![]()
![]()
![]()
Normalmente, los caracteres cualitativos son menos importantes que los cuantitativos para los cultivadores de peces destinados a la alimentación, pero hay dos razones por las cuales para aplicar programas de cría selectiva es interesante conocer cómo pueden ser controlados y explotados. La primera, y más importante, es que los pueden influir en el valor de la cosecha o en el costo de producción. Cuando el piscicultor consigue un producto más atractivo, los consumidores están dispuestos a pagar un precio más elevado y él obtiene mayores beneficios. A la inversa, algunos caracteres cualitativos, como las deformidades, pueden reducir el valor de la población, ya sea porque aumentan el costo de producción o porque los peces son tan poco atractivos que los consumidores se abstienen de comprarlos. En ambos casos, los cultivadores pueden mejorar la población por medio de la selección. En segundo lugar, conocer cómo pueden utilizarse los programas de cría selectiva para fijar (frecuencia = 100%) los caracteres cualitativos deseados y eliminar (frecuencia = 0%) los no deseados y obtener poblaciones puras, puede hacer más fácil comprender cómo mejorar los caracteres cuantitativos mediante la selección.
Quienes conozcan bien la genética mendeliana y sepan utilizar los programas de cría para fijar o eliminar caracteres cualitativos, y quienes sólo estén interesados en los caracteres cuantitativos pueden ignorar este capítulo y pasar directamente al Capítulo 4.
Antes de poner en práctica un programa de cría selectiva, el piscicultor debe darse cuenta de que sólo puede utilizar la selección para conseguir objetivos que son factibles desde el punto de vista biológico. La selección es un programa de cría que se basa en la variación fenotípica heredable y para que pueda aplicarse con éxito deben cumplirse dos condiciones. En primer lugar, el carácter que el cultivador desea modificar debe presentar variación; en segundo lugar, la variación debe ser heredable. Los mejoradores sólo podrán conseguir lo que los genes de los peces les permita conseguir y no podrán obtener una población de peces con unas características prefijadas eliminando los caracteres no deseados, como las espinas pectorales puntiagudas, a menos que se descubran ejemplares sin ese tipo de espinas y que, en tal caso, el carácter sea heredable, pues de otro modo la selección supondrá un esfuerzo costoso, baldío y frustrante. Si los caracteres son heredables, antes de comenzar el programa de mejora el criador debe poseer la información genética pertinente, que le permitirá elegir el programa más adecuado, y conocer los costos relativos de los caracteres (costos de producción) y su valor (valor de mercado).
Los caracteres cualitativos son mutuamente excluyentes, pues un pez tiene un carácter o tiene otro diferente. Dicho de otra forma, los peces de una población se agrupan en categorías fenotípicas discretas y no coincidentes. Teniendo en cuenta que la selección explota la variación fenotípica, es necesario que existan al menos dos caracteres en la población, uno de los cuales ha de ser el carácter deseado por el cultivador. Si existe un solo carácter, no habrá variación fenotípica y la selección no podrá modificarlo. Por ejemplo, si todos los peces de una población son de color negro, no existe variación fenotípica para el color corporal y, en consecuencia, no cabe emplear la selección para modificarlo.
Además de existir variación, la variación observada ha de ser heredable, condición indispensable para que la selección pueda alterar las secuencias fenotípicas. La distinción entre variación heredable y no heredable es importante porque muchos de los caracteres cualitativos que se observan en las piscifactorías y en los criaderos son deformidades no heredables. Algunas tienen una base genética, pero la mayor parte se deben a deficiencias nutricionales, perturbaciones ambientales, toxinas, enfermedades y accidentes, o son anomalías del desarrollo. Las deformidades causadas por factores no genéticos no pueden ser eliminadas mediante la selección. Sólo se puede eliminar una deformidad no heredable descubriendo el factor ambiental que la ocasiona y suprimiéndolo, o bien, modificando las condiciones de cultivo. Dada la dificultad, cuando no la imposibilidad, de establecer la causa de una deformidad no heredable, muchas veces debe ser ignorada, salvo cuando su frecuencia se aproxima al 0,5%
Antes de emplear la selección para fijar o eliminar caracteres cualitativos y conseguir poblaciones puras es necesario determinar la forma en que se heredan los caracteres. Si ya se ha determinado en un experimento anterior puede utilizarse esa información para proyectar el programa de mejora. Si conoce la forma de herencia, el cultivador podrá elegir el programa de cría apropiado y conseguir sus objetivos con rapidez y eficacia. Si no la conoce corre el riesgo de elegir un programa inadecuado, lo cual no le permitirá conseguir el objetivo deseado y supondrá un despilfarro de esfuerzos, instalaciones y dinero.
Los programas de investigación que permiten determinar la forma de herencia de los caracteres cualitativos no son muy complicados. Por lo general, consisten en efectuar apareamientos durante dos generaciones y determinar los coeficientes fenotípicos que producen (las figuras 3 y 5 ilustran algunos de los apareamientos y coeficientes que se utilizan). Este tipo de investigación no reviste gran complejidad, pero requiere esfuerzo, instalaciones y la posibilidad de analizar los resultados estadísticamente. Por ello, deben realizarla los científicos en las universidades o centros de investigación estatales.
Antes de iniciar un programa de cría selectiva, el cultivador debe evaluar si es necesario. La evaluación puede realizarla el propio piscicultor (este tipo de estudios son costosos y exigen técnicas muy complejas), cooperativas pesqueras o un centro de investigación local de piscicultura. El cultivador necesita saber si sus clientes quieren peces con una coloración corporal diferente o si están dispuestos a pagar un precio más alto por unos peces más atractivos. La evaluación le permitirá conocer también si el programa previsto puede abrir nuevos mercados. Sólo se debe iniciar un programa de mejora si es necesario o si permite aumentar los beneficios. La modificación del color corporal o de otros caracteres puede originar peces más atractivos, pero si la evaluación pone de manifiesto que los consumidores no desean que se modifique el color, no se debe iniciar el programa.
Por último, antes de iniciar un programa de cría selectiva, el piscicultor debe conocer el costo que comporta producir los caracteres alternativos. El valor de algunos de éstos es tan evidente que es innecesario realizar un estudio científico formal. Si un piscicultor desea utilizar la selección para eliminar una malformación heredable que disminuye la tasa de crecimiento o la viabilidad, no necesita realizar un estudio para determinar los valores relativos de los caracteres, porque ya sabe que la malformación comorta un costo económico.
En cambio, determinar los costos relativos de producción de algunos caracteres entraña una dificultad mayor y requiere hacerlo en experimentos científicos. El método a seguir consiste en evaluar los efectos de los caracteres en la tasa de crecimiento, supervivencia, producción de huevos, etc. Los efectos secundarios reciben el nombre de “efectos pleiotrópicos” y en agricultura y piscicultura pueden ser más importantes que el propio carácter, especialmente si afectan al crecimiento, la supervivencia o la fecundidad. Prácticamente, todos los colores corporales mutantes que se han investigado en los peces presentan efectos pleiotrópicos negativos, es decir, afectan negativamente a la tasa de crecimiento o a la supervivencia. En la figura 8 se ilustra el efecto pleiotrópico negativo del color corporal rojo sobre la viabilidad de la tilapia del Nilo.
