il en résulte des taux supérieurs de fixation d'azote, stimulant la croissance. Toutefois, dans un écosystème naturel, où les animaux se nourrissent de plantes, où les organismes pathogènes endommagent les arbres ou causent leur mort et où les végétaux se disputent la lumière, l'eau et les nutriments, un véritable accroissement de la production semble fort douteux. De plus, l'augmentation de la croissance et des rendements pourrait être compensée par des pertes plus élevées dues aux feux, aux insectes et aux maladies (voir questions 35 et 36).
A l'heure actuelle, peu de travaux ont été réalisés pour vérifier les effets de concentrations supérieures de CO2 sur les forêts ou d'autres communautés végétales naturelles sur de longues périodes. Par conséquent, l'effet net du changement climatique sur la croissance et le rendement des forêts est incertain. Sedjo et Solomon (1989) concluent que l'effet fertilisant dû au CO2 n'a pas encore été détecté chez les arbres, bien que des recherches approfondies aient été réalisées à ce sujet sur le terrain et en salles de culture.
Quand le régime des températures et des précipitations change, les aires de répartition naturelle des espèces animales et végétales changent aussi. Avec le réchauffement de la Terre, les espèces tendent à déplacer leurs aires vers des latitudes et des altitudes plus élevées. Pour chaque degré (1°C) de plus, les aires de répartition des arbres dans l'hémisphère nord ont la capacité de s'étendre de 100 km vers le nord tandis que les limites méridionales reculent. Ce phénomène a été observé depuis la dernière période glaciaire (Davis, 1989).
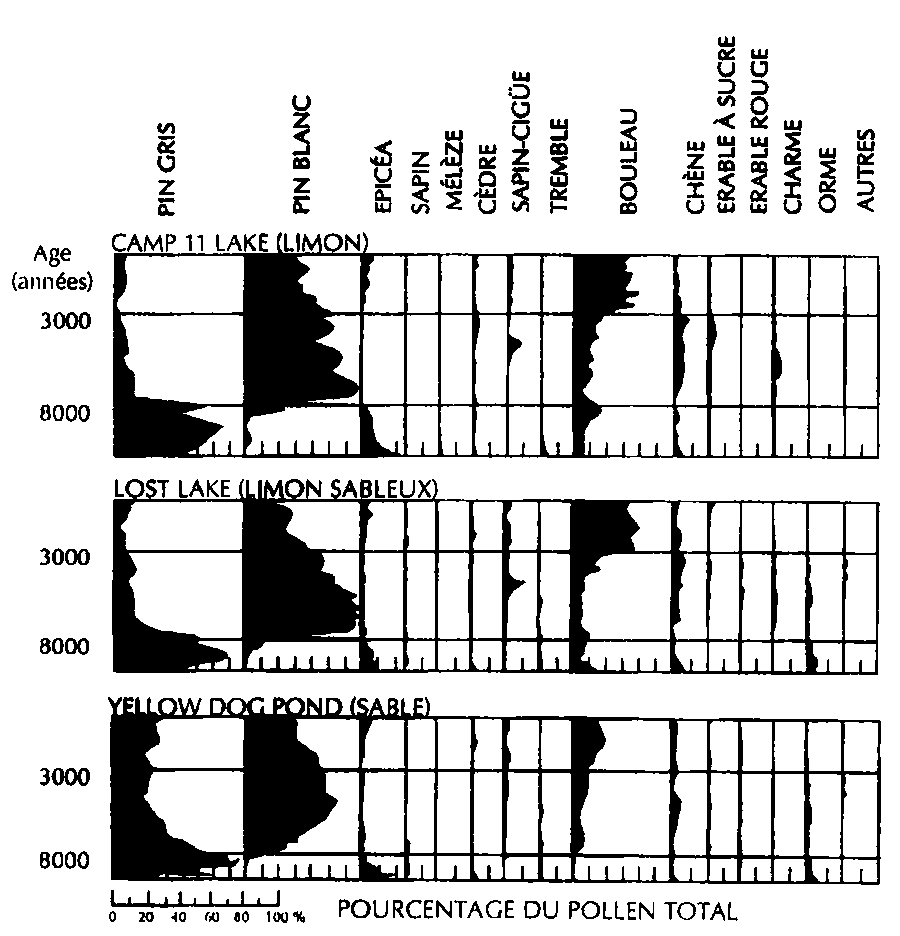
