![]()
![]()
![]()
por
Gösta Eriksson, Gene Namkoong, y James Roberds1
RESUMEN
Es generalmente aceptado que los programas de conservación, para que tengan éxito, deben contar con la variación genética. Sin embargo, hay un gran desacuerdo sobre los métodos a emplear para la conservación genética en estos programas. El origen de esta controversia radica en los supuestos que se hacen sobre los recursos genéticos y su dinámica. Un supuesto equivocado es que la evolución de las poblaciones naturales conduce a la adaptación máxima. Por el contrario, la selección, la deriva genética y la migración de genes son fuerzas evolutivas que actúan entre si de formas complejas de tal modo que nunca se puede obtener una adaptación máxima. Los continuos cambios del medio ambiente fuerzan también a la selección natural a actuar en múltiples direcciones. Además, la mayoría de los rasgos de importancia están controlados por una serie de loci genéticos que tienen alelos con efectos que son intercambiables. Por estas razones, los recursos genéticos actuales se deben considerar como punto de partida para las actividades de conservación genética y no como una meta de los programas de conservación genética.
El objectivo principal de la conservación genética debe ser crear las condiciones que favorezcan la evolución futura del recurso genético. Los métodos empleados deben garantizar el cumplimiento de los objectivos del programa, teniendo en cuenta tanto la dinámica como la estructura genética. El Sistema de Mejora Genética mediante Poblaciones Múltiples, que combina la conservación y la mejora genética, es el mejor método de dirigir los programas de conservación para este fin.
INTRODUCCION
Hay una amplia aceptación pública de que deben conservarse las especies y los genetistas forestales y el público bien informado están de acuerdo sobre la conservación de la variación genética como elemento necesario de cualquier programa de conservación. La variación no sólo es necesaria para garantizar la adaptabilidad actual de las especies sino también para la evolución continua de las especies. Sin embargo, hay un gran desacuerdo sobre los métodos de conservación genética. Siguen existiendo, por ejemplo, discusiones sobre el tamaño más efectivo de las muestras, los métodos in-situ vs. ex-situ, y la regeneración natural vs. la artificial. En este documento analizamos la variación genética desde el punto de vista de la evolución y describimos una estrategia programática que cumple los objetivos más importantes de la conservación genética forestal. Para desarrollar un programa eficaz tenemos que satisfacer tanto los principios científicos de los genetistas como los principios éticos. Analizamos en primer término estos principios y a continuación sugerimos algunas directrices para un programa de conservación.
LOS RECURSOS GENETICOS
Una causa importante de desacuerdo sobre los métodos de conservación son las suposiciones realizadas sobre la naturaleza del recurso genético y su dinámica. Una suposición incluye el número de genes que afectan a un rasgo. Si cada rasgo estuviera controlado por un solo locus, como se puede creer popularmente, en tal caso todo alelo sería importante en todo locus y cualquier pérdida disminuiría la capacidad de una población para responder a los desafíos ambientales. Esto ha llevado a poner el énfasis en favorecer las grandes colecciones y reservas para almacenar alelos y, cuanto más raro es el alelo, mayor debe ser la colección. Sin embargo, muchos rasgos de importancia están controlados por muchos loci genéticos y cada locus tiene efectos que controlan la expresión de muchos rasgos. De este modo, el efecto de un alelo en un locus puede ser sustituido con frecuencia por cualquiera de varios alelos en aquél o en otros loci que actúan en la misma o en otras vías metabólicas. Vamos a utilizar un ejemplo simplificado para ilustrar sobre este tipo de control genético. Supongamos que todos los alelos con un índice común contribuyen igualmente a la adaptabilidad, independientemente del locus. Los alelos con índices diferentes, hacen sin embargo contribuciones diferentes a la adaptabilidad. Entonces, el genotipo A1A1B1B2C1C2 tendrá la misma adaptabilidad que el A2A2B1B1C1C1, y así sucedería con todos los otros genotipos que tengan cuatro alelos con índice 1. Para la evolución hasta un producto final determinado hay muchas opciones, no dependiendo la evolución de unos genes individuales.
