![]()
![]()
Exercício 6.7 Uso da curva de selectividade para ajustar capturas
| L50% = 13.6 cm | S1 = 13.6*ln 3/(14.6-13.6) = 14.941 |
| L75% = 14.6 cm | S2 = ln 3/(14.6-13.6) = 1.0986 |
| S(L) = 1/[1+exp(14.941-1.0986*L)] |
Folha de exercícios 6.7
| class de comp. L1–L2 | ponto médio | amostra viciada observada | ogiva de selectiv. S(L) | amostra estimada não viciada |
| 6– 7 | 6.5 | 3 | 0.00041 | 7326 a) |
| 7– 8 | 7.5 | 143 | 0.00123 | 116491 |
| 8– 9 | 8.5 | 271 | 0.00367 | 73769 |
| 9–10 | 9.5 | 318 | 0.01094 | 29067 |
| 10–11 | 10.5 | 416 | 0.03212 | 12952 |
| 11–12 | 11.5 | 488 | 0.09054 | 5390 |
| 12–13 | 12.5 | 614 | 0.2300 | 2670 |
| 13–14 | 13.5 | 613 | 0.4726 | 1297 |
| 14–15 | 14.5 | 493 | 0.7288 | 676 |
| 15–16 | 15.5 | 278 | 0.890 | 312 |
| 16–17 | 16.5 | 93 | 0.960 | 97 |
| 17–18 | 17.5 | 73 | 0.986 | 74 |
| 18–19 | 18.5 | 7 | 0.995 | 7 |
| 19–20 | 19.5 | 2 | 0.998 | 2 |
| 20–21 | 20.5 | 2 | 0.999 | 2 |
| 21–22 | 21.5 | 0 | 1.000 | 0 |
| 22–23 | 22.5 | 1 | 1.000 | 1 |
| 23–24 | 23.5 | 1 | 1.000 | 1 |
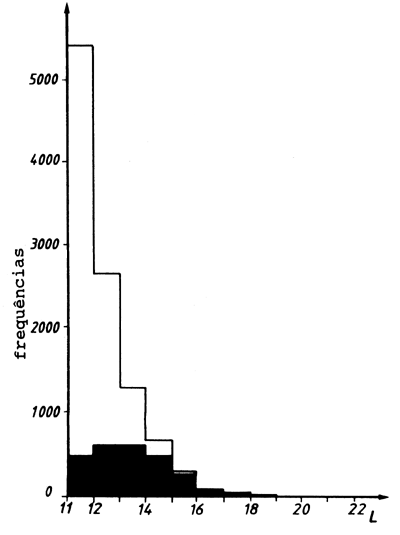
Fig. 18.6.7 Amostra viciada de salmonetes e amostra estimada não viciada,
corrigida para a selectividade. Fonte de dados: Ziegler, 1979
(ver Folha de exercícios 6.7)
Exercício 7.2 Amostragem aleatória estratificada contra amostragem aleatória simples e amostragem proporcional
Folha de exercícios 7.2
| estrato j | s(j) | s(j)2 | N(j) | ∑ Y(i,j)2 i | ∑ Y(i,j) i |
| 1 grande | 28.906 | 835.57 | 10 | 25413 | 423 |
| 2 médio | 8.569 | 73.43 | 30 | 9091 | 457 |
| 3 pequeno | 2.809 | 7.89 | 60 | 1524 | 252 |
| Total: | 100 | 36028 | 1132 |
Total 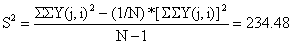
a)
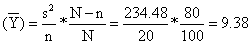
b) VARprop (Y) =
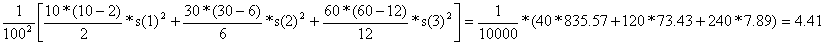
c) Amostragem estratificada óptima
| estrato j | s(j)*N(j) | 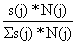 | 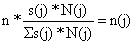 |
| 1 grande | 289.06 | 0.40 | 8 |
| 2 médio | 257.07 | 0.36 | 7 |
| 3 pequeno | 168.55 | 0.24 | 5 |
| Total: | 714.68 | 1.00 | n = 20 |
VARopt (Y) =
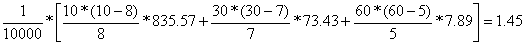
| aleatória | proporcional | óptima | |