Les donnés fournies par les fossiles révèlent que les végétaux ont subi des déplacements importants de leurs aires de répartition à la suite des changements climatiques. L'analyse des données des pollens fossiles renseigne également sur la composition de la végétation d'autrefois (Brubaker, 1975; Solomon et Bartiein, 1992) (Fig. 6.1). Durant les périodes interglaciaires du Pléistocène, en Amérique du Nord, les températures étaient de 2° à 3°C supérieures à celles d'aujourd'hui. Des arbres tel que le liquidambar d'Amérique, Liquidambar styraciflua, ou l'oranger des Osages, Maclura pomifera, aujourd'hui considérés comme des éléments typiques de la végétation forestière du Sud-Est des Etats-Unis, étaient présents près de Toronto au Canada. Durant la dernière période interglaciaire, qui s'est achevée il y a plus de 100 000 ans, les zones recouvertes aujourd'hui d'une végétation boréale dans le nord-ouest de l'Europe étaient à dominante tempérée. Plus récemment, en Suède, l'aire de répartition naturelle du bouleau, Betula pubescens, a réagi rapidement au réchauffement durant la première moitié du vingtième siècle en s'étendant vers le nord dans la toundra (Peters, 1990).
Les déplacements des aires naturelles des arbres pourraient être importants pour plusieurs raisons. Premièrement, il semblerait que le climat pourrait changer plus vite que certaines essences ne peuvent réagir par migration. Deuxièmement, les caractères édaphiques des nouveaux sites ne sont pas toujours adaptés à cette migration. Enfin, les zones climatiques futures, en provoquant le déplacement des écosystèmes forestiers, ne correspondraient pas aux frontières politiques actuelles et (ou) aux modes d'utilisation des sols (Izrael et al, 1990).
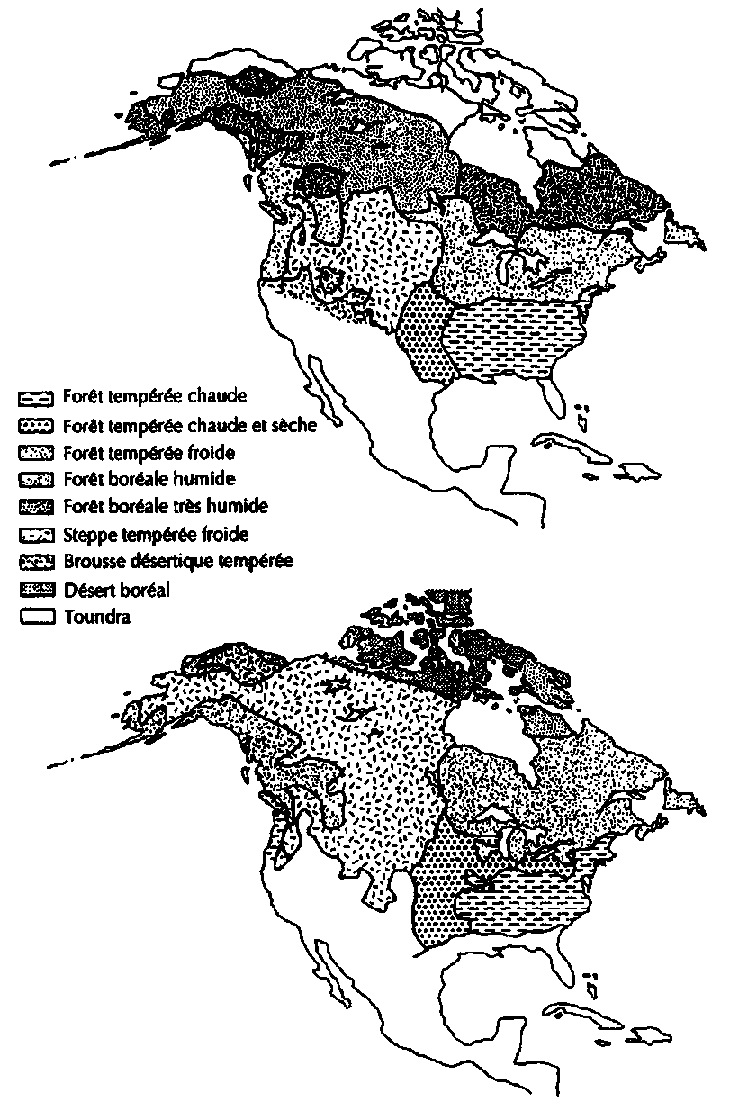
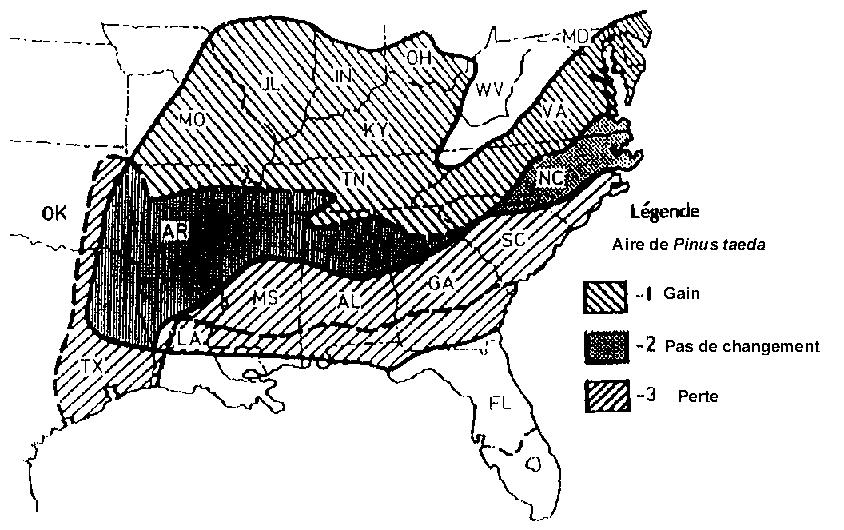
