![]()
![]()
![]()
Tout programme d'élevage sélectif a pour objectif final d'améliorer la valeur génétique de la population considérée, valeur qui est fonction des gènes dont sont dotés les poissons. Lorsqu'un exploitant entreprend d'améliorer la valeur génétique de ses poissons, il souhaite aussi en accroître la valeur monétaire, qui est tributaire de leurs phénotypes. A cet effet, il sélectionne (épargne) les poissons dotés de certains phénotypes et réforme (élimine) les autres, en souhaitant que ce programme d'amélioration lui permette d'obtenir une population d'une valeur génétique supérieure (la sélection étant une forme appliquée de la génétique, on appelle sélectionneur l'aquiculteur qui met en train un programme d'élevage sélectif). Si le programme réussit, la valeur des poissons de la génération suivante s'en trouvera accrue, du fait que leurs gènes leur permettront de se développer plus rapidement ou d'acquérir une couleur plus attrayante.
Bien que les programmes d'élevage sélectif aient pour objet de manipuler les gènes d'une population afin d'obtenir des poissons supérieurs, il est impossible d'examiner et de manipuler les gènes directement. En fait, on obtient indirectement des renseignements sur les gènes en étudiant les phénotypes (aussi appelés “caractères”) qui leur correspondent et en sont l'expression physique. Comme il s'agit de manipuler les gènes des poissons, il importe de comprendre comment les gènes se transmettent d'un parent à sa descendance et comment ils produisent des phénotypes. Le fait de bien comprendre ces processus permet de mieux saisir comment la sélection opère. Cela a son importance. En effet, si un pisciculteur comprend les raisons des tâches qui lui incombe, il y a plus de chances qu'il les accomplissent correctement et, donc, qu'elles soient couronnées de succès. De plus, une bonne compréhension de la façon dont les gènes suscitent l'expression de différents phénotypes permet au sélectionneur de choisir le programme d'amélioration qui lui permettra d'atteindre rapidement et efficacement l'objectif qu'il s'est fixé.
Le présent chapitre ne constitue pas un exposé exhaustif de génétique élémentaire. Seuls certains sujets sont abordés. La première section traite de la méiose, c'est-à-dire du processus par lequel les gènes d'un parent sont répartis dans ses gamètes (les oeufs et le sperme). La deuxième section précise la différence entre génotype et phénotype. Les autres sections portent sur l'héritabilité des phénotypes.
Les renseignements contenus dans le présent chapitre revêtent une importance particulière du fait que bon nombre des termes qui y sont mentionnés sont repris dans les chapitres 3 et 4, où l'on explique comment les programmes d'élevage sélectif doivent être mis en oeuvre et comment ils fonctionnent. S'il n'est pas indispensable d'être généticien pour exécuter avec succès un programme d'élevage sélectif, le fait de bien comprendre la nature héréditaire de l'expression des phénotypes permettra à l'exploitant de saisir plus aisément le reste du manuel et de devenir meilleur sélectionneur.
Le présent chapitre contient simplement des informations de base destinées à ceux qui souhaitent mieux comprendre comment la sélection modifie les phénotypes et améliore une population. Il s'adresse plus particulièrement - notamment les parties concernant les phénotypes quantitatifs -- aux agents de vulgarisation et aux aquiculteurs bénéficiant d'une formation supérieure. Il n'est pas destiné à la grande majorité des exploitants, ni à ceux qui s'intéressent uniquement aux aspects pratiques de la mise en oeuvre d'un programme d'élevage sélectif et ne se préoccupent pas de la façon dont il fonctionne. Il est souhaitable que les agents de vulgarisation lisent le présent chapitre, car ils y acquerront de solides connaissances en matière de pisciculture; de plus, une bonne compréhension de la façon dont opère la sélection au plan génétique leur facilitera la tâche lorsqu'il leur faudra concevoir et mettre en oeuvre des programmes d'élevage sélectif.
Ceux qui se désintéressent de ces questions ou qui possèdent déjà des connaissances suffisantes en matière de génétique fondamentale peuvent sauter le présent chapitre et passer directement aux chapitres suivants, où sont décrits les différents programmes d'élevage sélectif.
Les gènes sont situés sur des structures appelées “chromosomes”, elles-mêmes situées dans le noyau de chaque cellule. A quelques exceptions près, les chromosomes se présentent par paires, ce qui signifie que les gènes sont des unités “appariées”. Dans le présent ouvrage, nous avons pris pour hypothèse que les chromosomes des poissons d'élevage étaient toujours associés par paires. Dans chaque paire, l'un des chromosomes provient de la mère et l'autre, du père. Le nombre de chromosomes, quoique variable selon l'espèce, reste néanmoins constant au sein d'une même espèce.
Bien que chaque cellule d'un poisson contienne la totalité du génome (la configuration génétique du poisson, déterminée par l'ensemble des gènes situés sur chacun des chromosomes), les gènes présents dans les gamétocytes primaires sont ceux qui présentent en fait le plus grand intérêt pour le sélectionneur. Tous les gènes qui se trouvent dans les différentes cellules d'un poisson jouent bien entendu un rôle important, puisque ce sont eux qui produisent les divers phénotypes que le pisciculteur soumettra à la sélection; mais ce sont néanmoins les gènes présents dans les gamétocytes primaires qui revêtent la plus grande importance, car ces cellules se transforment en sperme et en oeufs, c'est-à-dire en gamètes qui contiennent les gènes primordiaux et sont à l'origine de la génération suivante de poissons.
Les gamétocytes primaires sont les cellules qui se développent dans les oeufs et le sperme par un processus appelé “méiose”. La méiose compte parmi les principaux processus biologiques, puisqu'elle augmente considérablement la variabilité génétique par le biais des enjambements chromosomiques et de la disjonction indépendante. En outre, les erreurs qui interviennent pendant la méiose, lors de la réplication des chromosomes, sont héritables. C'est d'ailleurs de cette façon que se créent de nouveaux gènes et allèles. La méiose aboutit à la création de sperme et d'oeufs haploïdes. Sans la méiose, la vie n'aurait pas dépassé le stade des organismes unicellulaires.
Pendant la phase initiale de la méiose, les chromosomes se répliquent et les homologues de chaque paire se regroupent (les homologues sont les deux chromosomes qui forment une paire; l'un d'eux provient du père et l'autre, de la mère). La réplication des homologues est un processus qui se déroule habituellement sans anicroche, a savoir qu'ils se répliquent parfaitement. Il arrive toutefois qu'une erreur se produise pendant la réplication et qu'un gène se réplique imparfaitement. C'est ce qu'on appelle une “mutation”. Les mutations qui interviennent au niveau des différents gènes sont rares; on n'en compte pas plus de 1 pour 10 000 à 100 000 réplications. Malgré cette rareté, vu que les mâles peuvent produire plusieurs centaines de millions à plus de un milliard de spermatozoïdes et les femelles, des dizaines à des centaines de milliers d'oeufs, il est presque assuré que chaque poisson produit des douzaines de gamètes présentant une ou plusieurs mutations. La mutation est un phénomène biologique d'une très grande importance, car elle augmente la variabilité génétique en créant de nouveaux gènes ou allèles qui peuvent, à leur tour, engendrer de nouveaux phénotypes.
Les homologues se répliquant avant de se regrouper par paires, il se forme un ensemble de quatre chromosomes appelé “tétrade”. Les chromosomes de ces tétrades sont extrêmement allongés et étirés et ont tendance à s'enrouler les uns autour des autres. En s'enroulant, les chromosomes se brisent et des morceaux des différents homologues se rassemblent. Cet échange de gènes d'un homologue à l'autre constitue ce qu'on appelle un “enjambement chromosomique”. Il s'agit là d'un phénomène génétique primordial, qui contribue à accroître considérablement la variabilité génétique en créant des combinaisons de gènes nouvelles et différentes à chaque génération. Ces combinaisons augmentent à leur tour la variabilité phénotypique, ce qui aide les sélectionneurs à produire du poisson de meilleure qualité.
Le deuxième processus important qui intervient pendant la méiose est la division réductionnelle. Durant ce processus, le nombre de chromosomes est réduit de moitié, les chromosomes passant de l'état diploïde (appariés) à l'état haploïde (non appariés). Lorsque les chromosomes sont regroupés par paires, on parle de cellule ou d'individu diploïde (2n). A l'inverse, lorsqu'il n'y a qu'un seul chromosome de chaque paire, on parle de cellule ou d'individu haploïde (n).