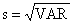 | 3.06 | 2.10 | 1.20 |
| repartições por estrato | |||
| 1 grande | ? | 2 | 8 |
| 2 médio | ? | 6 | 7 |
| 3 pequeno | ? | 12 | 5 |
Exercício 8.3 Modelo de captura por recruta de Beverton e Holt (captura por recruta, biomassa por recruta como uma função de F)
Folha de exercícios 8.3
| Tc = Tr = 0.2 | Tc = 0.3 | Tc = 1.0 | ||||
| F | Y/R | B/R | Y/R | B/R | Y/R | B/R |
| 0.0 | 0.00 | 8.28 | 0.00 | 8.00 | 0.00 | 4.53 |
| 0.2 | 1.36 | 6.81 | 1.33 | 6.67 | 0.79 | 3.96 |
| 0.4 | 2.28 | 5.71 | 2.26 | 5.65 | 1.41 | 3.51 |
| 0.6 | 2.91 | 4.85 | 2.92 | 4.86 | 1.89 | 3.15 |
| 0.8 | 3.34 | 4.18 | 3.39 | 4.24 | 2.28 | 2.85 |
| 1.0 | 3.64 | 3.64 | 3.73 | 3.73 | 2.60 | 2.60 |
| 1.2 | 3.84 | 3.20 | 3.98 | 3.31 | 2.86 | 2.39 |
| 1.4 | 3.97 | 2.84 | 4.15 | 2.97 | 3.08 | 2.20 |
| 1.6 | 4.06 | 2.54 | 4.28 | 2.68 | 3.27 | 2.05 |
| 1.8 | 4.11 | 2.28 | 4.38 | 2.43 | 3.43 | 1.91 |
| 2.0 | 4.14 | 2.07 | 4.44 | 2.22 | 3.57 | 1.79 |
| 2.2 | 4.15* | 1.88 | 4.49 | 2.04 | 3.69 | 1.68 |
| 2.4 | 4.14 | 1.73 | 4.51 | 1.88 | 3.80 | 1.58 |
| 2.6 | 4.13 | 1.59 | 4.53 | 1.74 | 3.89 | 1.50 |
| 2.8 | 4.10 | 1.47 | 4.54 | 1.62 | 3.98 | 1.42 |
| 3.0 | 4.08 | 1.36 | 4.54* | 1.51 | 4.05 | 1.35 |
| 3.5 | 4.00 | 1.14 | 4.52 | 1.29 | 4.21 | 1.20 |
| 4.0 | 3.91 | 0.98 | 4.48 | 1.12 | 4.33 | 1.08 |
| 4.5 | 3.82 | 0.85 | 4.44 | 0.99 | 4.42 | 0.98 |
| 5.0 | 3.74 | 0.75 | 4.39 | 0.88 | 4.50 | 0.90 |
| 100.0 | 2.39 | 0.02 | 3.35 | 0.03 | 5.15* | 0.05 |
A captura máxima sustentável (MSY) aumenta quando Tc aumenta, porque mais peixes sobrevivem a um tamanho maior antes de serem capturados. Da idade 0.2 ano para a idade 1.0 ano a produção de biomassa causada pelo crescimento individual supera as perdas causadas pela mortalidade. Isto, naturalmente, não é válido para valores altos de Tc. Se, por exemplo, Tc fosse maior que o tempo de vida da espécie em questão nenhum peixe, obviamente, seria capturado.
| curva A: | (Tc = 0.2) | MSY/R = 4.15 | (indicado por “*” na tabela) |
| curva B: | (Tc = 0.3) | MSY/R = 4.54 | |
| curva C: | (Tc = 1.0) | MSY/R = 5.15 |
Para F = 1, Y/R é 3.64 (curva A), 3.73 (curva B) ou 2.60 (curva C).
