Autolysis means "self-digestion". It has been known for many years that there are at least two types of fish spoilage: bacterial and enzymatic. Uchyama and Ehira (1974) showed that for cod and yellowtail tuna, enzymatic changes related to fish freshness preceded and were unrelated to changes in the microbiological quality. In some species (squid, herring), the enzymatic changes precede and therefore predominate the spoilage of chilled fish. In others, autolysis contributes to varying degrees to the overall quality loss in addition to microbially- mediated processes.
Production of energy in post mortem muscle
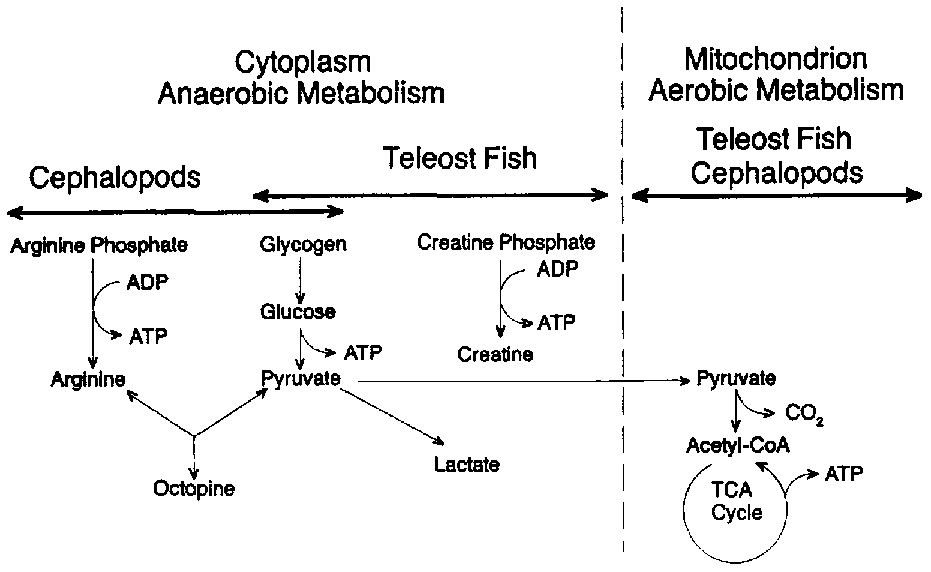
At the point of death, the supply of oxygen to the muscle tissue is interrupted because the blood is no longer pumped by the heart and is not circulated through the gills where, in the living fish, it becomes enriched with oxygen. Since no oxygen is available for normal respiration, the production of energy from ingested nutrients is greatly restricted. Figure 5.2 illustrates the normal pathway for the production of muscle energy in most living teleost fish (bony finfish). Glycogen (stored carbohydrate) or fat is oxidized or "burned" by the tissue enzymes in a series of reactions which ultimately produce carbon dioxide (CO2), water and the energy-rich organic compound adenosine triphosphate (ATP). This type of respiration takes place in two stages: an anaerobic and an aerobic stage. The latter depends on the continued presence of oxygen (02) which is only available from the circulatory system. Most crustaceans are capable of respiring outside the aquatic environment by absorption of atmospheric oxygen for limited periods.
Figure 5.2 Aerobic and anaerobic breakdown of glycogen in fish muscle
Figure 5.2 also illustrates that, under anaerobic conditions, ATP may be synthesized by two other important pathways from creatine phosphate or from arginine phosphate. The former source of energy is restricted to vertebrate muscle (teleost fish) while the latter is characteristic of some invertebrates such as the cephalopods (squid and octopus). In either case, ATP production ceases when the creatine or arginine phosphates are depleted. It is interesting to note that octopine is the end-product from the anaerobic metabolism of cephalopods and is not acidic (unlike lactate), thus any changes in post mortem pH in such animals are not related to the production of lactic acid from glycogen.