Au cours de la division réductionnelle, les homologues répliqués de chaque paire de chromosomes se séparent, puis le gamétocyte primaire se divise pour former deux cellules. Chez les mâles, ces cellules consistent dans les spermatocytes de deuxième ordre et chez les femelles, dans l'oocyte de deuxième ordre et le premier globule polaire. La séparation des homologues répliqués de chaque paire de chromosomes et la direction dans laquelle migrent les homologues répliqués sont indépendantes de ce qui se produit pour tous les autres chromosomes. Bien que chaque tétrade se divise selon des critères parentaux (l'homologue répliqué qui provient du père d'un poisson se sépare de l'homologue répliqué qui provient de sa mère), la division des homologues de chaque paire de chromosomes s'effectue au hasard, indépendamment de ce qui se produit dans toutes les autres tétrades. La division au hasard des chromosomes maternels et paternels, qu'on appelle “disjonction indépendante”, est un processus d'une extrême importance, car il engendre de nouvelles combinaisons chromosomiques et génétiques, ce qui augmente considérablement la variabilité génétique et, par conséquent, phénotypique. Les spermatocytes de deuxième ordre et l'oocyte de deuxième ordre sont haploïdes, puisque chacun ne contient qu'un homologue de chaque paire.
La méiose s'achève par la division équationnelle, au cours de laquelle les homologues répliqués de chaque chromosome se séparent et migrent dans une des deux spermatides ou encore dans l'oeuf ou le deuxième globule polaire. Comme dans le cas de la division réductionnelle, la direction que prennent les homologues répliqués de chaque chromosome est aléatoire et indépendante de ce qui se passe dans l'ensemble des autres chromosomes. La redistribution finale des chromosomes et des gènes contribue également à accroître la variabilité génétique.
Le résultat final de la méiose consiste dans la production de spermatozoïdes et d'ovules haploïdes. Chaque gamète contient un seul chromosome issu de chaque paire. Même si certains poissons peuvent produire des millions de gamètes, peu d'entre eux -- voire aucun -- ne sont identiques du fait des mutations, des enjambements chromosomiques et de la disjonction indépendante des chromosomes qui intervient pendant la division réductionnelle.
La réduction de moitié du nombre des chromosomes entre le stade diploïde des oocytes et des spermatocytes de premier ordre et le stade haploïde des ovules et des spermatozoïdes est une étape cruciale. Si elle n'avait pas lieu, le nombre de chromosomes doublerait à chaque génération. Pour une espèce donnée, le fait que les gamètes soient haploïdes permet le rétablissement du nombre normal de chromosomes correspondant au stade diploïde après fécondation d'un oeuf par un spermatozoïde.
Un gène (ou un ensemble de gènes) contient le plan d'exécution ou les instructions chimiques concernant la production d'une protéine. Cette protéine forme, ou contribue à former, divers phénotypes tels que la couleur du corps, le sexe, le nombre de rayons de la nageoire dorsale, la longueur d'une nageoire, la longueur du corps ou le poids. Lorsqu'un généticien décrit ce processus, il dit que le génotype du poisson contrôle ou produit le phénotype qui lui correspond.
Le génotype est la structure génétique du poisson. C'est le gène ou les gènes qui contrôlent un phénotype particulier. Les chromosomes étant associés par paires, les gènes le sont aussi (il existe certaines exceptions dont il ne sera pas fait mention dans le présent manuel). Le génotype est donc une entité appariée.
Un gène peut se présenter sous plusieurs formes, appelées “allèles”. Dans une population, s'il arrive qu'un gène se présente sous une seule forme - ce qui signifie qu'il n'y a qu'un seul allèle pour un locus donné (locus = gène) - il advient aussi qu'une douzaine d'allèles correspondent à un seul locus.
Etant donné que les chromosomes sont associés par paires, un individu peut posséder un ou deux allèles en un locus donné. Même si, dans une population, il existe dix allèles pour un gène particulier, un individu ne peut pas en posséder plus de deux (dans le présent manuel, nous prendrons pour hypothèse que tous les poissons sont diploïdes). Si, en un locus donné, les allèles d'une même paire sont identiques, on dit que le poisson est homozygote pour ce locus; s'ils sont différents, on dit que le poisson est hétérozygote pour ce locus. Les termes “homozygote” et “hétérozygote” (le génotype) font référence à des gènes particuliers, et non pas à l'ensemble du génome d'un poisson. Ce génome est constitué de dizaines de milliers de gènes correspondant à un mélange de loci homozygotes et hétérozygotes.
S'il importe d'établir une distinction entre individus homozygotes et hétérozygotes, c'est que les différentes formes d'un même gène (les allèles) produisent différentes formes de protéines correspondant à ce gène. Cela signifie que les divers allèles occupant un locus déterminent la production de couleurs corporelles ou de taux de croissance différents. Et ce sont ces différences qui intéressent les généticiens, parce qu'on peut en tirer parti par sélection pour obtenir des poissons d'un aspect plus attrayant ou qui se développent plus rapidement.
Le phénotype est l'expression physique de ce que le ou les gènes produisent; on peut le décrire (la couleur ou le sexe, par exemple) ou le mesurer (la longueur ou le poids, par exemple). Les sélectionneurs divisent les phénotypes en deux grandes catégories : les phénotypes qualitatifs et les phénotypes quantitatifs.
Les phénotypes qualitatifs sont les phénotypes que l'on décrit (la couleur, le sexe ou la disposition des écailles, par exemple). Ce sont aussi ceux qui sont les plus faciles à observer, simplement parce qu'un individu se range dans une catégorie discrète, descriptive et non chevauchante, ou bien dans une autre. Par exemple, si une population comprend des poissons bleus et jaunes, chaque poisson se range dans la catégorie bleue ou jaune.
La génétique des phénotypes qualitatifs est une génétique simple, souvent appelée “génétique mendélienne” en l'honneur de Gregori Mendel, son inventeur. Ces phénotypes sont généralement contrôlés par un ou deux gènes. Les diverses formes d'un phénotype (bleu ou jaune, par exemple) sont produites par les diverses formes d'un gène (les allèles). Le phénotype normal est souvent appelé “phénotype commun” ou “phénotype de type sauvage”; quant aux autres phénotypes, ils sont qualifiés de “mutants”.
Les phénotypes qualitatifs sont souvent qualifiés d'“esthétiques”, parce qu'ils influent en premier lieu sur l'apparence des individus. Cela ne signifie pas qu'ils soient sans importance. Ils peuvent en effet avoir un effet positif sur la santé des individus ou rendre le produit plus attrayant pour les consommateurs. C'est ainsi que le nanisme est un phénotype recherché dans le cas de nombreuses variétés de blé, car les tiges courtes sont plus résistantes que les tiges longues normales et n'ont en outre pas tendance à ployer ou à se rompre lorsque la plante se développe. De la même façon, l'absence de cornes est un phénotype recherché dans le cas de nombreuses variétés de bovins pour des raisons de sécurité et de santé, tout comme le plumage blanc est un phénotype recherché dans le cas de la volaille, car des plumes de couleur foncée, une fois enlevées, laissent sur la peau des traces sombres peu attrayantes. Les phénotypes qualitatifs peuvent aussi grandement augmenter la valeur de certains poissons d'élevage. Ainsi, aux Etats-Unis, la couleur corporelle des vairons à grosse tête a un effet considérable sur leur valeur marchande : les vairons à pigmentation normale (foncée) valent 6,05 $/kg, alors que les vairons de couleur rouge rosé valent 8,25 $/kg. Il suffit par ailleurs d'évoquer l'élevage de poissons ornementaux à l'échelle industrielle pour se rendre compte de l'importance des phénotypes qualitatifs. La valeur d'un poisson de ce genre est fonction de sa couleur, de sa livrée, de la forme de ses nageoires, de la forme de ses yeux, etc.
Il existe deux grandes catégories de phénotypes qualitatifs : les phénotypes autosomiques et les phénotypes liés au sexe. Les phénotypes autosomiques sont contrôlés par des gènes situés sur un autosome (chromosome non sexuel). Les phénotypes liés au sexe sont contrôlés par des gènes situés sur la paire de chromosomes qui déterminent le sexe. (Il existe certaines exceptions; certains poissons possèdent plus d'une paire de chromosomes sexuels, alors que d'autres en comptent un nombre impair -- un ou trois. Les principaux poissons comestibles d'élevage ont cependant tous une seule paire de chromosomes sexuels.)
