![]()
![]()
![]()
Par phénotypes quantitatifs, on entend les importants caractères “de production” tels que la longueur, le poids, l'indice de transformation ou le nombre d'oeufs par kg de poisson femelle. Pour ce qui est de ces caractères, l'exploitant ne peut classer les poissons en différentes catégories descriptives non chevauchantes comme dans le cas des phénotypes qualitatifs (par exemple les catégories “couleur rouge” et “pigmentation normale”). Chaque caractère correspond à un seul phénotype (la longueur, par exemple), et l'on détermine les différences entre individus en mesurant les poissons. La valeur numérique que l'on assigne à chaque poisson mesuré est une valeur phénotypique propre à l'individu considéré.
La valeur phénotypique procédant de la mesure (par exemple la longueur en millimètres) plutôt que d'un classement en catégories descriptives (par exemple la couleur), la sélection se fonde sur des valeurs numériques, et non pas sur l'aspect. Ainsi, lorsqu'un exploitant souhaite faire porter la sélection sur un phénotype qualitatif tel que la couleur, il épargne les poissons qui présentent une couleur particulière et réforme les autres; la décision d'épargner ou de réformer est uniquement fondée sur l'appartenance du poisson à telle ou telle catégorie descriptive. Par contre, lorsqu'il s'agit de faire porter la sélection sur un phénotype quantitatif, l'exploitant épargne les poissons dont la valeur mesurée est supérieure ou égale à une valeur numérique précise et réforme les autres; la décision de sélectionner ou de réformer est uniquement fondée sur des valeurs numériques.
Les programmes d'élevage sélectif portant sur des phénotypes qualitatifs visent à créer des populations génétiquement pures. Ce sont des programmes simples destinés à éliminer un allèle indésirable et à fixer l'allèle désiré, de façon à obtenir une population génétiquement pure qui produise uniquement le phénotype souhaité. Ce résultat est d'ordinaire obtenu en une seule génération (souvent en un seul jour de sélection); cela fait, le but de l'exploitant est atteint et le programme d'amélioration prend fin.
A l'inverse, les programmes d'élevage sélectif qui servent à améliorer des phénotypes quantitatifs sont plus complexes. Ils nécessitent des techniques plus élaborées et obligent à relever davantage de données. De plus, ils ne sont pas limités dans le temps et se poursuivent année après année, du fait qu'il est impossible d'obtenir des populations génétiquement pures dans le cas de ces phénotypes. Ces programmes ont au contraire pour objet d'améliorer graduellement la moyenne relative à la population à chaque génération. Il ne faut jamais oublier que les programmes d'élevage sélectif portant sur des phénotypes quantitatifs ne sont jamais finis et qu'il s'agit donc d'obtenir des améliorations progressives et limitées sur de nombreuses générations. Il est possible d'enregistrer des améliorations spectaculaires en une ou deux générations, mais cela oblige à réduire considérablement l'effectif de la population considérée, ce qui peut entraîner une dégénérescence par consanguinité susceptible d'annuler une bonne part des progrès résultant de la sélection.
Si les programmes d'élevage sélectif destinés à améliorer des caractères quantitatifs ne sont pas limités dans le temps, c'est aussi parce que leur abandon signifierait la perte d'une bonne part des gains obtenus. Il est difficile d'évaluer l'importance de cette perte, qui est fonction du caractère considéré, de son héritabilité et de la population concernée. Si le phénotype est uniquement subordonné à la variance génétique additive (h2 = 1,0), la moyenne ne variera pas en cas d'interruption du programme. Aucun des principaux phénotypes de production n'est toutefois subordonné à la seule variance génétique additive (tableau 6). En fait, l'abandon d'un programme d'élevage sélectif entraînera l'annulation d'une grande partie des améliorations obtenues, car cela signifiera également l'annihilation de la plus grande part des gains liés à la variance génétique de dominance, de la totalité des gains associés à la variance génétique épistatique et d'une partie des gains relevant des interactions des facteurs génétiques et des facteurs du milieu. Une partie des progrès attribuables à la variance génétique additive seront préservés, mais il est difficile d'en prévoir l'ampleur exacte.
Si le programme d'élevage sélectif a donné lieu à une amélioration considérable de la population avant son abandon, il se peut que la moyenne se stabilise à un niveau nettement supérieur au niveau initial. Si, par contre, l'amélioration a été minime (ce qui est généralement le cas), la moyenne finale sera probablement de très peu supérieure à la moyenne initiale, et cet écart sera en outre souvent masqué par les variations annuelles des facteurs du milieu.
Avant de mettre en train un programme d'élevage sélectif, tout exploitant devrait se poser la question suivante : ce genre de programme est-il approprié ou nécessaire? La réponse à cette question est tributaire des circonstances; elle peut différer d'un exploitant à l'autre, même voisins.
La décision de mettre en oeuvre ou non un programme d'élevage sélectif doit être prise à titre individuel, et en aucun cas à l'échelon régional. Les agents de vulgarisation ont souvent tendance à inciter l'ensemble des exploitants de la région qu'ils couvrent à adopter des techniques nouvelles. En théorie, cela peut sembler judicieux, car les agents savent qu'en cas d'adoption de ces techniques, la production augmentera, les revenus des exploitants progresseront et l'alimentation sera mieux assurée.
En fait, un programme d'élevage sélectif, comme tout processus technique, doit être mis en oeuvre sur une base individuelle. Dans la plupart des cas, ces programmes devraient être réservés aux exploitants qui sont en mesure de tenir des registres, d'assurer une gestion convenable et d'adopter durablement de nouvelles techniques et qui se montrent en outre désireux d'intégrer une certaine planification à long terme dans leurs programmes de gestion. De plus, ces exploitants doivent être prêts à consacrer le temps et l'argent nécessaires à la mise en oeuvre d'un tel programme.
Certains pisciculteurs n'auront jamais recours - ou ne devraient jamais être incités à recourir - à la sélection. Ce sont ceux qui ont de petites exploitations ou qui ne produisent du poisson que pour quelques familles, voire un petit village. Les exploitants qui sont incapables de tenir convenablement un registre, qui ne dispose pas d'un nombre suffisant d'étangs ou qui ne peuvent ou ne veulent consacrer l'argent nécessaire à la mise en oeuvre d'un programme d'amélioration ne devrait pas se lancer dans ce genre d'entreprise, ni d'ailleurs y être incités.
Les programmes d'élevage sélectif sont conçus en vue d'obtenir une amélioration de la productivité sur plusieurs générations (une génération correspond au remplacement des géniteurs par leur descendance) et d'augmenter graduellement le taux de croissance et le rendement sur une période de plusieurs années. Si un exploitant est incapable d'intégrer une certaine planification à long terme (1 à 10 ans) dans son programme piscicole, il interrompra le programme d'élevage sélectif au bout de quelques mois en invoquant son inefficacité, ce qui découragera les autres exploitants de se lancer dans l'aventure.
Les exploitants qui ne peuvent tenir un registre convenablement parce qu'ils ne savent pas lire ou écrire ou qui ignorent les techniques de mesure des poissons ne sont guère aptes à devenir des sélectionneurs et à exécuter un programme d'élevage sélectif. En général, ils en sont tout bonnement incapables. Toutefois, si ce sont des pisciculteurs sérieux qui travaillent sans compter et qui acceptent et adoptent les techniques nouvelles, ils peuvent mettre en train un programme d'élevage sélectif, dans la mesure où un agent de vulgarisation est disposé à leur prêter main-forte et à prendre en charge la tenue des registres et la conduite d'une bonne partie des travaux qu'occasionne un tel programme.
Enfin, les exploitants qui n'ont aucun aptitude à la gestion ne devraient jamais mettre en oeuvre un programme d'élevage sélectif; il en va ainsi de tous ceux qui inspectent rarement leurs étangs, qui nourrissent leurs poissons ou fertilisent leurs étangs uniquement lorsque l'agent de vulgarisation le leur rappelle ou qui ne font guère d'efforts pour accroître le rendement. Tout exploitant qui est incapable de gérer convenablement un étang à pisciculture ne sera pas davantage capable de gérer un programme d'élevage sélectif.
La plupart des programmes d'élevage sélectif concernant des phénotypes quantitatifs ont pour objectif premier d'améliorer le taux de croissance et, par conséquent, le rendement.
Il existe deux grands moyens d'intensifier le rendement. Le premier consiste à recourir à un programme d'amélioration génétique visant à améliorer les poissons qui sont élevés. Le deuxième consiste à améliorer les méthodes d'élevage. Or, les programmes d'amélioration génétique coûtent cher, alors que la modification des méthodes de gestion est souvent peu coûteuse.
En conséquence, s'il est facile d'augmenter le taux de croissance et le rendement en modifiant les techniques de gestion ou en en adoptant de nouvelles, les exploitants devraient en premier lieu privilégier cette démarche. Il s'agit, entre autres, de faire un usage approprié des engrais et de la chaux, de surveiller la densité d'empoissonnement, de placer les jeunes poissons les mieux développés dans des étangs d'affinage et de nourrir les poissons avec des rations préparées.
Si l'amélioration des méthodes de gestion constitue le tout premier moyen d'augmenter le rendement et le taux de croissance, c'est avant tout parce qu'elle ne coûte généralement pas cher et que les résultats sont immédiats. Il arrive que le rendement double ou triple lorsque les étangs sont convenablement fertilisés et que les poissons sont alimentés. Cet accroissement du rendement découlant de la fertilisation ou de l'alimentation des poissons se manifeste souvent dès la première période de croissance et la première récolte. Si les programmes d'élevage sélectif permettent également d'augmenter le rendement, cette augmentation s'effectue graduellement sur une période de plusieurs années. Pour cette seule raison, la toute première façon d'accroître la productivité devrait consister à améliorer les méthodes de gestion et les techniques d'élevage.
Par ailleurs, il n'est pas concevable de mettre en train un programme d'élevage sélectif et, une fois que le programme a débuté, de modifier les techniques d'élevage. Lorsqu'on opère une sélection portant sur des phénotypes quantitatifs, les phénotypes sélectionnés s'expriment dans des conditions d'élevage bien précises. Les poissons qui ont les meilleures performances dans des étangs fertilisés par fumure et à forte densité d'empoissonnement peuvent très bien différer de ceux qui ont les meilleures performances dans des étangs à faible densité d'empoissonnement où ils sont alimentés au moyen de rations préparées. Les poissons sélectionnés pour leur taux de croissance exceptionnel dans des conditions données contiennent des gènes qui leur permettent de se développer particulièrement bien dans ces conditions. En conséquence, si l'on modifie les méthodes d'élevage, il se peut que les gènes les mieux adaptés à ces nouvelles conditions aient déjà été réformés.
Il importe donc de s'assurer que les techniques de gestion qui sont employées pour produire du poisson dans une exploitation donnée ne seront pas modifiées ultérieurement. Si des modifications limitées (par exemple le fait de faire passer le taux d'empoissonnement de 2 000 à 2 500/ha dans des étangs d'affinage ou le poids moyen des alevins destinés au repeuplement de 10 à 15 g) ne devraient pas avoir de grosses conséquences, des modifications de plus grande ampleur (par exemple le fait de remplacer la fumure comme unique apport nutritif par des rations en granulés de qualité supérieure) auront une incidence considérable sur la gestion. Il convient d'ajourner tout programme d'élevage sélectif si d'importantes modifications sont planifiées ou prévues.
Il n'est pas nécessaire d'uniformiser les techniques d'élevage employées dans une région avant de mettre en oeuvre des programmes d'élevage sélectif. Il se peut en effet que certains exploitants préfèrent recourir à la fumure plutôt qu'aux aliments en granulés du fait qu'ils disposent de sources abondantes et bon marché de fumier, alors que d'autres préfèreront utiliser des rations en granulés afin de porter le rendement à son maximum. Il est possible de concevoir des programmes d'élevage sélectif adaptés à ces deux types d'exploitants, pour peu que ceux-ci soient décidés à consacrer à leur mise en oeuvre le temps et l'argent nécessaires ainsi qu'une superficie d'étang suffisante.
Quand doit-on mettre en oeuvre un programme d'élevage sélectif? Seulement lorsque l'exploitant a la volonté de consacrer le temps et les efforts nécessaires à son succès. Les exploitants qui ne sont pas disposés à déployer les efforts indispensables, que ce soit en matière de main-d'oeuvre ou d'investissements, ou qui risquent d'abandonner le projet passée la période d'enthousiasme initial devraient s'abstenir de mettre en train des programmes d'élevage sélectif.
Certains exploitants qui se disent intéressés par les programmes d'élevage sélectif ne se déclarent prêts à mettre en train un tel programme qu'à condition d'avoir au préalable réglé tous leurs autres problèmes. Cela signifie qu'ils ne mettront jamais en oeuvre un programme de ce genre, car ils ne parviendront jamais à résoudre la totalité des autres problèmes en question. Chaque fois qu'un problème est résolu, un autre obstacle à l'accroissement de la production surgit.
Pour mettre en oeuvre un programme d'élevage sélectif, l'exploitant doit être prêt à y consacrer le temps et l'argent nécessaires, et cela parce qu'il est convaincu que cette initiative lui permettra d'accroître sa production de denrées alimentaires et ses bénéfices et qu'il se rend par ailleurs compte qu'il s'agit là d'objectifs à long terme. Il doit enfin ne pas oublier que les programmes d'élevage sélectif ne sont pas limités dans le temps, du fait que des améliorations sont toujours possibles.
Avant de mettre en train un programme d'élevage sélectif, l'exploitant doit procéder à une évaluation destinée à confirmer son utilité. Il lui faut quantifier ses objectifs, connaître les techniques de mesure des phénotypes, définir le type de programme qu'il compte utiliser, savoir comment évaluer les résultats et prévoir une évaluation et une réévaluation annuelles du programme. A cet égard, il peut agir de son propre chef ou en consultation avec des agents de vulgarisation.