For most teleost fish, glycolysis is the only possible pathway for the production of energy once the heart stops beating. This more inefficient process has principally lactic and pyruvic acids as its end-products. In addition, ATP is produced in glycolysis, but only 2 moles for each mole of glucose oxidized as compared to 36 moles ATP produced for each mole of glucose if the glycolytic end products are oxidized aerobically in the mitochondrion in the living animal. Thus, after death, the anaerobic muscle cannot maintain its normal level of ATP, and when the intracellular level declines from 7-10 moles/g to < 1.0 mmoles/g tissue, the muscle enters rigor mortis. Post mortem glycolysis results in the accumulation of lactic acid which in turn lowers the pH of the muscle. In cod, the pH drops from 6.8 to an ultimate pH of 6.1-6.5. In some species of fish, the final pH may be lower: in large mackerel, the ultimate rigor pH may be as low as 5.8-6.0 and as low as 5.4-5.6 in tuna and halibut, however such low pH levels are unusual in marine teleosts. These pHs are seldom as low as those observed for post mortem mammalian muscle. For example, beef muscle often drops to pH levels of 5.1 in rigor mortis. The amount of lactic acid produced is related to the amount of stored carbohydrate (glycogen) in the living tissue. In general, fish muscle contains a relatively low level of glycogen compared to mammals, thus far less lactic acid is generated after death. Also, the nutritional status of the fish and the amount of stress and exercise encountered before death will have a dramatic effect on the levels of stored glycogen and consequently on the ultimate post mortem pH. As a rule, well- rested, well-fed fish contain more glycogen than exhausted fish. In a recent study of Japanese loach (Chiba et al., 1991), it was shown that only minutes of pre-capture stress resulted in a decrease of 0.50 pH units in 3 hours as compared to non-struggling fish whose pH dropped only 0.10 units in the same time period. In addition, the same authors showed that bleeding of fish significantly reduced the post mortem production of lactic acid.
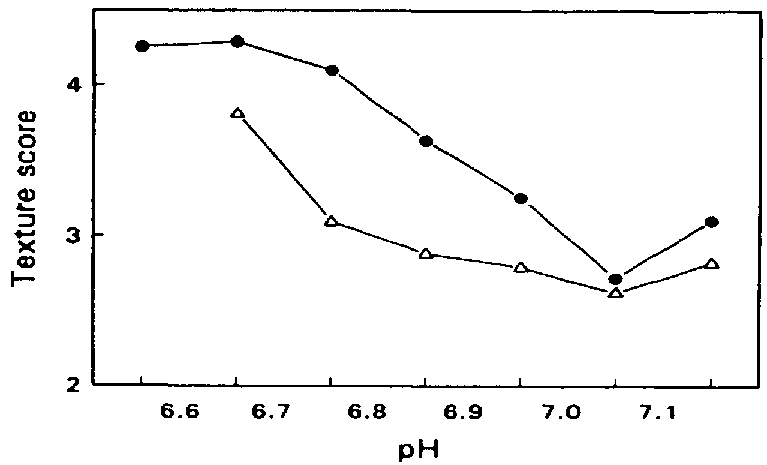
The post mortem reduction in the pH of fish muscle has an effect on the physical properties of the muscle. As the pH drops, the net surface charge on the muscle proteins is reduced, causing them to partially denature and lose some of their water- holding capacity. Muscle tissue in the state of rigor mortis loses its moisture when cooked and is particularly unsuitable for further processing which involves heating, since heat denaturation enhances the water loss. Loss of water has a detrimental effect on the texture of fish muscle and it has been shown by Love (1975) that there is an inverse relationship between muscle toughness and pH, unacceptable levels of toughness (and water-loss on cooking) occurring at lower pH levels (Figure 5.3).
Autotysis and nucleotide catabolism
As mentioned earlier, rigor mortis sets in when the muscle ATP level drops to < 1.0 mmoles/g. ATP is not only a source of high energy which is required for muscle contraction in the living animal, but also acts as a muscle plasticizer. Muscle contraction per se is controlled by calcium and an enzyme, ATP- ase which is found in every muscle cell. When intracellular Ca+2 levels are > 1 mM, Ca+2-activated ATP-ase reduces the amount of free muscle ATP which results in the interaction between the major contractile proteins, actin and myosin. This ultimately results in the shortening of the muscle, making it stiff and inextensible. A fish in rigor mortis cannot normally be filleted or processed because the carcass is too stiff to be manipulated and is often contorted, making machine-handling impossible (see also section 3.2 on bleeding and section 5.1 on sensory changes).