Les gènes autosomiques s'héritent et s'expriment de façon identique chez les mâles et les femelles (à moins que l'expression phénotypique nécessite une hormone sexuelle). A l'inverse, les gènes liés au sexe s'héritent et s'expriment de façon différente chez les mâles et les femelles. Jusqu'ici, tous les phénotypes qualitatifs des poissons comestibles que l'on est parvenu à déchiffrer étaient des phénotypes autosomiques. On a décelé des gènes liés au sexe uniquement chez les poissons ornementaux, et ce sont les guppys et les plattys qui ont fourni la plus grande partie des informations concernant ce type de transmission héréditaire. Comme tous les phénotypes qualitatifs qui ont été découverts chez les poissons comestibles d'élevage sont des phénotypes autosomiques, seule la génétique de ces phénotypes sera abordée dans la présente section, qui ne traite donc pas des phénotypes liés au sexe.
Par souci de simplicité, tous les exemples donnés dans la présente section se rapportent à des gènes ayant deux allèles. En fait, dans une population, il arrive qu'un gène ait une douzaine d'allèles. Chez les plattys, par exemple, la disposition des taches caudales est contrôlée par un gène à neuf allèles. Lorsqu'un gène a plus de deux allèles, il peut produire un très grand nombre de phénotypes qualitatifs, pour peu que son mode d'action le permette. Il est plus difficile d'obtenir des résultats convenables avec les phénotypes de ce genre; cela exige que l'on déploie des efforts considérables et qu'on réunisse beaucoup plus de données, même s'il ne s'agit de procéder qu'à une simple extrapolation des informations fournies dans la présente section. Heureusement, ces gènes sont rares et n'ont été décelés que chez les poissons ornementaux.
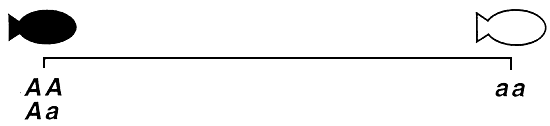
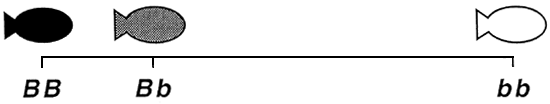
S'agissant des poissons comestibles, la plupart des phénotypes qualitatifs qui ont fait l'objet d'un déchiffrement génétique sont contrôlés par un seul gène autosomique présentant deux allèles par locus. Ce gène s'exprime généralement de façon additive ou non additive. En cas d'action génétique additive, chacun des deux allèles contribue au même degré à la production des phénotypes selon un mode unidirectionnel par étapes, et le phénotype hétérozygote occupe une position intermédiaire entre les deux phénotypes homozygotes. En cas d'action génétique non additive, l'un des allèles (l'allèle dominant) s'exprime plus fortement que l'autre (l'allèle récessif) et exerce donc une plus grande influence sur la production des phénotypes (figure 1).
Action génétique en dominance complète. Il y a action génétique en dominance complète lorsque l'allèle dominant est si vigoureux qu'il exprime son phénotype quel que soit le génotype. Un allèle dominant simple suffit donc à produire le phénotype dominant. Cela signifie que les génotypes homozygote dominant et hétérozygote produisent l'un et l'autre le phénotype dominant; les phénotypes contrôlés par ces génotypes sont par conséquent identiques. L'allèle récessif ne peut produire le phénotype récessif qu'à la condition qu'il n'y ait aucun allèle dominant, ce qui est le cas chez les poissons homozygotes récessifs uniquement. En conséquence, pour ce mode d'hérédité, on dénombre trois génotypes et seulement deux phénotypes possibles (figure 1 ) :
ACTION GÉNÉTIQUE EN DOMINANCE COMPLÈTE
ACTION GÉNÉTIQUE EN DOMINANCE IN COMPLÈTE
ACTION GÉNÉTIQUE ADDITIVE
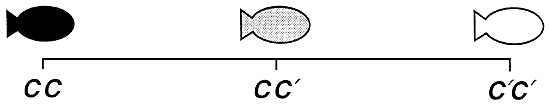
Figure 1. Représentation schématique des phénotypes qualitatifs contrôlés par un seul gène autosomique exerçant une action génétique en dominance complète ou incomplète ou encore une action génétique additive. Les génotypes sont indiqués au-dessous des phénotypes. Le gène A contrôle les couleurs blanche et noire par dominance complète, de sorte qu'il n'y a que deux phénotypes. Le noir, qui est le phénotype dominant, est produit à la fois par le génotype homozygote dominant (AA) et le génotype hétérozygote (Aa). Le blanc, qui est le phénotype récessif, est uniquement produit par le génotype homozygote récessif (aa). Le gène B contrôle les couleurs blanche et noire par dominance incomplète. Du fait que le mode d'action génétique est la dominance incomplète, le génotype hétérozygote (Bb) produit un phénotype unique (gris foncé), proche quoique légèrement différent du phénotype dominant (noir) produit par le génotype homozygote dominant (BB); quant au blanc, il correspond au phénotype récessif et s'exprime sous le contrôle du génotype récessif (bb). Le gène Ç contrôle les couleurs blanche et noire par action génétique additive. Aucun allèle n'étant dominant, le génotype hétérozygote (CC) produit un phénotype unique (gris) situé à mi-chemin des phénotypes noir et blanc produits par les deux phénotypes homozygotes (CC produit le phénotype “noir” et C'C'. le phénotype “blanc”). Lorsque le mode d'action génétique est additif, il n'y a pas d'allèle ou de phénotype dominant ou récessif.
| Phénotype | Génotype |
| Dominant | Homozygote dominant |
| Dominant | Hétérozygote |
| Récessif | Homozygote récessif |
Par exemple, chez les tilapias du Nil, les phénotypes correspondant à une pigmentation normale et à une pigmentation plus claire (rose) sont contrôlés par le gène B. L'allèle dominant B produit le phénotype dominant “pigmentation normale”, alors que l'allèle récessif b produit le phénotype récessif “rosé”. Le gène B agissant en dominance complète, la pigmentation normale est produite à la fois par le génotype homozygote dominant (BB) et par le génotype hétérozygote (Bb); quant à la couleur rose, elle est seulement produite par le génotype homozygote récessif (bb) (figure 2). Le tableau 1 énumère un certain nombre de phénotypes qualitatifs de poissons comestibles d'élevage courants, qui sont contrôlés par un seul gène autosomique agissant en dominance complète.
Tableau 1. Exemples de phénotypes de poissons comestibles d'élevage qui sont contrôlés par un seul gène autosomique agissant en dominance complète. Tous les phénotypes du présent tableau concernent la couleur corporelle, à l'exception du syndrome de difformité caudale (ou difformité de la aueue).
| Espèce | Phénotype récessif | Phénotype dominant |
| Carpe commune | bleu | pigmentation normale |
| doré | pigmentation normale | |
| gris | pigmentation normale | |
| pigmentation normale | bande jaune clair sur la nageoire dorsale; jaune sur la tête | |
| Tilapia du Nile | blond | pigmentation normale |
| miel | pigmentation normale | |
| couleur claire (rose) | pigmentation normale | |
| syndrome de difformité caudale | normal | |
| pigmentation normale | rouge | |
| Carpe de roseau | albinos | pigmentation normale |
| Ictalure tacheté | albinos | pigmentation normale |
| Truite arc-en-ciel | albinos | pigmentation normale |
| bleu métallique iridescent | pigmentation normale |
| PHENOTYPE | GENOTYPE |
 | |
| PIGMENTATION NORMALE | |
 | |
| PIGMENTATION NORMALE | |
 | |
| ROSE | |
Figure 2. Transmission héréditaire de la pigmentation normale et de la couleur rase du corps chez les tilapias du Nil. Ces phénotypes sont contrôlés par un seul gène autosomique agissant en dominance complète, appelé gène B : l'allèle dominant B l'emportant complètement sur l'allèle b, les génotypes BB et Bb produisent tous deux le phénotype dominant “pigmentation normale”. Le phénotype récessif “rose” s'exprime uniquement lorsque le poisson est homozygote récessif (bb). Les représentations graphiques des couleurs du corps dont il est fait usage dans la présente figure sont également utilisées aux figures 3 et 9.