Es conveniente utilizar la selección cuando no existen efectos pleiotrópicos negativos o cuando los costos adicionales de producción (menor tasa de crecimiento y/o menor viabilidad) se compensan con el incremento del valor de mercado del producto (el precio más elevado que están dispuestos a satisfacer los consumidores). En cambio, sería absurdo aplicarla si el incremento de los costos de producción superara el valor mayor de mercado del producto conseguido, porque el cultivador obtendría menos beneficios.
En el caso de los caracteres cualitativos, el objetivo de todos los programas de cría selectiva es conseguir una población pura, es decir, una población que contiene únicamente un alelo para el locus en cuestión porque se ha efectuado su fijación, eliminándose el otro alelo mediante selección. De esta forma, en cada gameto producido por los peces reproductores seleccionados sólo existirá el alelo deseado y, por tanto, todas las crías engendradas por estos reproductores tendrán el carácter deseado. Sólo la introducción accidental (o intencional) de peces de otra población o la mutación pueden producir el carácter no deseado una vez que se ha obtenido una población pura mediante selección.
El carácter no deseado y el alelo que lo produce se eliminan mediante el procedimiento de “descarte”, que impide la reproducción de los peces descartados. No es necesario dar muerte a estos peces; pueden ser cultivados y vendidos como alimento pero no se debe permitir que se reproduzcan.
La selección de los caracteres cualitativos se rige por tres normas básicas:
Si el carácter deseado está controlado por un único genotipo homocigótico, se conseguirá una población pura con una sola operación de selección.
Si el carácter deseado está controlado por dos o más genotipos, la selección no puede conseguir una población pura. Para fijar el alelo deseado y conseguir una población pura es necesario utilizar la prueba de la descendencia.
Si el carácter deseado está controlado por el genotipo heterocigótico, ningún programa de cría puede originar una población pura. Para conseguir una población en la que todos los peces tengan el carácter deseado se han de cruzar los dos caracteres homocigóticos, pero el procedimiento debe repetirse en todos los períodos de reproducción.
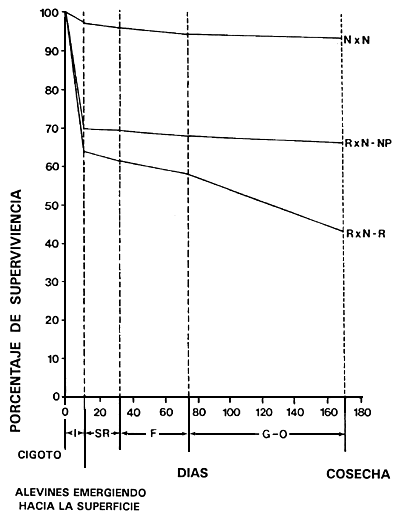
Figura 8. Efecto del color corporal rojo sobre la viabilidad de la tilapia del Nilo. Es un ejemplo de efecto pleiotrópico negativo. Se compara la viabilidad de los peces rojos (R × N-R) con la de los hermanos que poseen pigmentación normal (R × N-NP) y con una población control en la que no existen peces de color rojo (N × N). Se muestra la viabilidad en el transcurso de la incubación artificial de huevos (1), reversión del sexo (RS), producción de alevines (F) y crecimiento. En el momento de la cosecha, el peso medio de los peces era de 250 g.
Fuente: Tomado de El Gamal, A.R.A.L. 1987. Reproductive performance, sex ratios, gonadal development, cold tolerance, viability and growth of red and normally pigmented hybrids of Tilapia áurea and T.Nilotica. Disertación doctoral, Auburn University, Alabama, EE.UU.
El piscicultor utiliza este tipo de selección cuando quiere fijar el carácter recesivo y eliminar el carácter dominante y cuando los caracteres están producidos por un gen autosómico que muestra dominancia completa. El término “recesivo” no denota inferioridad y el término “dominante” no indica superioridad. Ambos términos se refieren únicamente a la forma en que se expresan los alelos y a los caracteres a los que dan lugar. Se ha descubierto que muchos caracteres recesivos son muy valiosos en diferentes plantas y animales cultivados y se han puesto en práctica programas de cría para conseguir su fijación. Los términos “inferior” y “superior” sólo deben aplicarse a los caracteres después de que hayan sido objeto de una evaluación biológica y económica.
Si el carácter deseado es el carácter recesivo producido por un gen que muestra dominancia completa, el programa de cría selectiva que debe utilizarse para fijarlo y producir una población pura es sencillo y bastará con una sola operación de selección. Todo lo que hay que hacer es reservar los peces que poseen el carácter recesivo y descartar los que presentan el carácter dominante.
Desde el punto de vista de su constitución genética, los peces con el carácter dominante no deseado son homocigóticos dominantes o heterocigóticos. Teniendo en cuenta que todos los peces que poseen al menos un alelo dominante expresan el carácter dominante no deseado, al descartar todos los peces con el carácter dominante se eliminarán todas las copias del alelo dominante. A la inversa, todos los peces que presentan el carácter recesivo deseado son homocigóticos recesivos, lo que significa que cuando se seleccionen ninguno de ellos poseerá una copia del alelo dominante no deseado. La población seleccionada sólo poseerá, por tanto, el alelo recesivo y se habrá conseguido una población pura.
Supongamos, por ejemplo, que un piscicultor posee tilapias del Nilo de pigmentación normal y de color rosado y decide cultivar solamente estas últimas. Puede conseguir una población pura con un programa de cría selectiva de una sola fase, que consistirá en descartar todos los peces de pigmentación normal. El color corporal rosado es el carácter recesivo y está originado por el alelo recesivo b (genotipo BB); el color corporal normal es el carácter dominante y está producido por el alelo dominante B (genotipos BB y Bb).
Al descartar todos los ejemplares de tilapia de pigmentación normal se eliminarán todos los alelos B de la población y sólo quedarán peces de color rosado, Como son homocigóticos recesivos bb, en la población de peces reproductores seleccionados sólo persistirá el alelo b. Así pues, con un único procedimiento de selección se habrá conseguido una población pura de color rosado (Figura 9).
Con este programa también se pueden fijar los caracteres recesivo o dominante si la acción génica es de dominancia incompleta, y el carácter homocigótico si la acción génica es aditiva. En ambos casos, existen dos genotipos homocigóticos que pueden ser identificados y aislados. Al descartar los caracteres no deseados, pueden crearse poblaciones puras en una sola operación de selección.