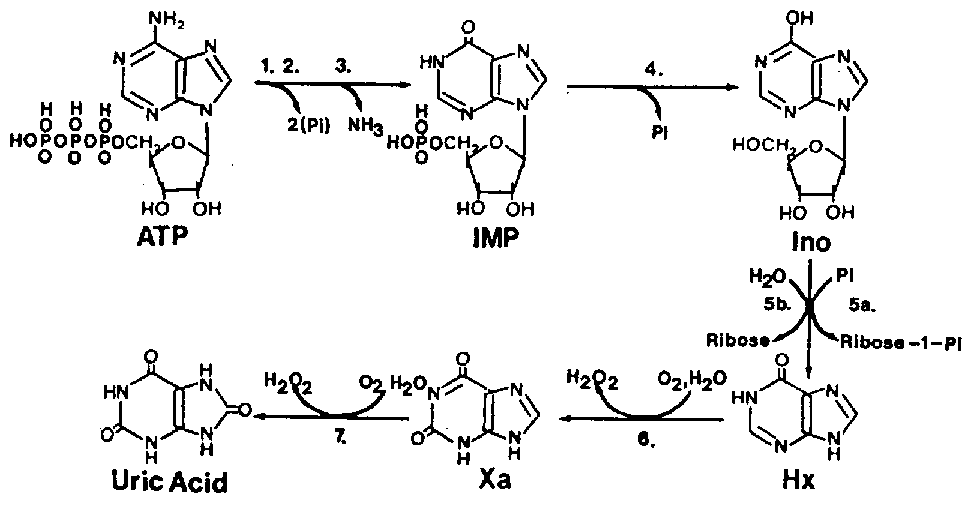
The resolution of rigor is a process still not completely understood but always results in the subsequent softening (relaxation) of the muscle tissue and is thought to be related to the activation of one or more of the naturally-occurring muscle enzymes, digesting away certain components of the rigor mortis complex. The softening of the muscle during resolution of rigor (and eventually spoilage processes) is coincidental with the autolytic changes. Among the changes, one of the first to be recognized was the degradation of ATP-related compounds in a more-or-less predictable manner after death. Figure 5.4 illustrates the degradation of ATP to form adenosine diphosphate (ADP), adenosine monophosphate (AMP), inosine monophosphate (IMP), inosine (Ino) and hypoxanthine (Hx). The degradation of ATP catabolites proceeds in the same manner with most fish but the speed of each individual reaction (from one catabolite to another) greatly varies from one species to another and often progresses coincidentally with the perceived level of spoilage as determined by trained analysts. Saito et al. (1959) were the first to observe this pattern and to develop a formula for fish freshness based on these autolytic changes:
Kayo ={[Ino] + [Hx]\ [ATP] + [ADP] + [AMP] + [IMP] + [Ino] + [Hx]} x 100
- where [ATP], [ADP], [AMP], [IMP], [Ino] and [Hx] represent the relative concentrations of these compounds in fish muscle measured at various times during chilled storage.
The K or "freshness" index gives a relative freshness rating based primarily on the autolytic changes which take place during post mortem storage of the muscle. Thus, the higher the K value, the lower the freshness level. Unfortunately, some fish species such as Atlantic cod reach a maximum K value well in advance of the shelf life as determined by trained judges, and K is therefore not considered reliable as a freshness index for all marine finfish. Also, the degradation of nucleotide catabolites is only coincidental with perceived changes in freshness and not necessarily related to the cause of freshness deterioration since only Hx is considered to have a direct effect on the perceived bitter off-flavour of spoiled fish (Hughes and Jones, 1966). It is now widely accepted that IMP is responsible for the desirable fresh fish flavour which is only present in top quality seafood. None of the nucleotide catabolites are considered to be related to the perceived changes in texture during the autolytic process except of course ATP whose loss is associated with rigor mortis.
