![]()
![]()
![]()
Los caracteres que tienen importancia para la producción son los caracteres cuantitativos, como la longitud, el peso, la conversión del pienso y los huevos por kg en las hembras,. Cuando se trata de estos caracteres, el piscicultor no separa los peces en categorías descriptivas no coincidentes como ocurre en el caso de los caracteres cualitativos (por ejemplo, peces de color rojo frente a otros de pigmentación normal). Antes bien, cada carácter es único (por ejemplo, la longitud) y las diferencias entre los peces se establecen mediante su medición. El valor numérico que se atribuye a un pez cuando se mide es su valor fenotípico.
El hecho de que el valor fenotípico no se atribuya en función de una categoría descriptiva (por ejemplo, el color) sino de la medición (por ejemplo, la longitud en milímetros), significa que la selección no se basa en el aspecto sino en valores numéricos. Por ejemplo, en un carácter cualitativo como el color, los piscicultores seleccionan peces de un color y descartan los de otro color; la decisión de seleccionarlos o descartarlos se basa únicamente en la categoría descriptiva. Pero cuando la selección tiene por objeto un carácter cuantitativo, seleccionan peces que alcanzan o superan un valor numérico yjdescartan los que quedan por debajo del mismo; la decisión de seleccionarlos o descartarlos se basa en valores numéricos.
Los programas de cría selectiva proyectados para los caracteres cualitativos tienen como finalidad originar poblaciones puras. Son programas sencillos para eliminar un alelo no deseado y fijar un alelo deseado, de manera que la población produzca únicamente el carácter deseado. Normalmente, esto se consigue en el transcurso de una generación (en muchos casos mediante una sola operación de selección) y en ese momento, conseguido el objetivo propuesto, el programa se da por concluido.
En cambio, los programas encaminados a mejorar caracteres cuantitativos son más complejos. Exigen un nivel tecnológico más elevado y registrar un mayor volumen de datos. Además, no tienen una duración determinada, pues al no ser posible producir poblaciones puras para estos caracteres, deben proseguir año tras año. La finalidad de estos programas es mejorar gradualmente la media de la población en cada generación. Es importante tener en cuenta que los programas de cría selectiva para caracteres cuantitativos no concluyen nunca e intentar obtener mejoras pequeñas pero graduales en el transcurso de muchas generaciones. Se pueden conseguir mejoras muy importantes en la primera generación o en las dos primeras generaciones, pero ello exigiría reducir drásticamente el tamaño de la población reproductora, ocasionando posiblemente depresión consanguínea, que malograría muchos de los beneficios conseguidos con la selección.
Otra razón por la que estos programas de cría selectiva tienen una duración ilimitada estriba en que si se interrumpen se pierde gran parte de la mejora conseguida. Es difícil prever la cuantía de la pérdida, pues depende del carácter, de la heredabilidad y de la población. Si el carácter está controlado únicamente por variación genética aditiva (h2 = 1,0), la media no se modificará si se interrumpe el programa de cría. Ahora bien, ningún carácter de interés productivo está controlado únicamente por variación genética aditiva (Cuadro 6).
Al interrumpir el programa de cría selectiva se perderá gran parte de la mejora conseguida, porque se perderá la mayor parte de la mejora debida a la variación genética de dominancia, toda la mejora que se debe a la variación genética epistática y una parte de la conseguida como consecuencia de la interacción entre el genotipo y el medio ambiente. Se conservará una parte de la mejora debida a la variación genética aditiva, aunque es difícil establecer la cuantía exacta.
Si la cría selectiva ha mejorado en gran medida la población antes de que se abandonara el programa, la media podría estabilizarse en un nivel bastante más alto que la media original. Pero si la mejora fue pequeña, caso muy probable, la media final podría ser sólo ligeramente más alta que la media original y la diferencia podría quedar enmascarada por los efectos ambientales anuales.
Antes de iniciar un programa de cría selectiva, el piscicultor debe preguntarse si es conveniente o necesario aplicarlo. La respuesta a esta pregunta depende de las circunstancias concretas y puede ser distinta incluso en el caso de dos piscicultores vecinos.
La decisión de aplicar o no un programa de cría selectiva no debe adoptarse a nivel regional, sino que debe ser de índole individual. Los agentes de extensión procuran (y consiguen) que todos los piscicultores de la región adopten nuevas tecnologías. Sin duda, es una actitud razonable porque el extensionista sabe que de esa forma aumentarán la producción, obtendrán mayores beneficios y habrá mayor disponibilidad de alimentos.
Como en cualquier otro aspecto de la tecnología, para adoptar un programa de cría selectiva es necesario analizar las circunstancias concretas de cada caso. En general, sólo deben utilizar estos programas los piscicultores que están en condiciones de efectuar y mantener registros de datos, son buenos administradores, adoptan y mantienen nuevas tecnologías y están dispuestos a incorporar la planificación a largo plazo en sus programas de gestión. Además, deben estar dispuestos a invertir el dinero y el tiempo adicional que exigen su puesta en práctica.
Algunos piscicultores (los que tienen piscifactorías pequeñas o producen peces para una o varias familias o para una pequeña aldea) nunca utilizarán la selección, ni se les debe exhortar a que lo hagan. Aquellos que no tienen los conocimientos necesarios para mantener un buen registro de datos, no cuentan con un número suficiente de estanques para dedicarlos al programa o no pueden o no quieren invertir dinero en él no deben utilizarla y no se les debe alentar a que lo hagan.
La finalidad del programa de cría selectiva es aumentar la productividad en el transcurso de varias generaciones (una generación se define como la sustitución de los peces reproductores por su descendencia). Está pensado para aumentar las tasas de crecimiento y el rendimiento de forma gradual a lo largo de muchos años. Si el piscicultor no tiene la posibilidad de incluir la planificación a largo plazo (1–10 años) en su programa de gestión abandonará el programa cuando sólo hayan transcurrido unos pocos meses y lo desaconsejará a otros piscicultores, argumentando que ese tipo de programas no dan buen resultado.
Aquellos piscicultores que no saben registrar bien los datos porque no saben leer o escribir, o que no saben medir los peces, no son buenos candidatos para poner en práctica un programa de cría selectiva, pues no sabrán aplicarlo correctamente. En cambio, si son serios, trabajan arduamente y aceptan y adoptan nuevas tecnologías, pueden llevarlo a la práctica si pueden contar con la colaboración de un agente de extensión y si asumen la responsabilidad de mantener un registro de datos y realizar el trabajo que conlleva la aplicación del programa.
Por último, si el piscicultor no es un buen administrador no debe aplicar un programa de cría selectiva. Aquellos que raramente examinan los estanques, que sólo alimentan a los peces o abonan los estanques cuando el agente de extensión les recuerda que deben hacerlo o que ponen poco empeño en aumentar la producción no deben utilizar estos programas. El piscicultor que no puede o no sabe manejar adecuadamente un estanque piscícola tampoco aplicará correctamente un programa de cría selectiva.
La principal finalidad de casi todos los programas de cría selectiva proyectados para mejorar los caracteres cuantitativos es aumentar la tasa de crecimiento, lo cual incrementará también el rendimiento.
Para aumentar el rendimiento existen básicamente dos procedimientos: el primero consiste en utilizar un programa de cría para conseguir la mejora de los peces que se están cultivando, y el segundo en mejorar el sistema de cultivo. Mientras que los programas de cría son costosos, muchas veces la mejora de la gestión no comporta gasto alguno.
Por consiguiente, si es posible aumentar la tasa de crecimiento y el rendimiento modificando las técnicas de manejo o adoptando otras distintas, éste debe ser el primer procedimiento a seguir. Para mejorar el cultivo, se pueden adoptar diversas medidas, como utilizar cal o fertilizantes, establecer en los estanques la densidad de población adecuada, almacenar alevines de mayor tamaño en los estanques de crecimiento o alimentar a los peces con una ración preparada.
La principal razón para intentar primero mejorar la gestión es que resulta menos costoso y da resultados inmediatos. Fertilizando adecuadamente los estanques y administrando pienso a los peces se puede duplicar o triplicar el rendimiento durante la primera temporada de crecimiento y la primera cosecha. Un programa de cría selectiva también permitirá aumentar el rendimiento, pero de forma gradual y a lo largo de muchos años. Por ello, para aumentar la productividad lo primero que hay que hacer es mejorar la capacidad de gestión y, en consecuencia, el sistema de cultivo de los peces.
Además, no es razonable modificar el sistema de cultivo una vez que se ha iniciado el programa de cría selectiva. Cuando se efectúa una selección para caracteres cuantitativos se intenta obtener dichos caracteres tal como se manifiestan en unas condiciones determinadas de cultivo. Es posible que los peces que dan mejores resultados en estanques abonados que tienen una alta densidad de población no sean los que mejor se comportan cuando la densidad de población es menor y son alimentados con una ración preparada. Si el carácter seleccionado es el crecimiento más rápido en unas condiciones determinadas de cultivo se seleccionan los peces cuyos genes dan los mejores resultados en esas condiciones. Por ello, es posible que al modificar el sistema de cultivo se hayan descartado aquellos genes que pueden dar buen resultado en las condiciones del nuevo sistema de gestión.
Es importante, pues, decidir si el programa de manejo que se está aplicando para la producción de peces en una determinada piscifactoría se utilizará también en el futuro. Probablemente, la introducción de pequeños cambios, como modificar la tasa de población en los estanques de 2.000 a 2.500 peces por hectárea o aumentar el tamaño medio de los alevines de 10 a 15 g, no tiene gran incidencia, pero los cambios de mayor entidad, como sustituir el abono orgánico como fuente única de nutrientes por alimentos granulados de gran calidad, constituyen una alteración sustancial del sistema de cultivo. Si se proyecta realizar cambios importantes es necesario posponer el programa de cría selectiva.
No es necesario normalizar las técnicas de cultivo utilizadas en una región antes de iniciar un programa de cría selectiva. Es posible que algunos piscicultores prefieran utilizar fertilizantes en lugar de alimentos granulados porque disponen de una fuente abundante y barata de estiércol; otros, en cambio, preferirán alimentar a los peces a base de grànulos para aumentar al máximo el rendimiento. Se pueden proyectar programas de cría selectiva para ambos tipos de piscicultores, siempre que estén dispuestos a invertir el tiempo y el dinero necesarios y dispongan de un número suficiente de estanques.
Sólo debe utilizarse un programa de cría selectiva si el piscicultor está decidido a dedicar el tiempo y el esfuerzo necesarios para aplicarlo satisfactoriamente. Si no quiere hacer el esfuerzo requerido, sea en concepto de mano de obra o de dinero, o se prevé que lo interrumpirá una vez que haya pasado el interés inicial, un piscicultor no debe comenzar un programa de cría selectiva.
A veces, los piscicultores se muestran interesados en aplicar un programa de cría selectiva pero afirman que no lo iniciarán hasta que no hayan solucionado todos los demás problemas. En realidad, nunca lo llevarán a la práctica, porque nunca podrán resolver todos los problemas. En efecto, cuando se supera una dificultad surge otro obstáculo para aumentar la producción.
El programa de cría selectiva debe iniciarse si el piscicultor está dispuesto a invertir el tiempo y el dinero requeridos porque tiene la convicción de que ése es el mejor procedimiento para aumentar la producción de alimentos y los beneficios, y si asume que se trata de objetivos a largo plazo. Por último, es importante también que comprenda que los programas de cría selectiva no tienen un plazo definido, porque siempre es posible realizar nuevas mejoras.
Antes de iniciar un programa el piscicultor deberá evaluar su conveniencia, cuantificar sus objetivos, saber cómo medir el o los caracteres, decidir qué tipo de programa aplicará, saber cómo evaluará los resultados y planificar la evaluación y reconsideración anual del programa. Esa labor la puede realizar por sí solo o en cooperación con los agentes de extensión.