Certaines études prévoient des déplacements des aires naturelles des écosystèmes végétaux (Fig. 6.2) et de certaines espèces d'arbres, pouvant résulter des variations de température et d'humidité dues aux niveaux de concentration des gaz à effet de serre dans l'atmosphère. Selon Miller et al (1987), Pinus taeda (loblolly pine), importante essence forestière industrielle dans le sud-est des Etats-Unis, se déplacerait de 350 km environ vers le nord dans le cas d'un réchauffement de la planète de 3°C (Fig. 6.3).
Des déplacements des aires naturelles des animaux et des végétaux auront lieu en fonction des besoins de chaque espèce et pas nécessairement de l'écosystème dans son ensemble. On peut donc s'attendre à quelques réajustements des associations d'espèces si le climat devait changer.
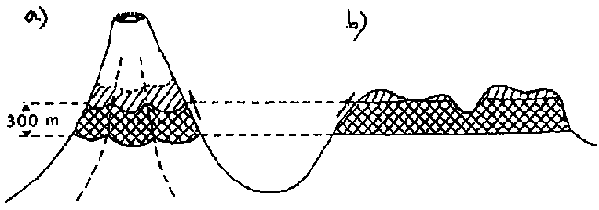
Le déplacement des espèces peut s'effectuer aussi bien en altitude qu'en latitude. Quand le climat se réchauffe, les espèces se déplacent vers le haut. Un faible accroissement en altitude correspond généralement à un déplacement important en latitude (Fig. 6.4). Par exemple, un refroidissement de 3°C associé à une augmentation d'altitude de 500 m correspond à un déplacement de 250 km en latitude. Du fait que les sommets des montagnes sont plus étroits que leurs bases, lorsque des espèces se déplacent vers le haut à la suite d'un réchauffement, elles occupent généralement des zones plus petites, ont des populations plus réduites et peuvent devenir plus vulnérables aux pressions génétiques et environnementales (Peters, 1990; Sombroek, 1990). Cela pourrait affecter la distribution et l'abondance des espèces endémiques dont les aires de répartition naturelle sont déjà confinées aux hautes altitudes dans les écosystèmes aussi bien tempérés que tropicaux (par exemple, les espèces endémiques des forêts de montagne en haute altitude dans les régions tropicales ou les îlôts de forêts boréales de haute altitude que l'on trouve au sud des Appalaches aux Etats-Unis).