La première étape consiste à déterminer si le programme est nécessaire. Les promoteurs persuadent souvent les exploitants d'apporter à leur élevage des modifications en fait superflues. Il se peut qu'un vendeur ait beaucoup apprécié la forme du corps de poissons qu'il a vus dans une autre province et estime qu'il serait bon que les exploitants procèdent à une sélection portant sur ce phénotype. S'il est manifeste que les consommateurs apprécient eux aussi cette forme et que cela soit confirmé par les études de marché, les exploitants peuvent alors envisager de mettre en oeuvre un programme d'élevage sélectif destiné à modifier la forme du corps de leurs poissons. Si, par contre, les consommateurs locaux n'y prêtent pas attention et qu'il n'y ait pas de demande pour cette forme modifiée, les exploitants doivent renoncer à mettre un tel programme en oeuvre. Des programmes d'amélioration lancés sans discernement constituent un gaspillage d'argent et de main-d'oeuvre et peuvent même aller à l'encontre du but recherché. Avant de mettre en train un programme d'élevage sélectif, l'exploitant doit toujours tenir compte de ce que le consommateur veut et de ce qu'il est prêt à payer pour cela.
Une autre étape de l'évaluation consiste à identifier les caractères qui se prêtent à la sélection. L'exploitant doit dresser une liste de tous les caractères qu'il souhaite améliorer, puis conserver uniquement ceux qui paraissent appropriés sur le plan de l'élevage et de la commercialisation des poissons. Certains caractères tels que le poids corporel sont inclus dans presque tous les programmes d'élevage sélectif. D'autres, comme la résistance aux maladies ou la facilité de capture, sont souvent retenus simplement parce que l'exploitant souhaite, en les améliorant, faciliter sa tâche ou la rendre moins risquée.
Certains caractères sont tout à fait superflus et devraient être immédiatement écartés. C'est ainsi qu'une usine de transformation du poisson peut inciter les exploitants à améliorer le rendement de carcasse ou à réduire la teneur en matières grasses des viscères, car cela aura pour effet d'augmenter les bénéfices qu'en tirent ses propriétaires. Toutefois, à moins que ces derniers aient l'intention d'acheter plus cher le poisson doté de ces caractères, ceux-ci ne devraient pas être intégrés dans un programme d'élevage sélectif.
Les exploitants doivent aussi écarter les caractères qu'il est plus facile d'améliorer par le biais des techniques de gestion. Par exemple, il n'est guère judicieux de recourir à l'élevage sélectif pour accroître la fécondité, car il est beaucoup plus simple et nettement moins onéreux d'augmenter la production d'oeufs en modifiant la gestion des écloseries. De plus, la plupart des poissons sont déjà extrêmement fécond. Faire passer la production d'oeufs de 4 000 à 4 500 oeufs/kg de femelle n'est vraiment ni primordial, ni nécessaire. Si l'exploitant souhaite produire plus d'oeufs, il lui suffit de faire frayer davantage de femelles ou des femelles plus grosses. L'amélioration de la production d'oeufs au moyen de l'élevage sélectif est coûteuse, et ses effets ne se font pas sentir pendant au moins une génération. A l'inverse, l'amélioration de la production d'oeufs par le biais de la modification de la gestion des écloseries est peu coûteuse et ses effets sont immédiatement perceptibles.
Les exploitants doivent s'efforcer d'améliorer uniquement les phénotypes qui ont les effets les plus positifs sur leurs bénéfices, soit parce que les poissons se développent plus vite ou plus efficacement, soit parce qu'ils ont un taux de survie supérieur, soit encore parce que les consommateurs sont disposés à les acheter plus cher.
Les responsables d'écloseries qui travaillent dans des centres de production de jeunes poissons peuvent envisager de procéder à une sélection portant sur des caractères susceptibles de faciliter la pisciculture ou d'en élargir le champ d'activité. C'est ainsi que le renforcement de la tolérance à la salinité peut permettre aux pisciculteurs d'élever des poissons dans des milliers d'hectares d'étangs d'eau saumâtre; par ailleurs, une sélection favorisant un frai précoce peut leur permettre de produire du poisson de taille marchande en une seule saison de croissance (et non en deux) et d'obtenir des poissons plus gros se vendant à un prix avantageux.
L'étape suivante de l'évaluation consiste, pour l'exploitant, à décider s'il produit des jeunes poissons génétiquement améliorés pour son propre usage ou s'il se spécialise dans l'élevage de jeunes poissons de ce genre en vue de leur distribution aux exploitations piscicoles locales. De cette décision dépendent en effet les montants qui doivent être investis dans le programme d'élevage sélectif ainsi que l'ampleur de ce dernier. Si l'exploitant entend réserver les poissons génétiquement améliorés qu'il produit à son propre usage, il n'aura besoin d'utiliser qu'une partie seulement de son exploitation pour mener à bien son projet. Au contraire, s'il souhaite produire de tels jeunes poissons pour les autres établissements locaux de pisciculture, il pourra être amené à destiner la plus grande partie, voire la totalité, de son exploitation à cette fin.
Si l'on cherche à ne retenir que quelques phénotypes parmi tous ceux qui apparaissent sur la liste initiale, c'est aussi en grande partie parce que l'efficacité du processus d'amélioration d'un caractère est inversement proportionnelle au nombre de phénotypes qui sont intégrés dans un programme d'élevage sélectif. En conséquence, s'il apparaît qu'un ou deux phénotypes ont nettement plus d'importance que les autres, ce sont seulement ceux-là qui devraient être retenus. Le maintien sur la liste d'autres caractères plus secondaires aura pour effet de ralentir le rythme d'amélioration des phénotypes importants. C'est pourquoi la plupart des programmes d'élevage sélectif devraient se limiter à l'amélioration d'un ou deux phénotypes. Certains programmes peuvent en inclure un troisième, et la mise en oeuvre des rares programmes qui en intègrent davantage devrait être réservée aux généticiens des stations de recherche.
Une fois que l'exploitant a réduit sa liste initiale et qu'il ne reste plus que le ou les deux phénotypes qu'il souhaite améliorer, il lui faut déterminer comment ces phénotypes seront définis et mesurés. Il convient de renoncer aux objectifs qui ne peuvent être définis avec précision et aux phénotypes qui sont difficiles à mesurer. Par exemple, tous les exploitants souhaitent améliorer la résistance aux maladies, mais que recouvre cette expression? Certains peuvent définir cette résistance comme le fait de ne pas tomber malade et d'autres, comme le fait de ne pas mourir. Ces définitions diffèrent considérablement. Si l'on définit le caractère comme le fait de ne pas tomber malade, qu'est-ce que cela signifie? Cela signifie-t-il que les poissons ont été infectés, mais ne présentent pas les signes cliniques de la maladie, ou qu'ils n'ont jamais été infectés? Si les poissons sont élevés dans le même étang, comment pourra-t-on distinguer les poissons infectés de ceux qui ne le sont pas? Quant à la définition se référant au fait de ne pas mourir, elle n'est pas non plus très précise, car la gestion sanitaire est tributaire de très nombreuses variables. Les poissons peuvent mourir pour des raisons tout autres qu'infectieuses lors d'un accès de maladie ou au cours d'une saison de croissance. Ce caractère étant difficile à mesurer, la plupart des exploitants devraient probablement renoncer à l'améliorer. De plus, si la résistance à une maladie particulière est héritable, elle se renforcera avec le temps sous l'effet de ce qu'on appelle la “domestication”. Il s'agit du processus par lequel l'exploitation et la gestion des exploitants soumettent naturellement les gènes des animaux à une sélection positive ou négative du simple fait de leur élevage. En conséquence, la résistance aux maladies est un caractère qui devrait disparaître des listes de la plupart des exploitants. Certes, quelques progrès ont été récemment accomplis en matière de sélection pour ce qui est de la résistance aux maladies, mais les programmes d'amélioration sont complexes et doivent être réservés aux scientifiques des stations de recherche.
Au nombre des autres caractères qui doivent être écartés figurent ceux que l'exploitant souhaite améliorer, mais qui sont difficiles à mesurer vu la façon dont les poissons sont élevés. Ainsi, les exploitants qui nourrissent leurs poissons avec des rations en granulés souhaitent améliorer l'indice de transformation, car cela permettrait aux poissons de transformer plus efficacement les aliments et entraînerait une forte progression du rendement et des bénéfices. Mais comment déterminer l'indice de transformation propre à chacun des poissons? Le seul moyen consiste à élever chaque poisson dans un bassin ou un hapa particulier, ce qui reviendrait extrêmement cher. Pour améliorer ce caractère, l'exploitant pourrait avoir recours à la sélection familiale, pour peu qu'il puisse élever des douzaines de familles dans différents hapas. Un tel programme serait fort coûteux, alors que l'amélioration de l'indice de transformation importe moins que celle du taux de croissance. Si le recours à des programmes d'élevage sélectif portant sur l'indice de transformation s'avère néanmoins indispensable, il faut laisser le soin de les mettre au point aux généticiens des stations de recherche.
Mêmes les caractères que tout exploitant souhaite améliorer et qui seront intégrés dans un programme d'élevage sélectif doivent être définis et mesurés avec précision. Tout exploitant souhaite améliorer le taux de croissance, mais comment ce taux doit-il être défini et comment sera-t-il mesuré? Cela a son importance, car c'est ce qui déterminera le degré d'amélioration qu'on peut attendre de la sélection. Le taux de croissance est généralement défini comme le poids ou la longueur du poisson à un âge donné; par conséquent, la première question qu'il faut se poser est la suivante : le taux de croissance doit-il être défini par le poids ou la longueur?
On peut mesurer sans difficulté des centaines de poissons au millimètre près, sans traumatisme notable pour ces derniers. Toutefois, la longueur comprend la longueur de la tête et de la queue. C'est ainsi que des poissons à queue trop longue pourraient être sélectionnés au détriment de poissons plus lourds; or, les consommateurs achètent leur poisson au kilogramme, et non pas au centimètre. Si le poids reflète plus exactement le mode d'achat et de vente du poisson, il est difficile de peser des centaines de poissons au dixième de gramme près. A moins que l'exploitant dispose d'une balance électronique, le processus est souvent très fastidieux et stressant pour le poisson. De plus, le poids noté pour chaque poisson comprend l'eau présente dans la bouche, les fèces et les aliments présents dans le tube digestif ainsi que le poids de la tête et des viscères, de sorte qu'il est lui aussi sujet à erreur.
Bien qu'il soit préférable d'évaluer le taux de croissance par le biais du poids, il existe une forte corrélation entre longueur et poids (autrement dit, les deux sont étroitement liés; lorsque l'un augmente, l'autre augmente aussi), de sorte qu'une évaluation de la croissance par le biais de la longueur constitue une bonne approximation de l'évaluation obtenue en mesurant le poids corporel. Si l'exploitant souhaite éliminer une partie de l'erreur liée à l'évaluation de la croissance par le biais de la longueur, il peut mesurer la longueur standard plutôt que la longueur totale, ce qui élimine l'effet de la longueur de la queue sur la sélection.
Si la mesure des poissons doit être effectuée par plus d'une personne, il convient de définir et de mettre au point à l'avance une méthode de mesure type qui permette d'obtenir des mesures uniformes. Sinon, les valeurs phénotypiques assignées aux poissons (leur longueur) seraient sujettes à l'erreur humaine. Si une seule personne mesure les poissons, il suffit qu'elle le fasse en employant toujours la même technique.
Si un processus de sélection portant sur la longueur contribue à améliorer le poids, on dit que le poids a été amélioré par “sélection indirecte”. Cela est dû au fait qu'il existe une corrélation positive (au plan génétique) entre ces deux phénotypes, de sorte que l'amélioration de l'un se traduit automatiquement par une amélioration de l'autre. De ce fait, toute sélection visant à améliorer le taux de croissance a souvent un effet positif sur d'autres caractères tels que le taux de survie, la résistance aux maladies ou l'indice de transformation. Les sélectionneurs tirent souvent parti de ces corrélations génétiques et, en faisant porter la sélection sur des phénotypes faciles à mesurer, profitent de la sélection indirecte pour améliorer d'autres phénotypes dont la mesure revient cher ou soulève des difficultés. Par exemple, le poids à la récolte des truites arc-en-ciel a progressé par sélection indirecte, alors que la sélection portait sur le poids des truitelles. Par ailleurs, on met en oeuvre des programmes d'élevage sélectif destinés à améliorer la résistance aux maladies par sélection indirecte, la sélection proprement dite concernant la réaction aux agressions et la résistance des membranes cellulaires.
Lorsqu'on cherche à améliorer le taux de croissance, il faut aussi se demander à quel âge il convient d'évaluer ce taux. La croissance des poissons n'est pas régulière. Si c'était le cas, l'âge auquel les mesures sont effectuées aurait relativement peu d'importance. Il semble judicieux de mesurer les poissons lorsqu'on les déplace ou qu'on les capture. Par exemple, on peut les mesurer lorsqu'on les transfère des étangs à alevins aux étangs d'affinage ou qu'on les capture pour les vendre. D'ordinaire, on parle à ce propos de mesure à un âge précis. Après un certain nombre de générations soumises à la sélection, on parle plus volontiers du temps nécessaire pour que le poisson atteigne sa taille marchande. Le fait de mesurer et de sélectionner les poissons à deux âges différents plutôt qu'à un seul aura pour effet d'accélérer l'amélioration du taux de croissance.
Il est indispensable de pouvoir définir précisément l'objectif poursuivi et d'être en mesure de mesurer avec exactitude le phénotype. Si l'exploitant est incapable d'accomplir ces tâches, il doit renoncer à mettre en oeuvre un programme d'élevage sélectif. En effet, s'il met en train un tel programme en se fondant sur des objectifs mal définis et que ses mesures phénotypiques manquent de précision, il se peut très bien que le taux de croissance diminue et que la réforme des allèles vraiment utiles empêche toute amélioration ultérieure.