El primer paso consiste en determinar si el programa es necesario. Con frecuencia, los promotores convencen a los piscicultores para que introduzcan unos cambios que en realidad no son necesarios. Un agente de ventas puede haberse sentido impresionado por la forma de un-pez que ha observado en otra provincia y cree que sería sumamente interesante que los piscicultores seleccionaran esa conformación corporal. Pero el programa no deberá iniciarse si no existe demanda de esa conformación modificada entre los consumidores locales. Aplicar de forma frivola un programa de cría es un despilfarro de dinero y de mano de obra y puede ser contraproducente. Antes de iniciarlo, el piscicultor debe de tener en cuenta las preferencias del consumidor y cuánto está dispuesto a pagar. Otro aspecto importante de la evaluación es establecer qué caracteres deben ser objeto de selección. El piscicultor puede preparar una lista de los caracteres que desearía mejorar y luego reducirla a unos cuantos caracteres convenientes desde el punto de vista del cultivo de los peces y de su comercialización. Algunos caracteres, como el peso corporal, deben figurar en cualquier programa de cría selectiva. Otros caracteres que pueden hacer de la piscicultura una actividad menos arriesgada o más fácil, como mayor resistencia a las enfermedades y mayor facilidad de captura, pueden figurar también en la lista inicial.
Algunos caracteres son totalmente innecesarios y deben eliminarse inmediatamente. Puede darse el caso de que una fábrica de elaboración de pescado esté interesada en que los piscicultores aumenten el rendimiento de la canal o disminuyan el contenido de grasa en las visceras con el fin de conseguir mayores beneficios. Esos caracteres sólo deben ser objeto de un programa de cría selectiva si los propietarios de la fábrica están dispuestos a pagar sumas mayores por los peces con un mayor rendimiento de la canal o con menos contenido graso.
Los piscicultores deben eliminar también los caracteres cuya mejora es más fácil de conseguir incidiendo en el sistema de manejo. Por ejemplo, no tiene mucho sentido utilizar la cría selectiva para aumentar la fecundidad porque es mucho más fácil y menos costoso aumentar la producción de huevos modificando el manejo del criadero. Además, hay que tener en cuenta que la mayor parte de los peces tienen una elevada fecundidad. Aumentar la producción de las hembras de 4.000 a 4.500 huevos por kg no supone un progreso sustancial y es innecesario. Para aumentar la producción de huevos basta con aparear a un mayor número de hembras o de mayor tamaño. Incrementar la producción de huevos mediante la cría selectiva es caro y para conseguirlo habrá de transcurrir al menos una generación, pero modificar el sistema de gestión es poco costoso y da resultados inmediatos.
Los piscicultores sólo deben mejorar los caracteres más importantes que permiten aumentar los beneficios, ya sea porque los peces crecen más rápida o eficientemente, porque tienen mayor supervivencia o porque los consumidores están dispuestos a pagar un precio más alto.
Los responsables de la gestión de los criaderos que trabajan en centros de producción de alevines pueden considerar la posibilidad de seleccionar caracteres capaces de aumentar la eficiencia del sector o abrir nuevas posibilidades a la piscicultura. Por ejemplo, si se consigue mayor tolerancia a la salinidad, se podrán cultivar peces en estanques de agua salobre en una superficie de varios miles de hectáreas y la selección para adelantar el período del desove puede permitir obtener en un solo período de crecimiento peces de tamaño comercial o de mayor tamaño, por los que pueden pedir un precio más elevado.
Un segundo aspecto de la evaluación consiste en decidir si el piscicultor va a producir alevines mejorados genéticamente para su propio uso o para una industria local de piscicultura. Esa decisión determina la cuantía de dinero que se debe invertir en el programa y el alcance del mismo. En efecto, en el primer caso sólo necesitará dedicar al proyecto una parte de su piscifactoría, mientras que si su intención es producir alevines mejorados genéticamente para la industria local deberá dedicar al proyecto todas sus instalaciones, o la mayor parte de ellas.
Una de las razones por las que debe reducirse al mínimo la lista de caracteres que serán objeto de mejora es que la eficiencia en la mejora de un carácter es inversamente proporcional al número de caracteres incluidos en el programa de cría selectiva. Por consiguiente, si uno ó dos caracteres revisten más importancia que los demás, la atención debe centrarse en esos caracteres. Si se incluyen caracteres que no son esenciales el ritmo de mejora de los caracteres importantes será más lento. Por ello, en los programas de cría selectiva sólo se debe intentar mejorar, por regla general, uno o dos caracteres. En algunos programas puede incluirse un tercer carácter y sólo se incorporará un mayor número de caracteres en los programas que lleven a cabo los genetistas en los centros de investigación.
Una vez que el piscicultor ha reducido la lista a uno o dos caracteres que desearía mejorar, debe determinar la forma de definirlos y medirlos. Se deben excluir los objetivos que no pueden definirse con precisión y los caracteres que son difíciles de medir. Por ejemplo, todos los piscicultores desean aumentar la resistencia a las enfermedades, pero cabe preguntarse qué se entiende por este concepto. Algunos dirán que consiste en que “los peces no enfermen”, mientras que para otros consistirá en que “los peces no mueran”. Sin duda, existe gran diferencia entre ambas definiciones. ¿Qué significa que “los peces no enfermen”? ¿Significa que los peces fueron infectados pero no mostraron signos clínicos de la enfermedad o que no resultaron infectados? Si los peces se cultivan en un estanque, ¿cómo se podrá establecer qué peces fueron infectados y cuáles no? Ni siquiera la expresión “que no mueran” es muy precisa, porque son muchas las variables que intervienen en la gestión sanitaria. Los peces pueden morir por causas distintas de la infección, en un brote de una enfermedad o durante el período de crecimiento. Dada la dificultad de medir este carácter, los piscicultores deben evitarlo en la medida de lo posible. Además, si la resistencia a una enfermedad determinada es heredable mejorará con el paso del tiempo, gracias a lo que se llama “selección por domesticación”. La domesticación es el proceso mediante el cual la piscifactoría y el sistema de gestión del piscicultor seleccionan a favor o en contra de los genes como consecuencia natural del cultivo. Así pues, la resistencia a la enfermedad no debe figurar en las listas de la mayoría de los piscicultores. Aunque se están registrando algunos avances en la resistencia a las enfermedades, estos programas de mejora son complicados y deben ser aplicados por científicos en los centros de investigación.
También deben evitarse los caracteres que son difíciles de medir debido al sistema de cultivo. Los piscicultores que administran a los peces alimentos granulados quieren mejorar la conversión del alimento, porque al mejorar este carácter los peces transformarán el alimento más eficientemente, con el consiguiente aumento del rendimiento y de los beneficios. Pero ¿cómo determinar la conversión del alimento en cada pez? Esto sólo se podría conseguir cultivando los peces en recipientes individuales, pero sería excesivamente costoso. El piscicultor podría recurrir a la selección familiar para mejorar este carácter si estuviera en condiciones de cultivar decenas de familias en los recipientes. Un programa de esa índole sería costoso y mejorar la conversión del alimento no es tan importante como aumentar la tasa de crecimiento. En caso de que sean necesarios, los programas de cría selectiva para mejorar la conversión del alimento deben aplicarlos los genetistas que trabajan en centros de investigación.
Incluso los caracteres que todos los piscicultores quieren mejorar y que deben figurar en un programa de cría selectiva deben ser cuidadosamente definidos y medidos. Todo piscicultor quiere mejorar el crecimiento, pero ¿cómo debe definirse y cómo debe medirse? Esto es importante, porque determinará el grado de progreso que se puede conseguir mediante la selección. Generalmente, la tasa de crecimiento se define como peso o como longitud a una edad determinada, de manera que lo primero que se ha de dilucidar es si el crecimiento debe definirse por el peso o por la longitud.
Es sencillo medir con precisión centenares de peces redondeando las cifras al milímetro más próximo y durante el proceso los peces sólo sufren un pequeño estrés. Sin embargo, la longitud incluye la medida de la cabeza y de la cola. Podría darse el caso de que se seleccionaran peces con una cola muy larga antes que los peces de mayor peso, pero los consumidores no compran el pescado por centímetros sino por kilogramos. Aunque el peso refleja mejor cómo se compran y se venden los peces, es difícil pesar con precisión centenares de peces. A menos que el piscicultor posea una balanza electrónica, el procedimiento puede ser muy tedioso y causar un gran estrés a los peces. Además, el peso registrado de cada pez incluye el agua que tiene en la boca, las heces y el alimento que contiene el tracto digestivo, así como el peso de la cabeza y de las visceras y, por tanto, también está sujeto a error.
Aunque lo mejor sería medir el crecimiento en función del peso, la correlación entre longitud y peso es muy estrecha (ambos factores están muy relacionados, pues el incremento del uno supone que también aumenta el otro), de manera que evaluar el crecimiento midiendo la longitud es un sistema adecuado de establecer el crecimiento midiendo el peso corporal. Si el piscicultor quiere reducir el índice de error que comporta evaluar el crecimiento por la longitud, puede tomar una longitud estándar en lugar de la longitud total, eliminando el efecto de la longitud de la cola en la selección.
En caso de que la medición de los peces la efectúe más de una persona, se habrá de establecer previamente un método estándar para la medición, a fin de que todos los encargados de ese cometido efectúen mediciones uniformes. En caso contrario, los valores fenotípicos atribuidos a los peces (longitud) estarán sujetos al error humano. Cuando una sola persona efectúa todas las mediciones, el único requisito es que utilice la misma técnica para todos los peces.
Cuando se mejora el peso mediante selección para longitud, se dice que se ha mejorado el peso por “selección indirecta”. Este fenómeno se debe a la correlación (genética) positiva entre los dos caracteres, debido a la cual cuando se mejora uno de los caracteres automáticamente se mejora también el otro. Por ello, muchas veces la selección para aumentar la tasa de crecimiento permite mejorar otros caracteres, como la supervivencia, la resistencia a las enfermedades y la conversión del alimento. Frecuentemente, los mejoradores aprovechan las correlaciones genéticas y utilizan la selección indirecta para mejorar caracteres cuya medición es difícil o costosa, seleccionando otros caracteres más fáciles de medir. Por ejemplo, se ha mejorado el peso de cosecha de la trucha arco iris mediante selección indirecta, al realizar la selección para peso de los alevines; también se están aplicando programas de cría selectiva para mejorar la resistencia a las enfermedades mediante selección indirecta, seleccionando para la respuesta al estrés o para la fortaleza de la membrana celular.
Otra pregunta a la que se debe dar respuesta antes de iniciar la mejora de la tasa de crecimiento es la siguiente: ¿A qué edad se evaluará el crecimiento? El crecimiento de los peces no es constante; si lo fuera, no tendría gran importancia a qué edad se efectuara la medición. Lo más juicioso es medirlos cuando se trasladan o cosechan, por ejemplo cuando se transfieren de los estanques de alevines a los estanques de crecimiento y cuando se cosechan para su envío al mercado. Generalmente, esto se define como medición a una edad determinada. Tras varias generaciones de selección, podría definirse como el tiempo necesario para alcanzar el tamaño comercial. Si los peces se miden y seleccionan a dos edades distintas en lugar de una sola, la tasa de crecimiento aumentará más rápidamente.
De importancia fundamental es saber definir exactamente el objetivo que se desea alcanzar y medir con precisión el carácter. Si el piscicultor no puede realizar estas tareas, no debe utilizar un programa de cría selectiva. Si lo hace con objetivos que no han sido bien definidos y mediciones imprecisas del carácter, el programa puede hacer disminuir la tasa de crecimiento e impedir futuras mejoras, al descartarse los alelos realmente valiosos.