Une étude récente menée par une équipe de scientifiques travaillant dans les Alpes autrichiennes indique que certaines espèces de plantes alpines se déplacent vers les sommets montagneux suivant un rythme allant de moins d'un mètre à près de quatre mètres par an. Ces données proviennent d'études réalisées en 1992 sur 26 sommets montagneux et comparées avec des inventaires historiques des espèces de plantes alpines sur les mêmes montagnes il y a 70-90 ans. Les Alpes centrales se sont réchauffées de 0,7°C durant cette même période (Grabherr et al, 1994).
L'élévation du niveau des mers, associée à des hausses de température, pourrait influer sur la répartition et l'abondance des forêts de mangroves. Ces forêts côtières fournissent d'abondants produits forestiers ligneux et non ligneux et des services associés. Outre qu'elles répondent aux besoins en produits ligneux des habitants des zones littorales dans les régions tropicales, elles offrent un riche habitat pour la pêche et l'aquaculture. Elles protègent également les zones littorales des tempêtes tropicales et de l'érosion et offrent des sites de reproduction à un grand nombre d'animaux sauvages (FAO, 1994; Gable et al, 1990).
Des déplacements futurs des aires naturelles des arbres et des formations forestières pourraient avoir des effets à la fois positifs et négatifs sur l'approvisionnement en bois d'oeuvre et autres produits forestiers, la distance jusqu'aux marchés, la diversité des espèces et leur sensibilité aux feux, aux ravageurs et aux maladies.
Figure 6.4 - Exemples de redistribution des essences dans les régions de hautes montagnes dans l'hypothèse d'une hausse de 2°C de la température moyenne annuelle: a = montagnes d'Afrique de l'Est: accroissement relativement modéré de la surface occupée et b = hautes terres de l'Ouganda: quasi-disparition de la zone de végétation de haute altitude (Source: Sombroek: 1990).
|
Encadré 6.1 Les forêts du monde seront-elles plus sèches dans l'avenir? Les taux de transpiration des plantes pourraient baisser dans une atmosphère enrichie en CO2. Le résultat pourrait être un monde avec une formation nuageuse réduite et moins de pluies, selon un rapport de la Terrestrial Initiative on Global Environmental Research (TIGER) du Natural Environmental Research Council du Royaume-Uni. Plusieurs groupes de chercheurs ont réussi à relier des modèles informatiques d'évolution des surfaces des terres à des modèles de prévisions climatiques. Un modèle climatique simple a prévu un accroissement de près de 10% du taux d'évaporation et de 3% des précipitations sur tes forêts tropicales ombrophiles dans un milieu ambiant à haute teneur en CO2. Toutefois, avec une description plus réaliste de la forêt ombrophile, une nouvelle séquence d'événements fut déclenchée, conduisant à une moindre quantité d'eau disponible pour la formation des nuages et une évaporation et une pluviométrie réduites (OMM, 1994) |
Rien ne permet d'affirmer que des espèces animales ou végétales puissent disparaître à cause du changement climatique. Du fait de leur mobilité, les animaux courent en général moins de risques car ils sont capables de se disperser vers des habitats plus favorables. Quant aux végétaux, ils sont fixes, et doivent compter sur la dispersion des semences à partir de zones qui ne sont plus favorables vers de nouvelles zones, d'où un déplacement progressif de leurs aires naturelles. Durant les périodes glaciaires du Pléistocène, de nombreuses essences forestières ont disparu des forêts boréales et tempérées de l'Europe car elles étaient incapables de déplacer leurs aires vers le sud en raison de la présence des Alpes, des Pyrénées et d'autres chaînes de montagnes orientées principalement est-ouest, constituant des obstacles naturels à la migration des végétaux. Par conséquent, les forêts d'Europe du Nord contiennent beaucoup moins d'espèces que celles que l'on trouve à des latitudes équivalentes en Asie et en Amérique du Nord.
