The bacterial flora on live fish
Microorganisms are found on all the outer surfaces (skin and gills) and in the intestines of live and newly caught fish. The total number of organisms vary enormously and Liston (1980) states a normal range of 102-107 cfu (colony forming units)/cm2 on the skin surface. The gills and the intestines both contain between 103 and 109 cfu/g (Shewan, 1962).
The bacterial flora on newly-caught fish depends on the environment in which it is caught rather than on the fish species (Shewan, 1977). Fish caught in very cold, clean waters carry the lower numbers whereas fish caught in warm waters have slightly higher counts. Very high numbers, i.e., 107 cfu/cm2 are found on fish from polluted warm waters. Many different bacterial species can be found on the fish surfaces. The bacteria on temperate water fish are all classified according to their growth temperature range as either psychrotrophs or psychrophiles. Psychrotrophs (cold-tolerant) are bacteria capable of growth at 0°C but with optimum around 25°C. Psychrophiles (cold-loving) are bacteria with maximum growth temperature around 20°C and optimum temperature at 15°C (Morita, 1975). In warmer waters, higher numbers of mesophiles can be isolated. The microflora on temperate water fish is dominated by psychrotrophic Gram-negative rod-shaped bacteria belonging to the genera Pseudomonas, Moraxella, Acinetobacter, Shewanella and Flavobacterium. Members of the Vibrionaceae (Vibrio and Photobacterium) and the Aeromonadaceae (Aeromonas spp.) are also common aquatic bacteria and typical of the fish flora (Table 5.4). Gram-positive organisms as Bacillus, Micrococcus, Clostridium, Lactobacillus and coryneforms can also be found in varying proportions, but in general, Gram-negative bacteria dominate the microflora. Shewan (1977) concluded that Gram-positive Bacillus and Micrococcus dominate on fish from tropical waters. However, this conclusion has later been challenged by several studies which have found that the microflora on tropical fish species is very similar to the flora on temperate species (Acuff et al., 1984; Gram et al., 1990; Lima dos Santos 1978; Surendran et al., 1989). A microflora consisting of Pseudomonas, Acinetobacter, Moraxella and Vibrio has been found on newly-caught fish in several Indian studies (Surendran et al., 1989). Several authors conclude, as Liston (1980), that the microflora on tropical fish often carry a slightly higher load of Gram-positives and enteric bacteria but otherwise is similar to the flora on temperate-water fish.
Aeromonas spp. are typical of freshwater fish, whereas a number of bacteria require sodium for growth and are thus typical of marine waters. These include Vibrio, Photobacterium and Shewanella. However, although Shewanella putrefaciens is characterized as sodium- requiring, strains of S. putrefaciens can also be isolated from freshwater environments (DiChristina and DeLong, 1993; Gram et al., 1990; Spanggaard et al, 1993). Although S. putrefaciens has been isolated from tropical freshwaters, it is not important in the spoilage of freshwater fish (Lima dos Santos, 1978; Gram, 1990).
Table 5.4 Bacterial flora on fish caught in clean, unpolluted waters
| Gram-negative | Gram-positive | Comments |
| Pseudomonas Moraxella Acinetobacter Shewanella putrefaciens Flavabacterium Cytophaga Vibrio Photobacterium Aeromonas |
Bacillus Clostridium Micrococcus Lactobacillus Coryneforms |
|
| Vibrio and
Photobactenum are typical of marine waters; Aeromonas is typical of freshwater |
In polluted waters, high numbers of Enterobacteriaceae may be found. In clean temperate waters, these organisms disappear rapidly, but it has been shown that Eschenchia coli and Salmonella can survive for very long periods in tropical waters and once introduced may almost become indigenous to the environment (Fujioka et al., 1988).
The taxonomy of S. putrefaciens has been rather confused. The organism was originally associated with the Achromobacter group but was later placed in the Shewan Pseudomonas group IV. Based on percentage of guanine+cytosine (GC%) it was transferred to the genus Alteromonas, but on the basis of 5SRNA homology it was reclassified to a new genus, Shewanella (MacDonnell and Colwell, 1985). It has recently been suggested that the genus Aeromonas spp. which was a member of the Vibrionaceae family be transferred to its own family, the Aeromonadaceae (Colwell et al., 1986).
Japanese studies have shown very high numbers of microorganisms in the gastrointestinal tract of fish, and as such numbers are much higher than in the surrounding water, this indicates the presence of a favourable ecological niche for the microorganisms. Similarly, Larsen et al. (1978) reported up to 107 cfu/g of vibrio-like organisms in the intestinal tract of cod and Westerdahl et al. (1991) also isolated high numbers of vibrio-like organisms from the intestines of turbot. hotobacterium phosphoreum which can be isolated from the surface can also be isolated in high numbers from the intestinal tract of some fish species (Dalgaard, 1993). On the contrary, some authors believe that the microflora of the gastrointestinal tract is merely a reflection of the environment and the food intake.