C'est ainsi que la sélection portant sur le taux de croissance dans certaines populations de carpes indiennes a donné lieu à un ralentissement de la croissance du fait d'une définition approximative et de mesures imprécises du taux de croissance. Ces carpes étaient élevées au moyen d'un système de production appelé “à lots multiples”. Ce mode de gestion consiste à procéder, dans chaque étang, à des récoltes à la senne périodiques sur une période de plusieurs années, de façon à prélever régulièrement les poissons ayant atteint une taille marchande. On déverse souvent de jeunes poissons dans l'étang en remplacement des poissons prélevés, de façon à permettre une récolte continue. Après plusieurs années de production continue, l'étang est drainé, car les gros poissons qui ont échappé au sennage répété ont un effet néfaste sur la production.
Lorsque ces étangs à carpes ont été drainés, les exploitants et le personnel des écloseries ont épargné les plus gros poissons, car ils ont décidé d'évaluer le taux de croissance par le biais de la taille corporelle au moment du drainage, qu'ils n'ont donc mesurée qu'au moment où l'étang de production à lots multiples a été drainé. S'ils ont épargné ces poissons, c'est aussi parce qu'ils produisaient davantage d'oeufs que les géniteurs plus petits.
Malheureusement, du fait de leur définition du taux de croissance et de la façon dont ils ont mesuré le caractère considéré (la taille au moment du drainage des étangs), le programme d'élevage sélectif a abouti à une régression du taux de croissance, car les poissons sélectionnés étaient en fait les plus âgés, et donc ceux dont la croissance était la plus lente. S'ils étaient les plus gros au moment du drainage des étangs, c'était tout simplement parce qu'ils étaient particulièrement habiles dans l'art d'échapper à la senne. Le programme d'élevage sélectif a ainsi abouti à la sélection de poissons qui non seulement se développaient lentement, mais encore étaient de véritables experts dans l'art d'échapper à la capture.
Une fois que l'exploitant a quantifié ses objectifs et défini la façon dont les phénotypes seront mesurés, il lui faut déterminer le type de programme d'élevage sélectif qui lui permettra d'atteindre au mieux les objectifs en question. Il existe deux types fondamentaux de programmes d'élevage sélectif : la sélection individuelle (aussi appelée “sélection massale”) et la sélection familiale. Lorsqu'on procède à une sélection individuelle, on ignore les relations familiales et l'on compare les individus entre eux. Les individus sont classés par ordre, et les meilleurs sont gardés comme géniteurs sélectionnés. Par contre, en cas de sélection familiale, les relations familiales sont d'une importance capitale : en effet, soit on compare et on classe par ordre les moyennes relatives aux diverses familles et des familles entières sont épargnées ou réformées, soit on classe par ordre les individus au sein de chaque famille et la sélection s'opère indépendamment dans chacune des familles. Le choix du programme d'élevage sélectif le plus efficace est fonction du phénotype et de la façon dont il sera mesuré, de la biologie du poisson et de l'héritabilité des caractères. Ces deux types de sélection (individuelle et familiale) sont examinés en détail dans la suite du présent chapitre.
L'exploitant doit impérativement dresser le plan qu'il adoptera pour atteindre l'objectif qu'il s'est fixé. Il lui faut ainsi préciser le nombre de poissons qui fraieront, le nombre de poissons de chaque famille qui seront élevés, le nombre d'étangs qui seront empoissonnés, la densité d'empoissonnement et le programme de gestion qui sera utilisé aux fins d'élevage des poissons.
Les programmes d'élevage sélectif coûtent cher, et il donc indispensable de pouvoir évaluer avec précision les effets de la sélection. A cet effet, le meilleur moyen consiste à suivre l'évolution de la moyenne relative au phénotype considéré au cours du temps. Pour ce faire, il faut comparer la moyenne relative à la population sélectionnée à celle d'une population témoin n'ayant pas fait l'objet d'une sélection. Si cette comparaison est indispensable, c'est que la moyenne est fonction à la fois du génome de la population (qu'on essaie d'améliorer par sélection) et du milieu. Les techniques qui permettent d'obtenir une population témoin et la méthode d'évaluation des effets de la sélection sont examinées plus loin dans le présent chapitre.
Enfin, l'exploitant doit effectuer le bilan de son programme d'élevage sélectif au moins une fois par an. Il lui faut évaluer les progrès enregistrés et déterminer s'il existe des moyens d'améliorer le programme en question. A l'inverse, il peut décider de mettre fin au programme si les résultats indiquent qu'il ne fonctionne pas ou que les gains obtenus n'en valent pas la peine. Il arrive souvent que des exploitants souhaitent interrompre un programme d'élevage sélectif pour aussitôt en mettre en train un autre, souvent par simple dépit. Il importe alors de les décourager d'agir ainsi. Un exploitant qui change de programme tous les ans ou tous les deux ans ne fera que dépenser son argent en pure perte. De plus, si le programme ne donne pas les résultats escomptés, il serait malavisé de persister et de dépenser de l'argent sans raison.
Une fois que l'exploitant a décidé d'améliorer un phénotype quantitatif par le biais de la sélection, il lui faut choisir le programme d'élevage sélectif le mieux adapté à son objectif. Il existe deux grands types de programmes d'élevage sélectif : les programmes fondés sur la sélection individuelle et les programmes fondés sur la sélection familiale. La sélection individuelle est plus simple, moins coûteuse et nécessite moins de relevés de données.
Dans le cas de la sélection individuelle, on mesure tous les individus, et la décision de sélectionner ou de réformer un poisson est uniquement fondée sur sa valeur phénotypique propre. Chaque valeur relative à un poisson donné est comparée à une “valeur limite”; les poissons dont la valeur phénotypique est égale ou supérieure à cette valeur limite sont épargnés, alors que les autres sont réformés. La valeur limite est une valeur phénotypique définie à l'avance, qui correspond souvent aux “10 % supérieurs” ou aux “5 % supérieurs”, ou encore à toute autre valeur appropriée. Les poissons qui sont épargnés constituent la première génération de géniteurs sélectionnés. Le choix de la valeur limite et la façon dont ce choix est effectué sont examinés plus loin dans le présent chapitre.
Lorsque la première génération de géniteurs sélectionnés fraient, ils engendrent la première génération de poissons sélectionnés, qu'on appelle “génération F,”; leur descendance est, à son tour, appelée “génération F2”, et ainsi de suite. Lorsque l'exploitant fait frayer les géniteurs qu'il a sélectionnés, il doit faire en sorte qu'ils puissent s'accoupler au hasard ou encore les apparier de façon aléatoire, à moins qu'il souhaite faire porter la sélection sur le taux de croissance “post-marchand” ou sur des phénotypes liés à la maturité. Le processus est alors répété pour chacune des générations successives (figure 19).
Les variables d'environnement exercent une influence considérable sur l'expression des phénotypes quantitatifs. Même lorsqu'un phénotype est doté d'une forte héritabilité, la part environnementale de la variance phénotypique est souvent plus importante que la part héritable. Si l'influence que le milieu exerce sur la variance phénotypique était la même pour tous les poissons, son impact sur la sélection serait minime, pour peu que la part héritable de la variance soit suffisante pour permettre à la sélection d'opérer. Malheureusement, bon nombre de sources à l'origine de cette variance d'environnement ont un effet différent sur les individus ou les familles. Cela oblige l'exploitant qui recourt à la sélection à maîtriser les variables d'environnement s'il veut éviter que certains individus ou certaines familles soient indûment avantagés. Si cela advenait, les poissons pourraient être sélectionnés du fait de conditions du milieu particulièrement favorables plutôt qu'en fonction de leur supériorité génétique. En ce cas, la sélection serait un échec.
Par exemple, dans une population donnée, les plus vieux poissons sont habituellement les plus gros, parce qu'ils ont bénéficié de la plus longue période de croissance. Il a été démontré qu'une simple différence d'âge de 24 heures pouvait donner aux poissons les plus vieux un avantage de taille définitif. Cela signifie qu'on sélectionnera les poissons les plus vieux, et non pas les poissons génétiquement supérieurs. En conséquence, si l'on ne maîtrise pas les différences de taille liées à l'âge, tout programme d'élevage sélectif visant à améliorer le taux de croissance sera en fait fondé sur la variance d'environnement (l'âge) et non pas sur la variance génétique additive, et se soldera donc par un échec. Il importe par conséquent que, dans la mesure du possible, tous les poissons fraient le même jour. Si ce n'est pas possible, la population doit être scindée en différentes cohortes regroupant des poissons d'âge identique, la sélection s'effectuant de façon indépendante dans chacune des cohortes. Une fois la sélection achevée, les géniteurs sélectionnés dans chaque cohorte peuvent être mélangés.
Au nombre des autres facteurs qui doivent être maîtrisés figurent l'âge et la taille des femelles ainsi que la variabilité des méthodes de gestion appliquées dans les différents étangs à jeunes poissons ou d'affinage. Par exemple, si chaque famille est regroupée dans un seul étang, la sélection individuelle ne permettra pas de distinguer les poissons dont la très grande taille résulte de leur supériorité génétique de ceux dont la très grande taille est due au fait qu'ils ont bénéficié de la plus faible densité d'empoissonnement ou de la nourriture la plus abondante. Si ces variables ne sont pas maîtrisées par l'exploitant, il peut très bien advenir que tous les plus gros poissons proviennent de la même famille et que leur très grande taille relève uniquement de la variance d'environnement. En ce cas, la sélection s'avèrera inopérante.
Même des variables environnementales apparemment négligeables comme la dimension des particules d'aliments peuvent constituer d'importantes sources de variance phénotypique. Des études concernant la carpe commune ont montré qu'en cas de calibrage inadéquat des particules d'aliments, certains individus se développent notablement plus vite que le reste de la population; cet avantage de développement ne peut être exploité dans le cadre d'un programme d'amélioration, car il s'agit d'un avantage lié au milieu, et non pas d'un avantage héritable.
Lorsqu'un exploitant met en oeuvre un programme d'élevage sélectif, il doit maîtriser la totalité des facteurs du milieu susceptibles d'avantager certains individus ou certaines familles; faute de quoi, la sélection sera moins efficace que dans le cas de programmes similaires exécutés par d'autres exploitants ou, même, échouera totalement. Vu que les programmes d'élevage sélectif nécessitent du temps et beaucoup d'efforts, l'incapacité de maîtriser ces variables se révèlera fort coûteuse.
Pour obtenir une population de géniteurs sélectionnés, l'exploitant doit décider de la valeur limite qu'il compte utiliser. Cette valeur s'exprime d'ordinaire en pourcentage (par exemple les 10 % supérieurs). On peut aussi l'exprimer sous la forme d'une taille minimale (500,0 g, par exemple), mais cela peut s'avérer hasardeux si, lors d'une année médiocre, aucun poisson n'atteint la valeur limite.
La valeur limite est généralement déterminée juste avant que ne débute la sélection. Il s'agit d'ordinaire d'épargner une fraction définie à l'avance de la population. Le moyen le plus simple de déterminer cette valeur en traumatisant le moins possible le poisson consiste à mesurer un échantillon aléatoire de 100 à 200 poissons. Une fois ces mesures faites, on les range par ordre. La valeur limite est en ce cas la valeur qui correspond au percentile souhaité. Par exemple, s'il s'agit d'épargner les 10 % supérieurs, la valeur limite correspond au 90e percentile. Si l'on a mesuré 200 poissons, il suffit simplement, une fois les mesures classées par ordre, de repérer la 20e mesure à partir de la valeur phénotypique la plus grande : c'est la valeur limite (figure 20).
Dimorphisme sexuel. Il est nécessaire de mettre la touche finale au processus de sélection. L'exploitant doit en effet déterminer si la sélection concernera la population dans son ensemble ou si elle portera sur les deux sexes séparément. Il s'agit d'une décision importante, car beaucoup d'espèces présentent un dimorphisme sexuel, l'un des sexes se développant davantage ou plus vite que l'autre. Si l'espèce ne présente pas de dimorphisme sexuel ou que la sélection ait lieu avant que ce dimorphisme se manifeste, il est alors possible de définir une seule valeur limite pour l'ensemble de la population. Par contre, si l'espèce présente un dimorphisme sexuel, il faut définir une valeur limite pour chacun des sexes si l'on ne veut pas que la population sélectionnée soit uniquement composée d'individus du sexe de plus grande taille (figure 21).
Bon nombre d'espèces présentent un dimorphisme sexuel pour ce qui est de la taille du corps. Ainsi, toutes les principales espèces d'élevage de tilapias présentent un dimorphisme sexuel prononcé. Après quelques mois, les mâles se développent beaucoup plus vite que les femelles, au point d'être souvent deux fois plus gros qu'elles après un an. C'est la raison pour laquelle les éleveurs de tilapias préfèrent élever des populations unisexuées entièrement composées de mâles plutôt que de femelles lors de leurs tentatives de maîtrise de la reproduction. La carpe chinoise et la carpe commune présentent aussi un dimorphisme sexuel quant à la taille, à savoir que les femelles se développent davantage que les mâles. Toutefois, chez la carpe, cette différence de taille ne se manifeste qu'après la maturité et, par conséquent, une seule valeur limite suffit lorsque la sélection porte sur le taux de croissance avant maturité.