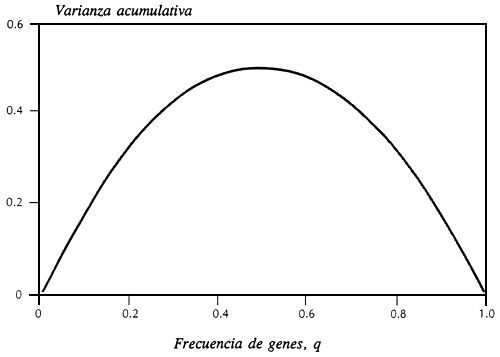
Los rasgos medios se pueden variar sustituyendo alelos que tengan efectos acumulativos, siendo ocasionados los mayores cambios por alelos que no son ni muy comunes ni muy raros. La Figura 1 demuestra que los alelos con frecuencia inferior a 0,01 o superior a 0,99 tienen poco efecto sobre la varianza genética acumulativa y por lo tanto poco efecto inmediato sobre la adaptabilidad. Por ello, no es necesario salvar todos los alelos en todos los loci y, de hecho, la evolución en las próximas generaciones no depende de los alelos raros, por lo que su salvación no es una preocupación inmediata. Existen preocupaciones legítimas en cuanto al papel de los alelos raros en la evolución a largo plazo, pero nosotros podemos manejar el recurso en razón de aquellas preocupaciones sin tener que crear siempre mayores colecciones y reservas para almacenar alelos singulares.
Figura 1. Dependencia de la varianza acumulativa sobre la frecuencia de genes en un locus único con dos alelos, siendo totalmente acumulativa la acción de los genes.
Otra causa de desacuerdo es la suposición de que la evolución en la regeneración natural siempre da el máximo de adaptabilidad y que la mínima intervención del hombre es la mejor estrategia de conservación. Sin embargo, los resultados de la selección reflejan los efectos de las condiciones previas y además, la selección no es la única fuerza de evolución que configura el recurso genético. Como los principales efectos de la selección operan al nivel del organismo individual, los efectos de la selección sobre rasgos específicos son dependientes como un rasgo está relacionado con otros rasgos que constituyen la adaptabilidad total. De este modo, los rasgos están a veces correlacionados negativamente lo que significa que el avance de un rasgo lleva al retroceso de otro rasgo. La plasticidad fenotípica permite también la supervivencia de los individuos incluso aunque no estén genéticamente adaptados al máximo a un ambiente específico. Además, algunos genes y rasgos de poca importancia para la adaptabilidad se cambian simplemente por un efecto de “tirón” de una correlación accidental. Más aún, ciertos genes individuales suelen afectar pleiotrópicamente a varios rasgos y por ello es raro poder afirmar que algún rasgo pueda estar a nivel óptimo. El continuo cambio del ambiente, con o sin acontecimientos importantes como p. ej. el cambio climático mundial, fuerza a la selección natural a actuar en múltiples direcciones. Por ello, nunca se alcanzará la adaptabilidad máxima por aquella misma razón.
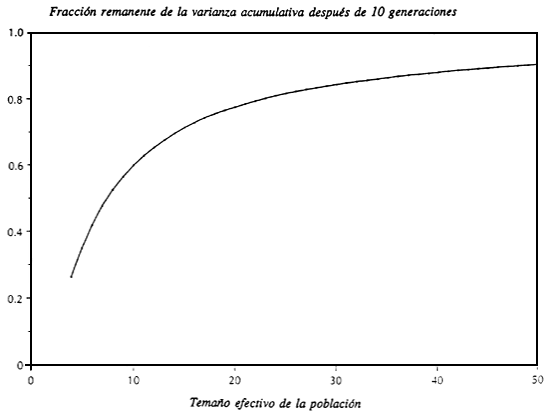
Entre las otras fuerzas de la evolución que afectan a la estructura del recurso genético se encuentran la deriva genética y su contrapartida, la migración. Poblaciones de pequeño tamaño hacen posible que los procesos al azar afecten fuertemente a las distribuciones genéticas y reduzcan la variación genética dentro de tales poblaciones. Esto se ilustra en la Figura 2 que revela que sólo algo más del 50% de la varianza acumulativa permanece después de 10 generaciones con un tamaño efectivo de población (No) = 8, perdiéndose más del 25 % de la varianza para No = 16. La deriva genética es una poderosa fuerza evolutiva que puede anular el efecto de la selección natural en pequeñas poblaciones.
Figura 2. Fracción remanente de la varianza acumulativa después de diez generaciones para distintos tamaños de población efectiva. Se supone que no hay otras fuerzas evolutivas que influyan en el resultado.