Microbial invasion
The flesh of healthy live or newly-caught fish is sterile as the immune system of the fish prevents the bacteria from growing in the flesh (Figure 5.X a). When the fish dies, the immune system collapses and bacteria are allowed to proliferate freely. On the skin surface, the bacteria to a large extent colonize the scale pockets. During storage, they invade the flesh by moving between the muscle fibres. Murray and Shewan (1979) found that only a very limited number of bacteria invaded the flesh during iced storage. Ruskol and Bendsen (1992) showed that bacteria can be detected by microscope in the flesh when the number of organisms on the skin surface increases above 106 cfu/cm² (Figure 5.6 b). This was seen at both iced and ambient temperatures. No difference was found in the invasive patterns of specific spoilage bacteria (e.g., S. putrefaciens) and non-spoilage bacteria.
Since only a limited number of organisms actually invade the flesh and microbial growth mainly takes place at the surface, spoilage is probably to a large extent a consequence of bacterial enzymes diffusing into the flesh and nutrients diffusing to the outside.
Fish spoil at very different rates (see also section 6.5), and differences in surface properties of fish have been proposed to explain this. Skins of fish have very different textures. Thus whiting (Merlangius merlangus) and cod (Gadus morhua) which have a very fragile integument spoil rapidly compared to several flatfish such as plaice that has a very robust dermis and epidermis. Furthermore, the latter group has a very thick slime layer, which includes several antibacterial components, such as antibodies, complement and bacteriolytic enzymes (Murray and Fletcher, 1976; Hjelmland et al., 1983).
Changes in the microflora during storage and spoilage/Specific spoilage organisms
Bacteria on fish caught in temperate waters will-enter the exponential growth phase almost immediately after the fish have died. This is also true when the fish are iced, probably because the microflora is already adapted to the chill temperatures. During ice storage, the bacteria will grow with a doubling time of approximately 1 day and will, after 2-3 weeks, reach numbers of 108-109 cfu/g flesh or cm² skin. During ambient storage, a slightly lower level of 107-108 cfu/g is reached in 24 hours. The bacteria on fish caught in tropical waters will often pass through a lag-phase of 1-2 weeks if the fish are stored in ice, whereafter exponential growth begins. At spoilage, the bacterial level on tropical fish is similar to the levels found on temperate fish species (Gram, 1990; Gram et al., 1990).
If iced fish are stored under anaerobic conditions or if stored in CO2 containing atmosphere, the number of the normal psychrotrophic bacteria such as S. putrefaciens and Pseudomonas is often much lower, i.e., 106-107 cfu/g than on the aerobically stored fish. However, the level of bacteria of psychrophilic character such as P. phosphoreum reaches a level of 107-108 cfu/g when the fish spoil (Dalgaard et al., 1993).
The composition of the microflora also changes quite dramatically during storage. Thus, under aerobic iced storage, the flora is composed almost exclusively of Pseudomonas spp. and S. putrefaciens after 1-2 weeks. This is believed to be due to their relatively short generation time at chill temperatures (Morita, 1975; Devaraju and Setty, 1985) and is true for all studies carried out whether on tropical or temperate-water fish. At ambient temperature (25°C), the microflora at the point of spoilage is dominated by mesophilic Vibrionaceae and, particularly if the fish are caught in polluted waters, Enterobacteriaceae.
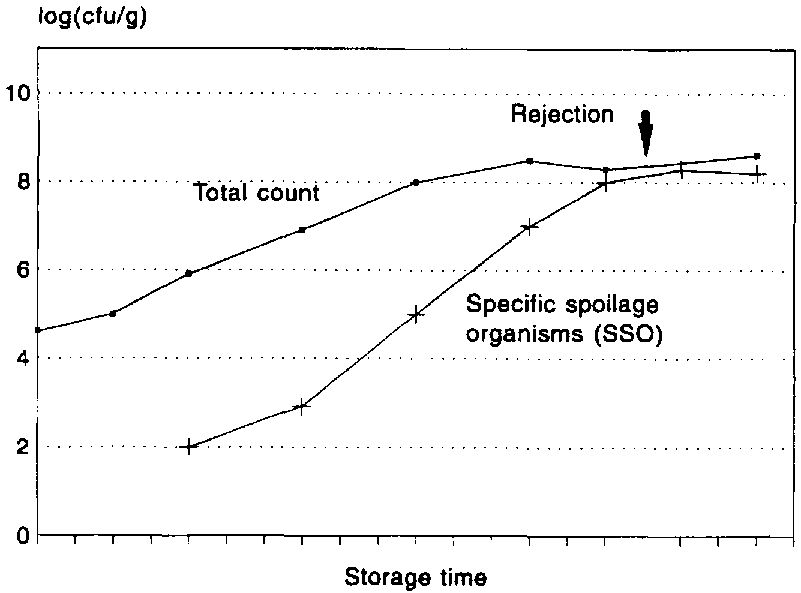
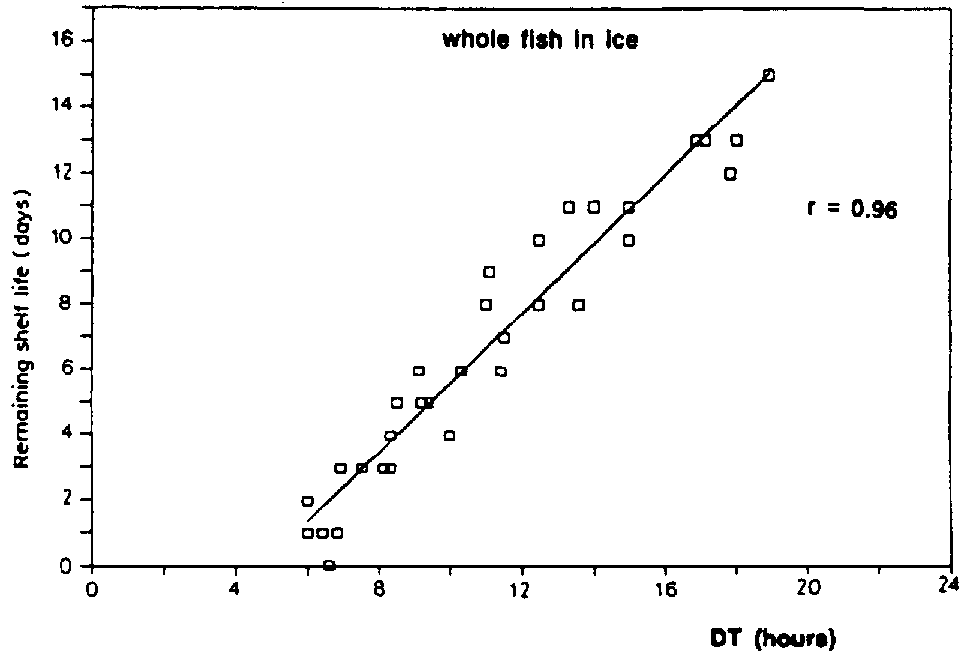