Figure
5.4 Post mortem ATP degradation in
fish muscle. Enzymes include: 1. ATP-ase; 2. myokinase; 3. AMP
deaminase; 4. IMP phosphohydrolase; 5a. nucleoside phosphorylase;
5b. inosine nucleosidase; 6,7. xanthine oxidase.
Source: Gill (1992)
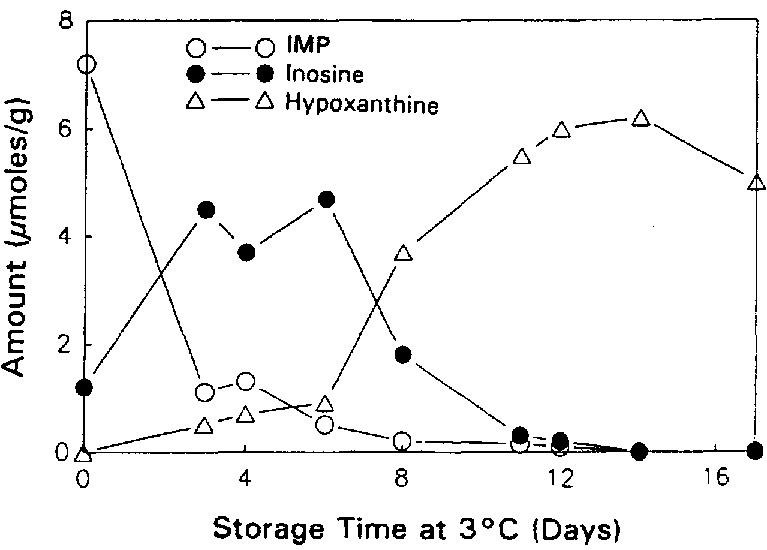
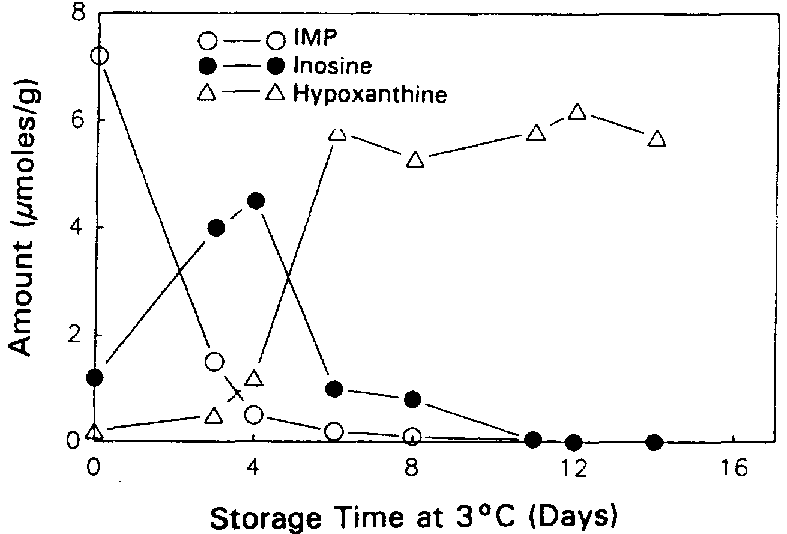
Surette et al. (1988) followed the autolysis of sterile and non-sterile cod as indicated by the ATP catabolites. The rates of formation and breakdown of IMP were the same in both sterile and non-sterile samples of cod tissue (Figures 5.5a and 5.5b), indicating that the catabolic pathway for the degradation of ATP through to inosine is entirely due to autolytic enzymes.