En général, les phytoécologues estiment que les espèces végétales ayant de vastes aires de répartition géographique et de nombreuses populations ont le plus de chances de survivre au changement climatique. Citons à titre d'exemple Pinus sylvestris qui est présent de l'Europe de l'Ouest à la Sibérie, Populus tremula et P. tremuloides, qui ont tous les deux des aires de répartition transcontinentales. Les espèces rares ou occupant des zones géographiques limitées risqueraient davantage de disparaître. Cela est particulièrement vrai pour les espèces vivant exclusivement en haute altitude qui, en définitive, seraient incapables de déplacer leurs aires de répartition plus haut en cas de réchauffement du climat. C'est le cas du sapin de Fraser, Abies fraseri, dont l'aire de répartition naturelle est limitée aux plus hautes altitudes dans six zones du Sud des Appalaches aux Etats-Unis (Fig. 6.5). Une autre catégorie de plantes menacés d'extinction sont celles à graines lourdes, donc à dispersion difficile.
Selon d'autres écologues, le risque d'extinction d'une espèce végétale et la perte consécutive de diversité biologique est minime car les plantes possèdent une variabilité génétique leur permettant de s'adapter aux conditions changeantes de l'environnement. La variation génétique est une condition préalable nécessaire à l'évolution et un mécanisme puissant permettant aux animaux et aux plantes de changer et de s'adapter (Eriksson et al, 1993).
De même que la structure, la composition et la biomasse des forêts réagissent au changement climatique, le comportement des feux sera lui aussi modifié (Fosberg et al, 1990). On prévoit en particulier une augmentation de la fréquence et de l'intensité des feux et un allongement de la saison des feux dans les zones déjà sujettes à ce phénomène.
Plusieurs forêts tropicales ombrophiles sont périodiquement victimes de sécheresses prolongées comme celles causées par l'oscillation australe El Niño (ENSO). Ces sécheresses peuvent changer radicalement les conditions des combustibles et l'inflammabilité de la végétation. Dès que les précipitations sont inférieures à 100 mm par mois, et qu'il ne pleut pas pendant deux semaines ou plus, la végétation forestière perd progressivement ses feuilles à mesure que la sécheresse s'intensifie. En outre, la teneur en humidité des combustibles de surface diminue, alors que le matériel ligneux tombé au sol et la litière de feuilles peu compacte contribuent au déclenchement et à la diffusion des feux de surface. Les combustibles aériens comme les plantes grimpantes et les lianes desséchées deviennent de véritables échelles propageant le feu jusqu'aux cimes (Goldammer et Seibert, 1990). Cet enchaînement d'événements favorise des incendies dévastateurs comme ceux qui ont eu lieu à Kalimantan Est, en Indonésie, en 1982-83 et qui ont entraîné la destruction de plus de 3,5 millions d'ha de forêts humides primaires et secondaires. Certains modèles de circulation générale (MCG) prévoient des vagues de sécheresse accrues dans certaines forêts tropicales. En conséquence, la fréquence des grands feux incontrôlés comme ceux intervenus à Kalimantan Est pourrait augmenter.
Certaines forêts tropicales ombrophiles, surtout celles des régions tropicales subéquatoriales (de 10 à 23° de latitude) sont sujettes aux ouragans. Les dégâts provoqués par ces tempêtes favorisent l'invasion de lianes qui peuvent contribuer à l'accumulation d'une biomasse foliaire dans les fissures du sol, particulièrement durant les vagues de sécheresse survenant de temps à autre. Cela entraîne une accumulation de combustible pouvant déclencher des feux (Mueller-Dombois et Goldammer, 1990).