A clear distinction should be made between the terms spoilage flora and spoilage bacteria since the first describes merely the bacteria present on the fish when it spoils whereas the latter is the specific group that produce the off-odours and off-flavours associated with spoilage. A large part of the bacteria present on the spoiled fish have played no role whatever in the spoilage (Figure 5.9). Each fish product will have its own specific spoilage bacteria and the number of these will, as opposed to the total number, be related to the shelf life. In Figure 5.10, it is shown that the remaining shelf life of iced cod can be predicted from the conductometric detection time (in TMAO broth), which is inversely correlated with the number of hydrogen sulphide-producing bacteria.
It is not an easy task to determine which of the bacteria isolated from the spoiled fish are those causing spoilage, and it requires extensive sensory, microbiological and chemical studies. First, the sensory, microbiological and chemical changes during storage must be studied and quantified, including a determination of the level of a given chemical compound that correlates with spoilage (the chemical spoilage indicator). Second, bacteria are isolated at the point of sensory rejection. Pure and mixed cultures of bacteria are screened in sterile fish substrates for their spoilage potential, i.e., their ability to produce sensory (off-odours) and chemical changes typical of the spoiling product. Finally, the selected strains are tested to evaluate their spoilage activity, i.e., if their growth rate and their qualitative and quantitative production of off-odours are similar to the measurements in the spoiled product (Dalgaard, 1993).
The latter step is particularly important, as some bacteria may produce the chemical compounds associated with spoilage but are unable to do so in significant amounts, and they are thus not the specific spoilage bacteria. When stored aerobically, levels of 108-109 cfu/g of specific spoilage bacteria are required to cause spoilage. The spoilage of packed fish is seen at a much lower level of 107 cfu P. phosphoreum per gramme. This relatively low level is probably due to the very large size (5 mm) of the bacterium resulting in a much higher yield of for example, TMA per cell (Dalgaard, 1993).
Spoilage potential and activity can be assessed in several fish substrates as sterile, raw fish juice (Lerke et al., 1963), heat-sterilized fish juice (Castell and Greenough, 1957; Gram et al., 1987; Dalgaard, 1993) or on sterile muscle blocks (Herbert et al., 1971). The latter is the most complicated but is also that yielding results comparable to the product. If any of the fish juices are chosen, it is important that the growth rate of the spoilage bacteria in the model system is equal to the growth rate in the product.
A qualitative test for the ability of the bacteria to produce H2S and/or reduce TMAO may also be used when the spoilage flora is screened for potential spoilage bacteria. A medium where the reduction of TMAO to TMA is seen as a redox indicator changes colour, and the formation of H2S is evident from a black precipitation of FeS which has been developed for this purpose (Gram et al., 1987).
Shewanella putrefaciens has been identified as the specific spoilage bacteria of marine temperate-water fish stored aerobically in ice. If the product is vacuum-packed, P. phosphoreum participates in the spoilage and it becomes the specific spoilage bacteria of CO2 packed fish (see section 6.3). The spoilage flora on iced tropical fish from marine waters is composed almost exclusively of Pseudomonas spp. and S. putrefaciens. Some Pseudomonas spp. are the specific spoilers of iced stored tropical freshwater fish (Lima dos Santos, 1978; Gram et al., 1990) and are also, together with S. putrefaciens, spoilers of marine tropical fish stored in ice (Gillespie and MacRae, 1975; Gram, 1990).
At ambient temperature, motile aeromonads are the specific spoilers of aerobically stored freshwater fish (Gorzyka and Pek Poh Len, 1985; Gram et al., 1990). Barile et al. (1985) showed that a large proportion of the flora on ambient-stored mackerel consisted of S. putrefaciens, indicating that this bacterium may also take part in the spoilage.