Para comprender los efectos de la migración es conveniente considerar el concepto de los entornos ambientales selectivos (SENs) introducido por Brandon (1990). Dentro de un SEN determinado, los árboles no cambian de categoría respecto a la adaptabilidad. Cuando existe un flujo de genes entre varios SENs, el efecto de la selección natural disminuye con respecto a lo que cabría esperar a priori. Esto produce una gran variación de la adaptabilidad dentro de cada población a través de los SENs, pero también se traduce en una menor adaptación para cualquier estación dada. Así, el intercambio entre distintos SENs produce probablemente la gran variación de los rasgos de adaptación observada en las poblaciones de Pinus sylvestris y Picea abies (Eriksson 1982). La deriva genética puede ser fuerte en algunas poblaciones de árboles tropicales, pero para muchas especies de árboles forestales de zonas boreales templadas de polinización anemófila, la migración es fuerte, pudiendo contrarrestar los efectos de la deriva, al menos periódicamente.
En resumen, las fuerzas evolutivas de selección, deriva y migración actúan de formas complejas, no llevándonos a confiar en que la naturaleza produzca la perfección. El estado actual del sistema genético puede considerarse como la condición inicial con que contamos para el futuro, pero no podemos considerarlo como sagrado, y su mantenimiento sin cambios no puede constituir la meta de la conservación. No obstante, es evidente que cualquiera que sea lo producido por la evolución, las especies que han sobrevivido han tenido éxito hasta aquí y son por tanto una solución viable para su historia pasada. Queda la duda sobre su carácter óptimo para las condiciones futuras, especialmente frente a los cambios ambientales (incluyendo el posible cambio climático mundial).
Para las especies de árboles forestales el desacuerdo sobre los métodos de conservación parece basarse, al menos en parte, en conceptos erróneos sobre la naturaleza del recurso genético. Si el recurso se entiende como un sistema dinámico, la estructura de cualquier especie debería considerarse como una de las muchas estructuras posibles, siendo la estructura actual, por su propia naturaleza, un estado transitorio. Los intentos de congelar el estado actual irían contra la dinámica natural del sistema y no cumplirían los objetivos de la conservación genética forestal.
ETICA
Como existen fuertes limitaciones de tiempo, personal y fondos para la conservación, siempre tendremos que elegir sobre los genotipos, poblaciones o especies en que invertir nuestro tiempo, fondos y conocimientos. Con frecuencia se proclama, como una perspectiva ética, la conocida “falacia naturalista” mediante la cual todo lo que existe naturalmente es considerado intrínsecamente bueno. A partir de aquí surgen dos directrices: todas y cada una de las cosas existentes deben conservarse y ninguna acción humana es mejor que no actuar. Estas directrices nos fuerzan entonces a no adoptar decisiones o a conservar solamente por capricho. Ninguno de estos resultados nos da una orientación razonable o unas prioridades de conservación que puedan justificarse con vistas a la inversión pública. Además, tal proclama otorga un peso indebido a lo que es en realidad un estado biológicamente transitorio, sin justificar la preferencia. La falacia estriba en proclamar que un estado particular del mundo es bueno sin justificarlo de otro modo que afirmando simplemente su verdad.
Para justificar éticamente la conservación, hay que aportar razones poderosas sobre por qué en un mundo de limitaciones, deben dirigirse las inversiones a los programas de conservación. Un problema básico es aclarar para el bien de quiénes conservamos. La inmensa mayoría estaría de acuerdo en incluir el bienestar de los seres humanos y que es el bien de las generaciones actuales y futuras el que justifica en gran parte la preocupación por la conservación. Esto obedece a la creencia de que los seres humanos tienen valores intrínsecos diferentes del valor de las herramientas que son instrumentos para nuestro bien. El problema para los conservacionistas consiste en si las entidades biológicas distintas de los seres humanos individuales, tienen un valor intrínseco y, de ser así, cómo incorporamos el bien intrínseco de estas otras entidades a los objetivos de la conservación. Existe una extensa bibliografía sobre el debate acerca de este tema pero, para los fines de este documento, puede señalarse que incluso aunque no asignásemos valores intrínsecos a otros seres, aún podría justificarse prácticamente la conservación de todas las especies (Norton 1987). Un programa de conservación basado exclusivamente en el valor intrínseco de los seres humanos actuales y futuros, no excluiría la conservación de otras especies de valor directo e indirecto para los seres humanos sino que podría incluir de hecho un enorme número de especies debido a su utilidad para el hombre, a su utilidad para otras especies que son directamente útiles para el hombre, etc. El basar la conservación de las especies en su utilidad directa e indirecta para los seres humanos evita también el problema de tratar de justificar cómo los grupos de individuos pueden reclamar un valor intrínseco. No parecen existir propiedades de grupos como las especies, de carácter distinto de las de sus miembros individuales, que otorguen al grupo una consideración especial. Así la rareza es una propiedad del grupo pero no implica de por si tanto un valor intrínseco como el de que la ordenación sea eficaz en el salvamento de genes al salvar los pocos genotipos remanentes. Un fundamento más sólido para la conservación de las especies se puede basar en sus valores para los seres humanos, en cuyo caso es mucho más fácil establecer también prioridades. Como advertencia previa debemos señalar que adoptamos un criterio amplio sobre lo que constituye el valor para los seres humanos actuales y futuros.