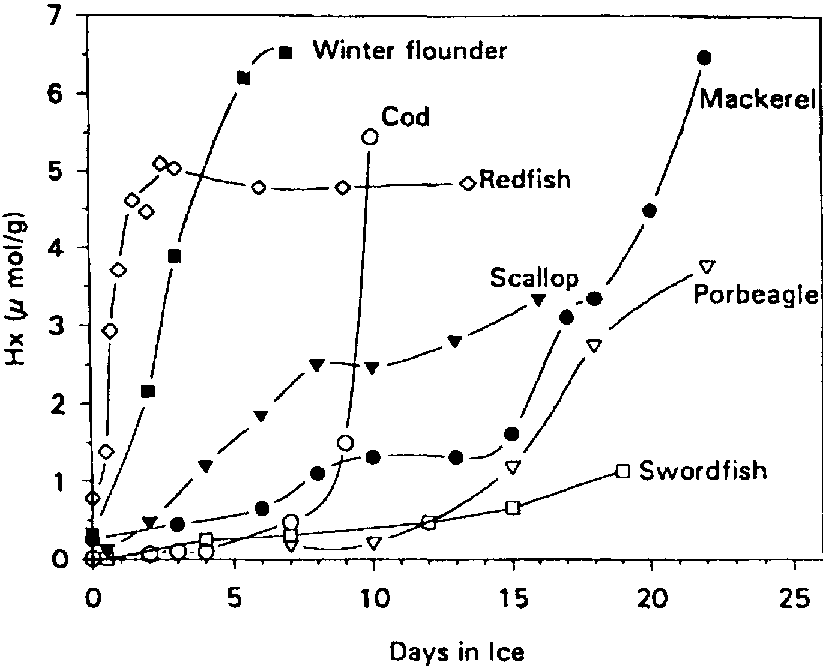
The conversion of Ino to Hx was accelerated by about 2 days for the non-sterile samples, suggesting that bacterial nucleoside phosphorylase (enzyme 5.a in Figure 5.4) plays a major role in the post mortem production of Hx in refrigerated cod (see also section 5.3). It is interesting to note that Surette et al. (1988) were not able to recover nucleoside phosphorylase from freshly killed cod, but Surette et al. (1990) later went on to isolate and purify this enzyme from a Proteus bacterium recovered from spoiled cod fillets. As mentioned earlier, large variations can be expected in the patterns of nucleotide degradation from one species to another. The variations in Hx among various types of fish are shown in Figure 5.6. It is clear therefore that Hx determination would likely not be useful for such species as swordfish and redfish.
Figure 5.5a Changes in IMP, Ino and Hx in sterile cod fillets at 3°C, adapted from Gill (1990)
Figure 5.5b Changes in IMP, Ino and Hx in non-sterile cod fillets at 3°C, adapted from Gill (1990)
There is little doubt that physical handling accelerates the
autolytic changes in chilled fish.
Surette et al. (1988) reported that the breakdown rate of the
nucleotide catabolites was greater in sterile fillets than in
non-sterile gutted whole cod. This is perhaps not surprising
since many of the autolytic enzymes have been shown to be
compartmentalized in discrete membrane-bound packages which
become broken when subjected to physical abuse and result in the
intimate mixing of enzyme and substrate. Crushing of the fish by
ice or other fish can seriously affect the edibility and
filleting yields even for fish which have a relatively low
bacterial load, demonstrating the importance of autolytic
processes. Iced fish should never be stored in boxes deeper than
30 cm and it is equally important to be sure that fish boxes are
not permitted to "nest" one on top of the other if
autolysis is to be minimized. Systems for conveying fish and for
discharge from the vessels must be designed so as to avoid
physical damage to the delicate tissues.
Several rapid methods have been developed for the determination of individual nucleotide catabolites or combinations including the freshness index. Two recent reviews should be consulted (Gill, 1990, 1992).
Autolytic changes involving proteolytic enzymes
Many proteases have been isolated from fish muscle and the effects of proteolytic breakdown are often related to extensive softening of the tissue. Perhaps one of the most notable examples of autolytic proteolysis is the incidence of belly-bursting in pelagic (fatty fish) species such as herring and capelin. This type of tissue softening is most predominant in summer months when pelagics are feeding heavily, particularly on "red feed" consisting of copepods and euphausiids. The low molecular weight peptides and free amino-acids produced by the autolysis of proteins not only lower the commercial acceptability of pelagics, but in bulk-stored capelin, autolysis has been shown to accelerate the growth of spoilage bacteria by providing a superior growth environment for such organisms (Aksnes and Brekken, 1988). The induction of bacterial spoilage in capelin by autolysis also resulted in the decarboxylation of amino-acids, producing biogenic amines and lowered the nutritive value of the fish significantly. This is particularly important since autolysis and bacterial growth greatly lower the commercial value of pelagics used for the production of fishmeal.