Table 5.5 gives an overview of the specific spoilage bacteria of fresh fish products stored in ice and at ambient temperature.
Table 5.5 Dominating microflora and specific spoilage bacteria at spoilage of fresh, white fish (cod) (from Huss, 1994)
| Storage tempe rature |
Packaging atmos- phere |
Dominating microflora |
Specific spoilage organisms (SSO) |
References |
| 0°C | Aerobic | Gram-negative
psychrotrophic, non-fermentative rods (Pseudomonas spp., S. putrefaciens, Moraxella, Acinetobacter) |
S. putrefaciens Pseudomonas3 |
2,3,4,9 |
| Vacuum | Gram-negative
rods; psychrotrophic or with psychrophilic character (S. putrefaciens, Photobacterium) |
S. putrefaciens P. phosphoreum |
1,9 | |
| MAP1 | Gram-negative
fermentative rods with psychrophilic character (Photobacterium ) Gram-negative non-fermentative psychrotrophic rods (1-10% of flora; Pseudomonas, S. putrefaciens) Gram-Positive rods (LAB2) |
P. phosphoreum | 1, 7 | |
| 5°C | Aerobic | Gram-negative
psychrotrophic rods (Vibrionaceae, S. putrefaciens) |
Aeromonas spp. S. putrefaciens |
10 |
| Vacuum | Gram-negative
psychrotrophic rods (Vibronaceae) S. putrefaciens) |
Aeromonas spp. S. putrefaciens |
10 | |
| MAP | Gram-negative psychrotrophic rods ( Vibrionaceae) | Aeromonas spp. | 6 | |
| 20-30°C | Aerobic | Gram-negative
mesophilic fermentative rods (Vibrionaceae, Enterobacteriaceae) |
Motile Aeromonas
spp. (A. hydrophila) |
2,4,5,8 |
1) Modified Atmosphere Packaging (CO2 containing)
2) LAB: Lactic Acid Bacteria
3) Fish caught in tropical waters or freshwaters tend to have a
spoilage dominated by Pseudomonas spp.
References: 1) Barile et al. (1985); 2) Dalgaard et al., (1993); 3) Donald and Gibson (1992); 4) orczyca and Pek Poh Len (1985); 5) Gram et al. (1987); 6) Gram et al. (1990); 7) Gram and Dalgaard (pers. comm.); 8) Jorgensen and Huss (1989); 9) Lima dos Santos (1978); 10) van Spreekens (1977)
Biochemical changes induced by bacterial growth during storage and spoilage
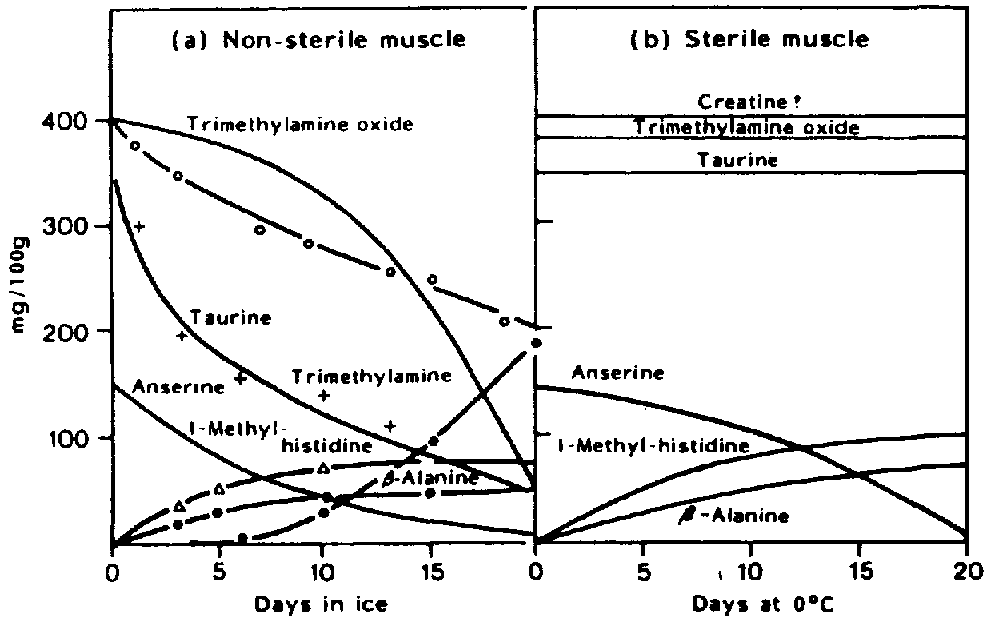
Comparison of the chemical compounds developing in naturally spoiling fish and sterile fish has shown that most of the volatile compounds are produced by bacteria (Shewan, 1962) as shown in Figure 5.11. These include trimethylamine, volatile sulphur compounds, aldehydes, ketones, esters, hypoxanthine as well as other low molecular weight compounds.
The substrates for the production of volatiles are the carbohydrates (e.g., lactate and ribose), nucleotides (e.g., inosine mono-phosphate and inosine) and other NPN molecules. The amino-acids are particularly important substrates for formation of sulphides and ammonia.