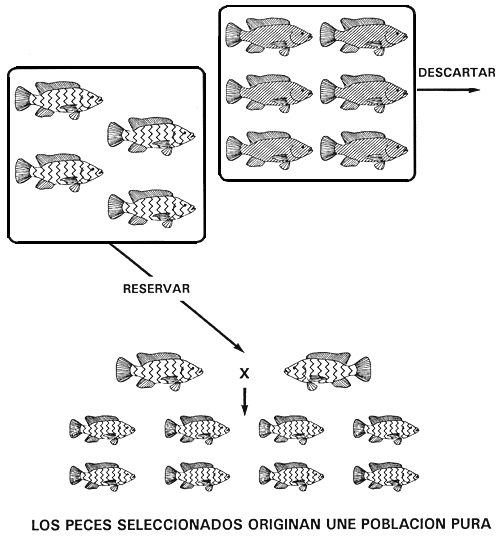
Figura 9. Programa de cría selectiva para obtener una población pura de tilapia del Nilo de color rosado. Si se descartan todos los peces de pigmentación normal (carácter dominante), se eliminarán todas las copias del alelo dominante B. Sólo existirán entonces peces de color rosado (carácter recesivo) y como son homocigóticos recesivos bb, la población seleccionada de peces reproductores de color rosado engendrará una línea pura, es decir, de color rosado. Las representaciones gráficas de los caracteres son las mismas que se utilizaron en la Figura 2. Este sencillo procedimiento permite fijar cualquier carácter recesivo.
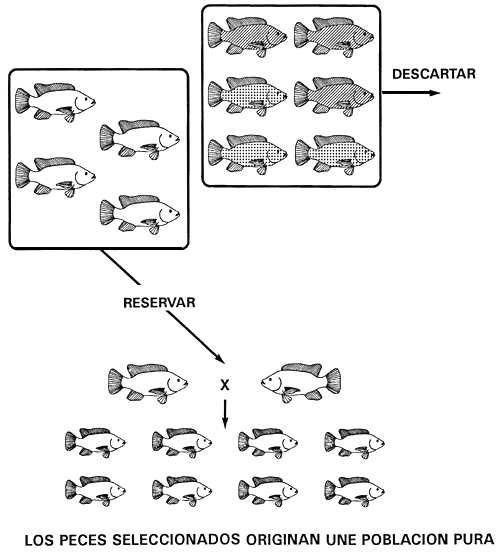
Figura 10. Programa de cría selectiva para conseguir una población pura de tilapia dorada de Mozambique. Si se descartan todos los peces melánicos (de color negro [GG] y bronceado [Gg]) se eliminarán todas las copias del alelo dominante G. así, sólo existirán peces de color dorado (carácter recesivo) y puesto que son homocigóticos recesivos gg, la población seleccionada de peces reproductores de color dorado engendrará una línea pura, esto es, de color dorado. Las representaciones gráficas de los caracteres son las mismas que las utilizadas en la Figura 4.
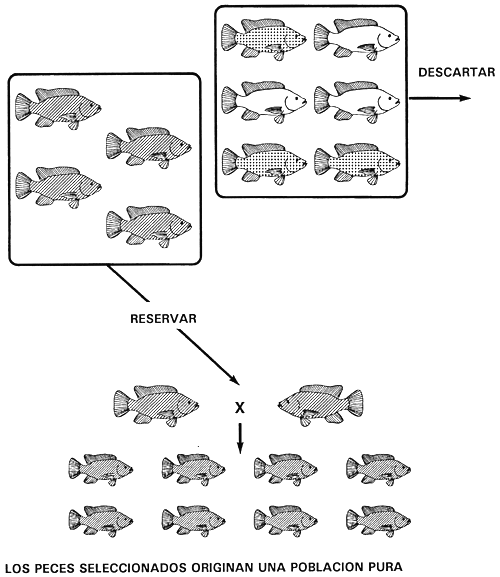
Figura 11. Programa de cría selectiva para obtener una población pura de tilapia de Mozambique de color negro. Si se descartan todos los peces de color bronce (carácter heterocigótico [Gg]) y de color dorado (carácter recesivo [gg]) se eliminarán todas las copias del alelo recesivo g. Sólo quedarán peces de color negro (carácter dominante) y puesto que son homocigóticos dominantes GG, la población seleccionada de peces reproductores de color negro engendrará una línea pura, de color negro. Las representaciones gráficas de los caracteres son las mismas que las utilizadas en la Figura 4.
Supongamos, por ejemplo, que un piscicultor tiene peces de color negro, bronceado y dorado en su población de tilapia de Mozambique. Para conseguir una población pura de peces de color dorado o negro sólo necesita efectuar una operación de selección. El color dorado es el carácter recesivo y está producido por el genotipo homocigótico recesivo (qg); el color negro es el carácter dominante y está controlado por el genotipo homocigótico dominante (GG); el color bronceado es el carácter heterocigótico (Gg).
Se puede obtener una población pura de tilapia dorada descartando todos los peces de color negro y bronceado (Figura 10). A la inversa, para obtener una población pura de tilapia negra basta con descartar todos los peces de color dorado y bronceado (Figura 11).
El mismo razonamiento vale también para los caracteres controlados por dos o más genes. Si el carácter deseado está controlado por un solo genotipo homocigótico, es posible obtener una población pura en una sola operación de selección.
Por ejemplo, si un cultivador que tiene una población de carpa común con los cuatro caracteres en cuanto a la configuración de las escamas quiere obtener una población pura de escamas reducidas, lo único que debe hacer es reservar los peces con el carácter escamas dispersas y descartar todos lo demás. Tres de los caracteres correspondientes a las escamas tienen un reducido número de escamas, pero sólo el carácter escamas dispersas está controlado por un genotipo homocigótico (ss. nn) y, por tanto, es el único que puede originar una línea pura. Los caracteres liso (ss, Nn) y escamas en línea (SS.Nn. y Ss,Nn) son caracteres no deseados, porque son heterocigóticos en el locus N, y, por consiguiente, si se aparean dos peces con los caracteres liso y/o escamas en línea, el 25% de la descendencia morirá (los peces homocigóticos NN, independientemente del genotipo en el locus S; véase la Figura 6). También los caracteres liso y escamas en línea son caracteres no deseados, porque los peces que los poseen sufren varios efectos pleiotrópicos negativos, entre ellos una baja tasa de crecimiento y una escasa viabilidad.
Si se descartan todos los peces lisos, con escamas en línea y cubiertos de escamas, se eliminarán todos los alelos S y N. En la población seleccionada sólo habrá peces con el carácter escamas dispersas y al ser homocigóticos recesivos (ss, nn) los alelos s y n serán los únicos que permanecerán en la población de peces reproductores seleccionados. En consecuencia, una única operación de selección originará una población pura de carpa común de escamas dispersas (Figura 12).