OBJETIVOS
En la sección anterior y en la precedente (Eriksson et al. 1993) hemos sostenido que la naturaleza se
comporta dinámicamente y esto representa que la conservación genética debe ser también dinámica. El
potencial para la evolución dinámica es especialmente importante en tiempos de grandes cambios
ambientales. De aquí se deduce que un objetivo importante es la creación de unas buenas condiciones para la evolución futura.
Un objetivo secundario, pero importante, es la captura de genes con frecuencias > 0,01 y la captura de las
adaptaciones existentes que pueden producirse en diferentes poblaciones. Esto último puede considerarse en
contradicción con nuestras afirmaciones anteriores. La razón de este objetivo es aumentar la probabilidad
de capturar la máxima varianza acumulativa posible en las poblaciones de recursos genéticos. Tal objetivo
está totalmente de acuerdo con nuestros argumentos anteriores. Un tercer objetivo es incorporar un
componente añadido de mejora genética. Esta es una función especialmente importante para los programas
de conservación que incluyan especies comercialmente importantes.
Los árboles no son los únicos organismos que constituyen un ecosistema forestal. Otras muchas especies dependen de ciertas especies arbóreas para su supervivencia y bienestar. Tales especies se denominan especies asociadas a diferencia de las arbóreas (especies principales) que son el centro de atención de un programa de conservación forestal. Para mantener la credibilidad, es importante incluir entre nuestros objectivos la conservación de las especies asociadas.
Algunos conservacionistas defienden la preservación de la actual estructura genética. La principal razón es que la estructura actual sirve como referencia para comparaciones futuras. Otros defienden la prioridad de las poblaciones en peligro. Ambos objetivos tienen su mérito pero son de menos importancia que nuestro objetivo principal.
Tal como se afirmó en la sección anterior, no es suficiente que un elemento genético sea raro para justificar su conservación por razones éticas.
CONOCIMIENTOS GENETICOS
El análisis de la estructura genética de una población proporciona la información básica necesaria para diseñar procedimientos eficaces de muestreo para los programas de conservación genética. La estructura puede variar dependiendo del rasgo para el que se determina. Con respecto a la estructura de la población, los rasgos pueden agruparse en dos clases principales: los marcadores moleculares y los rasgos métricos. Se sabe en la actualidad que la primera clase de rasgos muestra en muchas especies una variación limitada entre poblaciones mientras que los rasgos métricos muestran en un gran número de especies una varianza considerable, tanto entre poblaciones como dentro de ellas. Esto es consecuencia de la historia evolutiva previa de ambos tipos de rasgos. Los marcadores moleculares son neutrales o casi neutrales y reflejan la historia evolutiva a largo plazo mientras que muchos rasgos métricos están asociados con la adaptación y reflejan en consecuencia acontecimientos de selección recientes. Antes de poner en práctica un programa de muestreo de conservación es importante investigar las estructuras de población para ambos tipos de rasgos. Hay que evitar el muestreo basado exclusivamente en datos de marcadores moleculares.
Para la mayoría de las especies arbóreas nunca obtendremos un conocimiento adecuado sobre la estructura genética, ni para los marcadores moleculares ni para los rasgos métricos. La escasez de información no sirve de excusa para no actuar. Como no se dispone de los conocimientos necesarios que sirvan de base para un programa de conservación, habrá que depender de suposiciones bien fundadas sobre las posibles estructuras de la población. Estas suposiciones pueden basarse en las características ecológicas de las especies y también en su hábitat de distribución.
La investigación sobre estructura de la población debe centrarse en especies con diferentes combinaciones de rasgos ecológicos. Estas especies pueden emplearse como modelos en los que basar los programas de conservación de otras especies. Cuando se dispone de datos de suficientes especies, pueden desarrollarse directrices a utilizar con especies cuyas estructuras de población son desconocidas.