Microorganisms obtain far more energy from aerobic oxidation than from an anaerobic fermentation; thus the complete oxidation of 1 mole glucose (or other hexose) via Kreb's cycles yields 6 moles of CO2 and 36 moles of ATP. On the contrary, the fermentation of 1 mole glucose gives only 2 moles of ATP and two moles of lactic acid. The initial aerobic growth on fish is dominated by bacteria using carbohydrates as substrate and oxygen as terminal electron-acceptor with the concurrent production of CO2 and H2O.
Reduction of Trimethylamine Oxide (TMAO)
The growth of oxygen-consuming bacteria results in the formation of anaerobic or microaerophilic niches on the fish. This does, however, not necessarily favour the growth of anaerobic bacteria. Some of the bacteria present on fish are able to carry out a respiration (with the ATP advantage) by using other molecules as electron acceptor. It is typical of many of the specific spoilage bacteria on fish that they can use TMAO as electron acceptor in an anaerobic respiration. The reduced component, TMA, which is one of the dominant components of spoiling fish, has a typical fishy odour. The level of TMA found in fresh fish rejected by sensory panels varies between fish species, but is typically around 10-15 mg TMA-N/100 g in aerobically stored fish and at a level of 30 mg TMA-N/100 g in packed cod (Dalgaard et al., 1993).
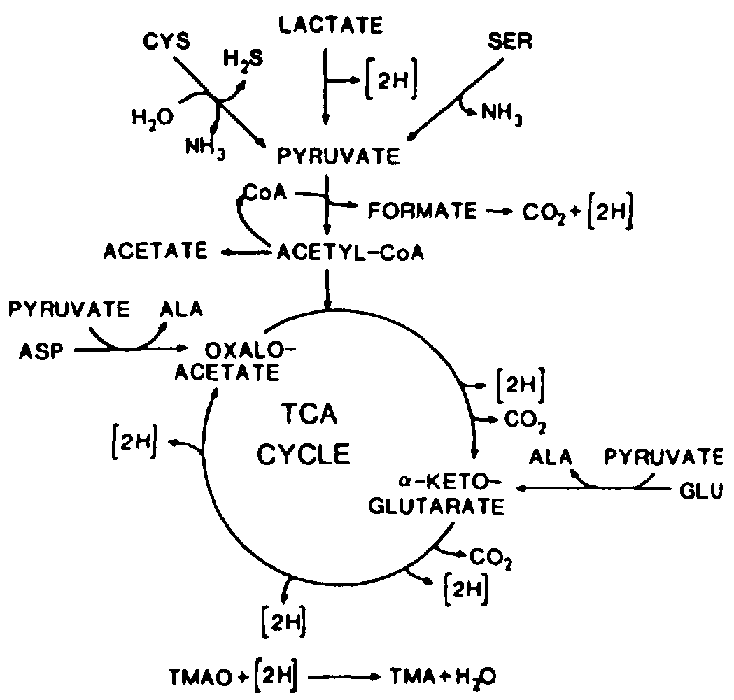
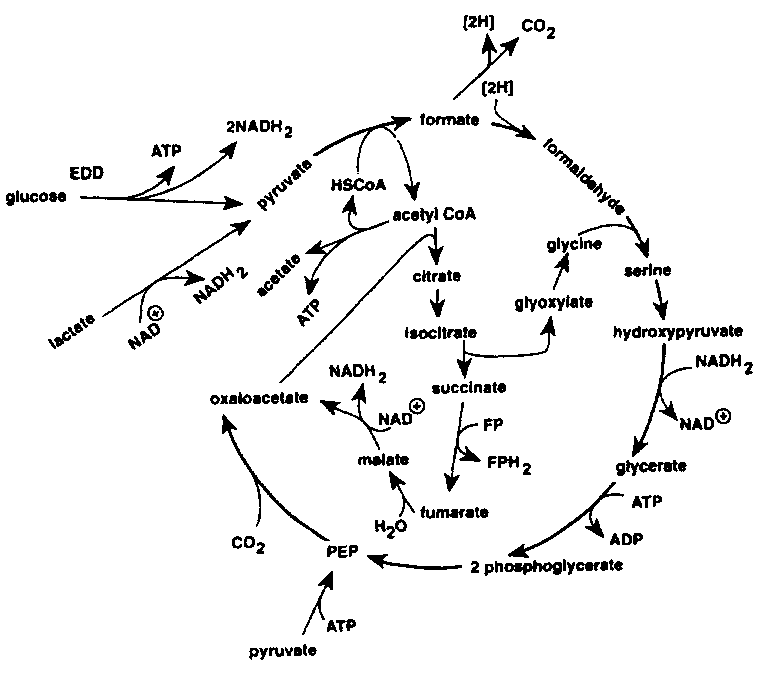
The TMAO reduction is mainly associated with the genera of bacteria typical of the marine environment (Alteromonas, Photobacterium, Vibrio and S. putrefaciens), but is also carried out by Aeromonas and intestinal bacteria of the Enterobacteriaceae. TMAO reduction has been studied in fermentative, facultative anaerobic bacteria like E. cold (Sakaguchi et al., 1980) and Proteus spp. (Stenberg et al., 1982) as well as in the non- fermentative S. putrefaciens (Easter et al., 1983; Ringo et al., 1984). During aerobic growth, S. putrefaciens uses the Kreb's cycle to produce the electrons that are later channelled through the respiratory chain. Ringo et al. (1984) suggested that during anaerobic respiration S. putrefaciens also uses the complete Kreb's cycle (Figure 5.12), whereas it has recently been shown that in the anaerobic respiration in S. putrefaciens, only part of the Kreb's cycle is used (Figure 5.13) and electrons are also generated by another metabolic pathway, namely the serine pathway (Scott and Nealson, 1994). S. putrefaciens can use a variety of carbon sources as substrate in its TMAO-dependent anaerobic respiration, including formate and lactate. Compounds like acetate and succinate that are used in the oxygen respiration cannot be used when TMAO is terminal electron acceptor (DiChristina and DeLong, 1994) and on the contrary, acetate is a product of the anaerobic TMAO reduction (Ringo et al., 1984; Scott and Nealson, 1994).