La selección no puede fijar un carácter controlado por más de un genotipo, es decir, no se puede fijar un carácter dominante y crear una población pura si el carácter está controlado por un gen que presenta dominancia completa. A la inversa, tampoco es posible eliminar mediante selección un carácter recesivo (y el alelo recesivo) cuando está controlado por un gen con dominancia completa. Aunque un programa de cría selectiva no puede tener éxito en estas circunstancias, muchas veces se utiliza, equivocadamente, para eliminar caracteres no deseados, controlados (supuestamente) por alelos recesivos.
Dos posibles razones explican que no se pueda conseguir el objetivo deseado. La primera es que la mayor parte de los caracteres no deseados son anomalías causadas por factores no genéticos (perturbaciones ambientales o anomalías del desarrollo). Muchos consideran que todos los caracteres anormales son caracteres mutantes producidos por alelos recesivos, pero esto sólo es cierto en algunos casos. Si el carácter no deseado está causado por un factor no genético, la selección no dará resultado positivo, porque ningún programa de cría selectiva puede fijar o eliminar caracteres no heredables.
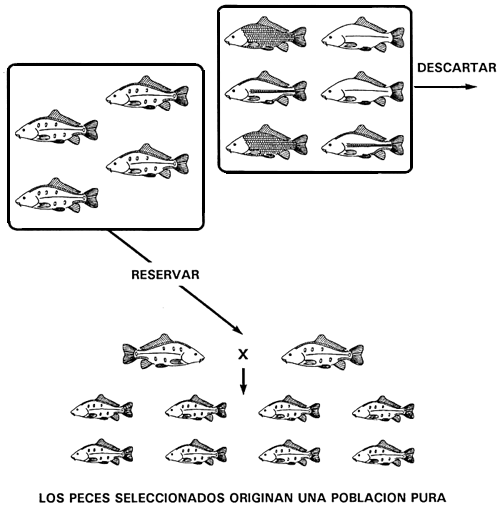
Figura 12. Programa de cría selectiva para producir una población pura de carpa común de escamas dispersas. Si se descartan todos los peces cubiertos de escamas, con escamas en línea y lisos (caracteres regulados por genotipos que poseen al menos un alelo dominante; véase la Figura 6), se eliminarán todas las copias de los alelos S y N. Sólo pervivirán los peces con el carácter escamas dispersas (recesivo) y dado que son homocigóticos recesivos ss, nn, la población seleccionada de peces reproductores de escamas dispersas originará una línea pura, que tendrá únicamente el carácter escamas dispersas. Aun en el caso de que un conjunto de caracteres sean controlados por dos o más genes, si el carácter deseado es el carácter recesivo (todos los loci determinantes del carácter son homocigóticos recesivos), la selección permitirá conseguir fácilmente una población pura.
Aun cuando una deformidad sea causada por un alelo recesivo, la selección no podrá originar una población libre de dicha anomalía si se trata de un gen autosómico con dominancia completa, porque el carácter dominante (normal) está regulado por uno de dos genotipos y es imposible distinguir los peces normales homocigóticos dominantes de los heterocigóticos.
Por ello, si el piscicultor descarta los peces recesivos (anómalos), la población seleccionada estará formada por peces con un solo carácter (normal), pero dos genotipos y todos los peces reproductores heterocigóticos normales seleccionados tendrán una copia del alelo recesivo no deseado. Al poseer ambos alelos, la población seleccionada no podrá originar una línea pura. Si se aparean dos peces reproductores heterocigóticos normales seleccionados, su descendencia tendrá el carácter anómalo no deseado (recesivo). Este tipo de selección reducirá la frecuencia de un carácter recesivo no deseado, pero no podrá eliminarlo y originar una población pura sin la deformidad.
Supongamos, por ejemplo, que un cultivador con una población de tilapia del Nilo de color rojo y de pigmentación normal decide producir únicamente tilapias de color rojo. No lo conseguirá descartando todos los peces de pigmentación normal, porque los peces reproductores rojos seleccionados engendrarán descendencia de color rojo y de pigmentación normal. La razón es que el rojo es el carácter dominante y está controlado por el alelo dominante R (genotipos RR y Rr), mientras que la pigmentación normal es el carácter recesivo y está producido por el alelo recesivo r.(genotipo rr). Como los genotipos homocigótico dominante y heterocigótico producen caracteres idénticos de color rojo, algunos de los peces reproductores rojos seleccionados (los heterocigóticos) poseerán una copia del alelo no deseado r. En el apareamiento de dos peces reproductores heterocigóticos de color rojo, una parte de la descendencia tendrá pigmentación normal (Figura 13).
Lo mismo ocurre con los caracteres controlados por dos o más genes. Si el carácter deseado lo produce más de un genotipo y si al menos uno de los genes puede estar en homocigosis o en heterocigosis, la selección no puede originar una población pura. Como en el caso de los caracteres controlados por un solo gen, algunos de los peces reproductores seleccionados poseerán una copia del alelo (o de los alelos) recesivo no deseado y si se aparean dos heterocigotos su descendencia poseerá el carácter (o caracteres) no deseado.
Por ejemplo, un cultivador con una población de carpa común con los cuatro caracteres relativos a la configuración de las escamas no podrá conseguir una población pura de carpa común cubierta de escamas mediante selección. La causa es que los peces reproductores cubiertos de escamas seleccionados poseerán dos genotipos -SS, nn o Ss, nn-y es imposible obtener peces heterocigóticos cubiertos de escamas a partir de peces homocigóticos cubiertos de escamas. Podrá eliminar el alelo dominante N descartando todos los peces con los caracteres escamas en línea y liso (se puede descartar un alelo dominante en una sola operación de selección); los peces que poseían dos copias del alelo N. fueron descartados de forma natural y murieron. Sin embargo, no podrá eliminar el alelo no deseado s descartando el carácter escamas dispersas que aún subsiste. En el apareamiento de dos peces reproductores heterocigóticos seleccionados cubiertos de escamas (Ss,nn), una parte de la descendencia serán peces con el carácter escamas dispersas y otra peces cubiertos de escamas (Figura 14).