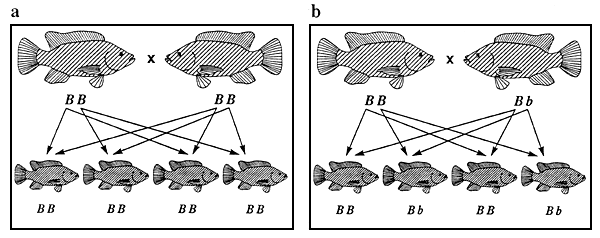
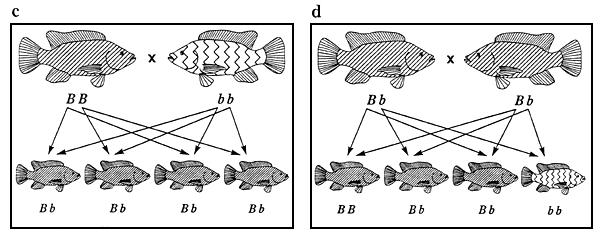
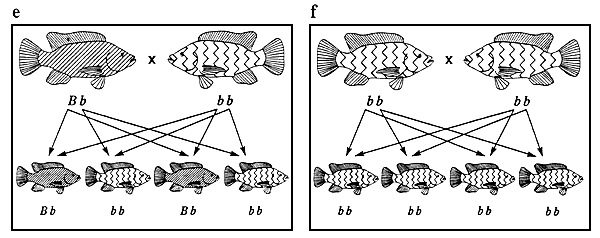
Figure 3. Ensemble des combinaisons de reproduction possibles entre des tilapias du Nil à pigmentation normale et de couleur rose et phénotypes se manifestant chez la descendance issue de chacun des accouplements. Les représentations graphiques des phénotypes sont identiques à celles de la figure 2. Les génotypes sont indiqués sous les poissons. Les flèches représentent les gamètes. Les combinaisons de reproduction a et f sont des exemples de combinaisons aboutissant à des populations génétiquement pures, ce qui constitue l'objectif de tout programme d'élevage sélectif.
La figure 3 illustre l'ensemble des combinaisons de reproduction possibles entre des tilapias du Nil de couleur rose et à pigmentation normale; les phénotypes se manifestant chez la descendance issue de chacun des accouplements sont également indiqués. Dans cette figure, les combinaisons de reproduction et la répartition phénotypique de la descendance issue de chaque croisement (par exemple 3 descendants à pigmentation normale pour un descendant de couleur rose dans le cas du croisement 3d) sont caractéristiques de l'ensemble des phénotypes qualitatifs contrôlés par un seul gène autosomique agissant en dominance complète (pour peu qu'aucun génotype ne soit létal).
On se fonde sur la répartition phénotypique de la descendance issue des différentes combinaisons de reproduction pour déchiffrer les gènes qui contrôlent les phénotypes. Des modes d'action génétique différents donnent lieu à une répartition phénotypique différente. Lors d'une expérience de génétique, le croisement 3c est l'un des premiers qu'on effectue. La descendance obtenue est appelée “descendance F1”. Lorsqu'ils sont parvenus à maturité, on croise ces poissons (croisement 3d) de façon à obtenir ce qu'on appelle la “descendance F2”. La répartition phénotypique de la descendance F2 est le rapport clé que l'on utilise pour déchiffrer la plupart des formes de transmission héréditaire.
A la figure 3, les combinaisons de reproduction 3a et 3f sont des exemples d'accouplements produisant des populations génétiquement pures. Tout programme d'élevage sélectif vise à créer une population de ce type (voir chapitre 3).
Action génétique en dominance incomplète. Il y a action génétique en dominance incomplète lorsque l'allèle dominant l'emporte sur l'allèle récessif, mais n'est toutefois pas assez vigoureux pour éclipser complètement l'allèle récessif dans le génotype hétérozygote. De ce fait, le phénotype dominant peut seulement s'exprimer lorsque le poisson dispose de deux exemplaires de l'allèle dominant (homozygote dominant). L'allèle récessif n'étant pas entièrement occulté par l'allèle dominant, le génotype hétérozygote produit un phénotype qui ressemble - sans être pour autant identique -- au phénotype dominant. Comme dans le cas d'une dominance complète, le phénotype récessif s'exprime uniquement chez les poissons homozygotes récessifs. Etant donné que le génotype hétérozygote produit un phénotype proche mais néanmoins différent du phénotype dominant, le mode d'action génétique en dominance incomplète se traduit par l'existence de trois génotypes et de trois phénotypes, à raison d'un unique phénotype par génotype (figure 1 ) :
| Phénotype | Génotype |
| Dominant | Homozygote dominant |
| Dominant | Hétérozygote |
| Récessif | Homozygote récessif |
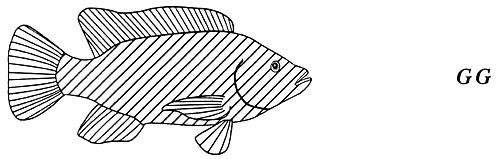
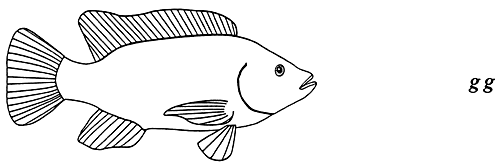
Par exemple, les couleurs noire (pigmentation normale), bronze et dorée du corps des tilapias du Mozambique sont contrôlées par le gène G. L'allèle G dominant produit un poisson mélanique (de couleur foncée), mais n'empêche pas totalement l'expression de l'allèle g. récessif au stade hétérozygote en raison du mode d'action en dominance incomplète du gène G_. Les génotypes homozygote dominant et hétérozygote contrôlent des phénotypes uniques : les poissons GG sont noirs, alors que les poissons Gg, sont de couleur bronze. Quant à la couleur dorée, elle est contrôlée par le génotype homozygote récessif gg. (figure 4). Le tableau 2 fournit des exemples de phénotypes qualitatifs contrôlés par un seul gène autosomique agissant en dominance incomplète chez des poissons comestibles d'élevage courants.
La figure 5 illustre l'ensemble des combinaisons de reproduction possibles entre les tilapias du Mozambique de couleur noire, bronze et dorée ainsi que les différents phénotypes de la descendance issue de chaque croisement. Les combinaisons de reproduction et la répartition phénotypique de la descendance issue de chacun des croisements illustrées dans cette figure (par exemple 1 noir/2 bronze/1 doré dans le cas du croisement 5d) sont caractéristiques de la totalité des phénotypes qualitatifs contrôlés par un seul gène autosomique agissant en dominance incomplète (pour peu qu'il n'y ait pas de génotype létal). Toujours à la figure 5, les combinaisons de reproduction 5a et 5f sont des exemples de combinaisons produisant des populations génétiquement pures, et la répartition phénotypique de la descendance issue du croisement 5d est la clé qui sert à décrypter le mode de transmission héréditaire.
Action génétique additive. A une exception près, tous les phénotypes qualitatifs contrôlés par un seul gène autosomique qui ont été identifiés chez les poissons comestibles d'élevage relèvent du mode de dominance complète ou incomplète. Cette unique exception consiste en un gène qui contrôle, par action génétique additive, les couleurs dorée, “alezan” (correspondant au phénotype hétérozygote) et normale du corps des truites arc-en-ciel. Lorsque les phénotypes sont soumis à une action génétique additive, il n'existe pas d'allèle dominant ou récessif. Les deux allèles participent au même titre à la production des phénotypes, de sorte que le génotype hétérozygote contrôle un phénotype qui occupe une position intermédiaire entre les phénotypes produits par les deux génotypes homozygotes. En conséquence, lorsque le mode de transmission héréditaire consiste en l'action génétique additive, on dénombre trois génotypes et trois phénotypes, à raison d'un phénotype unique par génotype.