Figure 5.12 Anaerobic reduction of TMAO by Shewanella putrefaciens (formerly Alteromonas) as suggested by Ringo et al. (1984)
Contrary to this, sugars and lactate are the main substrates generating electrons when Proteus spp. reduces TMAO. The reduction is accompanied by a production of acetate as the main product (Kjosbakken and Larsen, 1974).
TMAO is, as mentioned in section 4.4, a typical component of marine fish, and it has recently been reported that also some tropical freshwater fish contain high amounts of TMAO (Anthoni et al., 1990). However, TMA is not necessarily a characteristic component during spoilage of such fish because spoilage is due to Pseudomonas spp. (Gram et al., 1990).
The development of TMA is in many fish species paralleled by a production of hypoxanthine. Hypoxanthine can, as described in section 5.2. be formed by the autolytic decomposition of nucleotides, but it can also be formed by bacteria; and the rate of bacterial formation is higher than the autolytic. Both Jorgensen et al. (1988) and Dalgaard (1993) showed a linear correlation between the contents of TMA and hypoxanthine during iced storage of packed cod (Figure 5.14). Several of the spoilage bacteria produce hypoxanthine from inosine or inosine mono-phosphate, including Pseudomonas spp. (Surette et al., 1988) S. putrefaciens (van Spreekens, 1977; Jorgensen and Huss, 1989; Gram, 1989) and P. phosphoreum (van Spreekens, 1977).
In cod and other gadoid fishes, TMA constitutes most of the so-called total volatile bases, TVB (also called total volatile nitrogen, TVN) until spoilage. However, in the spoiled fish where the TMAO supplies are depleted and TMA has reached its maximum level, TVB levels still rise due to formation of NH3 and other volatile amines. A little ammonia is also formed in the first weeks of iced storage due to autolysis. In some fish that do not contain TMAO or where spoilage is due to a non-TMAO reducing flora, a slow rise in TVB is seen during storage, probably resulting from the deamination of amino-acids.
Volatile sulphur-compounds are typical components of spoiling fish and most bacteria identified as specific spoilage bacteria produce one or several volatile sulphides. S. putrefaciens and some Vibrionaceae produce H2S from the sulphur containing amino-acid l-cysteine (Stenstroem and Molin, 1990; Gram et al., 1987). On the contrary, neither Pseudomonas nor P. phosphoreum produce significant amounts of H2S. Thus, hydrogen sulphide, which is typical of spoiling iced cod stored aerobically, is not produced in spoiling CO2 packed fish (Dalgaard et al., 1993). Methylmercaptan (CH3SH) and dimethylsulphide ((CH3)2S) are both formed from the other sulphur-containing aminoacid, methionine. Taurine, which is also sulphur-containing, occurs as free amino-acid in very high concentrations in fish muscle. It disappears from the fish flesh during storage (Figure 5.11) but this is because of leakage rather than because of bacterial attack (Herbert and Shewan, 1975). The formation of compounds in naturally-spoiling cod as compared to sterile muscle is shown in Figure 5.15.
The volatile sulphur-compounds are very foul-smelling and can be detected even at ppb levels, so even minimal quantities have a considerable effect on quality.
Ringo et al. (1984) have shown that cysteine is used as substrate in the Kreb's cycle when electrons are transferred to TMAO, and the formation of H2S and TMA is thus to some extent a linked reaction (Figure 5.12).
Contrary to the iced spoilage by S. putrefaciens and the ambient spoilage by Vibrionaceae which is dominated by H2S and TMA, the spoilage caused by Pseudomonas spp. is characterized by absence of these compounds (Gram et al., 1989, Gram et al., 1990). Fruity, rotten, sulphydryl odours and flavours are typical of the Pseudomonas spoilage of iced fish. Pseudomonas spp. produce a number of volatile aldehydes, ketones, esters and sulphides (Edwards et al., 1987; Miller et al., 1973 a, 1973 b). However, it is not known which specific compounds are responsible for the typical off-odours (Table 5.6). The fruity off-odours produced by Pseudomonas fragi originate from monoamino-monocarboxylic amino-acids.