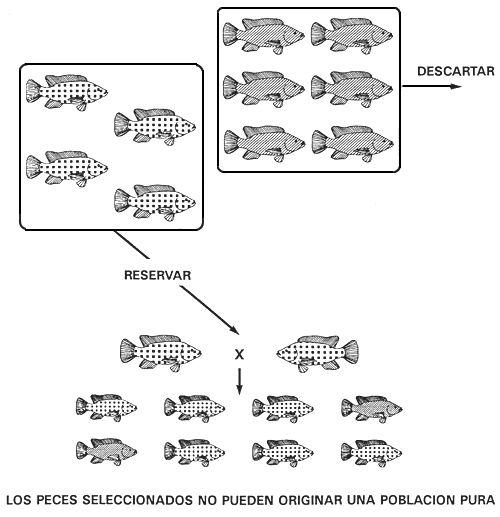
Figura 13. Programa de cría selectiva utilizado (sin resultados positivos) para conseguir una población pura de tilapia del Nilo de color rojo. Los peces con cuadrados son los de color rojo y los peces de pigmentación normal los que están cubiertos de franjas (en la figura 15 se utilizarán las mismas representaciones gráficas del color corporal). Si se descartan todos los peces de pigmentación normal (carácter recesivo) sólo habrá peces de color rojo (carácter dominante) en la población seleccionada. Como el color corporal rojo está producido por un alelo dominante que presenta dominancia completa, el rojo está causado por los genotipos homocigótico dominante (RR) y heterocigótico (Rr). Cada pez reproductor heterocigótico (Rr) reservado aportará una copia del alelo r a la población seleccionada y no podrá ser eliminado mediante selección. En el desove de los peces reproductores rojos, el apareamiento de dos heterocigotos (Rr) produce algunos individuos de pigmentación normal (el apareamiento que se representa en la figura). No es posible eliminar los caracteres recesivos descartándolos, pues reaparecerán cuando desoven los peces reproductores seleccionados. Así pues, si el carácter deseado es el carácter dominante y puede ser producido tanto por el genotipo homocigótico dominante como por el genotipo heterocigótico, la selección no puede originar una línea pura.
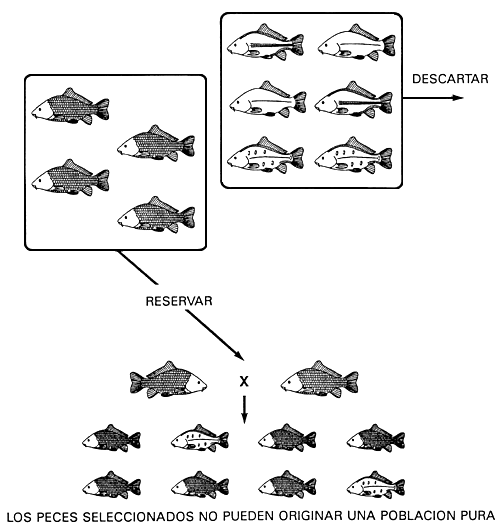
Figura 14. Programa de cría selectiva utilizado para producir (sin conseguirlo) una población pura de carpa común cubierta de escamas. Si se descartan todos los peces con los caracteres escamas en línea, escamas dispersas y liso, la población seleccionada estará formada por peces cubiertos de escamas. La operación de selección eliminará todas las copias del alelo dominante no deseado N porque se descartarán todos los heterocigotos, es decir, los caracteres en línea y liso (los homocigotos fueron descartados por la selección natural porque el genotipo NN es letal). Lamentablemente, este programa de cría selectiva no originará una población pura porque no se puede eliminar el alelo recesivo no deseado s descartando el carácter recesivo (escamas dispersas). Como el carácter cubierto de escamas está producido por un alelo que presenta dominancia completa, continuarán existiendo peces cubiertos de escamas, tanto homocigóticos (SS, nn) como heterocigóticos (Ss, nn). Todos los peces reproductores heterocigóticos seleccionados cubiertos de escamas aportarán una copia del alelo recesivo s a la población seleccionada y, por tanto, no podrá ser eliminado mediante la selección. Durante el desove de los peces reproductores con escamas seleccionados, el apareamiento de dos heterocigotos engendrará una parte de la descendencia con el carácter escamas dispersas (apareamiento que se representa en la figura). Como en el caso de un carácter producido por un solo gen, si el carácter deseado está producido por más de un genotipo y uno de los genotipos es heterocigótico, no será posible obtener una población pura mediante selección.
Prueba de la descendencia: Puesto que la selección para descartar un carácter recesivo no puede fijar un alelo dominante y originar una población pura (cuando la acción génica es de dominancia completa), para conseguirla habrá que utilizar otro programa de cría. La prueba de la descendencia es el único procedimiento que permite eliminar un alelo recesivo no deseado. Consiste en averiguar el genotipo de uno de los padres determinando los caracteres de su descendencia. Una vez que se han descifrado los caracteres de los padres dominantes, se utiliza la selección para conservar los homocigotos y descartar los heterocigotos, con lo cual se eliminarán todas las copias del alelo recesivo, se fijará el alelo dominante y se obtendrá una población pura.
Para ello se empareja y aparea a peces que tengan el carácter dominante con peces de prueba (cuyo genotipo se conoce). Normalmente, los peces de prueba poseen el carácter recesivo, porque son homocigóticos recesivos (no es necesario realizar experimentos para establecer el genotipo de un fenotipo recesivo). Como el pez de prueba puede producir gametos que tengan tan sólo un alelo recesivo, el pez dominante es el progenitor que determina los caracteres de su descendencia (véanse las figuras 3c y 3e). Si el parental dominante es homocigoto, toda la descendencia tendrá el carácter dominante, pero si es heterocigoto, la mitad de la descendencia tendrá el carácter dominante y la otra mitad el carácter recesivo (Figura 3e).
El hecho de que un descendiente posea el carácter recesivo permite asegurar que el padre dominante es heterocigoto y debe ser descartado. Si no se detecta ningún descendiente con el carácter recesivo en una muestra al azar de un mínimo de 20 ejemplares, se puede afirmar que el parental dominante es homocigoto y conservarlo para fines de reproducción. Una vez que se ha realizado la prueba de la descendencia con un número suficiente de machos y hembras y se ha establecido que son homocigotos dominantes, se descartan todos los demás peces (incluida la descendencia obtenida a raíz de la prueba de descendencia). Ahora será posible conseguir una población pura que producirá únicamente descendencia con el carácter dominante.
Por ejemplo, un cultivador de tilapia con una población de tilapia del Nilo de color rojo y de pigmentación normal puede crear una población pura de color rojo aplicando la prueba de la descendencia para identificar y conservar los peces rojos homocigóticos (fifi) e identificar y descartar a los peces de color rojo heterocigóticos (Rr). Antes de iniciar el programa de la prueba de descendencia, el piscicultor debe descartar todos los peces de pigmentación normal, excepto los que servirán como peces de prueba.
El piscicultor realiza la prueba de la descendencia en los peces rojos apareando a uno de ellos con un pez de prueba (en este caso los peces de prueba son tilapias de pigmentación normal porque son homocigóticos recesivos, rr). Los parentales de color rojo homocigóticos (RR) sólo engendrarán descendencia de color rojo y serán reservados, mientras que los parentales de color rojo heterocigóticos (Rr) engendrarán peces de color rojo y de pigmentación normal en igual número y serán descartados. Después de realizar la prueba de la descendencia en una generación, el piscicultor tendrá una población seleccionada de peces reproductores de color rojo que sólo engendrará descendencia de ese color, porque todos los peces reproductores seleccionados de color rojo son homocigóticos fifi (Figura 15).