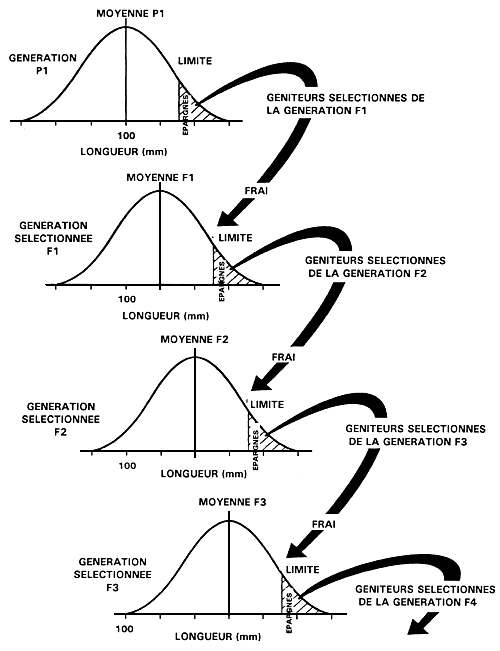
Figure 19. Représentation schématique d'un programme de sélection individuelle destiné à améliorer le taux de croissance par le biais d'une sélection portant sur la longueur. La figure présente un programme de sélection s'étalant sur quatre générations et montre comment la longueur peut être améliorée avec le temps.
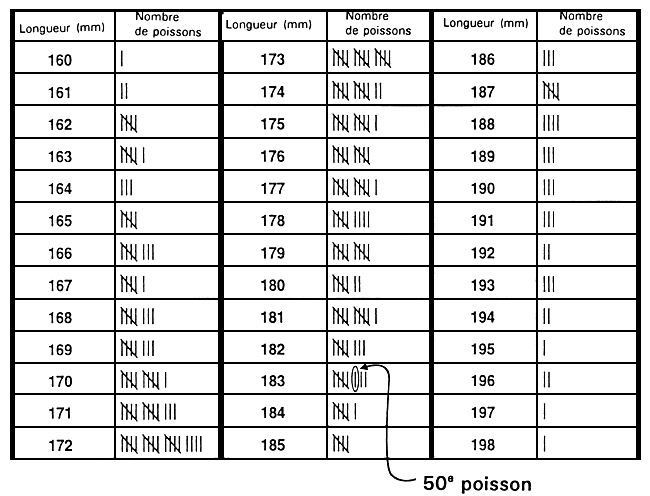
Figure 20. Mode de détermination rapide et simple d'une valeur limite. Le programme d'élevage sélectif en question a pour but d'augmenter la longueur à 10 mois par le biais de la sélection des 20 % supérieurs de la population. Pour déterminer la longueur qui correspond au 80e percentile, un échantillon de 250 poissons âgés de 10 mois a été prélevé dans un étang qui avait été empoissonné au préalable avec 5 000 poissons engendrés le même jour. Chacun des poissons de l'échantillon a été mesuré au millimètre près, et les valeurs phénotypiques ont été consignées sur la fiche illustrée ci-dessus. La valeur limite a été déterminée en dénombrant les poissons à partir du plus long d'entre eux (198 mm) afin de repérer le poisson correspondant 80e percentile. L'objectif consistant à sélectionner les 20 % supérieurs, la longueur du 50e poisson en partant du plus long indiquera la valeur limite (20 % de 250 = 50). Le 50e poisson est long de 183 mm, ce qui constitue en l'occurrence la valeur limite. Les poissons d'une longueur égale ou supérieure à 183 mm seront donc épargnés et deviendront les géniteurs sélectionnés, alors que les autres seront réformés.
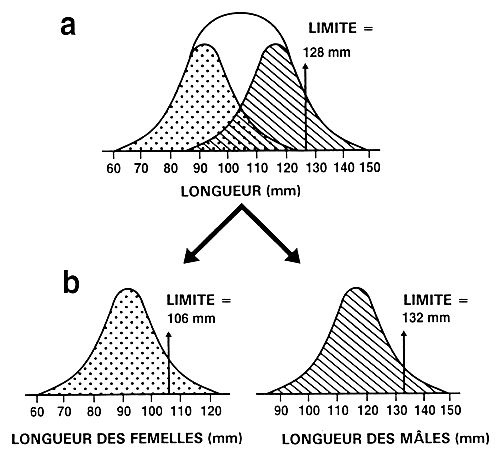
Figure 21. Effet du dimorphisme sexuel sur la sélection portant sur la taille. Le graphique “a” indique la distribution de la longueur des poissons d'une population où les mâles se développent davantage que les femelles; la distribution de la longueur des mâles correspond à la partie hachurée et celle de la longueur des femelles, à la partie en pointillés. En “a”, une seule valeur limite (128 mm) a été utilisée dans le cadre d'un programme d'élevage sélectif visant à augmenter la longueur. Par suite du dimorphisme sexuel, tous les géniteurs retenus étaient des mâles. En conséquence, l'exploitant soit se trouve dans l'impossibilité de produire une descendance, soit a recours à des femelles non sélectionnées. Le graphique “b” indique comment procéder à la sélection dans le cas des espèces présentant un dimorphisme sexuel. Il faut déterminer une valeur limite distincte pour chaque sexe et procéder indépendamment à la sélection des mâles et des femelles. On voit que l'exploitant a sélectionné les meilleurs mâles et les meilleures femelles en se fondant sur des valeurs limites de 132 mm et de 106 mm respectivement.
Si, en raison du dimorphisme sexuel, on procède indépendamment à la sélection des individus des deux sexes, la méthode permettant de déterminer la valeur limite (figure 20) doit être appliquée séparément pour les mâles et les femelles. Si l'on procède à une sélection touchant plusieurs cohortes, il faut déterminer la valeur limite propre à chacune de ces cohortes.
En général, les programmes les plus simples et, généralement, les plus efficaces sont ceux qui concernent un seul phénotype. Comme cela a été mentionné antérieurement, le degré possible d'amélioration de chacun des phénotypes est inversement proportionnel au nombre de phénotypes pris en considération par un programme d'élevage sélectif.
Si l'exploitant souhaite améliorer plus d'un phénotype, il peut avoir recours à la sélection en tandem, à la réforme indépendante ou à la méthode de l'indice de sélection. La sélection en tandem est inefficace et, donc, déconseillée. La méthode de l'indice de sélection est trop complexe pour la plupart des pisciculteurs, mais les aquiculteurs qui travaillent dans des stations de recherche et les responsables d'écloseries qui travaillent dans des centres de production d'alevins peuvent souhaiter l'employer. Les exploitants qui entendent procéder à une sélection portant sur deux ou, au plus, trois phénotypes doivent recourir à la réforme indépendante.
Sélection en tandem. Pour effectuer une sélection en tandem, il faut d'abord une sélection portant sur le premier phénotype, étalée sur plusieurs générations; on interrompt alors cette sélection initiale, et l'on met en train un nouveau programme de sélection portant sur le deuxième phénotype. Fondamentalement, il s'agit d'un programme d'élevage sélectif en deux étapes destiné à améliorer dans un premier temps un phénotype puis, après abandon du programme initial, changement soudain d'orientation et mise en train d'un deuxième programme de sélection, d'en améliorer un autre. Si l'amélioration du premier phénotype revêt une certaine importance et que le programme d'élevage sélectif donne de bons résultats, il n'est guère judicieux de l'interrompre. Pour cette raison, et aussi parce que cela prend beaucoup trop de temps d'essayer d'améliorer deux phénotypes au moyen de ce système “bout-à-bout”, la sélection en tandem doit être évitée par les exploitants désireux d'améliorer deux phénotypes ou plus. C'est une méthode par trop inefficace.
Réforme indépendante. La réforme indépendante est probablement le programme d'élevage sélectif le plus simple qu'un exploitant puisse utiliser pour améliorer deux ou trois phénotypes. Il s'agit d'un programme d'amélioration consistant à procéder à une sélection simultanée portant sur deux phénotypes ou plus. On établit une valeur limite pour chaque phénotype indépendamment de l'autre (ou des autres), et chaque poisson doit atteindre ou dépasser chacune de ces valeurs limites pour être épargné. Tout poisson qui ne satisfait pas à l'une des valeurs limites est réformé. Ce type de programme d'amélioration est illustré à la figure 22.
Supposons qu'un exploitant souhaite améliorer à la fois le taux de croissance, par une sélection portant sur la longueur, et la conformation du corps, par une sélection portant sur la largeur du corps au niveau de la première épine de la nageoire dorsale. La méthode permettant d'établir les valeurs limites est la même que celle mentionnée précédemment. L'exploitant peut soit établir des valeurs phénotypiques définies à l'avance, soit sélectionner les 10 ou 20 % supérieurs (ou tout autre pourcentage approprié). S'il a recours à des percentiles définis à l'avance, il détermine alors les valeurs limites qui leur correspondent en mesurant un échantillon aléatoire de 100 à 200 poissons et en identifiant les valeurs en question (voir figure 20). Dans notre exemple, l'exploitant a établi des valeurs limites de 243 mm pour la longueur et de 130 mm pour la largeur du corps au niveau de la première épine de la nageoire dorsale. Une fois ces deux valeurs limites établies, il mesure chaque poisson et épargne uniquement ceux qui atteignent ou dépassent chacune des deux valeurs en question. Dans le cadre de ce programme d'amélioration, les décisions concernant six des poissons mesurés ont été les suivantes :
| Poisson | Longueur (mm) | Largeur (mm) | Epargné/réformé |
| 1 | 254 | 129 | réformé |
| 2 | 243 | 130 | épargné |
| 3 | 241 | 138 | réformé |
| 4 | 240 | 120 | réformé |
| 5 | 280 | 129 | réformé |
| 6 | 250 | 131 | épargné |
Bien que la réforme indépendante soit plus efficace que la sélection en tandem et qu'elle soit facile à mettre en oeuvre, elle a pourtant deux inconvénients. Premièrement, lorsqu'on procède à une sélection simultanée et indépendante portant sur deux phénotypes, il arrive qu'on obtienne finalement très peu de poissons sélectionnés. Par exemple, lorsque la sélection porte sur deux phénotypes et que la valeur limite est fixée aux 10 % supérieurs pour les deux caractères, seulement 1 % de la population sera épargnée (0,1 × 0,1 = 0,01 ). Si l'on ajoute un troisième phénotype et que sa valeur limite soit identique, seul un poisson sur mille sera épargné. La rigueur de ce type de sélection est illustrée à la figure 22.
La forte restriction du nombre de poissons susceptibles de satisfaire à l'ensemble des valeurs limites soulève deux difficultés. Premièrement, il peut s'avérer impossible de produire assez de jeunes poissons sélectionnés pour l'affinage qui doit avoir lieu la saison suivante. Deuxièmement, et c'est peut-être là une difficulté encore plus grande, on peut aboutir, après une ou deux générations, à une consanguinité telle qu'elle risque d'annihiler les progrès accomplis grâce à la sélection.
On peut résoudre le problème suscité par des valeurs limites distinctes en décidant à l'avance de l'ampleur de la sélection, c'est-à-dire du pourcentage des poissons qui seront sélectionnés. Cela fait, on recalcule les pourcentages limites qui permettent d'atteindre cet objectif. Par exemple, si l'on désire conserver 15 % de la population et procéder à une sélection portant sur deux phénotypes d'importance identique, il suffit de calculer la racine carrée de 0,15, soit 0,39, et de sélectionner les 39 % supérieurs pour chacun des deux phénotypes (0,39 × 0,39 = 0,15). Si l'on souhaite mettre davantage l'accent sur l'un des phénotypes, il suffit de fixer une valeur limite plus élevée (en percentile) pour ce caractère et de réduire en conséquence la valeur limite relative au second caractère. Ainsi, si l'on désire épargner 10 % de la population, on peut fixer, pour les deux phénotypes, les valeurs limites au 25e et 40e percentile respectivement (0,4 × 0,25 = 0,1).
Deuxième inconvénient de la réforme indépendante, on est tenu de réformer des poissons qui s'avèrent exceptionnels s'agissant d'un des phénotypes. C'est le cas du poisson № 5 de l'exemple ci-dessus, qui est réformé alors que sa longueur est de très loin supérieure à la valeur limite relative à ce phénotype. On peut contourner la difficulté en ayant recours à la réforme indépendante modifiée. En ce cas, on épargne les poissons qui atteignent ou dépassent chacune des valeurs limites, mais aussi ceux qui dépassent l'une des valeurs limites et n'atteignent pas tout à fait l'autre (figure 23).
En raison de la facilité avec laquelle ils peuvent être mis en oeuvre et du peu de relevés de données qu'ils nécessitent, les programmes de réforme indépendante et de réforme indépendante modifiée sont sans conteste les programmes d'élevage sélectif que devraient mettre en oeuvre la plupart des exploitants désireux d'améliorer simultanément deux ou même trois phénotypes.
Indice de sélection. Il s'agit là du programme d'élevage sélectif le plus efficace que l'on puisse mettre en oeuvre pour améliorer deux phénotypes ou plus. Malheureusement, il est aussi plus complexe et plus coûteux que les programmes de réforme indépendante. L'usage de l'indice de sélection doit être réservé aux aquiculteurs travaillant dans des stations de recherche ou aux responsables d'écloseries travaillant dans des centres de production d'alevins. Les exploitants et la plupart des aquiculteurs devraient se limiter à la réforme indépendante s'ils souhaitent améliorer deux phénotypes. La méthode de l'indice de sélection ne s'impose réellement que lorsqu'il s'agit d'améliorer simultanément trois phénotypes ou plus.
La méthode de l'indice de sélection est un programme d'élevage sélectif consistant à évaluer simultanément deux phénotypes ou plus et à assigner une valeur numérique globale à chaque poisson. Les poissons sont ensuite classés par ordre en fonction de ces valeurs numériques. Le classement est établi de sorte qu'un poisson correspondant au 50e percentile pour chaque phénotype reçoive une valeur I de 100,0. En conséquence, les poissons dont la valeur I est supérieure à 100 sont considérés comme supérieurs à la moyenne et ceux dont la valeur I est inférieure à 100, comme inférieurs à la moyenne. L'élaboration d'un indice de sélection est le but ultime de tout programme d'élevage sélectif perfectionné à grande échelle.