Assim, independentemente do tamanho actual da malha usado, espera-se um crescimento na captura para um aumento do esforço (F).
Quanto menor for o tamanho da malha em uso, menor será o ganho na captura com o aumento do esforço.
Exercício 8.4 Conceito de captura relativa por recruta (Beverton e Holt)
Folha de exercícios 8.4
| E | Lc = 118 cm (Y/R)' | Lc = 150 cm (Y/R)' | (F) |
| 0 | 0 | 0 | 0 |
| 0.1 | 0.019 | 0.022 | 0.020 |
| 0.2 | 0.035 | 0.043 | 0.045 |
| 0.3 | 0.048 | 0.062 | 0.077 |
| 0.4 | 0.059 | 0.079 | 0.120 |
| 0.5 | 0.067 | 0.093 | 0.180 = M |
| 0.6 | 0.071 | 0.105 | 0.270 |
| 0.7 | 0.071*) | 0.112 | 0.42 |
| 0.8 | 0.068 | 0.116 | 0.72 |
| 0.9 | 0.063 | 0.117*) | 1.62 |
| 1.0 | 0.056 | 0.114 | ∞ |
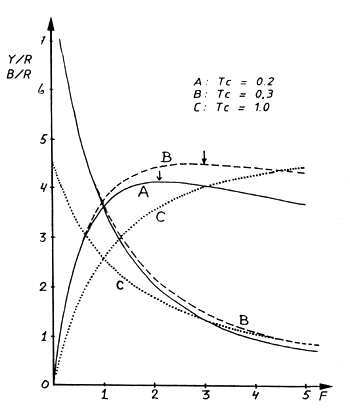 | 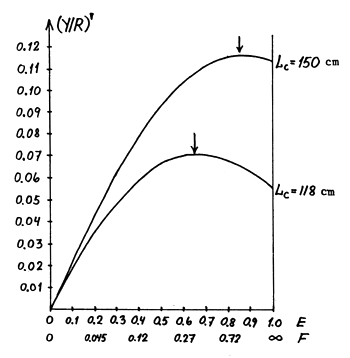 |
| Fig. 18.8.3 Curvas de capturas por recruta e biomassa por recruta em função de F a idades diferentes à primeira captura do peixe-pónei. Fonte de dados: Pauly, 1980. (Ver Folha de exercícios 8.3) | Fig. 18.8.4 Curvas de capturas relativas por recruta em função da taxa de exploração (E) para dois valores diferentes de 50 % do comprimento de retenção do espadarte. Fonte de dados: Berkeley e Houde, 1980. (Ver Folha de exercícios 8.4) |
Exercício 8.6 Modelo de projecção baseado em idade (análise de Thompson e Bell
Folha de exercícios 8.6
a) Não houve alteração no esforço de pesca:
| gr. idade | peso médio (g) | mort. cerco praia | mort. rede emalh. | mort. nat. | mort. total | o manacial | capt. cerco praia | capt. rede emal. | c.peso cerco praia | c.peso rede emal. | capt. peso total |
| t | W | FB | FG | M | Z | N*10-3 | CB | CG | YB | YG | YB+YG |
| 0 | 8 | 0.05 | 0.00 | 2.00 | 2.05 | 1000 | 21.3 | 0 | 170 | 0 | 170 |
| 1 | 283 | 0.40 | 0.00 | 0.80 | 1.20 | 129 | 30.0 | 0 | 8486 | 0 | 8486 |
| 2 | 1155 | 0.10 | 0.19 | 0.30 | 0.59 | 39 | 2.9 | 5.7 | 3383 | 6428 | 9810 |
| 3 | 2406 | 0.01 | 0.59 | 0.20 | 0.80 | 21 | 0.15 | 8.7 | 356 | 21002 | 21358 |
| 4 | 3764 | 0.00 | 0.33 | 0.20 | 0.53 | 9.7 | 0 | 2.5 | 0 | 9312 | 9312 |
| 5 | 5046 | 0.00 | 0.09 | 0.20 | 0.29 | 5.7 | 0 | 0.44 | 0 | 2241 | 2241 |
| 6 | 6164 | 0.00 | 0.02 | 0.20 | 0.22 | 4.3 | 0 | 0.08 | 0 | 471 | 471 |