Précisons en quoi consiste la différence entre action génétique additive et dominance incomplète. Lorsque le mode d'action génétique est la dominance incomplète, le phénotype hétérozygote est proche de l'un des phénotypes homozygotes (le phénotype dominant), alors qu'il est à mi-chemin de ces deux phénotypes lorsque le mode d'action génétique est de type additif (figure 1 ). Le fait que les phénotypes qualitatifs sont décrits, et non pas mesurés, rend possible une certaine confusion entre dominance incomplète et action génétique additive. Toutefois, une éventuelle erreur d'identification du mode d'action génétique n'aura aucune incidence pratique sur le bon déroulement du programme d'amélioration concerné. En effet, les deux modes d'action génétique se caractérisant par l'existence de trois génotypes dont chacun contrôle un unique phénotype, ce sont les mêmes programmes d'amélioration qu'on utilise pour influer sur les phénotypes engendrés par dominance incomplète ou action génétique additive.
Les combinaisons de reproduction et la répartition phénotypique de la descendance issue des différents croisements indiquées à la figure 5 peuvent également servir à illustrer ce qu'il advient dans le cas des phénotypes contrôlés par un seul gène autosomique agissant de façon additive. En effet, les répartitions phénotypiques relevant de la dominance incomplète et de l'action génétique additive sont identiques. La seule différence réside dans l'aspect du phénotype hétérozygote, qui ressemble fortement au phénotype dominant en cas de dominance incomplète ou qui se situe à mi-chemin des deux phénotypes homozygotes dans le cas d'une action génétique additive.
| PHENOTYPE | GENOTYPE |
 | |
| NOIR | |
 | |
| BRONZE | |
 | |
| DORÉ | |
Figure 4. Transmission héréditaire des couleurs corporelles noire, bronze et dorée chez les tilapias du Mozambique. Ces phénotypes sont contrôlés par un seul gène autosomique agissant en dominance incomplète, appelé gène G. Du fait que l'allèle dominant G ne l'emporte pas totalement sur l'allèle récessif g, le génotype hétérozygote produit un phénotype peu différent, mais cependant distinct, de celui que produit le génotype homozygote dominant. Les poissons homozygotes dominants (GG) sont noirs (phénotype dominant), les poissons hétérozygotes (Gg) sont de couleur bronze (phénotype hétérozygote) et les poissons homozygotes récessifs (gg) sont de couleur dorée (phénotype récessif). Les représentations graphiques des couleurs du corps utilisées dans cette figure sont reprises aux figures 5, 10, 11, 17 et 18.
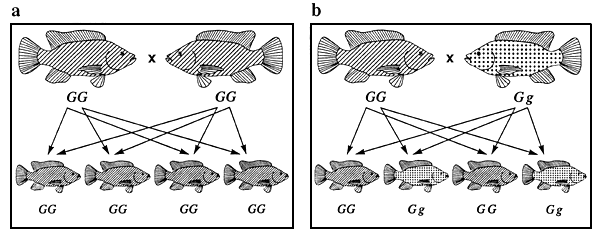
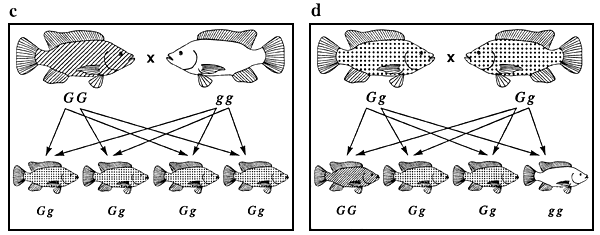
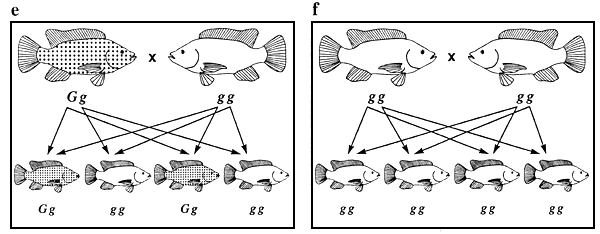
Figure 5. Ensemble des combinaisons de reproduction possibles entre les tilapias du Mozambique de couleur noire, bronze et dorée et différents phénotypes se manifestant chez la descendance issue de chacun des croisements. Les représentations graphiques des phénotypes sont les mêmes que celles utilisées à la figure 4. Les génotypes sont indiqués au-dessous des poissons. Les flèches représentent les gamètes. Les combinaisons de reproduction a et f sont des exemples de combinaisons produisant des populations génétiquement pures, ce qui constitue l'objectif de tout programme d'élevage sélectif.
Tableau 2. Exemples de phénotypes contrôlés par un seul gène autosomique agissant en dominance incomplète chez certains poissons comestibles d'élevage.
| Espèce | Phénotype dominant | Phénotype Heterozygote | Phénotype récessif |
| carpe commune | létal | de couleur claire | pigmentation normale |
| tilapia bleu | létal | ensellure (anomalie de la nageoire dorsale) | normal |
| tilapia du Mozambique | noir (pigmentation normale) | de couleur bronze | doré |
Certains phénotypes qualitatifs sont contrôlés par deux gènes autosomiques. Lorsque l'expression d'un ensemble de phénotypes est contrôlée par deux gènes, cela donne généralement lieu à une certaine interaction, et l'un des gènes influe sur l'expression de l'autre. Autrement dit, l'un des gènes modifie l'expression des phénotypes contrôlés par l'autre gène. Cette interaction des gènes est appelée “épistasie”.
Quoique la plupart des cas d'épistasie relevés chez les poissons concernent des poissons ornementaux, certains cas ont été découverts chez d'importantes espèces de poissons comestibles d'élevage. Les deux principaux cas consistent dans la disposition des écailles de la carpe commune et la couleur de la chair du quinnat. La carpe commune étant l'une des principales espèces comestibles d'élevage d'Asie et d'Europe, on peut soutenir que la disposition des écailles chez la carpe commune correspond au plus important ensemble de phénotypes qualitatifs qui ait été identifié chez une espèce aquicole. La disposition des écailles influe en outre sur la coloration - et donc la valeur - de la carpe commune ornementale (koï).
Chez la carpe commune, les quatre phénotypes relatifs à la disposition des écailles, à savoir les phénotypes “à écailles” (disposition normale des écailles), “miroir”, “à ligne” et “cuir”, sont contrôlés par deux gènes (S et N) liés par ce qu'on appelle une “épistasie dominante”. Le gène S détermine la disposition de base des écailles selon un mode de dominance complète. L'allèle dominant S suscite l'expression du phénotype “à écailles” (génotypes SS et Ss), alors que l'allèle récessif s produit le phénotype correspondant à des écailles réduites, que l'on appelle “miroir” (génotype ss). Le gène N modifie les phénotypes produits par le gène S. Il y a deux allèles au locus N. L'allèle dominant N modifie les phénotypes de la façon suivante : au stade homozygote (NN), l'allèle N provoque la mort de l'embryon; au stade hétérozygote (Nn), l'allèle N transforme le phénotype “à écailles” en phénotype “à ligne” et le phénotype “miroir” en phénotype “cuir”. L'allèle récessif n n'a aucun effet sur les phénotypes contrôlés par le gène S. Ces cinq phénotypes (y compris la mort) et la génétique sous-jacente sont illustrés à la figure 6.
Il arrive qu'un ensemble de phénotypes qualitatifs soient contrôlés par plus de deux gènes. Ainsi, chez les combattants siamois, la couleur du corps correspond à un ensemble de phénotypes contrôlés par l'interaction épistatique de quatre gènes. Ce nombre élevé de gènes complique beaucoup les travaux concernant ces phénotypes. Heureusement, chez les poissons comestibles, aucun phénotype qualitatif contrôlé par plus de deux gènes n'a été jusqu'ici découvert.