Similarly, bulk-stored herring used for fishmeal has been found to contain carboxy- peptidases A and B. chymotrypsin, and trypsin; and preliminary studies have shown that proteolysis can be inhibited by the addition of potato extracts which not only slowed the proteolysis but resulted in lower microbial growth and preservation of the nutritional value of the meal (Aksnes, 1989).
More recently, Botta et al. (1992) found that autolysis of the visceral cavity (belly-bursting) of herring was related more to physical handling practices than to biological factors such as fish size, amount of red feed in the gut or roe content. In particular, it was shown that for herring, freezing/thawing, thawing time at 15°C and time of iced storage, had a far greater influence on belly-bursting than biological factors.
Cathepsins
Although several proteolytic enzymes have been discovered in the fish tissues, it has perhaps been the cathepsins which have been described most often. The cathepsins are "acid" proteases usually found packaged in tiny, submicroscopic organelles called Iysozomes. In living tissue, Iysozomal proteases are believed to be responsible for protein breakdown at sites of injury. Thus cathepsins are for the most part inactive in living tissue but become released into the cell juices upon physical abuse or upon freezing and thawing of post mortem muscle.
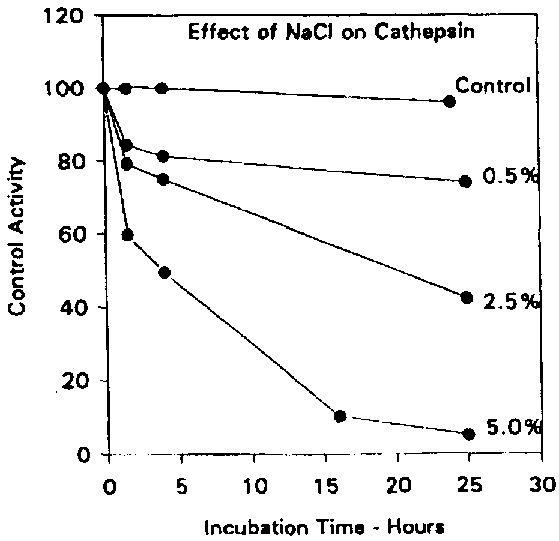
Cathepsins D and L are believed to play a major role in the autolytic degradation of fish tissue since most of the other cathepsins have a relatively narrow pH range of activity far too low to be of physiological significance. Reddi et al. (1972) demonstrated that an enzyme believed to be cathepsin D from winter flounder was active over a pH range of 3-8 with a maximum near pH 4.0, although no attempt was made to confirm the identity of the enzyme using synthetic substrates or specific inhibitors. Nevertheless, the enzyme was far less active in the presence of ATP, suggesting that such an enzyme would only be active in post mortem fish muscle. Also, the enzyme activity was inhibited strongly by the presence of salt (Figure 5.7) with virtually no activity remaining after a 25-hour incubation in the presence of 5% sodium chloride. It is therefore unlikely that Reddi's enzyme was active in salted fish products.
Cathepsin L has been implicated in the softening of salmon muscle during spawning migration. It is likely that this enzyme contributes more to autolysis of fish muscle than cathepsin D since it is far more active at neutral pH, and has been shown to digest both myofibrillar proteins (actomyosin) as well as connective tissue. Yamashita and Konogaya (1990) produced strong evidence implicating cathepsin L rather than other cathepsins in the softening of salmon during spawning. They demonstrated that electrophoresis of purified myofibrils treated with cathepsin L resulted in patterns which were almost identical to patterns of proteins recovered from muscle from spawning fish.