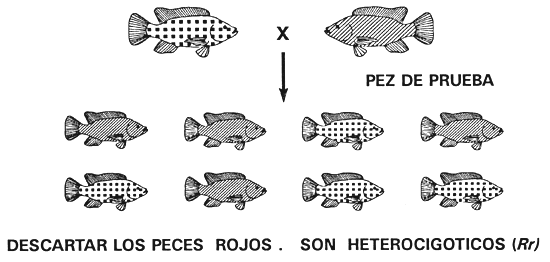
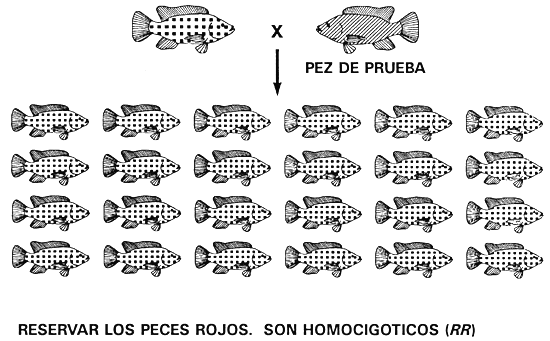
Figura 15. Diagrama esquemático de la prueba de descendencia, utilizada para obtener una población pura de tilapia del Nilo de color rojo. La prueba se aplica para identificar y descartar los peces rojos heterocigóticos (Rr) e identificar y reservar a los peces rojos homocigóticos (RR). Para averiguar el genotipo de un pez de color rojo se aparea a un pez de prueba (un pez de pigmentación normal [rr]) y se determinan los caracteres de su descendencia. En el apareamiento de la parte superior, se clasifica el pez rojo como heterocigótico (Rr) y se descarta, porque la mitad de su descendencia posee pigmentación normal. En el apareamiento de la parte inferior, se considera que el pez rojo es homocigótico (RR) y se reserva porque el muestreo al azar de al menos 20 de sus crías no denotó la existencia de ningún pez pigmentado. Mediante la prueba de descendencia se obtendrá una población de peces reproductores seleccionados de color rojo, todos los cuales son homocigóticos RR. Esos peces sólo engendrarán descendientes de color rojo. Las representaciones gráficas de los caracteres son las mismas utilizadas en la Figura 13.
El principio es el mismo en el caso de que el carácter deseado esté controlado por dos o más genotipos y por más de un gen. Por ejemplo, si un cultivador de carpas cuya población de carpa común tiene las cuatro configuraciones de escamas quiere conseguir una población pura cubierta de escamas deberá efectuar la prueba de la descendencia. Los peces cubiertos de escamas son producidos por los genotipos SS, nn y Ss, nn. En este caso, se utilizan como peces de prueba las carpas con escamas dispersas porque son homocigóticos recesivos (ss, nn). Antes de comenzar la prueba de la descendencia, deberá descartar todos los peces con los caracteres en línea y liso y la mayor parte de los peces de escamas dispersas, aunque conservará algunos de ellos como peces de prueba.
La prueba de la descendencia se efectúa para identificar y reservar peces cubiertos de escamas homocigóticos en el locus S (Ss, nn) e identificar y descartar peces cubiertos de escamas heterocigóticos en el locus S (SS,nn). Si el parental cubierto de escamas es homocigótico en el locus S (SS,nn), toda la descendencia tendrá el carácter cubierto de escamas, pero si es heterocigótico (Ss, nn), la mitad de la descendencia tendrá el carácter cubierto de escamas y la otra mitad el carácter escamas dispersas. Después de realizar la prueba de la descendencia en el transcurso de una generación, el piscicultor tendrá una población seleccionada en la que todos los peces reproductores cubiertos de escamas serán SS, nn, y que sólo engendrará peces cubiertos de escamas (Figura 16).
Si en la prueba de la descendencia se puede efectuar el desove artificial por presión un solo pez de prueba es suficiente para llevar a cabo la prueba en un número de peces que tengan el carácter dominante. Además, no es necesario criar varios miles de peces de cada familia, sino tan sólo una muestra al azar de 100–200 huevos fecundados, si la supervivencia hasta la fase temprana de alevín es buena. Así pues, la prueba de descendencia no es un método muy costoso, pues puede realizarse en un pequeño criadero, en acuarios, en pequeñas piscinas de plástico o en jaulas suspendidas en un estanque que no ha de tener una extensión superior a 0,04 ha. Mientras se están criando las familias, los peces reproductores dominantes deberán permanecer en jaulas o tanques aislados, a fin de reservarlos o descartarlos una vez se haya examinado la progenie. Pueden mantenerse varios peces en un mismo tanque si se marcan con chapas en las aletas. Para obtener una población pura sólo habrá que efectuar la prueba de la descendencia en una generación y no será preciso realizarla de nuevo salvo que se adquieran en otra piscifactoría peces con el alelo recesivo no deseado o que éste aparezca por mutación.
Cuando el carácter deseado es heterocigótico, ningún programa de cría puede originar una población pura, porque los peces con el carácter deseado no pueden engendrar una línea pura. Los heterocigotos producen dos tipos de gametos en número igual. Por ello, cuando se aparean originan una descendencia con tres caracteres (la selección para el carácter heterocigótico sólo puede realizarse en el caso de caracteres controlados por genes autosómicos con dominancia incompleta o con acción génica aditiva). Descartar los dos caracteres homocigóticos y seleccionar únicamente el carácter heterocigótico entrañará el fracaso del programa de cría selectiva, porque sólo la mitad de la descendencia engendrada por los peces reproductores seleccionados tendrá el carácter deseado.
Por ejemplo, un piscicultor que posee una población de tilapia de Mozambique con los caracteres negro, bronceado y dorado no podrá conseguir una población pura de peces de color bronceado. Si utiliza un programa de cría selectiva descartará los peces dorados (carácter recesivo) y negros (carácter dominante) y conservará los de color bronceado (los heterocigotos). Los peces reproductores seleccionados de color bronceado engendrarán una descendencia con los tres caracteres (Figura 17).