Pour élaborer un indice de sélection, il faut disposer d'informations sur l'héritabilité, la corrélation génétique entre phénotypes et l'importance économique des divers phénotypes; malheureusement, il n'existe pas d'informations de ce genre concernant la plupart des populations de poissons comestibles d'élevage. On peut tourner la difficulté en modifiant l'indice de sélection et en établissant ce qu'on appelle un “facteur d'importance” pour chacun des phénotypes.
Les techniques qu'on emploie pour élaborer un indice de sélection fondé sur les facteurs d'importance sont exposées au tableau 11. Dans ce tableau, l'indice concerne trois phénotypes : la longueur à 12 mois, la largeur du corps au niveau de la première épine de la nageoire dorsale et le rendement de carcasse (exprimé de façon approchante par le rapport de la longueur du tronc - qui va de l'arrière de la tête à l'extrémité du pédoncule caudal - à la longueur totale). Après calcul de la valeur I propre à chaque poisson, les poissons sont classés par ordre en fonction de ces valeurs, et les 10, 15 ou 20 % supérieurs sont sélectionnés. Quant aux autres, ils sont réformés. La figure 24 présente un programme d'élevage sélectif fondé sur l'indice de sélection établi au tableau 11.
Ce type de programme d'élevage sélectif demande beaucoup plus de travail que la réforme indépendante. S'agissant de cette dernière, l'exploitant établit une valeur limite pour chaque phénotype (en se fondant d'ordinaire sur un échantillon de 100 à 200 poissons), ce qui lui permet d'épargner ou de réformer rapidement chaque poisson. Dans le cas de la méthode de l'indice de sélection, il faut mesurer tous les phénotypes pour chacun des poissons, puis identifier ceux-ci au moyen d'une étiquette ou d'une marque et les placer dans un bassin ou un hapa jusqu'à ce que l'ensemble des mesures soient effectuées. Une fois les mesures achevées, on identifient de nouveau les poissons, et on les épargne ou on les réforme selon la valeur I qui leur correspond. Ce procédé a non seulement l'inconvénient de nécessiter une main-d'oeuvre considérable, mais la manipulation répétée stresse en outre les poissons. Pour toutes ces raisons, tout exploitant désireux d'améliorer deux ou même trois phénotypes devrait s'en tenir à la réforme indépendante ou à la réforme indépendante modifiée.
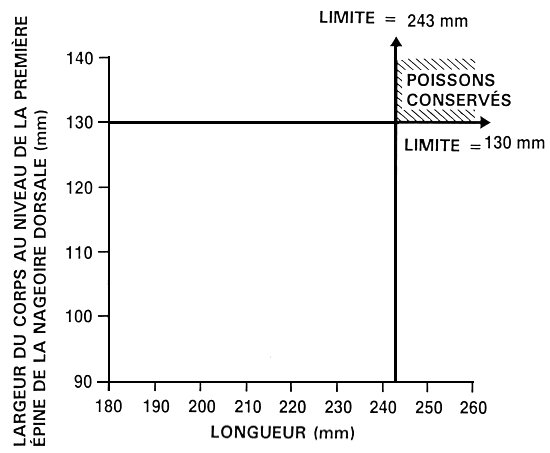
Figure 22. Représentation schématique d'un programme de réforme indépendante. Pour ce qui est du programme d'élevage sélectif présenté ici, un exploitant procède à une sélection qui vise à améliorer le taux de croissance (portant sur la longueur) et la conformation du corps (portant sur la largeur du corps au niveau de la première épine de la nageoire dorsale). Il a établi des valeurs limites distinctes de, respectivement, 243 et 130 mm. Les poissons qui atteignent ou dépassent ces deux valeurs limites sont épargnés et deviennent les géniteurs sélectionnés, et tous les autres sont réformés. La figure permet de se faire une idée précise des pourcentages relatifs de poissons épargnés (partie hachurée) et de poissons réformés (partie non hachurée) dans le cas d'un programme d'élevage sélectif de ce type.
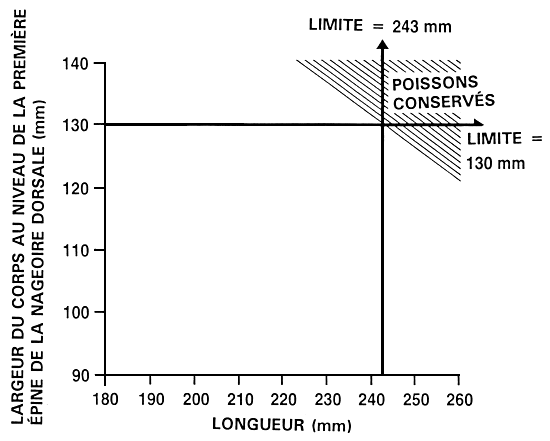
Figure 23. Représentation schématique d'un programme de réforme indépendante modifiée. Pour ce qui est de ce programme d'élevage sélectif, l'exploitant procède à une sélection qui vise à améliorer le taux de croissance (portant sur la longueur) et la conformation du corps (portant sur la largeur du corps au niveau de la première épine de la nageoire dorsale). Comme dans le cas du programme d'amélioration illustré à la figure 22, il a fixé des valeurs limites distinctes de, respectivement, 243 mm et 130 mm. En ce cas, l'exploitant épargne non seulement les poissons qui atteignent ou dépassent les deux valeurs limites, mais aussi les poissons qui dépassent une valeur limite et n'atteignent pas tout à fait l'autre (partie hachurée du graphique). Tous les autres poissons sont réformés. La réforme indépendante modifiée permet de sélectionner un plus grand nombre de poissons et de ne pas réformer les individus qui ont une valeur phénotypique tout à fait exceptionnelle pour un des caractères.
Tableau 11. Techniques permettant de calculer un indice de sélection à partir des facteurs d'importance
| Etape 1 | Déterminer les phénotypes qui seront pris en considération dans l'indice de sélection. Dans le présent exemple, on cherche à élaborer un indice de sélection en vue d'améliorer trois phénotypes: 1 ) la longueur à 12 mois (mm) 2) la largeur du corps au niveau de la première épine de la nageoire dorsale (mm) 3) le rendement de carcasse (exprimé de façon approchante par le rapport de la longueur du tronc -- qui va de l'arrière de la tête à l'extrémité du pédoncule caudal - à la longueur totale) | ||
| Etape 2 | Déterminer les facteurs d'importance relatifs aux phénotypes à l'aide de la formule suivante: | ||
| facteur d'importance (I) = importance relative du phénotype/moyenne relative au phénotype pour l'ensemble de la population | |||
| Etape 2a | Déterminer l'importance relative des divers phénotypes. A cet effet, on assigne une valeur arbitraire à chacun d'eux. Ces valeurs vont de 1 à 99 % (pour un total de 100 %) et représentent la part de la pression de sélection qui sera assignée à un phénotype (si l'on assigne une valeur de 0 % à un phénotype, cela signifie qu'on ne souhaite pas l'améliorer et qu'il ne sera pas intégré dans le programme; à l'inverse, si l'on assigne une valeur de 100 % à un phénotype, cela signifie qu'on entend améliorer uniquement ce phénotype et que tous les autres seront automatiquement exclus du programme d'amélioration). | ||
| Pour ce qui est de l'importance relative, les valeurs assignées sont les suivantes : | |||
| Phénotype | Relative improtance | ||
| longueur | 60% | ||
| largeur du corps | 30% | ||
| rendement de carcasse | 10% | ||
| Etape 2b. | Déterminer la moyenne propre à chaque phénotype. | ||
| L'exploitant mesure un échantillon aléatoire de 200 poissons et détermine les moyennes suivantes : | |||
| Phénotype | Moyenne | ||
| longueur | 195 mm | ||
| largeur du corps | 100 mm | ||
| rendement de carcasse | 55% | ||
| Etape 2c | Calculer le facteur d'importance de chaque phénotype à l'aide de la formule indiquée au début de l'étape 2. Les facteurs d'importance relatifs aux trois phénotypes retenus sont : | ||
| Ilargeur(IL) = 60 %/195 mm = 0,3076923 | |||
| Ilargeurdu corpe(IB) = 30 %/100 mm = 0,3 | |||
| Iredement de carcasse (ID) = 10 %/55 % = 0,1818182 | |||
| Etape 3 | Calculer l'indice de sélection qui permettra de déterminer la valeur génétique I de chaque poisson. On détermine la valeur de chaque poisson en fonction de ses valeurs phénotypiques et des facteurs d'importance des divers phénotypes, en se fondant sur la formule suivante : | ||
| Ipaisson = (IL)(longueur) + (IB)(largeur du corps) + (!D)(rendement de carcasse) | |||
| Etape 4 | Déterminer la valeur génétique d'un poisson présentant les valeurs phénotypiques suivantes : longueur = 201 mm; largeur = 119 mm; rendement de carcasse = 54 %. | ||
| I = (0,3076923X201 mm) + (0,3)(119 mm) + (0,1818182)(54 %) | |||
| I = 107,364 | |||
La sélection familiale diffère de la sélection individuelle en ce que la décision d'épargner ou de réformer concerne cette fois des familles et que les valeurs phénotypiques des individus n'importent que dans la mesure où elles sont liées à la moyenne relative à leur famille. Le plus souvent, on a recours à la sélection familiale quand la sélection individuelle s'avère inefficace du fait de la faible héritabilité du phénotype considéré (généralement moins de 0,15). Lorsque l'héritabilité et, par conséquent, la part héritable de la variance phénotypique sont faibles, l'essentiel des différences mesurables entre individus sont dues à des sources de variance non héritables. La sélection à l'échelon familial permet généralement de neutraliser une fraction appréciable de la variance d'environnement, ce qui facilite l'identification des différences génétiques et la sélection des poissons dont la supériorité résulte de la variance héritable.
On a également recours à la sélection familiale lorsque les causes environnementales de la variance ne peuvent être maîtrisées, ce qui a souvent pour effet de rendre toute amélioration par la sélection individuelle difficile ou impossible. Par exemple, si l'on n'est pas en mesure de faire frayer les poissons de façon synchrone et que la reproduction s'étale sur plusieurs semaines ou plusieurs mois, on utilise souvent la sélection familiale. Ce mode de sélection permet de maîtriser les différences de taille dues à l'âge.
La sélection familiale est le seul type de programme d'amélioration qui puisse être utilisé lorsqu'il est nécessaire de tuer un animal pour déterminer son phénotype. Ainsi, si l'on souhaite améliorer le rendement de carcasse ou réduire la teneur en matières grasses des viscères, on est obligé de tuer les poissons pour obtenir des données précises. Une fois les poissons morts, il est impossible de les faire frayer (à moins qu'on soit en mesure de conserver les gamètes par cryogénie). Il faut donc recourir à la sélection familiale pour améliorer de tels phénotypes.
La sélection familiale se divise en deux grandes catégories : la sélection interfamiliale et la sélection intrafamiliale. Pour procéder à une sélection interfamiliale, on commence par établir les valeurs moyennes propres à chaque famille, puis on classe ces valeurs par ordre. Ensuite, on épargne ou on réforme des familles entières (figure 25). Lorsqu'on a recours à ce type de sélection, on ne fixe généralement pas à l'avance une valeur limite minimale. L'exploitant décide plutôt d'épargner les 10, 15 ou 20 meilleures familles.
Il n'est pas nécessaire d'épargner toute la famille. Si l'effectif familial est important, on peut se contenter d'épargner des échantillons aléatoires et égaux dans chacune des familles sélectionnées. Il importe que l'échantillon soit prélevé au hasard dans chaque famille si l'on désire procéder à une véritable sélection interfamiliale. L'exploitant ne doit pas délibérément choisir les meilleurs poissons dans chacune des familles sélectionnées. Même si des familles entières sont épargnées, il convient d'égaliser les effectifs des familles sélectionnées de sorte qu'aucune d'entre elles n'ait une part plus importante que les autres dans la population soumise à la sélection.