Furthermore, the cathepsin L autolytic activity correlated well with the texture of the muscle as measured instrumentally. The linear correlation between cathepsin L activity and breaking strength of the muscle was excellent; r = 0.86 and -0.95 for fresh and frozen/thawed tissue, respectively. It is interesting that, in all cases, the autolytic ability as measured by cathepsin L activity was higher in frozen/thawed tissue than in fresh tissue. Freezing and thawing often break down cell membranes allowing autolytic membrane-bound enzymes to react with their natural substrates. The enzyme and its naturally occurring inhibitor were further studied by the same authors (Yamashita and Konogaya, 1992). Cathepsin L has also been associated with the production of a jelly- like softening of flounder (Toyohara et al., 1993 a) and the uncontrollable softening of Pacific hake muscle which has been parasitized by Myxosporidia (Toyohara et al., 1993 b).
Figure 5.7 Effect of NaCl on the catheptic activity. Adapted from Reddi et al. (1972)
The tissues of such infected fish have little commercial value, but at present it is not known if it is the parasite or the host which secretes the proteolytic enzymes which autolyze the muscle.
In addition to their detrimental effect on texture, catheptic enzymes induce intentional autolytic changes in fermented fish products. For example, cathepsins are believed to be responsible for major textural changes during the fermentation of salted preserved Japanese squid and Crucian carp (Makinodan et al., 1991, 1993).
Calpains
A second group of intracellular proteases called "calpains" or "calcium activated factor" (CAF) has recently been associated with fish muscle autolysis and is found in meats, finfish and crustaceans. Tenderness is probably the most important quality characteristic of red meat. It has been known for nearly a century that post mortem aging of red meat results in the tenderization process. Calpains have been found primarily responsible for the post mortem autolysis of meat through digestion of the z-line proteins of the myofibril. Although toughness is seldom a problem with unfrozen fish muscle, softening through autolysis is a serious problem limiting the commercial value. The calpains are intra-cellular endopeptidases requiring cysteine and calcium; m-calpain requiring 5-50 mM Ca+2, m-calpain requiring 150-1000mM Ca+2. Most calpains are active at physiological pH, making it reasonable to suspect their importance in fish- softening during chilled storage.
Studies have shown that in crustacean muscle, calpains are associated with molt- induced textural changes to the muscle and carry out non-specific generalized digestion of the myofibrillar proteins. However, vertebrate muscle calpains have been shown to be very specific, digesting primarily tropinin-T, desmin, titin and nebulin, attacking neither vertebrate actin or myosin (Koohmaraie, 1992). In contrast, fish calpains digest myosin (specifically the myosin heavy chain) to form an initial fragment with approximate molecular weight of 150 000 Da (Muramoto et al., 1989). The same authors demonstrated that fish calpains were far more active at low temperatures than were mammalian calpains and that the rates of cleavage were species-specific, being most active against myosins with lowest heat stabilities. Thus, fish species adapted to colder environmental temperatures are more susceptible to calpain autolysis than those from tropical waters. Although calpain has been identified in several fish species including carp (Toyohara et al., 1985), tilapia and shrimp (Wang et al., 1993), as well as tuna, croaker, red seabream and trout (Muramoto et al., 1989) to name a few, little work has to date demonstrated a "cause and effect" relationship between calpain activity and instrumental measurements of texture.
Collagenases
To this point, all of the post mortem autolytic changes described have involved changes within the muscle cell per se. However, the flesh of teleost fish is divided into blocks of muscle cells separated into "flakes" or myotomes by connective tissue called myocommata (Figure 3.3). Each muscle cell or fibre is surrounded with connective tissue which attaches to the myocommata at the ends of the cells by means of fine collagenous fibrils. During chilled storage, these fibrils deteriorate (Bremner and Hallett, 1985). More recently, it was shown that instrumental measurements of texture of chilled trout muscle decreased as the amount of type V collagen was solubilized, presumably due to the action of autolytic collagenase enzymes (Sato et al., 1991). It is these enzymes which presumably cause "gaping" or breakdown of the myotome during long-term storage on ice or short term storage at high temperature. For Atlantic cod, it has been shown that upon reaching 17°C, gaping is inevitable presumably because of degradation of the connective tissue and rapid shortening of the muscle due to high temperature rigor.
The relatively short shelf life of chilled prawns due to softening of the tissue has also been shown to be due to the presence of collagenase enzymes (Nip et al., 1985). The source of the collagenase enzymes in prawn is thought to be the hepatopancreas (digestive organ).