L'accumulation de combustibles résultant des effets directs des tempêtes tropicales peut également augmenter les risques d'incendie. En 1988, l'ouragan Gilbert a balayé une partie de la Péninsule du Yucatan au Mexique et endommagé plus d'un million d'hectares de forêts tropicales. Le volume de combustible créé par les débris a augmenté le risque de feux incontrôlés. L'année suivante, plus de 120 000 ha de la plus grande surface de forêts tropicales du Mexique ont brûlé (Ciesla, 1993). Un des effets plus incertains du changement de climat prévu est la possibilité d'une fréquence et d'une intensité accrues des tempêtes tropicales. Cela renforcerait l'accumulation de combustibles.
Dans certaines zones forestières reculées, c'est très souvent la foudre qui provoque des incendies. Une étude a été effectuée à l'aide de plusieurs MCG en vue de déterminer la fréquence de la foudre pour un régime climatique deux fois plus riche en CO2 Cette étude prévoit un accroissement de la fréquence de la foudre à toutes les latitudes avec une augmentation globale moyenne de 26% (Fosberg et al, 1990).
Une augmentation des pertes dues aux insectes et aux maladies dans les forêts pourrait être l'un des premiers effets apparents du changement climatique. Preuve en sont les épidémies liées aux ravageurs résultant du stress provoqué par les sécheresses périodiques ou les pluies excessives. Des études menées par Kristiansen (1993) et Sauerbeck (1992) sur les effets potentiels du changement climatique sur les ravageurs et les maladies en agriculture fournissent un cadre de référence pour l'identification des effets potentiels sur le secteur forestier. Ceux-ci peuvent être aussi bien positifs que négatifs.
Les effets négatifs prévus sur la santé des forêts comprennent les aspects suivants:
* En un emplacement donné, une hausse des températures pourrait se traduire par un nombre supérieur de générations d'insectes nuisibles par an, augmentant ainsi leur potentiel destructeur. Ceci est particulièrement vrai pour les insectes qui ont déjà plus d'une génération par an. Un exemple d'insecte destructeur des forêts tropicales dont le nombre de générations pourrait augmenter dans l'hypothèse d'un réchauffement est la chenille du pin, Dendrolimus punctatus, important défoliateur des pins tropicaux en Chine méridionale et en Asie du Sud-Est (Fig. 6.6).* Les ratios des espèces de ravageurs à leurs ennemis naturels pourraient changer en faveur des ravageurs, avec une augmentation de leur capacité de reproduction entraînant des dégâts plus importants.
* Des anomalies climatiques plus marquées sont prévues dans le cadre du changement du climat. Une plus grande fréquence des sécheresses, des orages, du gel intense ou des périodes de pluviosité excessive induira un stress supplémentaire sur les arbres et les forêts, les rendant plus sensibles aux attaques des ravageurs et des maladies. Les anomalies climatiques peuvent également augmenter la vulnérabilité des arbres à la pollution atmosphérique d'origine anthropique. Les cas de dépérissement de la forêt pourraient aussi devenir plus fréquents. Ceux-ci résultent d'une interaction complexe des forêts avec le climat, le site, les ravageurs et les maladies, et, dans de nombreux cas, les activités humaines (Mueller-Dombois, 1992). Citons à titre d'exemple le dépérissement de Metrosideros polymorpha dans les îles Hawaï, aux USA, de Azadirachta indica dans la région sahélienne en Afrique, de Acacia nilotica au Soudan et de Eucalyptus spp. en Australie et en Amérique du Sud (Ciesla et Donaubauer, 1994).* Des rapports carbone/azote plus élevés dans les arbres dûs à des niveaux supérieurs de CO2 pourraient augmenter la consommation de feuillage par les insectes comme l'ont montré des études en laboratoire. Lincoln et al (1984) ont montré que les quantités d'aliments ingérés par les larves des lépidoptères augmentaient avec des accroissements correspondants du CO2 atmosphérique. Des défoliateurs des forêts comme les tordeuses du sapin, Choristoneura spp. en Amérique du Nord ou les chenilles du pin, Dendrolimus spp. en Asie pourraient être affectés de la même façon. En conséquence, les attaques pourraient causer une défoliation plus grave. Plus récemment, Lincoln (1993) a montré des réactions semblables chez les larves d'un autre groupe d'insectes phyllophages, les lophyres du pin, Neodiprion sp. (Hyménoptères: Diprionidae).