Por ejemplo, la selección para tasa de crecimiento en algunas poblaciones de carpas indias originó peces que crecían más lentamente, porque no se había definido con precisión la tasa de crecimiento y no se habían medido bien los peces. Las carpas se cultivaron con el sistema de producción denominado “de lotes múltiples”. Cuando se aplica este sistema de manejo, cada estanque se explota muchas veces en un período de varios años para retirar continuamente los peces de tamaño comercial. El estanque se repuebla frecuentemente con alevines para sustituir a los peces que se han retirado, de manera que puedan cosecharse peces constantemente. Al cabo de varios años de producción continua, el estanque se ha agotado porque los peces de gran tamaño que han escapado repetidamente a las redes afectan negativamente a la producción.
Cuando se vaciaron estos estanques de carpas, los piscicultores y el personal del criadero conservaron los peces de mayor tamaño porque definieron la tasa de crecimiento como el tamaño corporal en el momento del vaciado y sólo la midieron cuando se hubo vaciado un estanque de producción de lotes múltiples. Además, conservaron esos peces porque podrían producir mayor número de huevos que los peces reproductores de menor tamaño.
Lamentablemente,'debido a la manera de definir la tasa de crecimiento y a la forma en que se llevó a cabo la medición del carácter (tamaño en el momento de vaciar los estanques) el programa de cría selectiva redujo la tasa de crecimiento, porque los peces seleccionados eran los de mayor edad y, además, de crecimiento lento. Eran los peces de mayor tamaño cuando se vaciaron los estanques simplemente porque escapaban a las redes. Así pues, el programa no sólo originó peces de crecimiento más lento sino que sirvió para seleccionar peces expertos en el arte de escapar.
Una vez que el piscicultor ha cuantificado sus objetivos y determinado cómo efectuará la medición de los caracteres, debe decidir qué programa utilizará. Existen dos tipos básicos de programas de cría selectiva: la selección individual (llamada también “selección masal”) y la selección familiar. En la selección individual, se ignoran las relaciones familiares y se establecen comparaciones entre individuos. Se clasifican los peces y se utiliza los mejores como peces reproductores seleccionados. En la selección familiar, son de la máxima importancia las relaciones familiares; se comparan y clasifican las medias familiares y se reservan o descartan familias enteras, o se establecen clasificaciones individuales dentro de cada familia y se efectúa la selección independientemente en cada una de ellas. Cuál sea el programa más eficiente dependerá del carácter y de cómo sea medido, así como de la biología de los peces y de la heredabilidad del carácter. En este capítulo se examinarán con más detenimiento la selección individual y familiar.
El piscicultor debe trazar el plan que seguirá para alcanzar sus objetivos. En él debe incluir diversos aspectos, como el número de peces que apareará, el número de peces por familia que cultivará, cuántos estanques poblará, la densidad de población y el sistema de cultivo.
En vista de que los programas de cría selectiva son costosos, es importante evaluar con precisión los efectos de la selección. El mejor procedimiento para hacerlo consiste en comparar la media del carácter en el transcurso del tiempo. Para hacerlo de forma adecuada, se debe compararla media de la población seleccionada con la de una población control que no ha sido objeto de selección. Hay que hacerlo así porque la media está determinada tanto por el genoma (que se está intentando mejorar mediante selección) como por el medio. En una sección posterior del presente capítulo se examinarán las técnicas para crear una población control y cómo se utiliza para evaluar los resultados de la selección.
Finalmente, el piscicultor debe revisar el programa de cría selectiva al menos una vez al año. Debe evaluar cómo se está aplicando y si es posible mejorarlo. También puede decidir interrumpirlo si no da resultados o si éstos no están en consonancia con el esfuerzo desplegado. Con frecuencia, los piscicultores se sienten frustrados y desean poner fin al programa de cría selectiva para comenzar otro, pero una decisión de este tipo es desaconsejable. Si un piscicultor aplica un nuevo programa cada año o cada dos años lo único que conseguirá será malgastar el dinero. Por otra parte, si el programa no da resultado sería estúpido persistir y gastar el dinero innecesariamente.
Cuando el piscicultor ha decidido mejorar un carácter cuantitativo mediante selección, debe elegir el programa de cría selectiva más eficiente. Existen dos categorías principales de programas de cría selectiva: la selección individual y la selección familiar. La selección individual es más sencilla y menos costosa y requiere un menor registro de datos.
En la selección individual se miden todos los individuos y la decisión de seleccionar o descartar un pez se basa únicamente en su valor fenotípico. Se compara cada uno de los peces con un valor denominado “valor límite” y se conservan los peces cuyo valor fenotípico es igual o mayor que dicho límite, descartándose aquellos cuyo valor fenotípico queda por debajo del límite. El valor límite es un valor fenotípico predeterminado que se expresa frecuentemente como “el mejor 10%”, “el mejor 5%”, etc. Los peces que se conservan pasan a ser la primera generación de peces reproductores seleccionados. En otra sección de este mismo capítulo se analiza el valor límite y la forma de determinarlo.
La primera generación de peces reproductores seleccionados producen, al aparearse, la primera generación de peces seleccionados, que recibe el nombre de “generación F,”. A su descendencia se le da el nombre de “generación F2”, y así sucesivamente. El piscicultor debe permitir que los peces reproductores seleccionados se apareen al azar o aparearlos de forma aleatoria, a menos que desee efectuar selección para tasa de crecimiento después de alcanzada el tamaño comercial o para caracteres relacionados con la madurez. Este proceso se repite en las generaciones subsiguientes (figura 19).
Las variables ambientales tiene una influencia significativa en la producción de caracteres cuantitativos. Incluso cuando la heredabilidad de un carácter es elevada, el componente ambiental de la variación fenotípica puede ser mayor que el componente heredable. Si la influencia ambiental sobre la variación fenotípica fuera idéntica en todos los peces, no tendría gran importancia durante la selección, a condición de que hubiera un grado suficiente de variación heredable como para que pudiera efectuarse la selección. Pero lo cierto es que muchas fuentes de variación ambiental influyen de forma distinta en los individuos o en las familias; cuando utiliza la selección, el piscicultor debe controlar esas variables ambientales, pues, de otro modo, supondrán una ventaja para determinados individuos o familias. Si tal cosa ocurre, es posible que los peces seleccionados estén en mejor situación debido a una ventaja ambiental pero no porque sean genéticamente superiores y la selección no dará los resultados esperados. Por ejemplo, los peces de mayor edad suelen ser también los de mayor tamaño porque su período de crecimiento ha sido más prolongado. Se ha demostrado que una diferencia de edad de sólo 24 horas puede suponer para los peces de más edad una ventaja con respecto al tamaño que no pierden nunca, lo que supone que la selección elegirá los peces de mayor edad y no los que tienen superioridad genética. Por consiguiente, si no se controlaran las diferencias de tamaño que guardan relación con la edad, el programa de cría selectiva pensado para aumentar la tasa de crecimiento explotaría la variación ambiental (edad) pero no la variación genética aditiva y no tendría éxito. Por ello, cuando sea posible se ha de efectuar el desove de todos los peces el mismo día. Si eso no es factible, se dividirá la población en cohortes de peces de la misma edad para que la selección tenga lugar independientemente en cada cohorte. Una vez realizada la selección, se pueden mezclar los peces reproductores seleccionados de cada cohorte.
Otros factores que deben controlarse son la edad y el tamaño de las hembras y las diferencias de manejo de los estanques de alevines o de crecimiento. Por ejemplo, si cada familia ocupa un estanque distinto, la selección individual no podrá distinguir qué peces han alcanzado el mayor tamaño por su superioridad genética y cuáles porque la densidad de población era menor y recibieron más alimento. Si el piscicultor no controla estas variables, puede ocurrir que todos los peces de mayor tamaño procedan de una misma familia y que sean los más grandes por efecto de la variación ambiental. En tal caso, la selección no producirá mejora alguna.
Incluso variables ambientales insignificantes como el tamaño de las partículas de alimento pueden ser causas importantes de variación fenotípica. Los estudios que se han realizado sobre la carpa común han puesto de manifiesto que si las partículas de alimento no son del tamaño adecuado, algunos individuos crecen mucho más deprisa que el resto de la población. La ventaja que presentan estos peces con respecto al tamaño no puede ser aprovechada en un programa de cría porque es una ventaja ambiental no heredable.
Al aplicar un programa de cría selectiva, el piscicultor debe controlar todos los factores ambientales que pueden reportar una ventaja a determinados individuos o familias. En caso contrario, la selección será menos eficaz que en otros programas similares o no tendrá efecto alguno. Teniendo en cuenta que los programas de cría selectiva requieren una inversión considerable de esfuerzo y de tiempo, la ausencia de control de estas variables tendrá un costo importante.
Para crear una población de peces reproductores seleccionados, el piscicultor debe decidir dónde situar el valor límite. Generalmente, se expresa en porcentaje, por ejemplo, el 10% mejor. Puede expresarse también como tamaño mínimo, como 500,0 g, pero este criterio puede ser arriesgado, pues es posible que en un año desfavorable ningún pez alcance el valor límite.
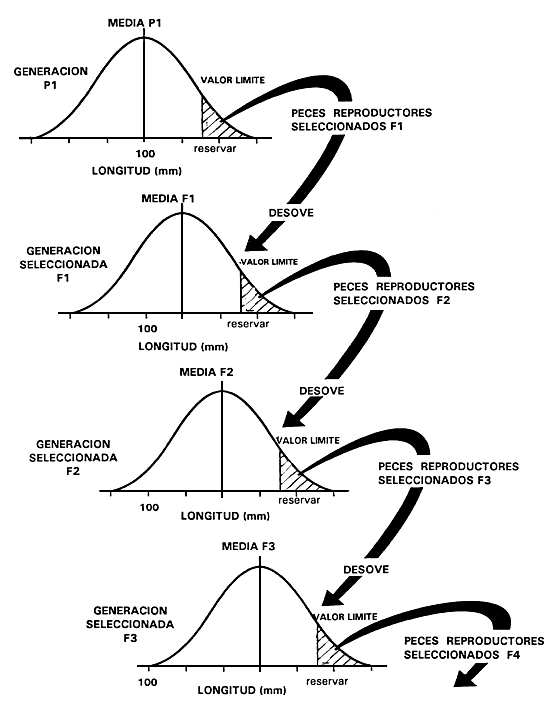
Figura 19. Diagrama esquemático de la selección individual para mejorar la tasa de crecimiento mediante selección para aumento de longitud. La figura muestra cuatro generaciones de selección e indica cómo se puede aumentar la longitud en el transcurso del tiempo.
Normalmente, el valor límite se establece inmediatamente antes de que tenga lugar la selección y se basa en la reserva de un porcentaje de la población determinado previamente. La forma más sencilla de establecer el valor límite y la que causa menos estrés a los peces consiste en medir una muestra al azar de 100–200 peces. Una vez medidos, se clasifican y el valor que corresponde al percentil deseado es el valor límite. Por ejemplo, si se reserva el 10% mejor, el valor límite es el percentil 90. Si se miden 200 peces, para obtener el valor límite basta con retroceder 20 mediciones a partir del valor fenotípico más alto una vez que han sido clasificados (Figura 20).
Dimorfismo sexual: Es necesario un último refinamiento del procedimiento de selección. El piscicultor debe decidir si la selección se efectuará en el conjunto de la población o separadamente para cada sexo. Esta es una decisión importante porque muchas especies presentan dimorfismo sexual, que se traduce en que uno de los dos sexos alcanza mayor tamaño o crece más rápidamente. Si la especie no presenta dimorfismo sexual o si la selección se ha de efectuar antes de que comience el dimorfismo, puede crearse un único valor límite para el conjunto de la población. En caso de que exista dimorfismo sexual, se establecerán valores límite distintos para cada sexo, pues de lo contrario la población seleccionada podría estar formada únicamente por el sexo que alcanza el mayor tamaño (Figura 21).