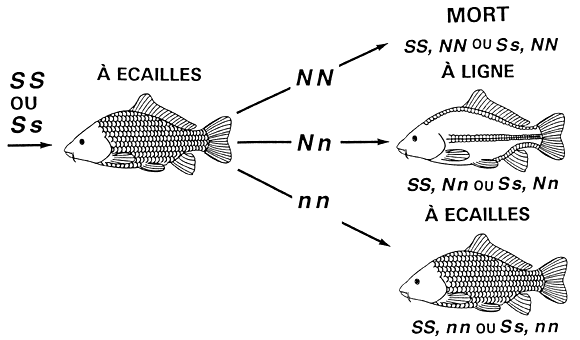
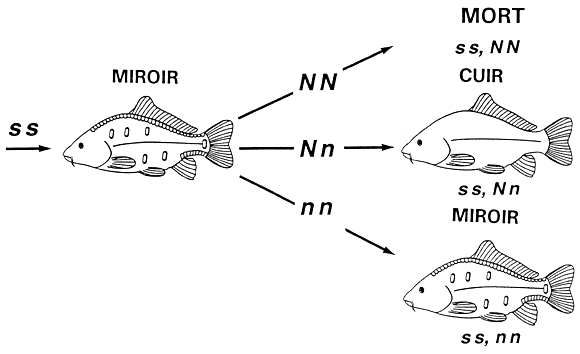
Figure 6. Transmission héréditaire de la disposition des écailles chez la carpe commune. La disposition des écailles est déterminée par l'interaction épistatique des gènes S et N. Le gène S contrôle l'expression du phénotype “à écailles” (génotypes SS et Ss) et du phénotype “miroir” (génotype ss). Le gêne N modifie ces phénotypes. Le génotype NN provoque la mort du poisson (génotypes SS,NN, Ss,NN et ss,NN). Le génotype Nn transforme le phénotype “à écailles” en phénotype “à ligne” (génotypes SS,Nn et SS,Nn) et le phénotype “miroir” en phénotype “cuir” (génotype ss,Nn). Enfin, le génotype nn ne modifie pas les phénotypes produits par le gène S, de sorte que les poissons à écailles ont un génotype SS,nn ou Ss,nn, alors que les poissons miroirs ont un génotype ss.nn.
Les phénotypes quantitatifs sont les phénotypes que l'on mesure, comme la longueur, le poids, le nombre d'oeufs par kg de femelle ou l'indice de transformation. Ils diffèrent des phénotypes qualitatifs en cela que les individus ne se répartissent pas en catégories discrètes non chevauchantes. Lorsqu'un généticien décrit un phénotype quantitatif, il crée une catégorie unique (le poids, par exemple). Les poissons ne sont pas regroupés en catégories discrètes telles que “légers” ou “lourds”, mais les individus sont plutôt rangés dans un continuum, leur valeur phénotypique étant déterminée par l'unité de mesure employée par le pisciculteur (le millimètre, le centimètre, le gramme, le kilogramme, etc.).
Comme la valeur phénotypique des individus s'exprime par une mesure (la longueur en millimètres, par exemple) plutôt que par une catégorie descriptive (la couleur, par exemple), les différences entre deux individus sont davantage une question de degré (millimètres) que de genre (la couleur). De ce fait, dans une population donnée, les phénotypes quantitatifs constituent ce qu'on appelle des distributions continues, que l'on peut décrire au moyen de graphiques (figure 7).

Figure 7. Distribution d'un phénotype quantitatif dans une population donnée. Le graphique a illustre une distribution parfaite, qui se traduit par une “courbe en cloche” dont le sommet coïncide avec la moyenne. Le graphique b correspond à la distribution de la longueur à 7 mois relevée dans une population de carpes communes.
C'est du fait de leur complexité génétique beaucoup plus grande que les phénotypes quantitatifs, au contraire des phénotypes qualitatifs, ne permettent pas de répartir les individus dans des catégories nettes et précises. En effet, alors que les phénotypes qualitatifs sont généralement contrôlés par un ou deux gènes (très peu d'entre eux sont contrôlés par plus de deux gènes), les phénotypes quantitatifs sont contrôlés par des douzaines à des centaines de gènes. On en connaît d'ailleurs rarement le nombre exact. De plus, pendant la méiose, les gènes sont battus comme un jeu de cartes par suite des phénomènes d'enjambement et de disjonction indépendante des chromosomes; par suite de la combinaison des phénomènes, chaque descendance est assurée de recevoir un message génétique légèrement différent.
Les phénotypes quantitatifs subissent aussi la forte influence de diverses variables d'environnement, ce qui participe d'ailleurs à leur distribution continue. Parmi ces variables, si certaines vont de soi, comme la densité d'empoissonnement, d'autres sont souvent négligées, comme la taille et l'âge de la mère. Certaines de ces variables interviennent à l'échelon familial (la date de naissance et l'âge de la mère, par exemple), alors que d'autres concernent au premier chef l'individu (l'accès à la nourriture, par exemple).
L'influence simultanée des facteurs génétiques et de ces facteurs du milieu aboutit à la création de catégories phénotypiques uniques, où le seul moyen de décrire un individu consiste à le mesurer. Les phénotypes quantitatifs correspondant à des catégories uniques à distribution continue, il n'est pas possible d'analyser des rapports ou de déterminer quelle fraction d'une population possède un phénotype particulier, comme on pouvait le faire dans le cas des phénotypes qualitatifs. Au lieu de cela, on calcule des valeurs de population et on les compare aux valeurs phénotypiques individuelles ou familiales. Dans une population donnée, les phénotypes quantitatifs sont décrits par la moyenne (il s'agit de la moyenne arithmétique) et l'écart-type (la racine carrée de la variance). La moyenne indique la tendance centrale et l'écart-type précise comment les valeurs relatives à la population se répartissent de part et d'autre de cette moyenne.
Pour mener à bien les tâches pratiques d'amélioration génétique dans des exploitations piscicoles de moyenne importance, il importe de savoir calculer la moyenne, de façon à pouvoir évaluer les résultats des programmes d'élevage sélectif mis en oeuvre. Quant à l'écart-type, à l'inverse des scientifiques et des chercheurs, les pisciculteurs ne sont pas tenus de savoir le calculer. Le tableau 3 indique la méthode de calcul de la moyenne.
Les phénotypes quantitatifs étant contrôlés par des dizaines à des centaines de gènes, l'expression simultanée et/ou séquentielle de ces gènes ne permet pas de procéder à leur identification précise et de déchiffrer leur mode de transmission héréditaire. Il faut par conséquent adopter une autre démarche pour parvenir à utiliser et à comprendre les phénotypes quantitatifs. Compte tenu de leur complexité génétique, ces phénotypes sont d'un usage beaucoup plus difficile que les phénotypes qualitatifs; ce sont pourtant eux (et notamment le poids et le taux de fécondité) qui jouent le premier rôle en agriculture et en aquiculture, et la valeur génétique d'une population de poissons comestibles d'élevage est essentiellement fonction des gènes qui les contrôlent. Cette importance des phénotypes quantitatifs est mise en évidence par le fait qu'on les appelle souvent “phénotypes de production”.
Tableau 3. Méthode de calcul de la moyenne relative à un phénotype quantitatif. Il s'agit en ce cas de la longueur moyenne. On utilise en général d'un échantillon aléatoire de 30 à 200 poissons.
| Etape 1. | Mesurer la longueur des individus (30 poissons en l'occurrence) au millilètre près. | |||||||||
| 98 | 103 | 106 | 111 | 104 | 91 | 87 | 114 | 103 | 107 | |
| 101 | 104 | 97 | 105 | 108 | 100 | 110 | 104 | 113 | 105 | |
| 95 | 97 | 107 | 108 | 99 | 111 | 112 | 105 | 113 | 103 | |
| Etape 2. | Faire la somme des mesures, c'est-à-dire des valeurs phénotypiques. | |||||||||
| 98 + 103 + 106 + 111 + 104 + 91 + 87 + 114 + 103 + 107 + 101 + 104 + 97 + 105 + 108 + 100 + 103 + 113 + 105 + 95 + 97 + 107 + 108 + 99 + 111 + 112 + 105 + 113 + 103 = 3,120 | ||||||||||
| Etape 3. | Diviser le total obtenu à l'étape 2 par le nombre de poissons mesurés (30 en ce cas). | |||||||||
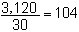 | ||||||||||
| La longueur moyenne de la population est de 104 mm. | ||||||||||
Attendu que les phénotypes quantitatifs relatifs à une population présentent une distribution continue, la seule façon d'utiliser et d'améliorer ces caractères consiste à analyser leur variance et à y distinguer une part héritable et une part non héritable. La variance phénotypique consiste dans la variabilité que présente un phénotype dans une population donnée; la moyenne correspond à la valeur phénotypique moyenne, alors que la variance décrit la façon dont les individus sont répartis de part et d'autre de cette moyenne (l'écart-type, déjà mentionné précédemment, est la racine carrée de la variance). La variance phénotypique (Vp) est la somme de trois éléments : la variance génétique (VG), la variance d'environnement (VE) et la variance d'interaction des facteurs génétiques et de l'environnement VG-E). On peut la représenter au moyen de la formule suivante :
VP = VG + VE + VG-E
La variance génétique est évidemment l'élément sur lequel les sélectionneurs essaient d'agir dans le cadre des programmes d'amélioration. Cette forme de variance est elle-même la somme de trois composantes qui ne peuvent être exploitées qu'à l'aide de programmes d'amélioration génétique différents.