* Une fréquence plus élevée des attaques d'insectes et de maladies due au stress causé aux arbres par le changement climatique entraînerait un accroissement des volumes de combustibles accumulés dans les forêts, augmentant le risque de feux incontrôlés. Dans les forêts de conifères de l'ouest de l'Amérique du Nord, les attaques récentes de plusieurs espèces de scolytes (famille des Scolytidae) ont porté les volumes de combustibles inflammables à des niveaux dangereusement élevés, provoquant des feux importants de forte intensité, en particulier l'incendie du Parc national de Yellowstone aux Etats-Unis en 1988. Bien que l'on estime que ces attaques sont liées à l'élimination des incendies et non au changement climatique, elles illustrent les conséquences possibles du changement climatique prévu (Hessburg et al, 1994, Département de l'agriculture des Etats-Unis, 1994).
Au nombre des effets positifs possibles, il faut signaler:
* Les taux de croissance plus élevés prévus par certains scientifiques en raison de hausses de température ou d'un accroissement des concentrations de CO2 pourraient permettre aux forêts de supporter des dégâts plus importants causés par les insectes et les maladies sans que leur croissance ou leur rendement ne soient réduits.* La vigueur accrue des arbres et des forets poussant dans un milieu à forte concentration de CO2 pourrait les rendre plus résistants aux attaques des insectes et des maladies.
* Un taux élevé de CO2 pourrait être bénéfique à la santé et à la productivité des plantes en modifiant leur morphologie et leur physiologie au détriment des organismes pathogènes.
On pense que le risque d'attaques dévastatrices par les insectes et les maladies dans les forêts tropicales est minime en comparaison aux forêts tempérées et boréales en raison de leur diversité intrinsèque. Si cela peut être vrai dans les forêts tropicales d'origine naturelle, il faut tenir compte du fait que de nombreux pays tropicaux, pour couvrir leurs besoins de produits ligneux, dépendent de plantations monospécifiques, souvent de variétés exotiques à croissance rapide. Beaucoup de ces plantations sont établies avec un matériel à base génétique étroite, souvent incapable de s'adapter à des conditions changeantes de l'environnement. En 1990, on estimait à 30,7 millions d'ha les plantations forestières dans 90 pays tropicaux (FAO, 1993). Un total de 23% était composé d'Eucalyptus spp. et 10% de diverses essences de Pinus. Celles-ci sont sujettes aux attaques de ravageurs très divers, dont beaucoup introduits par accident. On doit à Hutacharern et al (1990) une excellente étude des insectes et des maladies des forêts et plantations de la région Asie-Pacifique. Cette étude indique clairement que dans les forêts aménagées des régions tropicales, un grand nombre d'insectes et de ravageurs pourraient réagir au changement climatique.
|
Encadré 6.2. Le dépérissement de Juniperus procera au Kenya - un exemple des effets d'un changement climatique régional? Dès le début des années 80, on a observé de nombreux cas de dépérissement et de mortalité de Juniperus procera, composante importante des forêts d'altitude au Kenya. En certains endroits, jusqu'à 90% des arbres ont été touchés. C'est dans les forêts plus sèches de basse altitude que sont enregistrés les taux de dépérissement et de mortalité les plus importants. Les peuplements de haute altitude, qui reçoivent plus de pluies et se développent sur de meilleurs sols, semblent être dans un état de santé satisfaisant. On ignore la cause de cette situation. Une hypothèse serait que ces forêts ont souffert d'une tendance au réchauffement et au dessèchement à long terme au niveau régional, qui a affecté les sites de basse altitude au point de les rendre impropres à la croissance de cette essence (Cisela et al, 1994). En conséquence, l'aire de répartition en altitude de cette essence pourrait se réduire dans l'avenir. |
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}