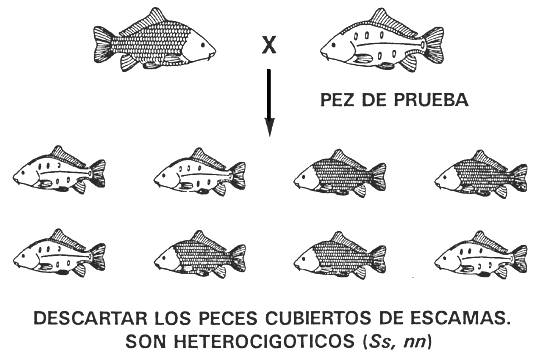
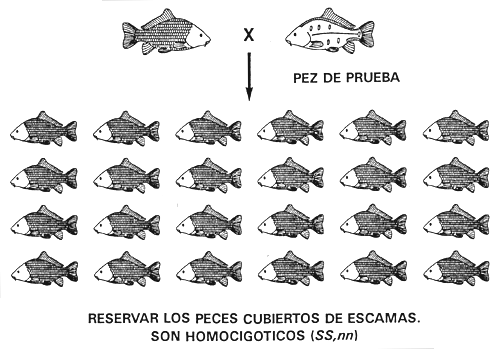
Figura 16. Diagrama esquemático del programa de prueba de la descendencia que debe utilizarse para conseguir una población pura de carpa común cubierta de escamas. La prueba de descendencia sirve para identificar y descartar los peces cubiertos de escamas heterocigóticos (Ss, nn) e identificar y conservar los peces cubiertos de escamas homocigóticos (SS,nn). Para averiguar el genotipo de un pez cubierto de escamas se aparea a un pez de prueba (que ha de tener el carácter escamas dispersas [ss,nn]) a fin de determinar los caracteres de la descendencia. En el apareamiento de la parte superior, el pez cubierto de escamas es heterocigótico (Ss,nn) y se descarta porque la mitad de su descendencia presenta el carácter escamas dispersas. En el apareamiento de la parte inferior, se considera que el pez cubierto de escamas es homocigótico (SS,nn) y se reserva, porque en una muestra al azar de al menos 20 de sus crías no se detectó el carácter escamas dispersas. La prueba de la descendencia originará una población seleccionada de peces reproductores cubiertos de escamas, todos los cuales han sido acreditados como homocigóticos SS, nn, que sólo engendrará peces cubiertos de escamas.
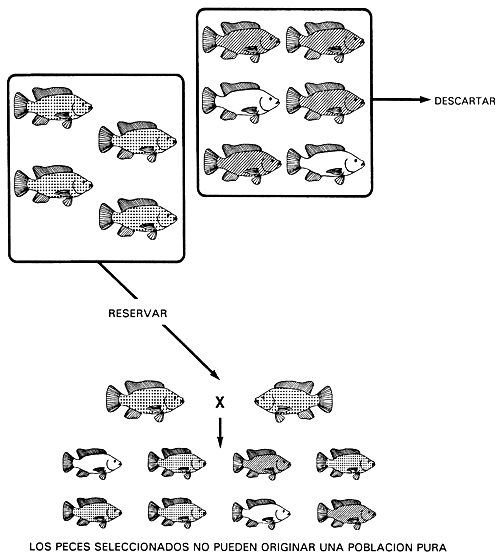
Figura 17. Programa de cría selectiva utilizado para obtener, sin éxito, una población pura de tilapia de Mozambique de color bronceado. En este caso, se descartan los peces homocigóticos de color negro (GG) y dorado (gg) y se conservan los de color bronceado (Gg). Dado que éstos son heterocigotos, la población seleccionada no podrá engendrar una línea pura. Sólo la mitad de la descendencia será de color bronceado. Las representaciones gráficas del color corporal son las mismas que se utilizaron en la Figura 4.
Para obtener únicamente peces de color bronceado deberá aparear peces dorados y negros. En este tipo de apareamiento (entre los dos homocigotos; se ilustra en la Figura 5c) toda la descendencia será heterocigótica (Figura 18).
La única condición necesaria para desarrollar este programa de cría consiste en identificar con seguridad el sexo de los peces. Si el piscicultor comete un error en la identificación del sexo de un solo pez nacerán crías de color dorado o negro. Para este plan de apareamiento hacen falta al menos dos estanques para la descendencia. En uno de ellos se depositarán hembras doradas y machos negros y en el otro hembras negras y machos dorados.
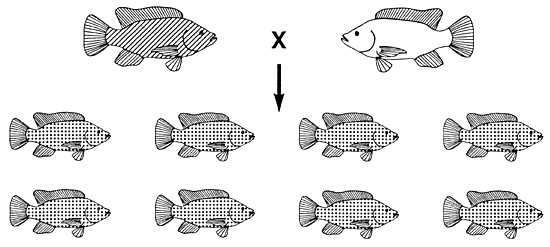
Figura 18. Programa de cría que debe utilizarse para producir una población de tilapia de Mozambique de color bronceado. Sólo se obtendrán peces de color bronceado (Gg) apareando peces negros (GG) y dorados (gg). Si el piscicultor quiere cultivar únicamente peces de color bronceado, debe aplicar este programa de apareamiento en cada período de reproducción. Las representaciones gráficas del color corporal son las mismas que en la Figura 4.
El volumen de datos que es necesario registrar en el caso de los caracteres cualitativos es mucho menor que cuando se trata de caracteres cuantitativos. Una de las informaciones que se debe consignar es la descripción de caracteres anormales o “mutantes” que se observan cada año. Los piscicultores no tienen una memoria infalible y el hecho de que los peces anómalos sean muy inusuales les hace ver, muchas veces, más de los que hay en realidad. El recuento de esos caracteres indicará al piscicultor qué tipo de anomalías se han registrado y le permitirá saber si alguna de ellas constituye un problema. Además, estos datos le darán información sobre otros caracteres cualitativos que existen en la población piscícola.,
El piscicultor deberá disponer de una hoja de datos para registrar la fecha, el nombre del carácter (puede inventar un nombre si aún no existe) y una breve descripción del mismo. Registrará, además, el número de peces que presentan dicho carácter y el número total de peces producidos, para conocer la frecuencia de cada deformidad. Si la frecuencia aumenta en el transcurso del tiempo, sabrá que se enfrenta con un problema potencial. En el cuadro 7 se ofrece un ejemplo de hoja de datos.
Cuadro 7. Ejemplo de hoja de datos para registrar los caracteres anómalos y/o otros caracteres cualitativos que se observen.
| Fecha: 23 de marzo de 1995 | |||
| Especie: tilapia del Nilo | |||
| Volumen producido: 25.500 | |||
| Carácter | Descripción | Número | Frecuencia |
| Ausencia de cola | ausencia de cola; | 3 | 0,000117 |
| ausencia de pedúnculo | |||
| caudal | |||
| Semiopérculo | opérculo derecho corto; | 5 | 0,000196 |
| agallas parcialmente | |||
| expuestas | |||
| Cuerpo achaparrado | enano; tronco | ||
| anormalmente corto | 2 | 0,000078 | |
Si el piscicultor va a poner en práctica un programa de cría selectiva para fijar un carácter cualitativo, debe registrar la fecha(s) en que efectuó la selección, el número de peces reservados y descartados y el estanque(s) en que depositó los peces reproductores. Cuando desoven los peces reproductores seleccionados, registrará si engendraron una población pura. En el Cuadro 8 se reproduce una hoja de datos de estas características.
Cuadro 8. Ejemplo de hoja de datos para un programa de cría selectiva. En este caso, se trata de un programa para conseguir una población pura de tilapia de Mozambique.
| Fecha: 1 de junio de 1995 | |
| Especie: iilapia de Mozambique | |
| Programa de cría: originar una población pura de peces dorados | |
| Carácter descartado | Número descartado |
| negro | 456 |
| bronceado | 935 |
| Carácter reservado | Número reservado |
| dorado | 204 hembras y 196 machos |
¿Se consiguió un población pura?