La variance génétique (VG) est la somme de la variance génétique additive (VA), de la variance génétique de dominance (VD) et de la variance génétique épistatique (V,). Comme ci-dessus, on peut la représenter par la formule :
VG = VA + VD + Vl
Les termes “additive”, “dominance” et “épistatique” ne font pas référence à des sortes particulières d'action génétique comme cela était le cas pour les modes de transmission héréditaire concernant les phénotypes qualitatifs. Les expressions correctes, à savoir “variance génétique additive”, “variance génétique de dominance” et “variance génétique épistatique” (et non pas action génétique), renvoient à des éléments précis de la variance, qui relèvent du génome entier et non pas d'un ou deux gènes seulement.
La variance génétique additive est l'élément génétique dû aux effets additifs de l'ensemble des allèles du poisson considéré. Autrement dit, c'est la somme des valeurs correspondant à la part prise par chacun des allèles dans l'expression du phénotype. Certains allèles jouent un rôle important, d'autres, un rôle moindre, d'autres encore ne prennent aucune part dans cette expression et d'autres enfin exercent même une influence négative. On additionne alors l'ensemble des valeurs correspondant aux parts prises respectivement par les divers allèles et on obtient ainsi la variance génétique additive propre au poisson considéré.
La variance génétique de dominance est l'élément génétique dû à l'interaction des deux allèles occupant chacun des loci. De ce fait, cette variance ne peut être transmise de façon héréditaire.
Même s'il peut paraître assez déroutant que certaines formes du patrimoine génétique ne soient pas héritables, il s'agit cependant d'une notion simple. La variance génétique de dominance résulte de l'interaction des deux allèles occupant chacun des loci et est par conséquent subordonnée à l'état diploïde (2n). Si elle n'est pas héritable, c'est tout simplement parce que chacun des parents participe au processus de reproduction par le biais d'un gamète haploïde (n). Les gamètes ne contiennent pas de paires d'allèles (2n), et l'état diploïde fait place à l'état haploïde pendant la méiose. Lors de la division réductionnelle, la disjonction indépendante des chromosomes provoque la séparation de l'ensemble des paires alléliques, ce qui explique la destruction, pendant la méiose, des effets génétiques de dominance propres à chacun des parents. La variance génétique de dominance propre à un individu étant détruite au cours de la division réductionnelle, elle ne peut donc être transmise à la descendance par les gamètes et n'est donc pas héritable. Les effets liés à cette sorte de variance réapparaissent au moment de la fécondation, lorsqu'un spermatozoïde haploïde féconde un ovule haploïde pour former un zygote diploïde. Après fécondation, les gènes sont de nouveau associés par paires, ce qui suscite une interaction entre les deux allèles occupant chacun des loci et, par conséquent, la réapparition de la variance génétique de dominance. C'est ainsi que les effets liés à cette sorte de variance disparaissent et réapparaissent sous des formes différentes à chaque génération.
La variance génétique épistatique est l'élément génétique dû à l'interaction des allèles occupant des loci différents (et non pas au sein d'une même paire d'allèles). C'est un mélange de variance héritable et non héritable. La part de l'interaction des allèles présents dans le gamète fécondant est héritable, alors que la part de l'interaction impliquant les allèles répartis dans les autres spermatozoïdes ou dans les globules polaires ne l'est évidemment pas. La fraction de la variance génétique épistatique qui est héritable varie en outre d'un gamète à l'autre en raison des phénomène d'enjambement chromosomique et de disjonction indépendante. Ces phénomènes ont d'ailleurs tendance à effacer la plus grande partie de la variance génétique épistatique pendant la méiose, de sorte qu'un petit échantillon aléatoire seulement de cette variance est transmis du parent à sa descendance; en conséquence, seule une petite fraction aléatoire de la variance génétique épistatique est héritable.
Au plan pratique, il importe de savoir ce qui distingue la variance génétique additive, la variance génétique de dominance et la variance génétique épistatique et de comprendre comment elles sont transmises, car on utilise des programmes d'amélioration de différents types pour tirer parti de ces divers éléments de la variance génétique. De plus, le choix du programme d'amélioration à mettre en oeuvre et l'efficacité avec laquelle il permettra d'améliorer le phénotype sont subordonnés aux fractions respectives de la variance phénotypique que l'on peut attribuer à ces divers éléments.
Les deux éléments principaux sont la variance génétique additive et la variance génétique de dominance. La plupart des sélectionneurs prennent pour hypothèse que la variance génétique épistatique joue un rôle accessoire. Cette hypothèse est fondée sur le fait qu'il est difficile de sélectionner des combinaisons d'allèles lorsqu'on ne sait pas quelles sont les combinaisons souhaitables. En outre, l'amélioration qu'on peut attendre d'une sélection portant sur des effets épistatiques est généralement limitée et plafonne rapidement.
La variance génétique additive et la variance génétique de dominance sont par essence opposées. La première est liée à des allèles particuliers, et donc au stade haploïde, alors que la seconde est associée aux paires alléliques, et donc au stade diploîde. Un parent, parce qu'il produit des gamètes haploïdes, peut transmettre les effets génétiques additifs qui lui sont propres à sa descendance, alors qu'il n'est pas en mesure de transmettre les effets de dominance, qui sont annihilés pendant la méiose. Ces derniers effets sont produits dans chaque zygote après fécondation. Ainsi, les effets additifs dépendent de chacun des parents, alors que les effets de dominance sont fonction de croisements particuliers. Les effets additifs étant transmis par un parent à sa descendance, on qualifie souvent la variance génétique additive de “variance des valeurs génétiques”.
La variance génétique additive étant transmissible d'une génération à l'autre, on a recours à la sélection pour tirer parti de cet élément de la variance et améliorer ainsi la population considérée, alors qu'on a recours à l'hybridation dans le cas de la variance génétique de dominance du fait que cet élément de la variance n'est pas héritable et est fonction du croisement impliqué.
Héritabilité. La variance génétique additive pouvant être transmise par un parent à sa descendance de manière prévisible et sûre, il suffit de connaître la part en pourcentage de la variance phénotypique qui est imputable à la variance génétique additive pour être en mesure de prévoir le degré d'amélioration que l'on peut attendre de la sélection. Le sélectionneur a même la possibilité de pratiquer une sélection sur mesure afin d'obtenir un degré d'amélioration déterminé à l'avance par génération.
La part proportionnelle de la variance génétique additive est appelée “héritabilité” et peut être représentée à l'aide de la formule suivante :
h2 = VA/VP
où h2 est le symbole de l'héritabilité, VA, la variance génétique additive et VP, la variance phénotypique. L'héritabilité, qui s'exprime en pourcentage (de 0 à 100 % ou de 0,0 à 1,0), quantifie la fraction de la variance phénotypique qui est transmise héréditairement d'une façon prévisible et sûre.
La détermination de l'héritabilité d'un phénotype quantitatif sert en premier lieu à prévoir les résultats d'un programme d'élevage sélectif donné à l'aide de la formule :
R = Sh2
où R est le résultat de la sélection (gain par génération), S, le différentiel de sélection (la supériorité des géniteurs sélectionnés par rapport à la moyenne relative à la population, obtenue simplement en soustrayant cette moyenne de la moyenne relative aux géniteurs sélectionnés) et h2, l'héritabilité. Le tableau 4 indique comment on peut utiliser l'héritabilité pour prévoir le résultat de la sélection.