Table 5.6 Typical spoilage compounds during spoilage of fresh fish stored aerobically or packed in ice or at ambient temperature
| Specific spoilage organism | Typical spoilage compounds |
| Shewanella
putrefaciens Photobacterium phosphoreum Pseudomonas spp. Vibrionaceae anaerobic spoilers |
TMA, H2S,
CH3SH, (CH3)2S, Hx TMA, Hx ketones, aldehydes, esters, non-H2S sulphides TMA, H2S NH3, acetic, butyric and propionic acid |
As mentioned above, TVB will continue to rise even after TMA has reached its maximum. This latter rise is due to proteolysis commencing when several of the free amino-acids have been used. Lerke et al. (1967) separated fish juice into a protein and a non-protein fraction and inoculated spoilage bacteria in each fraction and in the whole juice. The non-protein fraction of a fish juice spoiled as the whole juice whereas only faint off-odours were detected in the protein fraction of the juice. Although some authors have used the number of proteolytic bacteria as indicators of spoilage, it must be concluded that the turnover of the protein fraction is not of major importance in spoilage of fresh fish.
Some of the compounds typically formed by bacteria during spoilage of fish are shown in Table 5.7 together with the substrate used for the formation.
Table 5.7 Substrate and off-odour/off-flavour compounds produced by bacteria during spoilage of fish
| Substrate | Compounds produced by bacterial action |
| TMAO cysteine methionine carbohydrates and lactate inosine, IMP amino-acids (glycine, serine, leucine) amino-acids, urea |
TMA H2S CH3SH, (CH3)2S acetate, CO2, H2O hypoxanthine esters, ketones, aldehydes NH3 |
The formation of TMA is accompanied by a formation of ammonia during anaerobic storage of herring and mackerel (Haaland and Njaa, 1988). Prolonged anaerobic storage of fish results in vigorous production of NH3 owing to further degradation of the aminoacids, and in the accumulation of lower fatty acids as acetic, butyric and propionic acid. The very strong NH3- producers were found to be obligate anaerobes belonging to the family Bacteroidaceae genus Fusobacterium (Kjosbakken and Larsen, 1974; Storroe et al., 1975, 1977). These organisms grow only in the spoiled fish extract and have little or no proteolytic activity relying on already hydrolysed proteins.
During iced storage of fresh fatty fish, changes in the lipid fraction is caused almost exclusively by chemical action, e.g., oxidation, whereas bacterial attack on the lipid fraction contributes little to the spoilage profile. During storage of lightly preserved fish, lipid hydrolysis caused by bacteria may be part of the spoilage profile.
5.4 Lipid oxidation and hydrolysis
The two distinct reactions in fish lipids of importance for quality deterioration are:
- oxidation
- hydrolysis
They result in production of a range of substances among which some have unpleasant (rancid) taste and smell. Some may also contribute to texture changes by binding covalently to fish muscle proteins. The various reactions are either nonenzymatic or catalyzed by microbial enzymes or by intracellular or digestive enzymes from the fish themselves. The relative significance of these reactions, therefore, mainly depends on fish species and storage temperature.
Fatty fish are, of course, particularly susceptible to lipid degradation which can create severe quality problems even on storage at subzero temperatures.
Oxidation
The large amount of polyunsaturated fatty acid moieties found in fish lipids (see section 4.2) makes them highly susceptible to oxidation by an autocatalytic mechanism (Figure 5.16). The process is initiated as described below by abstraction of a hydrogen atom from the central carbon of the pentadiene structure found in most fatty acid acyl chains containing more than one double bond:
-CH = CH-CH2-CH = CH- (r) -CH = CH-CH-CH = CH- +H.
Contrary to the native molecule, the lipid radical (L ) reacts very quickly with atmospheric oxygen making a peroxy-radical (LOO ) which again may abstract a hydrogen from another acyl chain resulting in a lipid hydroperoxide (LOOM) and a new radical L. This propagation continues until one of the radicals is removed by reaction with another radical or with an antioxidant (AH) whose resulting radical (A) is much less reactive. The hydroperoxides produced in relatively large amounts during propagation are tasteless, and it is therefore perhaps not surprising that the widely used "peroxide value" (section 8.2) usually correlates rather poorly to censorial properties.
Figure 5.16 Autoxidation of polyunsaturated lipid
The hydroperoxides are readily broken down, catalyzed by heavy metal ions, to secondary autoxidation products of shorter carbon chain-length. These secondary products - mostly aldehydes, ketones, alcohols, small carboxylic acids and alkanes - give rise to a very broad odour spectrum and in some cases to a yellowish discoloration. Several of the aldehydes can be determined as "thiobarbituric acid- reactive substances" (section 8.2).
Metal ions are very important in the first step of lipid autoxidation - the initiation process- in catalyzing the formation of reactive oxygen species as for example the hydroxyl radical (OH ). This radical immediately reacts with lipids or other molecules at the site where it is generated. The high reactivity may explain that free fatty acids have been found to be more susceptible to oxidation than the corresponding bound ones, because the amount of iron in the aqueous phase is probably greater than the amount bound to the surface of cellular membranes and lipid droplets.
Fatty acid hydroperoxides may also be formed enzymatically, catalyzed by lipoxigenase which is present in variable amounts in different fish tissues. A relatively high activity has been found in the gills and under the skin of many species. The enzyme is unstable and is probably important for lipid oxidation only in fresh fish. Cooking or freezing/thawing rather effectively destroys the enzyme activity.