METODOS
La creación de unas buenas condiciones para la evolución futura se definió anteriormente como un objetivo primordial para la conservación genética. El método de conservación elegido para una especie debe garantizar por tanto su potencial evolutivo. Esto se consigue muy fácilmente desarrollando una colección de meta- poblaciones dentro de la especie. La conservación no debe limitarse a la preservación de la varianza acumulativa existente sino que debe promover si es posible un aumento de este tipo de varianza. El Sistema de Mejora Genética Mediante Poblaciones Múltiples (MPBS) desarrollado por Namkoong (1976, 1984) proporciona un método para cumplir este objectivo. De acuerdo con el MPBS, una población combinada de mejora y recursos genéticos se divide en unas 20 subpoblaciones, cada una con 50 entes genéticos aproximadamente. Las ventajas de dividir la población en subpoblaciones pueden resumirse como sigue:
En este contexto, la conservación genética es algo más que la mera conservación genética, convirtiéndose en un proceso dinámico que puede incluir y apoyarse en la mejora genética de árboles a largo plazo. En este proceso, aumenta la varianza entre poblaciones mientras se mantiene la varianza existente dentro de las poblaciones. Para un tamaño efectivo de población de 50 la tasa de pérdida de varianza acumulativa es del 1 % por generación. Con una gestión razonable parte de tal pérdida se puede evitar mediante selección. La frecuencia de mutaciones en los loci que controlan los rasgos métricos se ha calculado en algunas especies entre el 10-2 – 10-3 por generación (Lande y Burrowclough 1987). De este modo, se espera que las mutaciones de rasgos adaptables compensen parte de la pérdida de varianza acumulativa ocasionada por la deriva genética.
En su forma más detallada el MPBS requiere el establecimiento de subpoblaciones en una gran variedad de condiciones ambientales. Las plantaciones fuera del ámbito actual de las especies pueden ayudar también a prevenir acontecimientos futuros dudosos. Esta forma del MPBS será utilizada con gran probabilidad para especies arbóreas de importancia comercial debido a los elevados costos que representa. No obstante, las especies que tienen gran importancia para la existencia de otras numerosas especies, son también candidatos para esta forma del MPBS. Las ventajas del MPBS se pueden obtener también con la conservación genética in situ de especies menos importantes comercialmente, siempre que el muestreo de las poblaciones del recurso genético se realice para conseguir la máxima adaptabilidad existente. Dependiendo de las condiciones económicas, puede prescribirse una gestión intensiva que permita seleccionar los individuos deseados para que sirvan como árboles-padre para la próxima generación.
La investigación ha demostrado que muchas especies amenazadas dependen especialmente de la características del hábitat. Para cumplir el objetivo de la conservación genética de las especies secundarias, debe ser una meta la diversidad de hábitats. La expansión de algunas subpoblaciones en la variante in situ de la variante del MPBS incluyendo varios centenares de hectáreas, junto con un plan de ordenación para conseguir la máxima diversidad de hábitats dentro de estas grandes subpoblaciones debe servir para la conservación genética de la mayoría de las especies secundarias (cf. Varela y Eriksson 1995).
En conclusión, el MPBS es el método más eficaz para la conservación genética de árboles forestales pero tiene el inconveniente de los costes que representa la plantación. También se pueden diseñar variantes in situ del MPBS a utilizar con especies que actualmente se consideran menos importantes para servir al objetivo principal de la conservación genética de árboles forestales: crear buenas condiciones para la evolución futura.
REFERENCIAS
Brandon (1990). Adaptation and Environment. Princeton Univ. Press, Princeton, New Jersey. 214pp.
Eriksson, G. (1982). Ecological genetics of conifers in Sweden. Silva Fenn. 16: 149–156.
Eriksson, G.; Namkoong, G. & Roberds, J. (1993). Dynamic gene conservation for uncertain futures. For. Ecol. Manag. 62: 15–37.
Lande, R. & Burrowclough, G.F. (1987). Effective population size, genetic variation, and their use in population management. In Viable Populations for Conservation (Ed. M.E. Soulé), New York, Cambridge Univ. Press: 87–123.
Namkoong, G. (1976). A multiple index selection strategy. Silvae Genet. 25:199–201.
Namkoong, G. (1984). Strategies for gene conservation in forest tree breeding. In Plant Gene Resources: A Conservation Imperative. (Eds. C.W. Yeatman, D. Kafton, and G. Wilkes). AAAS Selected Symposium 87. Westview Press, Boulder CO.: 79–89.
Norton, B.C. (1987). Formal and substantive priority systems. In Why preserve Natural Variety, (Ed. M. Cohen). Princeton Univ. Press, Princeton, New Jersey: 243–257.
Varela, M.C. and Eriksson, G. (1995). Multipurpose gene conservation in Quercus suber - A Portuguese example. Silvae Genet. In press.
Recursos Genéticos Forestales No. 23. FAO, Roma (1995)
Manuscrito recibido en mayo de 1995
![]()
![]()
![]()