Muchas especies presentan dimorfismo sexual para el tamaño corporal. Todas las especies cultivadas importantes de tilapia presentan un acusado dimorfismo sexual. A los pocos meses de vida, los machos crecen mucho más deprisa que las hembras y al año de edad su tamaño puede duplicar el de estas últimas. Esta es la razón por la que los cultivadores de tilapia quieren cultivar poblaciones monosexuales de machos y no de hembras, en su intento de controlar la reproducción. También la carpa común y las carpas chinas presentan dimorfismo sexual para el tamaño corporal, siendo las hembras las que alcanzan mayor tamaño. Sin embargo, lá diferencia de tamaño en las carpas no aparece hasta que alcanzan la madurez, por lo cual la selección para tasa de crecimiento en las carpas en el período anterior a la madurez sólo exige un valor límite.
Si la selección se realiza independientemente en los dos sexos debido al dimorfismo sexual, el procedimiento que se indicó para establecer el valor límite (Figura 20) debe aplicarse por separado para los machos y las hembras. En caso de que la selección se realice para cada cohorte o para varias de ellas, el proceso ha de tener lugar en cada cohorte.
En general, los programas más eficientes y sencillos son aquellos en los que el objeto de la selección es la mejora de un solo carácter. Como ya se ha indicado, la eficiencia en la selección de cada carácter es inversamente proporcional al número de caracteres incluidos en el programa de cría selectiva.
Si un piscicultor desea mejorar más de un carácter, puede recurrir a la selección en tándem, al descarte independiente o al índice de selección. La selección en tándem es ineficaz y no se recomienda. El índice de selección es demasiado complicado para la mayoría de los piscicultores, pero pueden emplearlo los acuicultores de los centros de investigación y los directores de criaderos que trabajan en centros de producción de alevines. Los piscicultores que quieran seleccionar 2, o a lo sumo 3, caracteres deben utilizar el descarte independiente.
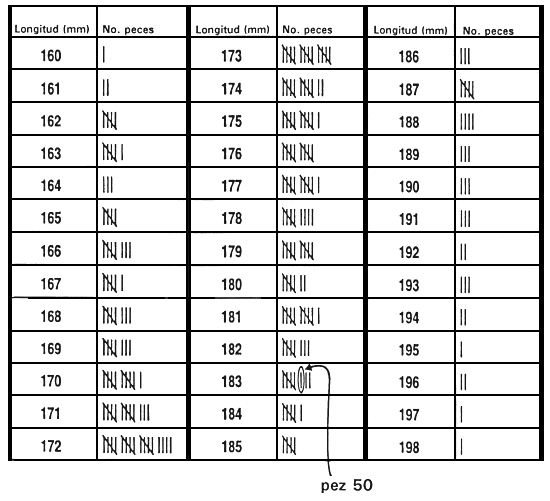
Figura 20. Una forma rápida y sencilla de establecer el valor límite. El objetivo de este programa de cría selectiva es aumentar la longitud a los 10 meses seleccionando el 20% mejor de la población. Para determinar la longitud que corresponde al percentil 80, se obtuvo una muestra de 250 peces de 10 meses de edad en un estanque en el que había 5.000 peces producidos el mismo día. Se midió cada uno de los peces de la muestra redondeando al milímetro más próximo y se registraron los valores fenotípicos en esta hoja de datos. Para establecer el valor límite se realizó una cuenta hacia atrás a partir del pez de mayor tamaño (198 mm),'para hallar el pez correspondiente al percentil 80. Dado que la meta es seleccionar el 20% mejor, la longitud del pez 50 será el valor límite (el pez 50 es el 20% de 250 peces). Como el pez 50 tiene un valor de 183 mm, el valor límite es 183 mm. Se conservarán los peces _<.183 mm, que serán los peces reproductores seleccionados, y se desecharán los peces < 183 mm.
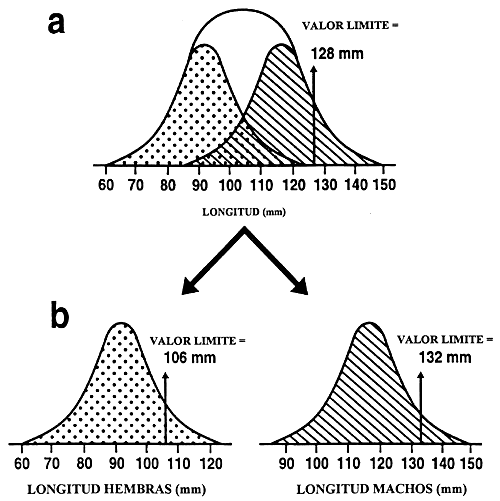
Figura 21. Efecto del dimorfismo sexual en la selección para el tamaño. El gráfico muestra la distribución de la longitud en una población de peces en la que los machos alcanzan mayor tamaño que las hembras: el gráfico rayado ilustra la distribución de la longitud de los machos en la población y el gráfico de puntos la que presentan las hembras. En el gráfico a, se utilizó un único valor límite (128 mm) en un programa de cría selectiva para mejorar la longitud. A causa del dimorfismo sexual, todos los peces reproductores seleccionados eran machos. Debido a ello, el piscicultor no podrá producir descendencia alguna u obtendrá y apareará hembras no seleccionadas. El gráfico b muestra cómo debe realizarse la selección en especies que presentan dimorfismo sexual. Se deben establecer valores límite distintos para cada sexo y la selección debe realizarse por separado para los machos y las hembras. El gráfico b indica que el piscicultor seleccionó los machos mejores estableciendo un valor límite de 132 mm, y seleccionó las mejores hembras estableciendo un valor límite de 106 mm.
Selección en tándem: En la selección en tándem, se selecciona primero un carácter durante varias generaciones, luego se interrumpe la selección de dicho carácter y se inicia un nuevo programa de cría para seleccionar un segundo carácter. Se trata, en esencia, de un programa de cría selectiva en dos fases, concebido para mejorar primero un carácter, para luego interrumpir dicho programa y comenzar un segundo programa de cría selectiva con el fin de mejorar otro carácter. Si la mejora del primer carácter se considera importante y el programa funciona correctamente sería poco juicioso interrumpirlo. Por esa razón y porque la mejora de dos caracteres lleva un tiempo muy largo con este procedimiento, el piscicultor no debe utilizar la selección en tándem para mejorar dos o más caracteres. Es sumamente ineficaz.
Descarte independiente: Es, tal vez, el programa de cría selectiva más sencillo para mejorar dos o tres caracteres. Es un programa de cría en el que se seleccionan simultáneamente dos o más caracteres estableciendo valores límite para cada carácter y sólo se conservan aquellos peces que alcanzan o superan los valores límite. Se descartan los peces que no alcanzan alguno de los valores límite. La Figura 22 ilustra este tipo de programa de cría.
Consideremos, por ejemplo, el caso de un piscicultor que desea aumentar la tasa de crecimiento mediante selección para longitud, y la conformación corporal mediante selección para profundidad corporal en la primera espina de la aleta dorsal. Para establecer los valores límite se utiliza el procedimiento indicado anteriormente. El piscicultor establece unos valores fenotípicos previamente determinados o selecciona el 10% mejor, el 20% mejor, etc. Si establece percentiles predeterminados, determina los valores límite que corresponden a dichos percentiles midiendo una muestra al azar de 100–200 peces y hallando dichos valores, tal como se ilustró en la Figura 20. En este ejemplo, el piscicultor ha establecido valores límite de 243 mm para la longitud y 133 mm para la profundidad corporal en la primera espina de la aleta dorsal. Una vez que ha fijado los valores límite, mide cada uno de los peces y sólo conserva los que alcanzan o superan dichos valores. En este programa de cría, la decisión de conservar o descartar seis peces fue la siguiente:
| Pez | Longitud (mm) | Profundidad (mm) | Conservar / descartar | |
| 1 | 254 | 129 | descartar | |
| 2 | 243 | 130 | conservar | |
| 3 | 241 | 138 | descartar | |
| 4 | 240 | 120 | descartar | |
| 5 | 280 | 129 | descartar | |
| 6 | 250 | 131 | conservar |
Aunque el descarte es más eficaz y más fácil de aplicar que el sistema de tándem, este programa de cría tiene dos inconvenientes. En primer lugar, cuando se seleccionan dos caracteres de forma simultánea e independiente, el número de peces seleccionados puede ser muy reducido. Por ejemplo, si se selecciona para dos caracteres y el valor límite es el 10% mejor para ambos caracteres, sólo se conservará el 1 % de la población (0,1 × 0,1 = 0,01). Si se añade un tercer carácter, situándose en el mismo nivel el valor límite, sólo se conservará un pez de cada mil. En la Figura 22 puede apreciarse el carácter fuertemente restrictivo de este tipo de selección.
El reducido número de peces que cumplen o superan todos los valores límite puede ocasionar dos problemas. Por un lado, es posible que no pueda producirse un número suficiente de alevines seleccionados para que alcancen el estado adulto en la temporada siguiente. Por otro, es posible que, al cabo de una o dos generaciones, se registre un elevado nivel de consanguinidad, lo que podría suponer que se perdiera gran parte de la mejora obtenida mediante la selección.
El problema planteado por el establecimiento de valores límite independientes puede obviarse decidiendo previamente hasta qué punto será estricta la selección, esto es, qué porcentaje de peces serán seleccionados. Una vez que se ha determinado este extremo, se vuelven a calcular los porcentajes límite necesarios para producir ese valor. Por ejemplo, si se desea conservar el 15% de la población y seleccionar dos caracteres que revisten la misma importancia, hay que hallar la raíz cuadrada de 0,15, que indicará que es necesario seleccionar el 39% mejor para ambos caracteres (0,30 × 0,30 = 0,15). Si se desea hacer más hincapié en un carácter, se fija un valor límite más elevado (en percentiles) para dicho carácter y se reduce el valor límite para el segundo carácter. Por ejemplo, si se desea conservar el 10% de la población, los valores límite pueden ser los percentiles 25 y 40 para dos caracteres (0,4 × 0,5 = 0,1).
Una segunda consecuencia negativa del descarte independiente es que se han de descartar peces que destacan para un carácter. Así, en el ejemplo anterior, el pez № 5 se descartó pese a que superaba ampliamente el valor límite para la longitud. Para solventar este problema se utiliza el descarte independiente modificado, en el cual se conservan los peces que alcanzan o superan ambos valores límite y aquellos que superan uno de los valores límite pero no alcanzan por un margen escaso el valor límite del segundo carácter (Figura 23).
Por la facilidad con que pueden realizarse y porque no exigen registrar un gran volumen de datos, el descarte independiente o el descarte independiente modificado son los programas de cría selectiva que deben aplicar los piscicultores que quieren mejorar simultáneamente dos o tres caracteres.
Indice de selección: El índice de selección es el programa de cría selectiva más eficiente para mejorar dos o más caracteres. Lamentablemente, es también más complejo y más costoso que el descarte independiente. Este tipo de programa de cría selectiva sólo deben utilizarlo acuicultores que trabajan en centros de investigación o administradores de criaderos que trabajan en centros de producción de alevines. Para mejorar dos caracteres, los piscicultores y la mayor parte de los acuicultores deben recurrir al descarte independiente. De hecho, el índice de selección sólo es necesario cuando se quieren mejorar tres o más caracteres simultáneamente.
Un índice de selección es un programa de cría selectiva en el que se evalúan simultáneamente dos o más caracteres y se asigna a cada pez un valor numérico global. A continuación, se ordenan los peces de acuerdo con dichos valores numéricos. La clasificación se establece de manera que un pez que se encuentra en el percentil 50 para cada uno de los caracteres obtiene un valor I de 100,0. Por consiguiente, se considera que los peces que tienen valores I > 100 están por encima de la media y los que poseen valores I < 100 están por debajo de la media. El objetivo último de todo programa de cría selectiva complejo a gran escala es construir un índice de selección.