Sí, el 23 de junio de 1995, los peces reproductores seleccionados engendraron únicamente peces de color dorado
Sí, el 5 de julio de 1995, los peces reproductores seleccionados engendraron únicamente peces de color dorado
Sí, el 17 de julio de 1995, los peces reproductores seleccionados engendraron únicamente peces de color dorado
Para realizar una prueba de la descendencia el piscicultor registrará la fecha de cada apareamiento y sus resultados e indicará qué peces reservó y descartó y dónde colocó los peces reproductores seleccionados. Cuando estos desoven, deberá registrar si engendraron una población pura. El cuadro 9 reproduce una hoja de datos de estas características. El piscicultor debe registrar también dónde permaneció cada ejemplar y cada familia hasta que los peces fueron reservados o descartados durante la prueba de la descendencia.
Por último, si quiere producir heterocigotos, debe registrar en qué estanques se encuentra cada homocigoto, el número de peces de cada estanque y cuándo se depositaron; el número de peces reproductores a los que se les hizo desovar y en qué estanques; y el resultado conseguido. En el Cuadro 10 se reproduce una hoja de datos de estas características.
Cuadro 9. Ejemplo de hoja de datos para la prueba de la descendencia. En este caso, la finalidad es descartar peces rojos heterocigóticos (Rr) y reservar peces rojos homocigóticos (RR) para obtener una población pura de tilapia del Nilo de color rojo. Sólo figuran los datos de cada apareamiento para los peces 1–5.
| Fechas: 21 de mayo–15 de junio de 1995 | |||
| Especie: tilapia del Nilo | |||
| Prueba de la descendencia: prueba de la descendencia de peces rojos para descartar heterocigotos y reservar homocigotos. Se utilizaron como peces de prueba individuos de pigmentación normal. | |||
| Fecha | Peces reproductores rojos | Proporción de la descendencia (rojo:normal) | Descartados/reservados |
| 1 de junio | 1 | 156:0 | Reservados |
| 1 de junio | 2 | 73:70 | Descartados |
| 1 de junio | 3 | No desovaron | Descartados |
| 2 de junio | 4 | 75:0 | Reservados |
| 2 de junio | 5 | 56:61 | descartados |
| Número total de peces rojos reservados: 75 hembras y 49 machos | |||
| ¿Se obtuvo una población pura? | |||
| Sí, el 25 de julio de 1995 los peces reproductores seleccionados engendraron únicamente peces de color rojo | |||
| Sí, el 31 de julio de 1995 los peces reproductores seleccionados engendraron únicamente peces de color rojo. | |||
Cuadro 10. Ejemplo de hoja de datos que puede ser usado para recopilar los datos de un programa de cría dirigido a la produción de heterocigotos para engorda. En este caso en un programa de cría que cruza peces dorados con negros para producir una población totalmente bronceada de tilapia de Mozambique.
| Fecha: 25 de julio – 1 agosto 1995 | Especie: tilapia de Mozambique | |||
| Programa de cría: Cruzar dorado × negro para producir 100% bronceado | ||||
| Carácter | Fecha | Estanque No. | Número de machos | Número de hembras |
| Negro | 1 de junio 1995 | 23 | 125 | 230 |
| Bronceado | 1 de junio 1995 | 32 | 119 | 198 |
| Ejemplo de hoja | ||||
| Produción de pez bronceado | ||||
| Fecha | Estanque No. | Carácter/sexo | Carácter/sexo | Descendencia |
| 1 agosto | 2 | 23 machos dorados | 45 hembras negras | 8,300 bronceado |
| 1 agosto | 3 | 50 hembras doradas | 30 machos negros | 9,500 bronceado |
| 1 agosto | 4 | 45 hembras doradas | 32 machos negros | 10,300 bronceado |
Los programas de cría selectiva a los que se ha hecho referencia en este capítulo no extraña gran dificultad técnica y el volumen de datos que deben registrarse es mínimo. Los programas necesarios para obtener poblaciones puras son sencillos y la mayor parte de ellos pueden culminarse en un solo período de reproducción. En algunos casos, el objetivo se puede conseguir en un solo día. La selección se concluye en el momento en que se obtiene una población pura. Si ello no es posible, se puede preparar un programa de cría para producir únicamente el carácter deseado.
Los programas aquí descritos se han aplicado en importantes especies cultivadas para conseguir poblaciones que sólo tuvieran el carácter deseado. Por ejemplo, el carácter no deseado aleta dorsal deformada de la línea de tilapia azul de la Universidad de Auburn, se eliminó en una sola operación de selección porque estaba producido por un alelo dominante. En los organismos de pesca estatales de Virginia Occidental y de Pensilvania (EE.UU.) se utilizan la selección y el apareamiento de homocigotos para producir truchas arco iris doradas (mediante selección) y palomino (mediante el apareamiento de homocigotos) para repoblar las aguas de propiedad pública. En Europa muchos cultivadores de carpa común fijaron en sus poblaciones la configuración de escamas dispersas descartando todos los demás caracteres relativos a las escamas. Por último, los cultivadores de peces ornamentales emplean habitualmente estos programas para obtener peces más valiosos.
Antes de efectuar la selección, el piscicultor o la cooperativa piscícola deben realizar un análisis económico que determine si es conveniente utilizar un programa de cría selectiva. Dos son los datos que deben conocerse: el valor de mercado de los caracteres y su costo de producción, el cual se establecerá analizando los efectos pleiotrópicos, como la tasa de crecimiento, la supervivencia, etc. Sí de la evaluación se desprende que la fijación de un color corporal más atractivo, etc., permitirá al piscicultor aumentar los beneficios, éste deberá adoptar un programa de cría selectiva. En cambio, si indican que el incremento de los costes de producción superará el valor de mercado del producto, no deberá ponerlo en práctica.
El piscicultor debe saber también si es innecesario utilizar la selección porque no es preciso corregir el problema planteado. En todas las poblaciones existen individuos con deformidades. En el medio natural, el número de peces con deformidades que se observa es muy reducido porque generalmente son devorados por los depredadores. En cambio, en los estanques son más numerosos porque los piscicultores excluyen de ellos a los depredadores. Sin embargo, su incidencia es escasa todavía. Muchas veces, los piscicultores creen que ello constituye un problema porque los peces de aspecto extraño son muy llamativos y pueden recordarlos. Si la frecuencia de una malformación es inferior a 1 de cada 250 peces, el problema no necesita ser corregido, pero si se manifiesta una malformación en 1 de cada 100–200 peces, habrá que estudiar el problema y, posiblemente, recurrir a la selección si la malformación es heredable.
Por último, cuando un piscicultor pone en práctica un programa de cría selectiva, debe disponer de 100 a 200 peces reproductores seleccionados. Es necesario reservar un número tan elevado de peces para poder aparear al menos 25 machos y 25 hembras en cada período de reproducción, de manera que la consanguinidad no alcance un nivel excesivo.
![]()
![]()
![]()