Tableau 4. Méthode de prévision du résultat de la sélection (l'héritabilité h2 du phénotype étant connue). Dans le présent exemple, on prévoit le résultat d'une sélection visant à accroître la longueur et l'on calcule ensuite la longueur moyenne prévue des individus de la prochaine génération.
| Soit: | h2 pour la longueur à 12 mois = 0,26 |
| longueur moyenne à 12 mois des individus de la population = 146 mm | |
| longueur moyenne à 12 mois des géniteurs sélectionnés = 162 mm | |
| Etape 1. | Calculer le différentiel de sélection (S). |
| S = longueur moyenne des géniteurs sélectionnés - longueur moyenne des individus de la population | |
| S = 162 mm - 146 mm = 16 mm | |
| Etape 2. | Calculer le résultat prévu de la sélection (R). |
| R = Sh2 | |
| R = (16 mm) (0,26) | |
| R = 4,16 mm | |
| Etape 3. | Calculer la moyenne de la génération F, de poissons sélectionnés. |
| moyenne de la génération F, = moyenne de la population + R = 146 mm + 4,16 mm = 150,16 mm |
La formule précédente démontre clairement que l'héritabilité est le facteur qui permet de déterminer la fraction en pourcentage du différentiel de sélection qui peut être gagnée par sélection, autrement dit quelle amélioration on peut espérer. En général, une héritabilité supérieure à 0,25 indique que la sélection procurera une amélioration importante, alors qu'une héritabilité inférieure à 15 % indique que la sélection sera inefficace. Une héritabilité supérieure ou égale à 0,3 est considérée comme forte.
Bien qu'il soit préférable de connaître l'héritabilité d'un phénotype quantitatif avant de mettre en oeuvre un programme d'élevage sélectif, cela n'est portant pas indispensable. Si l'on dispose de cette valeur, on peut s'en servir pour prévoir l'amélioration, choisir le différentiel de sélection nécessaire pour obtenir le résultat souhaité ou décider d'abandonner le programme, vu son inefficacité prévisible. Le tableau 5 indique comment utiliser l'héritabilité pour ajuster le différentiel de sélection en vue d'obtenir les résultats souhaités.
II est ouvent inutile de déterminer l'héritabilité, car il existe déjà des informations publiées sur le sujet. A ce jour, on a déterminé plusieurs centaines de valeurs de l'héritabilité chez les principales espèces de poissons comestibles d'élevage pour des phénotypes tels que le taux de croissance, l'indice de transformation, la résistance aux maladies, la fécondité, la taille des oeufs, le nombre d'oeufs, le rendement de carcasse, la conformation du corps ou la tolérance aux pesticides. Quoique les valeurs publiées puissent différer des valeurs relevées dans une population piscicole donnée (ces valeurs sont en effet propres à la population qui a fait l'objet d'une évaluation et aux conditions d'élevage utilisées pour l'expérience), elles sont généralement très proches de celles que l'on observe dans la plupart des populations. Le tableau 6 présente un certain nombre de valeurs de l'héritabilité qui ont été établie chez la carpe commune et le tilapia.
Tableau 5. Mode d'utilisation de l'héritabilité (h2) d'un phénotype aux fins d'ajustement du différentiel de sélection en vue d'obtenir le résultat souhaité.
| Soit : | h2 pour la longueur à 12 mois = 0,26 |
| longueur moyenne à 12 mois des individus constituant la population = 146 mm | |
| résultat souhaité de la sélection = 6 mm | |
| Etape 1. | Calculer le différentiel de sélection (S) nécessaire pour obtenir un résultat de 6 mm. |
| R = Sh2 | |
| 6 mm = S(0,26) | |
| S = 6 mm/0,26 = 23,08 mm | |
| Etape 2. | Calculer la longueur moyenne à 12 mois des géniteurs sélectionnés qui permettra d'obtenir un différentiel de sélection de 23,08 mm. |
| S = longueur moyenne des géniteurs sélectionnés - longueur moyenne des individus de la population | |
| 23,08 mm = longueur moyenne des géniteurs sélectionnés - 146 mm | |
| longueur moyenne des géniteurs sélectionnés = 146 mm + 23,08 mm = 169,08 mm |
Bien que la production des phénotypes soit fondée sur les plans d'exécution fournis par les gènes, elle est néanmoins conditionnée en partie par le milieu. Si ce dernier influe sur la production de tous les phénotypes, les phénotypes quantitatifs subissent cependant davantage l'influence des variables d'environnement que les phénotypes qualitatifs. Si le poisson ne dispose pas des éléments nutritifs indispensables, il ne parviendra pas à produire certaines protéines, et donc à exprimer certains phénotypes particuliers. C'est en particulier le cas des phénotypes qualitatifs qui sont tributaires de pigments que le poisson n'est pas en mesure de synthétiser. Par exemple, les éleveurs de poissons tropicaux ajoutent divers pigments végétaux aux aliments de façon à aviver l'éclat des couleurs de ces poissons, et les éleveurs de saumons ajoutent également des pigments aux aliments pour que la chair rosisse. Parmi les facteurs du milieu qui influent sur les phénotypes quantitatifs, certains vont de soi, comme la densité d'empoissonnement ou la qualité des aliments, alors que d'autres, moins évidents, sont rarement pris en considération (âge et taille des femelles, moment du frai, taille des particules d'aliments, pratiques alimentaires, etc.). Lorsqu'on met en train un programme d'élevage sélectif en vue d'améliorer un phénotype quantitatif, il faut impérativement maîtriser les variables d'environnement et les empêcher de fluctuer selon les individus, les familles ou les étangs. En l'absence d'une telle maîtrise, ces variables ne manqueraient pas d'exercer une influence différente sur l'expression phénotypique, et le pisciculteur ne saurait pas si les poissons qu'il a sélectionné sont les meilleurs du fait de leur supériorité génétique ou de la qualité exceptionnelle du milieu. La question est cruciale, car seuls les poissons supérieurs au plan génétique ont le pouvoir de transmettre cette supériorité à leur descendance, ce qui constitue l'objectif de tout programme d'élevage sélectif.
Tableau 6. Héritabilité (h2) de différents phénotypes chez la carpe commune, le tilapia du Nil, le tilapia bleu et le tilapia du Mozambique. L'indication de différentes valeurs de l'héritabilité pour un même phénotype (par exemple pour le poids à 1 an chez la carpe commune) est dû au fait que la valeur obtenue diffère selon le chercheur qui a procédé aux mesures, la population considérée ou les conditions de croissance. Une héritabilité réalisée est une héritabilité qui a été déterminée à l'issue d'un programme d'élevage sélectif.
| Espèce | Phénotype | H2 |
| Carpe commune | poids à 1 an | 0.0 |
| poids à 1 an | 0.34 | |
| poids à 2 ans | 0.49 | |
| poids à 2 ans | 0.15 | |
| poids à 3 ans | 0.60 | |
| poids à 4 ans | 0.24 | |
| gain pondéral (réalisé) | 0.21 | |
| gain pondéral | 0.0 | |
| longueur à 1 an | 0.25 | |
| longueur à 1 an | 0.04 | |
| longueur à 2 ans | 0.34 | |
| largeur du corps à 1 an | 0.55 | |
| largeur du corps à 2 ans | 0.42 | |
| largeur du corps à 3 ans | 0.69 | |
| forme du corps (longueur/poids) (réalisée) | 0.47 | |
| teneur en matières grasses | 0.47 | |
| 0.14 | ||
| Tilapia du Nil | poids à 4 semaines | 0.0 |
| poids à 4 semaines | 0.06 | |
| poids à 46 jours | 0.04 | |
| poids à 8 semaines | 0.0 | |
| poids à 8 semaines | 0.21 | |
| poids à 10 semaines | 0.0 | |
| poids à 10 semaines | 0.46 | |
| poids à 90 jours | 0.04 | |
| poids à 136 jours (femelles) | 0.71 | |
| poids à 136 jours (femelles) | 0.37 | |
| poids à 136 jours (mâles) | 0.71 | |
| poids à 136 jours (mâles) | 0.30 | |
| poids à 7 mois (réalisé) | 0.06 | |
| longueur à 45 jours | 0.10 | |
| longueur à 90 jours | 0.06 | |
| fécondité au premier frai | 0.0 | |
| fécondité au premier frai | 0.09 | |
| Tilapia bleu | gain pondéral à 40 semaines chez les femelles (réalisé) | 0.38 |
| gain pondéral à 40 semaines chez les mâles (réalisé) | 0.20 | |
| gain pondéral à 49 semaines chez les femelles (réalisé) | 0.10 | |
| gain pondéral à 49 semaines chez les mâles (réalisé) | 0.27 | |
| gain de longueur à 40 semaines chez les femelles (réalisé) | 0.87 | |
| gain de longueur à 40 semaines chez les mâles (réalisé) | 0.40 | |
| Tilapia du Mozambique | poids à 5 mois des femelles (réalisé) | 0.01 |
| poids à 5 mois des mâles (réalisé) | 0.10 |
![]()
![]()
![]()