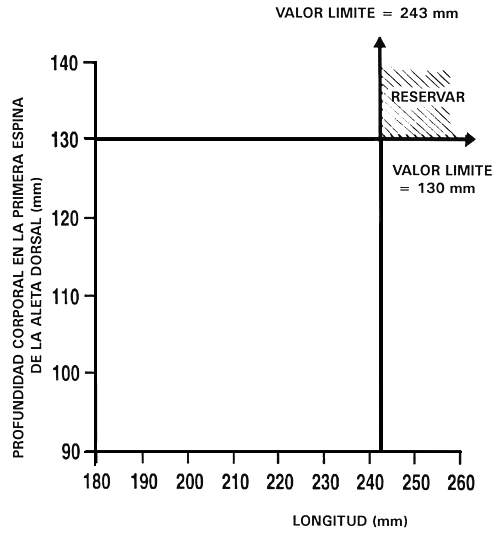
Figura 22. Diagrama esquemático del descarte independiente. En este programa de cría selectiva, un piscicultor está seleccionando para aumento de la tasa de crecimiento (mediante selección para longitud) e intenta mejorar también la conformación corporal (seleccionando para profundidad corporal en la primera espina de la aleta dorsal). Ha establecido valores límite independientes de 243 mm para longitud y 133 mm para profundidad corporal en la primera espina de la aleta dorsal. Sólo los peces que alcanzan o superan los dos valores límite serán reservados y pasarán a ser los peces reproductores seleccionados, descartándose todos los demás peces. El gráfico ilustra los porcentajes relativos de peces que se reservarán (parte rayada) y descartarán (parte no rayada).
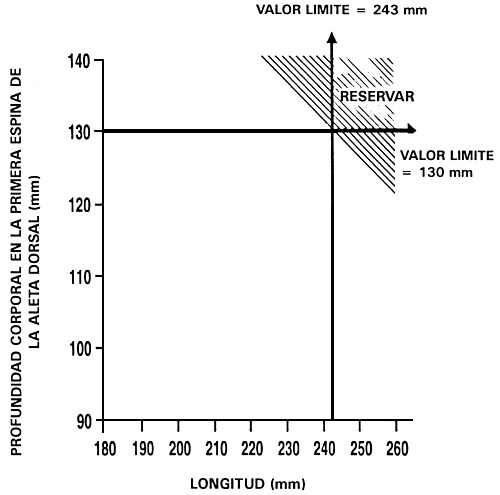
Figura 23. Diagrama esquemático del descarte independiente modificado. En este programa de cría selectiva, un piscicultor está efectuando la selección para aumento de la tasa de crecimiento (mediante selección para longitud) y trata de mejorar la conformación corporal (mediante selección de la profundidad corporal en la primera espina de la aleta dorsal). Al igual que en el programa de cría ilustrado en la Figura 22, el piscicultor ha establecido valores límite independientes de 243 mm para la longitud y 130 mm para la profundidad corporal en la primera espina de la aleta dorsal. En este caso, el piscicultor no sólo seleccionará peces que alcancen o superen ambos valores límite, sino también aquéllos que superen uno de los valores límite y no alcancen por escaso margen el valor límite establecido para el otro carácter (parte rayada del gráfico). Se descartan todos los demás peces. El descarte independiente modificado permite seleccionar un mayor número de peces y no descartar individuos con un valor fenotípico realmente notable para un carácter.
Para elaborar un índice de selección es necesario conocer las heredabilidades, las correlaciones genéticas entre los caracteres y la importancia económica de los mismos; lamentablemente, no se dispone de esta información para la mayor parte de las poblaciones de peces cultivados para la alimentación. Se puede soslayar este problema modificando el índice de selección y creando lo que se llaman “factores de importancia” para cada carácter.
En el Cuadro 11 se esbozan las técnicas que se utilizan para crear un índice de selección mediante factores de importancia. El índice del Cuadro 11 se ha concebido para mejorar tres caracteres: longitud a los 12 meses, profundidad corporal en la primera espina de la aleta dorsal y rendimiento de la canal (expresado por la relación longitud del tronco [longitud desde la parte posterior de la cabeza hasta el extremo del pedúnculo caudall/longitud total). Una vez que se ha computado un valor I para cada pez, se ordenan los peces en función de los valores y se seleccionan el 10% mejor, el 15% mejor o el 20% mejor, descartándose los demás. En la Figura 24 se describe un programa de cría selectiva que utiliza el índice de selección obtenido en el Cuadro 11.
Este tipo de programa de cría selectiva exige un trabajo más intenso que el descarte independiente. En este último, el piscicultor asigna un valor límite a cada carácter (normalmente sobre la base de una muestra de 100–200 peces) que le permitirá reservar o descartar con rapidez cada uno de los peces. En un índice de selección, se han de medir todos los caracteres para cada individuo, marcar todos los peces y depositarlos en un estanque hasta que se haya completado la medición. Entonces, se identifican de nuevo los peces y se reservan o descartan según sus valores I. Este proceso, además de ser intensivo en mano de obra, ocasiona estrés a los peces por efecto de la manipulación. Por todas estas razones, el piscicultor que desea mejorar dos o tres caracteres debe utilizar el descarte independiente o el descarte independiente modificado.
La diferencia entre la selección familiar y la selección individual es que en aquélla la decisión de conservar o descartar los peces se adopta a nivel familiar y los valores fenotípicos individuales sólo son importantes en relación con la media familiar. La selección familiar suele utilizarse cuando la selección individual es ineficaz a causa de la baja heredabilidad del carácter (generalmente <0,15). Cuando la heredabilidad es reducida, el componente heredable de la variación fenotípica también es reducido, lo que supone que la mayor parte de las diferencias medibles entre los individuos se deben a fuentes de variación no heredables. Al realizar la selección a nivel familiar, se puede excluir una parte sustancial de la variación ambiental, lo que facilita la tarea de identificar las diferencias genéticas y seleccionar los peces mejores en términos de la variación heredable.
La selección familiar se utiliza también cuando las causas ambientales de la variación no pueden ser controladas, lo que puede dificultar o imposibilitar la mejora mediante selección individual. Por ejemplo, cuando es imposible sincronizar el desove, y éste se produce en un período de varias semanas o de varios meses, se suele recurrir a la selección familiar. Este tipo de selección permite controlar las diferencias de tamaño relacionadas con la edad.
Cuadro 11. Técnicas que pueden utilizarse para crear un índice de selección utilizando factores de importancia.
| 1. | Determinar los caracteres que figurarán en el índice de selección. En este ejemplo, se crearáun índice de selección para mejorar tres caracteres: | |
1) longitud a los 12 meses (mm);, | ||
2) profundidad corporal en la primera espina de la aleta dorsal (mm); | ||
3) rendimiento de la canal (indicada por la relación entre la longitud del tronco [longitud desde la parte posterior de la cabeza hasta el extremo del pedúnculocaudall/longitud total). | ||
| 2. | Determinar los factores de importancia para los caracteres. Para ello se utiliza la siguiente fórmula: | |
| Factor de importancia (I) = importancia relativa de la media población/carácter | ||
| 2a. | Determinar la importancia relativa de los caracteres, para lo cual se asigna un valor arbitrario a cada uno de ellos. Estos valores van desde el 1 al 99% (y totalizan el 100%) y representan la parte de la presión de selección que se asignará a un carácter (si se le asigna un valor del 0% no se intentará mejorar y, por lo tanto, no se incluirá en el programa; en cambio, si se asigna a un carácter un valor del 100%, ello quiere decir que es el único que se pretende mejorar, de manera que todos los demás caracteres quedan excluidos automáticamente del programa de cría). | |
| Se asignan los siguientes valores de importancia relativa: | ||
| Carácter | Importancia relativa | |
| Longitud | 60% | |
| Profundidad corporal | 30% | |
| Rendimiento de la canal | 10% | |
| 2b. | Determinar la media para cada carácter. | |
| El piscicultor mide una muestra al azar de 200 peces y determina las siguientes medidas: | ||
| Carácter | Media | |
| Longitud | 195 mm | |
| Profundidad corporal | 100 mm | |
| Rendimiento de la canal | 55% | |
| 2c. | Calcular el factor de importancia para cada carácter utilizando la fórmula indicada en el apartado 2. El factor de importancia para cada carácter es el siguiente: | |
| l Longitud (IL) = 60%/195 mm = 0.3076923 | ||
| l Profundidad corporal (IB) = 30%/100 mm = 0,3 | ||
| I Rendlmlento de la canal (ID) = 10%/55% = 0,1818182 | ||
| 3. | Crear el índice de selección que se utilizará para determinar el valor reproductivo (l )para cada pez. El valor de cada pez se establece incluyendo sus valores fenotípicos y los factores de importancia en la siguiente fórmula: | |
| I pez = (Il) (longitud) + (IB) (profundidad corporal) + ID) (rendimiento de la canal) | ||
| 4. | Determinar el valor reproductivo de un pez con los siguientes valores fenotípicos: longitud = 201 mm; profundidad = 119 mm; rendimiento de la canal = 54%: | |
| I = (0,3076924) (201 mm) + (0,3) (119mm) + (0,1818182) (54%) I = 107,364 | ||
La selección familiar es el único tipo de programa de cría que puede utilizarse cuando es necesario matar a un animal para determinar su fenotipo. Por ejemplo, si se pretende mejorar el rendimiento de la canal o reducir el contenido de grasa en las visceras es necesario dar muerte al pez para efectuar una medición precisa. Una vez muerto, el pez no se puede aparear (salvo que se puedan preservar los gametos mediante crioconservación). Así pues, para mejorar esos caracteres se debe recurrir a la selección familiar.
Existen dos formas básicas de selección familiar: selección intrafamiliar y selección interfamiliar. En esta última, se determinan los valores medios para cada familia y a continuación se clasifican dichos valores. Se descartan o reservan, entonces, familias enteras (Figura 25). En este tipo de selección no se establece un valor límite, sino que el cultivador decide conservar las mejores 10, 15 o 20 familias.
No es necesario seleccionar a toda la familia. Si ésta es amplia, se puede reservar una muestra igual, al azar, de cada familia. Para que se pueda hablar de selección interfamiliar, la muestra ha de ser aleatoria. El piscicultor no puede elegir los mejores peces de cada una de las familias seleccionadas. Aun cuando se conserven familias enteras, el tamaño de las familias seleccionadas ha de ser uniforme, pues de lo contrario una de ellas contribuirá en mayor medida a formar la población reproductora seleccionada.
Como en el caso de la selección individual, el dimorfismo sexual puede complicar la selección para aumentar la tasa de crecimiento. Cuando una especie presenta dimorfismo sexual, es necesario pesar o medir por separado los machos y hembras de cada familia. La selección puede realizarse, entonces, de dos formas. Una vez que se conocen los valores medios para cada sexo, puede hallarse la media de los dos valores medios (en cada familia, será idéntica la aportación de los machos y las hembras a la siguiente generación) y se utilizará este valor medio global para reservar o descartar las familias. El segundo procedimiento consiste en realizar la selección por separado para cada sexo. En este caso, existe la probabilidad de que en algunas familias se descarten los machos y se seleccionen las hembras y viceversa.
Para llevar a cabo la selección familiar, el piscicultor debe identificar los peces por familias, para lo cual se pueden seguir dos métodos distintos. En el primero, se cultiva cada familia en tanques, jaulas o estanques separados hasta que alcanzan un tamaño que permite aplicar marcas o etiquetas familiares. Una vez marcados, los peces pueden ser cultivados comunalmente (se mezclan y se cultivan juntos) y en el momento de la cosecha se separan por familias para efectuar la selección. El segundo método consiste en cultivar los peces por separado desde el momento en que se produce la fecundación de los huevos hasta que se realiza la selección. Incluso después de ésta, si no es posible marcar los peces, se les debe mantener en unidades distintas. En caso de que puedan ser marcados, los peces reproductores seleccionados pueden ser cultivados comunalmente hasta el momento del apareamiento.
La selección interfamiliar es más costosa que la selección individual porque exige más instalaciones y mano de obra. También hace necesario consignar más datos. El coste del programa será mucho menor si los peces pueden ser marcados y cultivados comunalmente que si se cultiva cada familia por separado.