Automatic changes during frozen storage
The reduction of trimethylamine oxide (TMAO), an osmoregulatory compound in many marine teleost fish, is usually due to bacterial action (section 5.3) but in some species an enzyme is present in the muscle tissue which is able to break down TMAO into dimethylamine (DMA) and formaldehyde (FA):
(CH3)3 NO (r) (CH3)2NH + HCHO
It is important to note that the amount of formaldehyde
produced is equivalent to the dimethylamine formed but is of far
greater commercial significance. Formaldehyde induces
cross-linking of the muscle proteins making the muscle tough and
readily lose its water holding capacity. The enzyme responsible
for formaldehyde-induced toughening is called TMAO-ase or TMAO
demethylase and is most commonly found in the gadoid fishes (cod
family). Most of the TMAO demethylase enzymes reported to date
were membrane-bound and become most active when the tissue
membranes are disrupted by freezing or artificially by detergent
solubilization. Dark (red) muscle has a higher rate of activity
than white muscle whereas other tissues such as kidney, spleen
and gall bladder are extremely rich in the enzyme. Thus, it is
important that minced fish is completely free of organ tissue
such as kidney from gadoid species if toughening in frozen
storage is to be avoided. It is often difficult to ensure that
the kidney is removed prior to mechanical deponing since this
particular organ runs the full length of the backbone and is
adherent to it. The TMAO-ase enzyme has been isolated from the
microsomal fraction in hake muscle (Parkin and Hultin, 1986) and
the
Iysosomal membrane in kidney tissue (Gill et al., 1992). It has
been shown that the toughening of frozen hake muscle is
correlated to the amount of formaldehyde produced, and that the
rate of FA production is greatest at high frozen-storage
temperatures (Gill et al., 1979). In addition, it has been shown
that the amount of FA- induced toughening is enhanced by physical
abuse to the catch prior to freezing and by temperature
fluctuations during frozen storage. The most practical means of
preventing the autolytic production of FA is to store fish at
temperatures <-30°C to minimize temperature fluctuations in
the cold store and to avoid rough handling or the application of
physical pressure on the fish prior to freezing. The autolytic
changes affecting the edibility of fresh and frozen fish are
summarized in Table 5.3. Generally, the most important single
factor affecting autolysis is physical disruption of the muscle
cells. No attempt has been made here to deal with the alkaline
proteases associated with the softening of cooked surimi
products. An article by Kinoshita et al. (1990) deals with the
heat-activated alkaline proteases associated with the softening
in surimi- based products.
Table 5.3 Summary of Autolytic Changes in Chilled Fish
| Enzyme(s) | Substrate | Changes Encountered | Prevention/lnhibition |
| glycolytic enzymes | Glycogen | - production of
lactic acid, pH of tissue drops, loss of water-holding capacity in muscle - high temperature rigor may result in gaping |
- fish should
be allowed to pass through rigor at temperatures as close to 0°C as practically possible - pre-rigor stress must be avoided |
| autolytic
enzymes, involved in nucleotide breakdown |
ATP ADP AMP IMP |
- loss of fresh
fish flavour, gradual production of bitternes with Hx (later stages) |
- same as above - rough handling or crushing accelerates breakdown |
| cathepsins | Proteins, peptides | - softening of
tissue making processing difficult or impossible |
- rough handling
during storage and discharge |
| chymotrypsin, trypsin, carboxy- peptidases |
Proteins, peptides | - autolysis of
visceral cavity in pelagics (belly-bursting) |
- problem
increased with freezing/thawing or long- term chill storage |
| calpain | Myofibrillar proteins | - softening,
molt-induced softening in crustaceans |
- removal of
calcium thus preventing activation? |
| collagenases | Connective tissue | -
"gaping" of fillets - softening |
- connective
tissue degradation related to time and temperature of chilled storage |
| TMAO demethylase |
TMAO | -
formaldehyde-induced toughening of frozen gadoid fish |
- store fish at
temperature < -30°C - physical abuse and freezing/thawing accelerate formaldehyde- induced toughening |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}