| 7 | 7090 | 0.00 | 0.00 | 0.20 | 0.20 | 3.4 | 0 | 0 | 0 | 0 | 0 |
| Total | 54.4 | 17.4 | 12395 | 39454 | 51848 |
b) Tempo de defeso da pesca do cerco para a praia:
| gr. idade | peso médio (g) | mort. cerco praia | mort. rede emalh. | mort. nat. | mort. total | o manacial | capt. cerco praia | capt. rede emal. | c.peso cerco praia | c.peso rede emal. | capt. peso total |
| t | W | FB | FG | M | Z | N*10-3 | CB | CG | YB | YG | YB+YG |
| 0 | 8 | 0.00 | 0.00 | 2.00 | 2.00 | 1000 | 0 | 0 | 0 | 0 | 0 |
| 1 | 283 | 0.00 | 0.00 | 0.80 | 0.80 | 135 | 0 | 0 | 0 | 0 | 0 |
| 2 | 1155 | 0.00 | 0.19 | 0.30 | 0.49 | 61 | 0 | 6.9 | 0 | 10550 | 10550 |
| 3 | 2406 | 0.00 | 0.59 | 0.20 | 0.79 | 39 | 0 | 16.0 | 0 | 36560 | 36560 |
| 4 | 3764 | 0.00 | 0.33 | 0.20 | 0.53 | 17.8 | 0 | 4.6 | 0 | 16301 | 16301 |
| 5 | 5046 | 0.00 | 0.09 | 0.20 | 0.29 | 10.5 | 0 | 0.8 | 0 | 3923 | 3923 |
| 6 | 6164 | 0.00 | 0.02 | 0.20 | 0.22 | 7.8 | 0 | 0.14 | 0 | 824 | 824 |
| 7 | 7090 | 0.00 | 0.00 | 0.20 | 0.20 | 6.3 | 0 | 0 | 0 | 0 | 0 |
| Total | 0 | 29.5 | 0 | 68158 | 68158 | ||||||
Embora a captura total tenha aumentado, no caso em que se fechou a pesca do cerco para a praia, não é recomendável fechar esta pescaria sem se levar em conta os aspectos sócio-económicos.
Exercício 8.7 Modelo de projecção baseado em comprimentos (análise de Thompson e Bell)
Folha de exercícios 8.7
| classe compr L1–L2 | F(L1,L2) | N(L1) | biomassa média *Δt | captura C(L1,L2) | captura peso (L1,L2) | rend. (L1,L2) |
| 10–15 | 0.03 | 1000 | 6.47 | 9.94 | 0.19 | 0.19 |
| 15–20 | 0.20 | 890.56 | 17.02 | 63.54 | 3.40 | 3.40 |
| 20–25 | 0.40 | 731.70 | 31.97 | 112.28 | 12.79 | 19.18 |
| 25–30 | 0.70 | 535.20 | 45.18 | 152.08 | 31.62 | 47.44 |
| 30–35 | 0.70 | 317.95 | 50.39 | 102.75 | 35.27 | 70.55 |
| 35–40 | 0.70 | 171.15 | 48.27 | 64.08 | 33.79 | 67.59 |
| 40-L∞ | 0.70 | 79.60 | 61.10 | 55.72 | 42.77 | 85.55 |
| Total | 260.44 | 560.39 | 159.86 | 293.91 |
Exercício 8.7a Modelo de projecção baseado em comprimentos (curva de capturas, análise de Thompson e Bell)
Folha de exercícios 8.7a
| classe compr. | biomassa média | captura | captura peso | rend. | ||
| L1–L2 | F(L1,L2) | N(L1) | *Δt | C(L1,L2) | (L1,L2) | (L1,L2) |