Uno de los aspectos interesantes de la selección interfamiliar es que permite reducir la mano de obra necesaria para aumentar la tasa de crecimiento. Además, el piscicultor puede mejorar fácilmente la tasa de crecimiento mediante la selección para peso. Dado que la selección interfamiliar se basa en las medias familiares, no es necesario determinar valores individuales si se selecciona para aumento de peso. Se pueden pesar conjuntamente los peces de cada familia y obtener la media familiar dividiendo el valor obtenido entre el número de peces que se ha pesado.
Si cada familia se cultiva en un estanque de crecimiento distinto, es necesario replicar cada unidad al menos tres veces y determinar luego la media global de las tres unidades antes de seleccionar o descartar las familias. Si se cultiva cada familia en un estanque, no es posible determinar si el valor fenotípico medio de la familia representa el valor reproductor de dicha familia o la calidad del estanque. Todo aquel que ha cultivado peces sabe que cada estanque tiene unas características peculiares. Algunos son “buenos”, mientras que otros son “malos”. En algunos estanques el agua es de buena calidad, hay una buena floración de algas y se obtienen elevados rendimientos, mientras que en otros existen problemas respecto al oxígeno disuelto, la floración de algas es escasa y los rendimientos son bajos. Si se utiliza un estanque para cada familia, las diferencias entre las medias familiares no se deben a la variación heredable de las familias sino a las poblaciones de algas existentes en los estanques o a otros factores ambientales. La selección no dará resultado si son las distintas condiciones de los estanques la razón principal de la diferencia existente entre las medias familiares.
Los piscicultores deben poner buen cuidado en que las condiciones de cultivo sean iguales para todas las familias. Uno de los factores ambientales que pueden y deben controlar en la selección ¡nterfqmiliar es la densidad de población, que ha de ser idéntica en cada familia, pues de lo contrario las diferencias entre las medias familiares no se deberán a diferencias heredables sino a la diferente densidad de población.
Teniendo en cuenta que la selección se basa en medias familiares y que se reservan o descartan familias enteras, cabe la posibilidad de que en la selección se descarten algunos de los peces con los mejores caracteres. Por ejemplo, en la Figura 25 se descartó la familia K porque presentaba la media familiar más baja. El descarte de la familia completa supuso excluir a los peces de mayor tamaño de la población junto con sus hermanos. A la inversa, en este tipo de selección se incorporan individuos enanos a la población de peces reproductores seleccionados. Así, en la Figura 25 se seleccionó a la familia G porque su media familiar era la segunda más alta. Al seleccionarse a toda la familia, los individuos de menor tamaño de la población quedaron incluidos en el conjunto de peces reproductores seleccionados.
Esta forma de selección puede ser muy eficaz, aunque pueda parecer poco juicioso eliminar algunos peces que se habrían conservado e incluir otros que se habrían descartado en el caso de la selección individual. Al tomar la media familiar como base de la selección se neutralizan algunas de las causas ambientales de la variación fenotípica. Puede argüirse que los peces de gran tamaño de la familia K lo eran porque una determinada ventaja ambiental les permitió recibir más alimento del que les correspondía. Aunque se pueda sentir la tentación de quebrantar las normas de la selección interfamiliar para conservar algunos de los peces descartados y descartar algunos de los peces reproductores seleccionados, no se debe actuar así. Ningún sistema de selección es perfecto y todos son susceptibles de causar alguna frustración, pero tomar como base (al menos en parte) del programa de cría selectiva la preferencia personal en lugar de la genética sólo servirá para menoscabar su eficacia.
1. Determinar los valores fenotípicos de cada uno de los peces que forman la población. En este ejemplo, sólo se establecen dichos valores para 10 peces:
| Pez | Longitud (mm) | Profundidad corporal (mm) | Rendimiento de la canal |
| T-l | 198 | 103 | 54 |
| T-2 | 210 | 110 | 55 |
| T-3 | 180 | 108 | 57 |
| T-4 | 195 | 100 | 55 |
| T-5 | 197 | 101 | 56 |
| T-6 | 190 | 98 | 54 |
| T-7 | 188 | 97 | 53 |
| T-8 | 195 | 102 | 57 |
| T-9 | 185 | 95 | 52 |
| T-10 | 199 | 102 | 55 |
2. Determinar el valor I de cada pez utilizando los valores de importancia abtenidos en el cuadoll:
| IT-I | = | (0.3076923)(198) | + | (0.3)(103) | + | (0.1818182)(54) | = | 101.641 |
| IT-2 | = | (0.3076923)(210) | + | (0.3)(110) | + | (0.1818182)(55) | = | 105.777 |
| IT-3 | = | (0.3076923)(180) | + | (0.3)(108) | + | (0.l818182)(57) | = | 98.148 |
| IT-4 | = | (0.3076923)(195) | + | (0.3)(100) | + | (0.1818182)(55) | = | 100.000 |
| IT-5 | = | (0.3076923)(197) | + | (0.3)(101) | + | (0.1818182)(56) | = | 101.097 |
| IT-6 | = | (0.3076923)(190) | + | (0.3)(98) | + | (0.1818182)(54) | = | 97.680 |
| IT-7 | = | (0.3076923)(188) | + | (0.3)(97) | + | (0.1818182)(53) | = | 96.583 |
| IT-8 | = | (0.3076923)(195) | + | (0.3)(102) | + | (0.1818182)(57) | = | 100.964 |
| IT-9 | = | (0.3076923)(185) | + | (0.3)(95) | + | (0.1818182)(52) | = | 94.878 |
| IT-10 | = | (0.3076923)(199) | + | (0.3)(102) | + | (0.1818182)(55) | = | 101.831 |
3. Ordenar los peces según los valores L Seleccionar o descartar los peces basándose en esos valores. En este ejemplo, se reserva el mejor 20 % de la población:
| T-2 | = | 105.777 | → | RESERVAR | ||
| T-10 | = | 101.831 | ||||
| DESCARTAR | ← | T-l | = | 101.641 | ||
| T-5 | = | 101.097 | ||||
| T-8 | = | 100.964 | ||||
| T-4 | = | 100.000 | ||||
| T-3 | = | 98.148 | ||||
| T-6 | = | 97.680 | ||||
| T-7 | = | 96.583 | ||||
| T-9 | = | 94.878 | ||||
Figura 24. Protocolo esquemático necesario para elaborar un índice de selección. Como medida preliminar se han de determinar los factores de importancia a partir de las medias fenotfpicas y de la importancia relativa de cada carácter. En el Cuadro 11 se describen los métodos que se utilizan para calcular los factores de importancia para los tres caracteres. Para determinar el valor I de un pez, el primer paso es establecer los valores fenotípicos de cada pez en la población. En este caso, sólo se presentan 10 peces. El paso 2 consiste en calcular el valor I de cada pez. En el paso 3 se ordenan los peces en función de sus valores I y se seleccionan o descartan sobre la base de dichos valores. En este ejemplo, se conservó el 20% mejor de la población, de manera que se conservaron T-2 y T-10 porque eran los dos peces mejores; el resto se descartó. Obsérvese que el pez T-4, cuyos valores fenotípicos eran iguales a los de la media de la población, tenía un valor I de 100,0; por definición, el valor I de dicho pez debía ser 100,0.
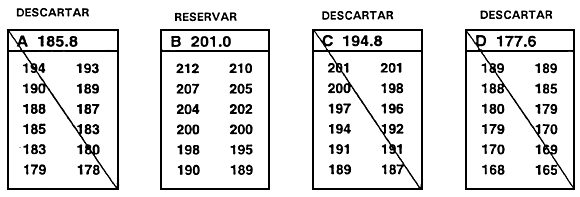
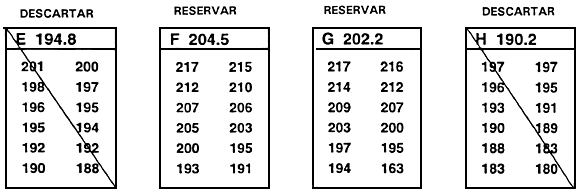
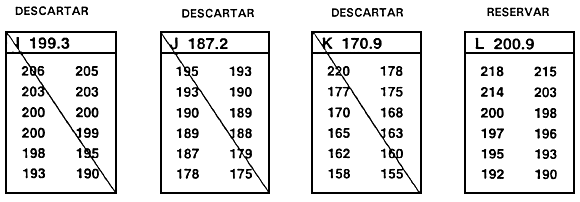
Figura 25. Diagrama esquemático de la selección interfamiliar. Se comparan las medias familiares y la selección se lleva a cabo a nivel familiar. Se reservan o descartan familias enteras (o muestras iguales y al azar). Esto se hizo seleccionando el 33% mejor de las familias (en este ejemplo sólo figuraban 12 familias). Para ello, se ordenaron las medias familiares y se reservaron las familias con las cuatro longitudes medias mayores (familias B, F, G y L). Se descartaron todos los peces de las ocho familias restantes. En este ejemplo, se midieron 12 peces procedentes de cada familia, redondeando al milímetro más próximo (los valores figuran debajo de la letra que identifica a la familia). En realidad, deberían medirse de 30 a 100 peces. El valor que figura junto a la letra que identifica a la familia es la media familiar. En este ejemplo, se parte del supuesto de que no existe dimorfismo sexual. En caso de que lo haya, es necesario establecer la media de ambos sexos en cada familia. A continuación, se puede efectuar la selección, basándose en los valores medios de los dos sexos o en los valores independientes de los machos y las hembras.
La selección intrafamiliar es otro tipo de selección familiar. En ella, se considera a cada familia como una subpoblación provisional y la selección se realiza independientemente dentro de cada familia (Figura 26). Cuando se miden los peces para decidir cuáles se han de reservar y cuáles deben ser descartados, se ordenan los peces de cada familia y se reservan los mejores de ellos. En esta forma de selección, el piscicultor conserva normalmente los mejores 10, 15 ó 20 peces de cada 15, 20 ó 30 familias. En caso de dimorfismo sexual, la selección se efectúa independientemente para cada sexo y el piscicultor conserva los 10, 15 o 20 machos mejores y las 10, 15 o 20 hembras mejores de cada familia.
En la Figura 26 se reservaron los cuatro mejores peces de cada familia. En teoría, se reservan o descartan los peces de cada familia basándose en la media familiar, pero para la selección intrafamiliar no es necesario calcular la media familiar. Al medir y ordenar todos los peces de cada familia (o todos los peces de una muestra al azar de 30–100 peces de cada familia), simplemente se reserva un número preestablecido.
Al efectuarse la selección independientemente en cada familia, es posible que los peces que se conservan de una familia sean de menor tamaño que muchos de los descartados de otra familia. Por ejemplo, en la Figura 26, tres de los peces de la familia K que se reservaron son de menor tamaño que muchos de los peces desechados de otras familias. Por otro lado, la figura 26 muestra que en una familia pueden ser descartados peces con cualidades notables si no alcanzan el valor límite establecido para dicha familia (número de peces que se deben conservar).
Como ocurría en la selección interfamiliar, descartar peces de gran tamaño e incluir peces más pequeños en la población de peces reproductores seleccionados puede resultar frustrante, pero el piscicultor debe resistir la tentación de reservar o descartar algunos peces atendiendo a su criterio personal. En la selección intrafamiliar es inevitable que algunos de los peces seleccionados de determinadas familias sean de menor tamaño que muchos de los peces descartados de otras. Es lógico que así sea porque normalmente se recurre a la selección intrafamiliar cuando existe una causa incontrolable importante de variación ambiental, como el momento del desove o la edad de la hembra, cuyos efectos se dejan sentir a nivel familiar pero no a nivel individual. Al subdividir la población en un amplio número de familias y efectuar la selección dentro de cada una de ellas se neutralizan esos efectos ambientales (que son los mismos para todos los peces de la familia) y de esta forma no influyen en la expresión fenotípica y en la selección.