| Etape 1. | Déterminer les valeurs phénotypiques propres à chacun des poissons de la population (composée en l'occurrence de 10 poissons). | |||
| Poisson | Longueur (mm) | Largeur du corps (mm) | Rendement de carcasse (%) | |
| T-1 | 198 | 103 | 54 | |
| T-2 | 210 | 110 | 55 | |
| T-3 | 180 | 108 | 57 | |
| T-4 | 195 | 100 | 55 | |
| T-5 | 197 | 101 | 56 | |
| T-6 | 190 | 98 | 54 | |
| T-7 | 188 | 97 | 53 | |
| T-8 | 195 | 102 | 57 | |
| T-9 | 185 | 95 | 52 | |
| T-10 | 199 | 102 | 55 | |
| Etape 2. | Déterminer la valeur I propre à chaque poisson à partir des facteurs d'importance établis au tableau 11 . | ||||||||
| IT-1 | = | (0.3076923)(198) | + | (0.3)(103) | + | (0.1818182)(54) | = | 101.641 | |
| IT-2 | = | (0.3076923)(210) | + | (0.3)(110) | + | (0.1818182)(55) | = | 105.777 | |
| IT-3 | = | (0.3076923)(180) | + | (0.3)(108) | + | (0.1818182)(57) | = | 98.148 | |
| IT-4 | = | (0.3076923)(195) | + | (0.3)(100) | + | (0.1818182)(55) | = | 100.000 | |
| IT-5 | = | (0.3076923)(197) | + | (0.3)(101) | + | (0.1818182)(56) | = | 101.097 | |
| IT-6 | = | (0.3076923)(190) | + | (0.3)(98) | + | (0.1818182)(54) | = | 97.680 | |
| IT-7 | = | (0.3076923)(188) | + | (0.3)(97) | + | (0.1818182)(53) | = | 96.583 | |
| IT-8 | = | (0.3076923)(195) | + | (0.3)(102) | + | (0.1818182)(57) | = | 100.964 | |
| IT-9 | = | (0.3076923)(185) | + | (0.3)(95) | + | (0.1818182)(52) | = | 94.878 | |
| IT-10 | = | (0.3076923)(199) | + | (0.3)(102) | + | (0.1818182)(55) | = | 101.831 | |
| Etape 3 | Classer les poissons en fonction des valeurs I qui leur correspondent. Sélectionner ou réformer les poissons selon leur place dans le classement. Dans le pré exemple, les 20% supérieurs de la population sont épargnés. |
| T-2 | = | 105.777 | → | EPARGNÉS | ||
| T-10 | = | 101.831 | ||||
| RÉFORMÉS | ← | T-l | = | 101.641 | ||
| T-5 | = | 101.097 | ||||
| T-8 | = | 100.964 | ||||
| T-4 | = | 100.000 | ||||
| T-3 | = | 98.148 | ||||
| T-6 | = | 97.680 | ||||
| T-7 | = | 96.583 | ||||
| T-9 | = | 94.878 | ||||
Figure 24. Méthode schématique permettant de mener à bien un programme fondé sur la méthode de l'indice de sélection. Il convient au préalable de déterminer les facteurs d'importance en fonction des moyennes phénotypiques et de l'importance relative des divers phénotypes. Les techniques permettant de calculer les facteurs d'importance relatifs aux trois phénotypes retenus sont décrites au tableau 11. Dans l'optique d'une détermination de la valeur I propre à chaque poisson, la première étape consiste à établir les valeurs phénotypiques relatives à chacun des poissons de la population. Dans le présent exemple, on s'est limité à 10 poissons. L'étape 2 consiste à calculer la valeur I propre à chaque poisson. Enfin, l'étape 3 consiste à classer les poissons en fonction des valeurs I qui leur correspondent, puis à les sélectionner ou à les réformer selon leur position dans ce classement. Dans l'exemple ci-dessus, où les 20 % supérieurs de la population devaient être sélectionnés, les poissons T-2 et T-10 ont été épargnés du fait qu'ils étaient les deux mieux classés, alors que les autres ont été réformés. On remarque que le poisson T-4, dont les valeurs phénotypiques sont identiques aux moyennes relatives à la population, a une valeur I égale à 100,0 (cette valeur I doit être égale à 100,0 par définition).
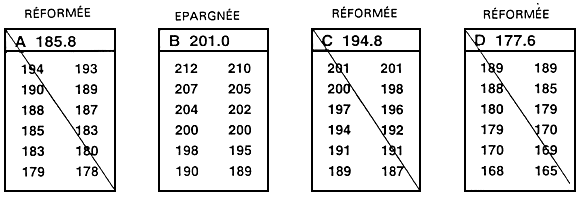
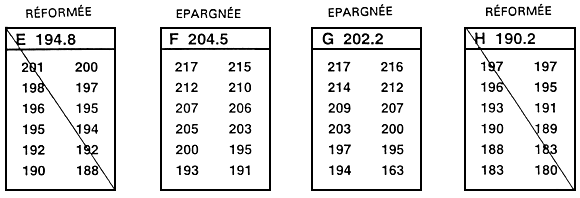
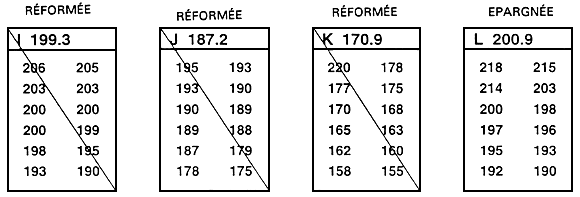
Figure 25. Représentation schématique du processus de sélection interfamiliale. On compare les moyennes relatives à des familles, et la sélection s'opère à l'échelon familial. Ce sont des familles entières (ou des échantillons aléatoires et d'effectifs égaux) qui sont épargnées ou réformées. Dans le présent exemple, il s'agit d'améliorer le taux de croissance par le biais d'une sélection portant sur la longueur. A cet effet, on sélectionne, parmi toutes les familles (12 en l'occurrence), les 33 % supérieurs. On procède en classant par ordre les moyennes relatives aux familles. Les quatre familles aux longueurs moyennes les plus fortes sont épargnées (familles B, F, G et L). Tous les poissons des huit autres familles sont réformés. Dans l'exemple, 12 poissons de chaque famille ont été mesurés au millimètre près (la liste de ces valeurs apparaît sous la lettre qui identifie la famille). Il faudrait en réalité mesurer 30 à 100 poissons. La valeur qui suit la lettre identifiant la famille correspond à la moyenne relative à la famille en question. Dans le présent exemple, on se fonde sur l'hypothèse d'une absence totale de dimorphisme sexuel. En présence d'un tel dimorphisme, il convient d'établir les moyennes relatives aux deux sexes dans chacune des familles. On peut alors procéder à la sélection, soit en se fondant sur les valeurs moyennes propres aux deux sexes, soit en traitant indépendamment les mâles et les femelles.
Comme dans le cas de la sélection individuelle, le dimorphisme sexuel peut compliquer la sélection destinée à améliorer le taux de croissance. Si l'espèce considérée présente un dimorphisme sexuel, les mâles et les femelles doivent être pesés ou mesurés séparément au sein de chaque famille. La sélection peut alors s'opérer de deux façons différentes. La première solution consiste, une fois connues les valeurs moyennes relatives à chaque sexe, à faire la moyenne de ces deux valeurs moyennes (les mâles et les femelles participant les uns et les autres pour moitié à la contribution de chaque famille à la génération suivante); on se fonde alors sur cette moyenne globale pour épargner ou réformer les familles. Deuxième solution, la sélection peut s'effectuer séparément pour chaque sexe; en ce cas, il est probable que les mâles de certaines familles seront réformés alors que les femelles seront épargnées, et vice-versa.
Pour mener à bien une sélection familiale, l'exploitant doit pouvoir assigner chaque poisson à une famille donnée. A cet effet, il existe deux façons de procéder. La première méthode consiste à élever chaque famille dans des bassins, des hapas ou des étangs distincts jusqu'à ce que les poissons aient une taille qui permette d'identifier leurs familles respectives à l'aide de marques ou d'étiquettes. Une fois marqués, les poissons peuvent être élevés ensemble; au moment de la récolte, ils seront de nouveau triés par famille, de manière à permettre la sélection. La deuxième méthode consiste à élever les poissons dans des bassins, des hapas ou des étangs distincts depuis le moment où les oeufs sont fécondés jusqu'à la sélection. Lorsque la sélection a eu lieu, il est souvent nécessaire que les poissons restent séparés si l'on ne peut les marquer. Si, par contre, le marquage peut avoir lieu, les géniteurs sélectionnés peuvent être rassemblés jusqu'au moment du frai.
La sélection interfamiliale coûte plus cher que la sélection individuelle, car elle nécessite davantage d'installations et de main-d'oeuvre. Elle nécessite aussi plus de relevés de données. Le fait de pouvoir marquer et élever en commun les poissons plutôt que d'être obligé d'élever chaque famille séparément contribue à abaisser considérablement les coûts.
Il est intéressant de noter que la sélection interfamiliale permet de réduire quelque peu la main-d'oeuvre que nécessite l'amélioration du taux de croissance et qu'elle permet en outre à l'exploitant d'atteindre cet objectif en faisant porter la sélection sur le poids. En ce cas, il n'est pas nécessaire de peser individuellement chaque poisson, puisque la sélection interfamiliale est fondée sur les moyennes familiales. Pour obtenir ces moyennes, il suffit de peser l'ensemble des poissons de chacune des familles et de diviser les valeurs obtenues par les effectifs correspondants.
Si chacune des familles est élevée dans un bassin ou un étang d'affinage distinct, il est nécessaire de répéter l'expérience dans au moins trois bassins ou étangs différents et de faire la moyenne globale des trois séries de résultats obtenus avant d'épargner ou de réformer les familles. Si chaque famille est élevée dans un seul étang ou bassin, il est en effet impossible de déterminer si la valeur phénotypique moyenne propre à une famille donnée reflète la valeur génétique de la famille en question ou encore la qualité de l'étang ou du bassin d'élevage. Tout pisciculteur sait fort bien que chaque étang ou bassin a ses particularités. Il y a de “bons” étangs et de “mauvais” étangs. Certains se caractérisent en permanence par une eau d'excellente qualité, une prolifération algale convenable et de bons rendements, alors que d'autres accumulent les problèmes d'oxygène dissous et de prolifération algale insuffisante et ne procurent que de faibles rendements. Si chaque famille est élevée dans un seul étang, les écarts entre les moyennes familiales peuvent résulter de la dissemblance des populations algales dans les différents étangs ou d'autres facteurs du milieu plutôt que de la variance héritable propre à chacune des familles. Si la plus grande part des écarts entre moyennes familiales est due à la disparité des conditions qui régnent dans les divers étangs, la sélection sera un échec.
Les exploitants doivent s'efforcer d'élever les différentes familles dans les mêmes conditions. Ainsi, la densité d'empoissonnement est un facteur du milieu que l'exploitant peut et doit maîtriser lorsqu'il procède à une sélection interfamiliale. Il importe que l'ensemble des familles soient élevées dans des conditions d'empoissonnement identiques; sinon, les écarts entre les moyennes familiales seront en partie dus aux différences de densité d'empoissonnement plutôt qu'à une disparité héritable.
Comme la sélection est fondée sur des moyennes familiales et que des familles entières sont épargnées ou réformées, il arrive que certains poissons dotés de phénotypes exceptionnels soient éliminés. A la figure 25, par exemple, la famille K, dont la moyenne familiale est la plus faible, a été réformée. Comme cela concerne l'ensemble de la famille, le plus gros poisson de la population a été éliminé avec ses frères et soeurs. Par contre, ce type de sélection a pour conséquence de maintenir des poissons de conformation médiocre dans la population de géniteurs sélectionnés. Par exemple, à la figure 25, la famille G, qui a la deuxième plus forte moyenne, a été épargnée. Comme cela concerne toute la famille, le plus petit poisson de la population a été sélectionné comme géniteur.
Même s'il peut sembler peu judicieux d'exclure certains poissons qui auraient été épargnés et d'en inclure d'autres qui auraient été réformés en cas de sélection individuelle, cette sorte de sélection peut néanmoins s'avérer très efficace. En fondant la sélection sur les moyennes familiales, on neutralise certains des facteurs environnementaux de la variance phénotypique. D'aucuns feront valoir que le gros poisson de la famille K a tout simplement acquis cette taille parce que des conditions d'environnement favorables lui ont permis d'avoir davantage de nourriture que sa juste part. Même si l'on peut être tenté d'infléchir les lois de la sélection interfamiliale en épargnant certains poissons réformés ou en réformant certains géniteurs sélectionnés, il faut s'abstenir de toute initiative en ce sens. Bien qu'aucune technique de sélection ne soit parfaite et que toutes engendrent certaines frustrations, le fait de fonder en partie un programme d'élevage sélectif sur des préjugés ou des fantaisies personnelles plutôt que sur la génétique ne manquera pas d'en réduire l'efficacité.
La sélection intrafamiliale est le second mode de sélection familiale. En ce cas, chaque famille est considérée comme une sous-population temporaire, et l'on procède à la sélection indépendamment au sein de chaque famille (figure 26). Après que les poissons ont été mesurés, on classe par ordre les poissons de chacune des familles et l'on épargne les meilleurs. Dans ce type de sélection, l'exploitant épargne généralement les 10, 15 ou 20 meilleurs poissons de chacune des 15, 20 ou 30 familles concernées. En cas de dimorphisme sexuel, il faut procéder à une sélection indépendante pour chaque sexe; l'exploitant épargne alors les 10,15 ou 20 meilleurs mâles et les 10, 15 ou 20 meilleures femelles de chaque famille.
A la figure 26, on a épargné les quatre meilleurs poissons de chaque famille. En théorie, dans chaque famille, la distinction entre poissons épargnés et poissons réformés devrait être fondée sur la moyenne familiale. En pratique, il n'est pas nécessaire de calculer ces moyennes pour mener à bien une sélection intrafamiliale. Il suffit de mesurer et de classer par ordre tous les poissons d'une même famille (ou d'un échantillon aléatoire de 30 à 100 poissons appartenant à la même famille), puis d'en épargner un nombre défini à l'avance.
Comme la sélection s'effectue indépendamment dans chaque famille, les poissons épargnés dans une famille donnée peuvent être plus petits que bon nombre des poissons réformés dans une autre famille. Ainsi, à la figure 26, trois des poissons épargnés de la famille K sont plus petits que bon nombre des poissons réformés dans les autres familles. A l'inverse, on constate aussi que des poissons exceptionnels peuvent être réformés s'il n'atteignent pas la valeur limite propre à la famille (le nombre fixé à l'avance de poissons à épargner).
Comme dans le cas de la sélection interfamiliale, même si la réforme de gros poissons et l'incorporation de petits poissons dans la population de géniteurs sélectionnés peuvent susciter certaines frustrations, l'exploitant doit résister à la tentation d'épargner ou de réformer certains poissons sur la base de préjugés personnels. Lorsqu'on a recours à la sélection intrafamiliale, il est inévitable que les poissons sélectionnés dans certaines familles soient plus petits que bon nombre des poissons réformés dans les autres. C'est en fait prévu. En effet, on a généralement recours à la sélection intrafamiliale lorsqu'une importante cause non maîtrisée de variance d'environnement telle que la date de frai ou l'âge des femelles est perceptible à l'échelon familial, mais ne se manifeste pas au plan individuel. Le fait de subdiviser la population en un grand nombre de familles et à procéder à la sélection au sein de chaque famille permet de neutraliser ces facteurs du milieu (ce sont les mêmes pour tous les poissons d'une même famille) et d'empêcher qu'ils influent sur l'expression phénotypique et la sélection.