The living cells possess several protection mechanisms directed against lipid oxidation products. An enzyme, glutathione peroxidase, exists which reduces hydroperoxides in the cellular membranes to the corresponding hydroxy-compounds. This reaction demands supply of reduced glutathione and will therefore cease post mortem when the cell is depleted of that substance. The membranes also contain the phenolic compound alfa- tocopherol (Vitamin E) which is considered the most important natural antioxidant. Tocopherol can donate a hydrogen atom to the radicals L or LOO functioning as the molecule AH in Figure 5.16. It is generally assumed, that the resulting tocopheryl radical reacts with ascorbic acid (Vitamin C) at the lipid/water interface regenerating the tocopherol molecule. Other compounds, for example the carotenoids, may also function as antioxidants. Wood smoke contains phenols which may penetrate the fish surface during smoking and thereby provide some protection against lipid oxidation.
Hydrolysis
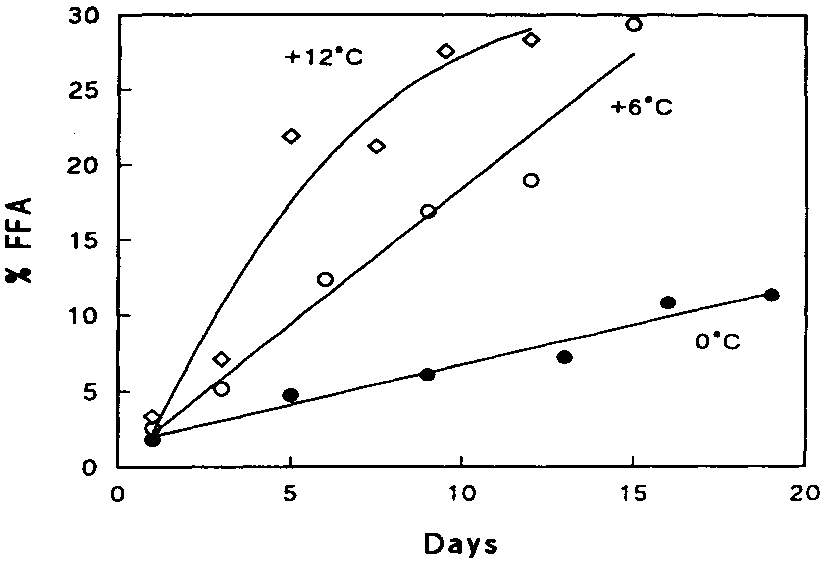
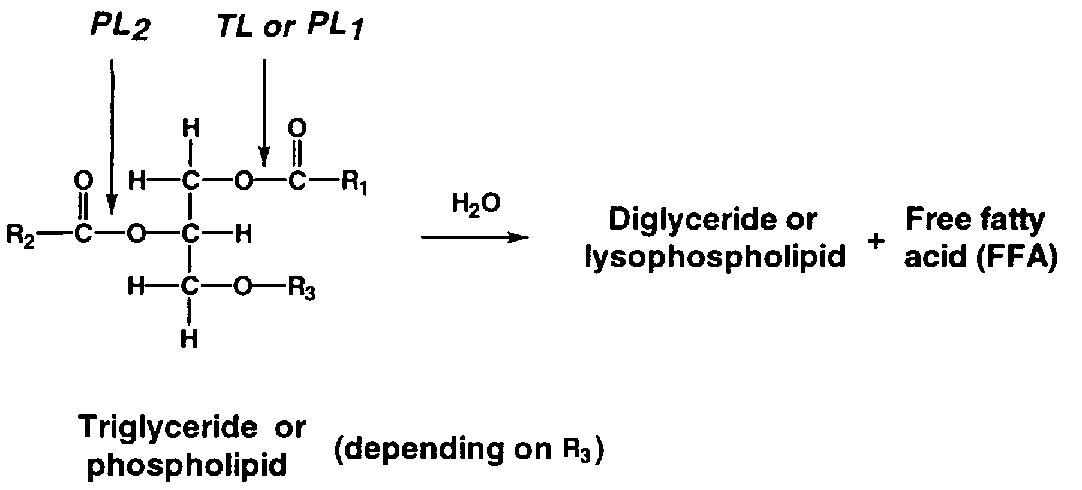
During storage, a considerable amount of free fatty acids (FFA) appears (Figure 5.17). The phenomenon is more profound in ungutted than in gutted fish probably because of the involvement of digestive enzymes. Triglyceride in the depot fat is cleaved by triglyceride lipase (TL in Figure 5.18) originating from the digestive tract or excreted by certain microorganisms. Cellular lipases may also play a minor role.
In lean fish, for example Atlantic cod, production of free fatty acids also occurs, even at low temperatures. The enzymes responsible are believed to be cellular phospholipases - in particular phospholipase A2 (PL2 in Figure 5.18) - although a correlation between activity of these enzymes and the rate of appearance of FFA has as yet not been firmly established. The fatty acids bound to phospholipids at glycerol-carbon atom 2 are largely of the polyunsaturated type, and hydrolysis therefore often leads to increased oxidation as well. Furthermore, the fatty acids themselves may cause a "soapy" off-flavour.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}