Las investigaciones realizadas con tilapias han puesto en evidencia que la mayor parte de las veces la selección individual para mejorar la tasa de crecimiento no ha resultado eficaz por diversas razones, una de ellas la imposibilidad de sincronizar el desove en la tilapia. De la investigación se desprende que al tener lugar el desove a lo largo de un período de varios meses, la selección intrafamiliar es el programa de cría selectiva que debe utilizarse para aumentar la tasa de crecimiento, ya que permite neutralizar la influencia que puede tener sobre el tamaño el momento del desove.
Como en la selección interfamiliar, el piscicultor debe marcar las familias o cultivarlas en unidades individuales, pero en la selección intrafamiliar no es necesario cultivar las familias en unidades idénticas, porque sólo se comparan los peces de una misma familia (peces cultivados en la misma unidad). Al no efectuarse comparaciones entre familias, las diferencias ambientales entre los estanques no influirán en el proceso de selección. Así pues, en la selección intrafamiliar cada familia se puede cultivar en un solo estanque y, por tanto, el cultivo es mucho menos costoso que en la selección interfamiliar.
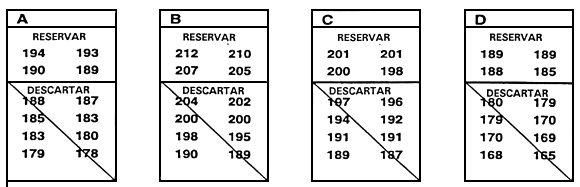
Figura 26. Diagrama esquemático de la selección intrafamiliar. En este tipo de selección se parte del supuesto de que las distintas familias son, provisionalmente, una subpoblación, y la selección tiene lugar independientemente dentro de cada una de ellas. En este ejemplo, el objetivo era aumentar la tasa de crecimiento mediante selección para longitud. Para ello se seleccionaron los cuatro peces mejores de cada familia, descartándose el resto. En este caso, se midieron 12 peces de cada familia. En realidad, deberían medirse de 30 a 100 peces, eligiendo los mejores de dicha muestra. Bajo la letra que identifica a la familia se indica la longitud individual (en milímetros). En este ejemplo se asume que no existe dimorfismo sexual. En caso de que exista, la selección se efectúa independientemente en machos y hembras y, en este ejemplo, se reservarían los dos mejores machos y las dos mejores hembras de cada familia.
La selección intrafamiliar puede realizarse en un único estanque. Si es posible marcar los peces, se pueden cultivar comunalmente después de que se les ha aplicado la marca familiar. En el momento de la cosecha se deben separar los peces de cada familia y puede realizarse la selección tal como se ha descrito. Si es posible disponer gran número de jaulas en un estanque, se puede cultivar cada familia en una jaula. El único requisito es que la tasa de crecimiento en las jaulas sea similar a la de los estanques.
Para superar algunos de los problemas que suscitan la selección interfamiliar y la intrafamiliar, es posible combinar ambas en un programa de cría integrado de dos fases (Figura 27). En este programa, se utiliza primero la selección interfamiliar para seleccionar las mejores familias y, a continuación, mediante la selección intrafamiliar se eligen los mejores peces de cada familia seleccionada. Si se utiliza esta combinación, el cultivo de los peces debe realizarse en las mismas condiciones que en la selección interfamiliar, porque la selección se efectúa primero a nivel interfamiliar. Se deben excluir todos los peces pequeños, de manera que en la población de peces reproductores seleccionados no existirán individuos enanos. En la fase de selección interfamiliar del programa de cría se descartarán algunos peces de gran tamaño, pero ya se ha indicado antes que posiblemente su tamaño no es heredable.
Una vez que se ha utilizado la selección familiar para crear la población de peces reproductores seleccionados, puede producirse descendencia mediante dos distintos protocolos de apareamiento. El primero y más sencillo consiste en aparear al azar los peces seleccionados. El segundo, que se utiliza con frecuencia, consiste en aparear los machos de la familia 1 con las hembras de la familia 2, los machos de la familia 2 con las hembras de la familia 3, los machos de la familia 3 con las hembras de la familia 4 etc., y aparear los machos de la última familia con las hembras de la familia 1. Este tipo de apareamiento (“apareamiento rotatorio”) tiene por objeto impedir la endogamia o reducirla al mínimo. En la segunda generación, y también en las generaciones sucesivas, se escalonan los apareamientos (los machos de la familia 1 × familia 2 se aparean con las hembras de la familia 3 × familia 4, etc.).
Como es lógico, este procedimiento exige marcar los peces reproductores seleccionados, utilizar un gran número de estanques de desove y registrar un importante volumen de datos. Aunque este tipo de apareamiento es conveniente para un programa de cría comercial en gran escala, no es necesario en la acuicultura de escala mediana, en la cual es suficiente el apareamiento al azar de los peces reproductores seleccionados.
Los programas de cría selectiva son costosos y, por ello, es importante poder evaluar con exactitud los resultados de la selección. La mejor manera de hacerlo consiste en comparar la media del carácter en el transcurso del tiempo con una población control no seleccionada. Para evaluar los resultados de la selección pueden aplicarse otras técnicas que, sin embargo, exigen criar o mantener otra población seleccionada (seleccionada en la dirección opuesta) o elaborar estadísticas muy complejas.
La razón por la que es necesario comparar la población seleccionada con una población control es que la media fenotípica de la población está determinada por su genoma (que se trata de mejorar durante la selección) y por el medio ambiente. El peso medio de la población seleccionada puede aumentar o permanecer invariable en función de determinados factores ambientales adversos, como un tiempo excepcionalmente frío o nuboso durante el período de crecimiento, que en la fábrica de piensos se haya utilizado sin saberlo harina de pescado de mala calidad para la fabricación del pienso o que el piscicultor aumente la densidad de población de, por ejemplo, 4.000 a 8.000 peces por hectárea. Si el piscicultor sólo conoce el peso medio de la población seleccionada y lo compara con el peso medio de la generación anterior llegará a la conclusión (tal vez incorrecta) de que la selección no está dando el resultado esperado. Puede ocurrir, a la inversa, que la selección no haya producido mejora genética pero que se registre un incremento del peso medio porque el tiempo es excepcionalmente bueno, porque la fábrica de piensos produce un alimento de mejor calidad o porque el piscicultor ha mejorado la gestión. Si únicamente se conoce el peso medio de la población seleccionada, al compararlo con el peso medio de la generación anterior se concluiría erróneamente que la selección había originado un aumento de la tasa de crecimiento.
Sólo la existencia de una población control permite establecer si esa conclusión es correcta. El peso medio debe aumentar con el paso del tiempo, a medida que el piscicultor aplica mejores métodos de manejo, y a través de la domesticación, pero si no existe una población control es imposible saber qué porcentaje del incremento ha sido consecuencia de la cría selectiva, qué porcentaje se debe a la mejor calidad de la gestión, etc.
En una piscifactoría grande, los peces reproductores control F1 pueden ser una muestra al azar de la generación P1 (población original) antes de que se iniciara la selección (Figura 28). Otra forma de conseguir peces reproductores control F, consiste en tomar una muestra de peces alrededor de la media de la generación P,. Se mantendrán en el criadero los peces control F, para cultivarlos con el mismo sistema con el que se cultivan los peces reproductores seleccionados F,. Es necesario que las condiciones de manipulación y desove de los peces reproductores de ambas poblaciones sean idénticos y es importante cultivar a su descendencia en las mismas condiciones o con las menores diferencias posibles, porque las diferencias en la técnica de cultivo pueden influir en el fenotipo y dificultar la evaluación precisa de los resultados de la selección. La única excepción en este sentido consiste en que en las generaciones subsiguientes (F2 etc.), los peces reproductores de la población control se eligen al azar, mientras que los peces reproductores seleccionados se eligen de entre los peces que forman la población seleccionada.
Otra forma de conseguir una población control consiste en mantener los peces reproductores P1 aparearlos durante varias generaciones y utilizar su descendencia como población control. El único inconveniente de este procedimiento puede radicar en que la edad y el tamaño de las hembras tengan una influencia notable en el crecimiento de la descendencia. Esto ocurre frecuentemente en las poblaciones piscícolas, pero normalmente los efectos desaparecen en un plazo de 20–180 días. Si la selección se realiza antes de que hayan desaparecido dichos efectos, no debe utilizarse este sistema para producir la población control.
La creación y mantenimiento son procesos idénticos en la selección individual y familiar.
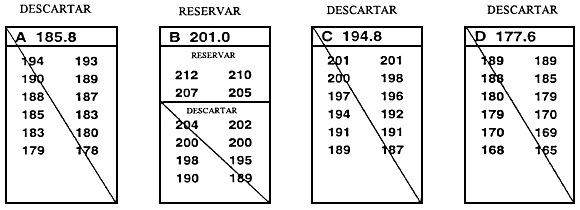
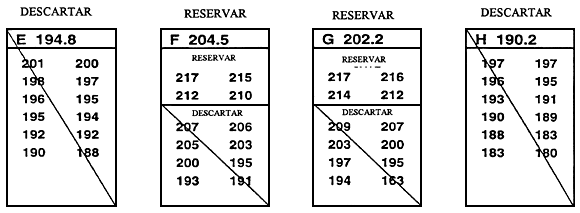
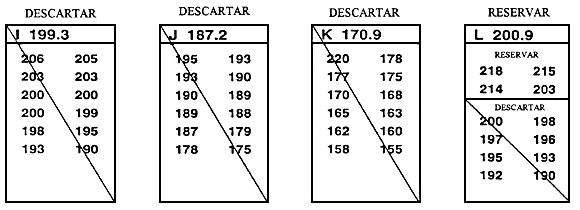
Figura 27. Diagrama esquemático de un programa de cría selectiva que combina la selección interfamiliar con la selección intrafamiliar. Este programa consta de dos fases. En este ejemplo, el objetivo era aumentar la tasa de crecimiento mediante selección para la longitud. Junto a la letra que identifica a cada familia figura el valor correspondiente a la media (en milímetros); bajo la letra se indican las longitudes individuales. En la primera fase se efectúa la selección interfamiliar y la segunda consiste en realizar la selección intrafamiliar en las familias seleccionadas. En este ejemplo, en la primera fase se reservaron mediante selección interfamiliar las cuatro mejores familias (B,F,G y L). A continuación, se reservaron mediante selección intrafamiliar los cuatro mejores peces de las familias B,F,G y L, descartándose todos los demás. Al igual que en las figuras 25 y 26, se asume que no existe dimorfismo sexual. Si lo hay, es necesario actuar como se indicó en las figuras 25 y 26. Este programa de selección de dos fases origina una población de peces reproductores seleccionados formada por los mejores individuos de las mejores familias.
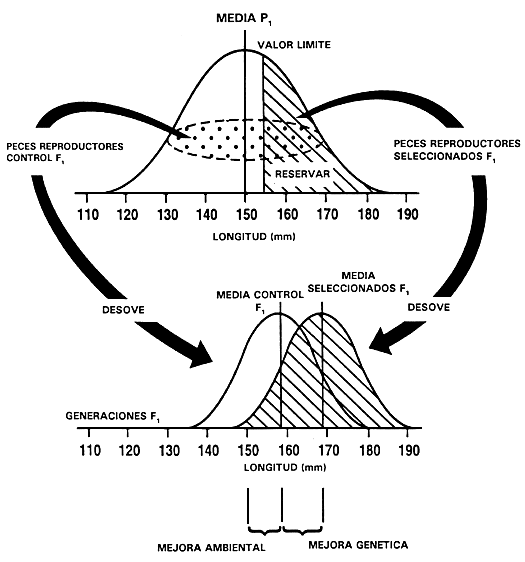
Figura 28. Se necesita disponer de una población control para evaluar la mejora conseguida mediante la selección. En este ejemplo, la longitud media de la generación P, es de 150 mm. Antes de iniciar la selección, se tomó una muestra al azar de dicha población para obtener los peces reproductores control F, (zona de puntos de la curva). Una vez conseguidos los peces reproductores control F, se seleccionó el 30% mejor de la población (valor límite = 155 mm). Los peces reproductores seleccionados P, y control F1 fueron cultivados en condiciones idénticas y se aparearon al alcanzar la edad madura, produciéndose las generaciones F1. Las generaciones seleccionada F1 y control F, se cultivaron con arreglo al mismo sistema. Al cosechar las generaciones F1( la longitud media de los peces seleccionados F, era de 169 mm y la de los peces control F1 de 159 mm. De no haber existido un control, los resultados habrían indicado que gracias a la selección la longitud media había pasado de 150 a 169 mm, lo que supone un incremento de 19 mm. Sin embargo, la existencia de una generación control F1 pone de manifiesto que una parte importante de la mejora de 19 mm se debió a factores ambientales. La media de los peces control F1 era de 159 mm, lo que supone una mejora de 9 mm por la acción de factores ambientales (159 mm - 150 mm); por tanto, la mejora genética fue de 10 mm (169 mm - 159 mm).