Les travaux de recherche concernant les tilapias ont montré que, dans la plupart des cas, la sélection individuelle destinée à améliorer le taux de croissance était inefficace. Parmi les diverses raisons avancées, l'une consistait dans l'incapacité de faire frayer les poissons simultanément. Vu que les tilapias se reproduisent sur une période de plusieurs mois, il est apparu que la sélection intrafamiliale constituait la méthode d'élevage sélectif la plus susceptible d'améliorer le taux de croissance, du fait qu'elle neutralisait l'influence de la date de frai sur la taille des poissons.
Comme dans le cas de la sélection interfamiliale, l'exploitant est tenu de marquer les poissons de chacune des familles ou encore de les élever dans des étangs ou des bassins d'affinage distincts. Toutefois, il n'est pas nécessaire, pour chacune des familles, de répéter l'expérience dans plusieurs étangs ou bassins d'affinage, car on compare uniquement les poissons d'une même famille (donc élevés dans le même bassin ou étang d'affinage). Comme on ne procède à aucune comparaison interfamiliale, les particularités environnementales des divers étangs n'influent pas sur le processus de sélection. En conséquence, chaque famille peut être élevée dans un seul étang ou bassin, ce qui explique que les coûts d'affinage associés à la sélection intrafamiliale soient bien moindres que ceux qui sont associés à la sélection interfamiliale.
Il est possible d'opérer une sélection intrafamiliale dans un seul étang. Si les poissons peuvent être marqués de façon à repérer leurs familles, on peut tous les regrouper. Au moment de la récolte, on sépare les poissons des différentes familles et on procède à la sélection de la façon décrite précédemment. Il est aussi possible, si un grand nombre de hapas peuvent être placés dans un étang, d'élever chaque famille dans un hapa distinct. Il suffit simplement que le taux de croissance soit identique dans les hapas et les étangs.
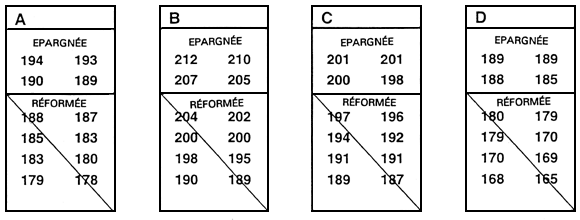
Figure 26. Représentation schématique du processus de sélection intrafamiliale. Dans le cas de ce type de sélection, on suppose que chaque famille constitue une sous-population temporaire, la sélection s'effectuant indépendamment dans chacune des familles. Dans le présent exemple, on essaie d'améliorer le taux de croissance en procédant à une sélection relative à la longueur. A cet effet, on sélectionne les quatre plus longs poissons de chaque famille. Les autres sont réformés. Dans le présent exemple, on a mesuré 12 poissons par famille. En réalité, il faut en mesurer 30 à 100 et sélectionner les meilleurs des spécimens constituant cet échantillon. Les longueurs des divers poissons (exprimées en mm) sont indiquées au-dessous de la lettre servant à identifier la famille. Dans le présent exemple, on suppose qu'il n'y a pas de dimorphisme sexuel. En cas de dimorphisme sexuel, la sélection s'opère indépendamment chez les mâles et les femelles. Dans l'exemple, les deux mâles et les deux femelles les plus longs de chaque famille seraient alors épargnés.
Une technique permettant d'atténuer les frustrations que suscite l'emploi de la sélection interfamiliale ou intrafamiliale consiste à les combiner de façon à obtenir un programme d'amélioration en deux étapes intégré (figure 27). En ce cas, on procède d'abord à une sélection interfamiliale de façon à sélectionner les meilleures familles, puis à une sélection intrafamiliale pour choisir les meilleurs poissons de chacune de ces familles. Il est indispensable d'élever les poissons selon la technique propre à la sélection interfamiliale, car la sélection porte d'abord sur les familles. La combinaison de ces deux formes de sélection permet d'éliminer tous les petits poissons et, donc, d'obtenir une population de géniteurs sélectionnés exemptes de “rebuts”. Si certains gros poissons seront réformés durant l'étape interfamiliale du programme d'amélioration, il ne faut pas oublier que cette supériorité de taille peut être non héritable, comme cela a été mentionné précédemment.
Une fois qu'on est parvenu à obtenir une population de géniteurs sélectionnés grâce à la sélection familiale, leur reproduction peut s'effectuer selon deux techniques d'accouplement possibles. La première, et aussi la plus simple, consiste à faire frayer ces géniteurs au hasard. La deuxième, qui est souvent employée, consiste à accoupler les mâles de la famille 1 avec les femelles de la famille 2, les mâles de la famille 2 avec les femelles de la famille 3, les mâles de la famille 3 avec les femelles de la famille 4, etc., et enfin les mâles de la dernière famille avec les femelles de la famille 1. Cette technique (appelée “accouplement en rotation”) vise à empêcher ou à réduire au minimum la consanguinité. A la deuxième génération, on décale les accouplements (on accouple les mâles issus du croisement de la famille 1 et de la famille 2 avec les femelles issues du croisement de la famille 3 et de la famille 4, et ainsi de suite) et on procède de même pour les générations suivantes.
Comme on peut s'en douter, cette technique nécessite un marquage des géniteurs sélectionnés, un nombre important d'étangs ou de bassins de frai ainsi que de nombreux relevés de données. Bien qu'il soit justifié de recourir à ce type d'accouplement dans le cadre d'un programme d'amélioration à grande échelle, cela n'est pas indispensable dans le cas d'une aquiculture de moyenne importance (en l'occurrence, l'accouplement aléatoire des géniteurs sélectionnés est suffisant).
Les programmes d'élevage sélectif sont coûteux, et il importe donc de pouvoir en évaluer les résultats avec précision. Le meilleur moyen consiste à comparer l'évolution de la moyenne phénotypique par rapport à celle d'une population témoin non soumise à la sélection. Il existe d'autres techniques permettant d'évaluer les résultats de la sélection, mais elles nécessitent la reproduction et l'élevage d'une autre population soumise à une sélection à rebours ou encore l'emploi de méthodes statistiques complexes.
S'il faut comparer la population sélectionnée avec une population témoin, c'est que la moyenne phénotypique relative à une population est fonction à la fois du génome (que l'on essaie d'améliorer par sélection) et du milieu. Supposons que, pendant la période de croissance, le temps soit exceptionnellement frais ou nuageux, que la fabrique d'aliments pour poissons ait mis, sans le savoir, de la chair de poisson de médiocre qualité dans ses produits ou que l'exploitant ait fait passer la densité d'empoissonnement de, disons, 4 000 à 8 000 poissons/ha; ces facteurs du milieu défavorables occasionneront un recul ou une stagnation du poids moyen de la population sélectionnée. Si l'on ne dispose que de cette donnée, on pourra seulement la comparer au poids moyen de la génération précédente et on en conclura que la sélection a échoué. Or, cela n'est pas nécessairement vrai. A l'inverse, il se peut que la sélection n'ait donné lieu à aucune amélioration génétique et que, cependant, le poids moyen ait augmenté à cause du temps exceptionnellement beau, de l'amélioration de la qualité des rations produites par la fabrique d'aliments pour poissons ou d'une gestion plus rigoureuse de l'exploitant. Là encore, si l'on ne dispose que du poids moyen de la population sélectionnée, on le comparera au poids moyen de la génération précédente et on en conclura, à tort, que la sélection a amélioré le taux de croissance.
En l'absence d'une population témoin, il est impossible de s'assurer du bien-fondé du diagnostic. Il est normal que le poids moyen augmente avec le temps, à mesure que l'exploitant améliore ses méthodes de gestion et par le biais de la domestication. Or, en l'absence d'une population témoin, il est impossible de distinguer la part de cette augmentation qui est due à l'élevage sélectif et la part qui est due, par exemple, à l'amélioration de la gestion.
Dans une grande écloserie, les géniteurs témoins de la génération F, peuvent consister en un échantillon aléatoire obtenu à partir de la population P1 (la population originale) avant toute opération de sélection (figure 28). Il est aussi possible de prélever un échantillon de poissons issus de spécimens proches de la moyenne de la génération P1. II faut laisser les géniteurs témoins de la génération F, à l'écloserie et les élever de la même manière que les géniteurs sélectionnés de la génération F1. Les géniteurs des deux populations doivent être traités et gérés de façon identique et doivent frayer dans les mêmes conditions. Leur descendance doit être elle aussi élevée de la même façon, ou de façon aussi peu différente que possible; en effet, toute modification des techniques d'élevage peut influer sur le phénotype et empêcher toute évaluation précise des résultats de la sélection. La seule exception consiste dans le fait qu'en ce qui concerne les générations suivantes (F2, etc.), les géniteurs de la population témoin sont choisis au hasard à chaque génération dans cette population, alors que les géniteurs sélectionnés sont choisis dans la population sélectionnée.
On peut aussi obtenir une population témoin en conservant les géniteurs P1, en les faisant frayer à différentes reprises sur plusieurs générations et en utilisant leurs descendances successives comme populations témoins. Cette façon de procéder peut cependant soulever des difficultés si l'âge et la taille des femelles exercent une influence appréciable sur le taux de croissance de la descendance. Bien que cela soit souvent le cas chez les poissons, les effets disparaissent généralement 20 à 180 jours après éclosion. Il ne faut pas employer cette méthode pour produire une population témoin si la sélection doit avoir lieu avant que ces effets aient disparu.
Quel que soit le mode de sélection (individuelle ou familiale), la façon d'obtenir et d'élever la population témoin ne varie pas. Il n'y a pas lieu de procéder différemment.
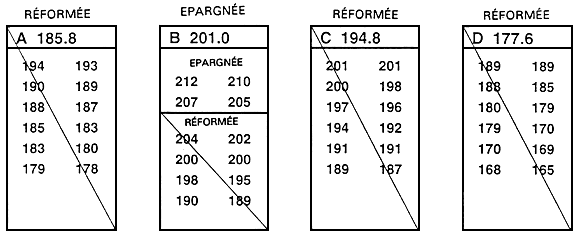
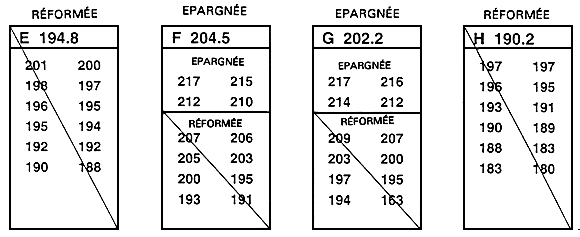
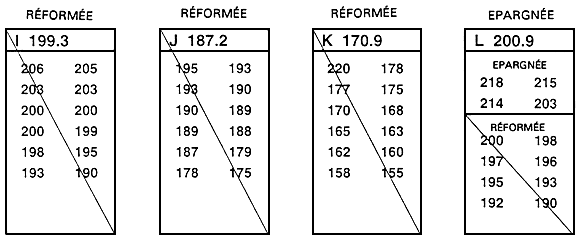
Figure 27. Représentation schématique d'un programme d'élevage sélectif combinant sélection interfamiliale et sélection intrafamiliale. Il s'agit d'un processus en deux étapes. Dans le présent exemple, on cherche à améliorer le taux de croissance en effectuant une sélection portant sur la longueur. La valeur qui suit la lettre servant à identifier chaque famille correspond à la moyenne (exprimée en mm); les longueurs des divers individus sont indiquées au-dessous. La première étape consiste à opérer une sélection interfamiliale et la deuxième, à réaliser une sélection intrafamiliale au sein des familles sélectionnées. Dans le présent exemple, lors de l'étape initiale, la sélection interfamiliale a conduit à épargner les quatre meilleures familles (B, F, G et L). Puis la sélection intrafamiliale a permis d'épargner les quatre poissons les plus longs de chacune de ces familles. Tous les autres poissons ont été réformés. Comme dans le cas des figures 25 et 26, on suppose que l'espèce considérée ici ne présente aucun dimorphisme sexuel. En cas de dimorphisme sexuel, on procède comme cela a été indiqué aux figures en question. Ce programme de sélection en deux étapes permet d'obtenir une population de géniteurs sélectionnés qui se trouvent être les meilleurs poissons des meilleures familles.
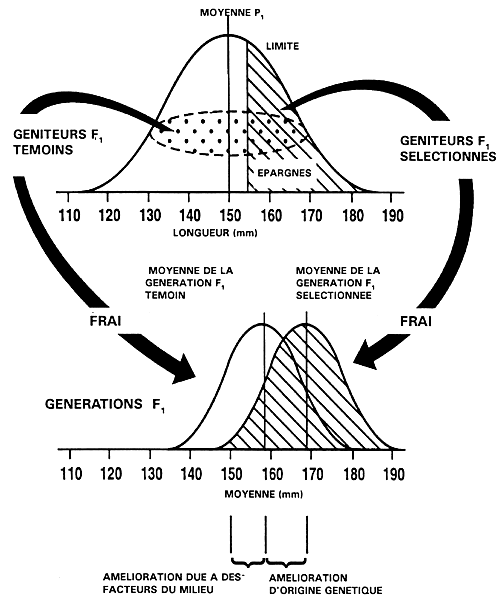
Figure 28. L'évaluation des résultats d'un processus de sélection nécessite une population témoin. Dans le présent exemple, la génération P1 a une longueur moyenne de 150 mm. Avant toute sélection, on a prélevé un échantillon aléatoire de la population afin d'obtenir les géniteurs témoins de la génération F1 (la partie pointillée de la courbe). Une fois ces géniteurs obtenus, on sélectionne les 30 % supérieurs de la population (limite = 155 mm). Tant les géniteurs sélectionnés que les géniteurs témoins de la génération F1 sont alors élevés de façon identique; lorsqu'ils sont parvenus à maturité, on les fait frayer afin d'obtenir les générations F,. Tant la génération F, sélectionnée que la génération F, témoin sont élevées selon des méthodes de gestion identiques. Après récolte, il a été établi que les poissons sélectionnés de la génération F, mesuraient en moyenne 169 mm, contre 159 mm pour les poissons témoins. En l'absence de population témoin, on aurait conclu que, sous l'effet de la sélection, la longueur moyenne était passée de 150 à 169 mm, soit une progression de 19 mm. Grâce à la génération F, témoin, il apparaît toutefois qu'une fraction importante de cette progression est due à des facteurs du milieu. La moyenne relative aux poissons témoins de la génération F, s'établissant à 159 mm, l'amélioration due aux facteurs du milieu est de 9 mm (159 mm - 150 mm) et l'amélioration d'origine génétique, de 10 mm (169 mm - 159 mm).