La creación y mantenimiento de una población control es el sistema más adecuado para evaluar los resultados de la selección, pero es costoso en tiempo, espacio y dinero. Por lo general, los piscicultores no pueden o no quieren mantener una población control. En tal caso, pueden comparar el rendimiento medio o el tamaño medio de los peces seleccionados con los cultivados en una piscifactoría de las proximidades o en un criadero de propiedad estatal. Para aplicar este plan, el piscicultor debe evaluar cuidadosamente los planes de gestión de las diferentes piscifactorías y del criadero estatal y elegir uno que sea análogo al que él utiliza. Además, debe seleccionar un piscicultor o administrador de un criadero que colabore con él y que previsiblemente vaya a seguir trabajando en esa esfera de actividad durante los diez años siguientes.
Cuadro 12. Cómo utilizar una población control para evaluar los resultados de la selección.
| Dadas: | longitud media de la población P, = 150 mm |
| longitud media de la generación seleccionada F1 = 165 mm | |
| longitud media de la generación control F1 = 159 mm | |
| La mejora total se calcula de la siguiente manera: | |
| Mejora total = (longitud media de la generación seleccionada F1 - longitud media de la generación P1) = 165 mm - 150 mm = 15 mm | |
| La mejora obtenida por efecto de los factores ambientales se calcula de la siguiente manera: | |
| Mejora debida a factores ambientales = (longitud media de la generación control F1 - longitud media de la generación P1) = 159 mm - 150 mm = 9 mm | |
| La mejora genética se calcula de la siguiente forma: | |
| Mejora genética = (longitud media de la generación seleccionada F1 - longitud media de la generación control F1) = 165 mm - 159 mm = 6 mm | |
| Mejora genética = (mejora total - mejora debida a factores ambientales) = 15 mm - 9 mm = 6 mm | |
En el cuadro 12 se muestra cómo puede utilizarse una población control para evaluar los resultados de la selección. En este ejemplo, la media de la generación P, era 150 mm y la media de la generación F1 de peces reproductores seleccionados, 165 mm. Si no se hubiera cultivado una población control, se sabría que el incremento promedio había sido de 15 mm, pero sería imposible determinar qué porcentaje de ese incremento se había conseguido gracias al programa de cría selectiva, cuál por la aplicación de un sistema mejor de manejo, etc. La existencia de una población control permite establecer que 9 de los 15 mm corresponden a la mejora obtenida por factores ambientales. Eso significa que si no se hubiera aplicado un programa de cría selectiva, la longitud media habría aumentado de 150 a 159 mm. La diferencia entre las generaciones seleccionada F1 y control F1 (diferencia entre la mejora total y la mejora debida a factores ambientales) es 6 mm, que es la mejora genética.
La existencia de una población testigo permite determinar también el porcentaje de la mejora que se debe a la selección. En el transcurso de una generación, la cría selectiva aumentó la longitud media en el 3,8% (6 mm/159 mm). Obsérvese que la mejora porcentual se evalúa utilizando como valor estándar el grupo de control contemporáneo (en este caso, la generación control F1).
Cuando se expresa en forma porcentual, la mejora genética puede expresarse como porcentaje por generación, como se hizo anteriormente, o como porcentaje anual. Si el intervalo entre generaciones es de un año, los valores son los mismos, pero si el intervalo es mayor o menor, los valores son menores o mayores, respectivamente. Por ejemplo, si la especie no madura y se aparea hasta los cuatro años de edad, la mejora porcentual por generación se divide entre cuatro para establecer la mejora porcentual anual.
Cuando un piscicultor compara los peces seleccionados con la población control debe comparar los valores medios globales obtenidos con peces cultivados al menos en tres estanques. No importa que utilice su propia población control o los peces de un piscicultor vecino. Si cada población se cultiva en un único estanque no podrá separar los efectos debidos a las condiciones del estanque de los efectos genéticos. La media de una población puede ser más alta porque en el estanque que ocupa la floración de algas es mayor.
Uno de los procedimientos que se sigue para no tener que cultivar las dos poblaciones en estanques iguales consiste en marcar el grupo control y cultivarlo conjuntamente con la población seleccionada en un mismo estanque. En tal caso, basta con depositar en un estanque de 30 a 50 peces control.
Para evaluar con total precisión los resultados de un programa de cría selectiva no basta con comparar la media de la población seleccionada con la de la población control. Las medias se han de comparar estadísticamente. Lamentablemente, los análisis estadísticos de los datos de un programa de cría selectiva exigen un conocimiento completo del proyecto experimental y unos conocimientos básicos de estadística. Por ello, prácticamente todos los piscicultores deberán recurrir a la asistencia de un experto en análisis de datos. Si eso no es factible, el piscicultor no debe renunciar al programa de cría selectiva por la imposibilidad de efectuar un análisis estadístico de los resultados. La media aproximada será una buena indicación de lo ocurrido.
Los programas de cría selectiva necesarios para mejorar los caracteres cuantitativos son más difíciles, complejos y largos que los que deben aplicarse en el caso de los caracteres cualitativos. De hecho, estos programas son de duración indefinida; si se interrumpe la selección, la población pierde gran parte de la mejora conseguida. Además, exigen un sistema de registro de datos más complejo y una cierta labor de análisis matemático/estadístico de los datos recogidos. En el Capítulo 5 se presentan ejemplos de programas de cría sencillos y poco costosos para mejorar diversos caracteres cuantitativos como la tasa de crecimiento, así como ejemplos de registros de datos que se deben mantener.
Los programas de cría selectiva son métodos experimentados y eficaces para aumentar la tasa de crecimiento y el rendimiento, pero entrañan unos gastos y son de un nivel tecnológico elevado. Si es posible aumentar los rendimientos mediante la utilización adecuada de cal, fertilizantes y/o piensos o introduciendo mejoras por lo que respecta a la densidad de población o la gestión de la calidad del agua, deben utilizarse primero estos métodos porque producen una mejora inmediata y son relativamente poco costosos.
Es bastante reducido el número de piscicultores a los que se debe aconsejar que utilicen programas de cría selectiva, porque para llevarlos a la práctica hacen falta estanques, mano de obra, dinero y registro de datos. También exigen una planificación a largo plazo y una dosis de paciencia. Los piscicultores que no pueden o no quieren dedicar una parte de su piscifactoría al programa de cría selectiva, aportar la mano de obra necesaria y registrar y mantener datos no deben iniciarlos. Tampoco se debe alentar a iniciar programas de cria a los piscicultores que no pueden efectuar una planificación a largo plazo (1–10 años).
Para utilizar programas de cría selectiva con objeto de aumentar la tasa de crecimiento, no es necesario que los piscicultores se conviertan en genetistas. No sólo no necesitan conocer la heredabilidad de la longitud y el peso, sino tampoco qué es la heredabilidad. Sin embargo, deben ser buenos administradores.
Antes de iniciar un programa de cría selectiva, el piscicultor debe emprender un estudio para establecer si es necesario y qué caracteres deben ser objeto de mejora. En la mayoría de los casos, lo más lógico es que la finalidad de la mejora sea aumentar la tasa de crecimiento. Por regla general, ésta ha de ser la única meta de los programas de cría selectiva en las piscifactorías de tamaño medio, y ello porque la mejora que puede obtenerse para un carácter es inversamente proporcional al número de caracteres que se incorporan al programa de mejora. Además, el aumento de la tasa de crecimiento mejorará también otros caracteres productivos importantes mediante la selección indirecta.
Tras decidir qué carácter desea mejorar, el piscicultor debe establecer cómo y cuando medir el carácter y qué programa de cría selectiva utilizará para crear la población de peces reproductores seleccionados (selección individual, intrafamiliar o interfamiliar). En el Cuadro 13 se exponen las diferencias entre estos programas. Siempre que sea posible, debe utilizarse la selección individual, porque además de ser más fácil y menos costosa exige un menor registro de datos.
Cuando desea mejorar dos caracteres, el piscicultor debe recurrir al descarte independiente o al descarte independiente modificado. Nunca debe utilizar la selección en tándem. Aunque el índice de selección es el programa de cría más eficiente para mejorar dos caracteres, no se recomienda para las piscifactorías de tamaño medio.
El piscicultor que ponga en práctica un programa de cría selectiva debe disponer de los medios necesarios para evaluar los resultados de su trabajo. Si cuenta con el espacio y los recursos necesarios, puede mantener una población control. De no ser así, puede comparar la media de la población seleccionada en el transcurso del tiempo con la de una piscifactoría de las proximidades o con la de un criadero estatal.
Una vez establecida la mejora conseguida con el programa de cría selectiva, debe reevaluar el programa. Ha de examinar todas sus facetas y si existen formas más sencillas y eficientes de realizarlo. Evaluará también si la inversión que ha efectuado produce los dividendos esperados. Sin duda, el piscicultor debe abandonar el programa de cría selectiva si no funciona adecuadamente, pero no debe emitir un juicio apresurado sobre los resultados de la selección. Si se ejecuta adecuadamente, el programa de cría selectiva producirá una ganancia pequeña pero constante.
Cuadro 13. Comparación de las estrategias básicas e indicación de algunas de las ventajas y desventajas de la selección individual, intrafamiliar e interfamiliar.
| Tipo de selección | Estrategia | Ventajas | Desventajas |
| Individual | Elegir los mejores ejemplares; las relaciones familiares carecen de importancia | El más indicado cuando H2 ≥ 0,25; poco costoso; puede realizarse en un reducido número de estanques; relativamente fácil de aplicar para 2 o 3 caracteres; todos los peces seleccionados son los de mayor tamaño; es fácil mantener una población reproductora importante; es el que exige un menor registro de datos. | Ineficaz cuando H2.≤ 0,15 porque VE hace difícil elegir los mejores peces; el desove asincrónico puede causar problemas. |
| Intrafamiliar | Elegir los mejores ejemplares de cada familia | El más indicado cuando h2≤ 0,15 y VEe, afecta a la familia más que a los individuos; puede utilizarse cuando el desove es asincrónico; relativamente fácil mantener una población reproductora numerosa; menos costoso que la selección interfamiliar | Moderadamente costoso, requiere gran número de estanques; difícil incorporar 2 o 3 caracteres; los peces pequeños pueden ser peces reproductores seleccionados. Exige un gran volumen de registro de datos. |
| Interfamiliar | Elegir las mejores familias basándose en medias familiares; no se tienen en cuenta los valores individuales | El más indicado cuando H2.≤ 0,15 y VE afecta a los individuos más que a las familias; puede utilizarse cuando es necesario dar muerte a los peces. | Muy costoso; requiere gran número de estanques; difícilincorporar 2 o 3 caracteres; los peces pequeños pueden ser peces reproductores seleccionados; puede ocasionar elevados niveles de endogamia. Exige registrar un gran volumen de datos |
![]()
![]()
![]()