Si la création et la préservation d'une population témoin constituent le meilleur moyen d'évaluer les résultats de la sélection, cela prend du temps, de l'espace et de l'argent. La plupart des exploitants n'ayant pas la volonté ou la possibilité d'utiliser une population témoin, que peuvent-ils faire? Ils peuvent comparer le rendement moyen ou la taille moyenne des poissons qu'ils ont sélectionnés à ceux des poissons élevés dans une ferme aquicole voisine ou une écloserie gouvernementale. En ce cas, l'exploitant doit évaluer avec soin les plans d'aménagement appliqués dans les diverses fermes aquicoles et écloseries gouvernementales et s'efforcer d'en choisir un qui s'approche de celui qu'il emploie. Il doit en outre choisir un exploitant ou un responsable d'écloserie qui soit disposé à coopérer avec lui et qui poursuivra ses activités aquicoles pendant 10 ans au moins.
Tableau 12. Utilisation d'une population témoin aux fins d'évaluation des résultats d'un processus de sélection
| Soit : | longueur moyenne des poissons de la génération P1 = 150 mm | |
| longueur moyenne des poissons sélectionnés de la génération F1 = 165 mm | ||
| longueur moyenne des poissons témoins de la génération F1 = 159 mm | ||
| On calcule l'amélioration totale de la façon suivante : | ||
| Amélioration totale = longueur moyenne des poissons sélectionnés de la génération F, - longueur | ||
| moyenne des poissons de la génération P1 = 165 mm - 150 mm = 15 mm | ||
| On calcule ensuite l'amélioration due aux facteurs du milieu de la façon suivante : | ||
| Amélioration due aux facteurs du milieu = longueur moyenne des poissons témoins de la génération F, - longueur moyenne des poissons de la génération P1 = 159 mm - 150 mm = 9 mm | ||
| On calcule enfin l'amélioration d'origine génétique de la façon suivante : | ||
| amélioration d'origine génétique = longueur moyenne des poissons sélectionnés de la génération F, - longueur moyenne des poissons témoins de la génération F1 = 165 mm - 159 mm = 6 mm | ||
| ou | ||
| amélioration d'origine génétique = amélioration totale - amélioration due aux facteurs du milieu = 15 mm - 9 mm = 6 mm | ||
Le tableau 12 indique comment utiliser une population témoin pour évaluer les résultats d'un processus de sélection. Dans l'exemple du tableau, la moyenne relative à la génération P1 est de 150 mm et celle relative aux poissons sélectionnés de la génération F,, de 165 mm. En l'absence d'une population témoin, on constaterait que la moyenne a augmenté de 15 mm, mais on serait dans l'incapacité de déterminer quelle part de cette amélioration est due au programme d'élevage sélectif et quelle part est due, par exemple, aux progrès accomplis en matière de gestion. Puisqu'on dispose d'une population témoin, on peut préciser que, sur les 15 mm de progression, 9 sont dus à des facteurs du milieu. Cela signifie qu'en l'absence de programme d'élevage sélectif, la longueur moyenne serait passée de 150 à 159 mm. La différence de longueur entre les poissons sélectionnés et les poissons témoins de la génération F, (ou entre l'amélioration totale et l'amélioration due aux facteurs du milieu) est de 6 mm, ce qui correspond à l'amélioration d'origine génétique.
L'existence d'une population témoin permet aussi d'évaluer le pourcentage d'amélioration imputable à la sélection. Le programme d'élevage sélectif, en une génération, a fait progresser la longueur de 3,8 % (6 mm/159 mm). On notera que pour procéder à cette évaluation, on utilise la valeur relative au groupe témoin contemporain (en l'occurrence la génération F1 témoin) comme valeur étalon.
Il existe deux façons d'exprimer l'amélioration d'origine génétique en pourcentage. La première consiste à l'exprimer en pourcentage par génération, comme précédemment, et la deuxième, en pourcentage par année. Si l'intervalle entre générations correspond à une année, ces deux pourcentages coïncident. Si, par contre, cet intervalle est plus long ou plus court, le deuxième pourcentage sera lui aussi, respectivement, plus élevé ou plus faible. Par exemple, si les poissons de l'espèce considérée ne parviennent à maturité et ne fraient qu'à quatre ans, il faut alors diviser par quatre le pourcentage d'amélioration par génération pour déterminer le pourcentage d'amélioration par an.
Pour comparer les poissons sélectionnés à la population témoin, l'exploitant doit comparer les valeurs moyennes globales déterminées au moyen de poissons élevés dans au moins trois étangs distincts. Cela ne soulève aucune difficulté s'il dispose de sa propre population témoin ou qu'il utilise les poissons d'un voisin. Si chaque population est élevée dans un seul et même étang, l'exploitant ne sera pas en mesure de distinguer les effets associés aux étangs et les effets génétiques. Une population peut ainsi avoir une moyenne plus élevée en raison d'une meilleure prolifération algale dans son étang d'élevage.
Pour éviter de devoir élever les deux populations dans des étangs différents, il est possible de marquer le groupe témoin et de le placer avec la population sélectionnée dans un unique étang. En ce cas, l'effectif de la population témoin peut se limiter à 30 à 50 poissons.
Il n'est pas possible d'obtenir une évaluation vraiment précise des résultats d'un programme d'élevage sélectif par simple comparaison des moyennes de la population sélectionnée et de la population témoin. Il faut en fait procéder à une comparaison statistique des moyennes. Malheureusement, toute analyse statistique de ce genre nécessite une connaissance poussée de la planification des expériences et une compréhension élémentaire des statistiques. En conséquence, presque tous les exploitants ont besoin de l'assistance d'experts pour mener à bien une analyse de données. Toutefois, s'ils ne peuvent disposer d'une telle assistance, ils ne doivent pas renoncer à mettre en oeuvre un programme d'élevage sélectif simplement parce que les résultats ne feront pas l'objet d'une analyse statistique. Les moyennes brutes donneront une bonne indication de l'évolution en cours.
Les programmes d'élevage sélectif permettant d'améliorer les phénotypes quantitatifs sont plus ardus, plus complexes et plus longs à mettre en oeuvre que ceux qui servent à fixer des phénotypes qualitatifs. En fait, ces programmes d'amélioration ne sont pas limités dans le temps, car toute interruption de la sélection se traduit généralement par l'annulation d'une grande partie des progrès accomplis. De plus, ils nécessitent une tenue de registres plus contraignante ainsi qu'un certain nombre d'analyses mathématiques et statistiques des données recueillies. Au chapitre 5 sont mentionnés différents exemples de programmes d'élevage sélectif simples et peu coûteux qui peuvent être mis en oeuvre pour augmenter le taux de croissance ainsi que des exemples du genre de registres qu'il faut tenir.
Les programmes d'élevage sélectif sont des méthodes éprouvées d'amélioration du taux de croissance et du rendement. Ces programmes ne sont cependant pas gratuits, et les techniques qu'ils mettent en oeuvre en vue d'augmenter les rendements sont relativement complexes. Si le rendement peut être amélioré par des apports appropriés de chaux, d'engrais et/ou d'aliments pour animaux, par une densité d'empoissonnement mieux adaptée ou par une meilleure gestion de la qualité de l'eau, il est alors préférable d'adopter dans un premier temps cette démarche, qui donne lieu à des améliorations immédiates sans être très onéreuse.
Assez peu d'exploitants devraient être encouragés à mettre en oeuvre des programmes d'élevage sélectif. Ces programmes requièrent des étangs, de la main-d'oeuvre, des crédits et de nombreux relevés de données. Ils nécessitent en outre une planification à long terme et beaucoup de patience. Les exploitants qui n'ont pas la possibilité ou la volonté d'affecter une partie de leur établissement à la réalisation d'un programme d'amélioration, d'y consacrer la main-d'oeuvre nécessaire et de recueillir, de consigner et de mettre à jour les données indispensables ne devraient pas mettre en train des programmes d'élevage sélectif, au même titre que les exploitants incapables d'élaborer une planification à long terme (sur 1 à 10 ans).
Il n'est pas nécessaire que les exploitants deviennent des généticiens pour mettre en oeuvre des programmes d'élevage sélectif destinés à améliorer le taux de croissance de leurs poissons. Ils n'ont pas besoin de connaître l'héritabilité en matière de longueur ou de poids, ni même de savoir ce qu'est l'héritabilité. Mais ils doivent être de bons gestionnaires.
Avant de mettre en train un programme d'élevage sélectif, l'exploitant doit faire une étude pour s'assurer du bien-fondé de ce programme et identifier les phénotypes à améliorer. Pour la plupart des exploitants, la décision la plus logique consiste à améliorer le taux de croissance. En règle générale, les programmes d'élevage sélectif mis en oeuvre dans des établissements piscicoles de moyenne importance devraient se limiter à l'amélioration du taux de croissance. En effet, la possibilité d'amélioration d'un phénotype est inversement proportionnelle au nombre de caractères pris en considération dans le programme d'amélioration. De plus, l'augmentation du taux de croissance a pour effet d'améliorer d'autres importants phénotypes de production par le biais de la sélection indirecte.
Une fois que l'exploitant a pris une décision à propos du phénotype qu'il entend améliorer, il lui faut déterminer comment le phénotype sera mesuré, à quel moment procéder et quel genre de programme d'élevage sélectif (sélection individuelle, interfamiliale ou intrafamiliale) lui permettra d'obtenir une population de géniteurs sélectionnés adéquate. Le tableau 13 précise ce qui distingue ces divers programmes d'amélioration génétique. Dans la mesure du possible, il est préférable de recourir à la sélection individuelle, car c'est une méthode simple, peu coûteuse et qui nécessite très peu de relevés de données.
Si l'exploitant souhaite améliorer deux phénotypes, il lui faut employer la réforme indépendante ou la réforme indépendante modifiée. Il ne doit jamais utiliser la sélection en tandem. Si la méthode de l'indice de sélection constitue le programme d'élevage sélectif le plus efficace lorsqu'il s'agit d'améliorer deux phénotypes, il est néanmoins déconseillé de le mettre en oeuvre dans des établissements piscicoles de moyenne importance.
Tout exploitant qui met en oeuvre un programme d'élevage sélectif doit pouvoir évaluer les résultats de son travail. S'il dispose de ressources et d'un espace suffisants, il peut entretenir une population témoin. Sinon, il pourra périodiquement comparer la moyenne relative à la population qu'il a sélectionnée avec celle d'une population élevée dans une ferme piscicole voisine ou une écloserie gouvernementale.
Lorsque l'exploitant fait le bilan des améliorations résultant du programme d'élevage sélectif qu'il a mis en oeuvre, il lui faut procéder à une réévaluation de ce programme. Il doit examiner tous ses aspects et déterminer s'il n'existe pas des façons de faire plus simples et plus efficaces. Il lui faut aussi s'assurer que son investissement lui procure bien les dividendes escomptés. Bien qu'il soit logique qu'un exploitant mette fin à un programme d'élevage sélectif qui a échoué, il ne doit pas juger trop hâtivement les résultats de la sélection. Ce genre de programme, pour peu qu'il soit judicieusement mis en oeuvre, donne lieu à des améliorations réelles, quoique limitées.
Tableau 13. Comparaison des stratégies de base et de certains des avantages et inconvénients de la sélection individuelle, de la sélection interfamiliale et de la sélection intrafamiliale
| Type de sélection | Stratégie | Avantages | Inconvénients |
| individuelle | Sélection des meilleurs individus; les relations familiales n'ont aucune incidence. | A employer de préférence lorsque h2 ≥ 0,25; peu coûteuse; nécessite peu d'étangs; assez facile à utiliser aux fins d'amélioration de deux ou trois phénotypes; les poissons sélectionnés sont les poissons les plus grands; permet de préserver une importante population d'élevage; faible quantité de données requise; tenue de registres restreinte. | Inefficace lorsque h2 ≤ 0,15, car VE rend la sélection des meilleurs poissons difficile; le décalage des périodes de frai peut poser des problèmes. |
| intrafamiliale | Sélection des meilleursindividus au sein de chaque famille. | A employer de préférence lorsque h2 ≤ 0,15 et que VEinflue davantage sur les familles que sur les individus; utilisable en cas de reproduction non synchrone; permet assez facilement de préserver une importante population d'élevage; moins coûteuse que la sélection interfamiliale. | Assez coûteuse; nécessite de nombreux étangs; convient mal à l'amélioration de deux ou trois phénotypes; des petits poissons peuvent être sélectionnés comme géniteurs; nécessite beaucoup de données et une tenue de registres rigoureuse. |
| interfamiliale | Sélection des meilleuresfamilles fondée sur les moyennes familiales; les valeurs propres aux individus ne sont pas prises en considération. | A employer de préférence lorsque h2 ≤ 0,15 et que VEinflue davantage sur les individus que sur les familles; peut être utilisée quand les poissons doivent être tués. | Très coûteuse; nécessite de nombreux étangs; convient mal à l'amélioration de deux ou trois phénotypes; des petits poissons peuvent être sélectionnés comme géniteurs; peut aboutir à un fort degré de consanguinité; nécessite beaucoup de données et une tenue de registres rigoureuse. |
![]()
![]()
![]()