| 10–15 | 0.06 | 1000 | 6.44 | 19.79 | 0.38 | 0.38 |
| 15–20 | 0.40 | 881.22 | 16.25 | 121.30 | 6.50 | 6.50 |
| 20–25 | 0.80 | 668.94 | 27.08 | 190.22 | 21.66 | 32.50 |
| 25–30 | 1.40 | 407.39 | 29.97 | 201.75 | 41.95 | 62.93 |
| 30–35 | 1.40 | 162.40 | 22.02 | 89.80 | 30.82 | 61.65 |
| 35–40 | 1.40 | 53.36 | 12.56 | 33.36 | 17.59 | 35.19 |
| 40-L∞ | 1.40 | 12.84 | 5.80 | 10.57 | 8.12 | 16.24 |
| Total | 120.13 | 666.79 | 127.05 | 215.41 |
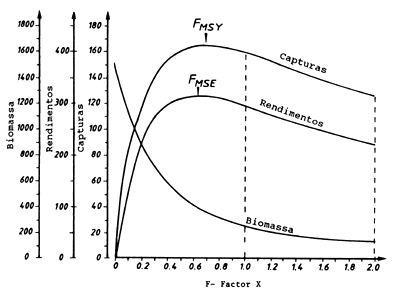
Fig. 18.8.7a Análise de Thompson e Bell, estimação da biomassa média, das capturas e rendimentos (os valores de X = 1 e X = 2
Exercício 9.1 Modelo de Schaefer e modelo de Fox *
Folha de exercícios 9.1
| ano i | capturas (tons. s/cabeça) Y(i) | esforço (barco/dias) f(i) (x) | SCHAEFER Y/f (y) | FOX ln(Y/f) (y) |
| 1969 | 546.7 | 1224 | 447 | 6.103 |
| 1970 | 812.4 | 2202 | 369 | 5.911 |
| 1971 | 2493.3 | 6684 | 373 | 5.922 |
| 1972 | 4358.6 | 12418 | 351 | 5.861 |
| 1973 | 6891.5 | 16019 | 430 | 6.064 |
| 1974 | 6532.0 | 21552 | 303 | 5.714 |
| 1975 | 4737.1 | 24570 | 193 | 5.263 |
| 1976 | 5567.4 | 29441 | 189 | 5.242 |
| 1977 | 5687.7 | 28575 | 199 | 5.293 |
| 1978 | 5984.0 | 30172 | 198 | 5.288 |
| valor médio | 17286 | 305.2 | 5.666 | |
| desvio padrão | 11233 | 102.9 | 0.3558 | |
| intersecção (Schaefer: a, Fox: c) | 444.6 | 6.1508 | ||
| declive (Schaefer: b, Fox: d) | -0.008065 | -0.000028043 | ||
| variância do declive: sb2 = ((sy/sx)2-b2) / (10-2) | 2.361*10-6 | 2.7113*10-11 |
| desvio padrão do declive, sb | 0.0015364 | 0.000005207 |
| distribuição de Student t(10-2) | ||
| limites de confiança do declive: | ||
| b+t(n-2)*sb superior | -0.0045 | -0.00001601 |
| b-t(n-2)*sb inferior | -0.0116 | -0.00004007 |
| variânçia do ponto de intersecção: sa2 = sb2*[sx2*(n-1)/n+x2] | 973.4 | 0.01152 |
| desvio padrão do ponto de intersecção: | 31.20 | 0.1073 |
| limites de confiança do ponto de intersecção: | ||
| a+t(n-2)*sa superior | 517 | 6.40 |
| a-t(n-2)*sa inferior | 372 | 5.90 |
| MSY Schaefer: -a2/(4b) | 6128 tons. | |
| MSY Fox : -(1/d)*exp(c-1) | 6154 tons. | |
| f(MSY) Schaefer: -a/(2b) | 27565 | |
| f(MSY) Fox: -1/d | barco/dias | 35660 |
| barco/dias |
*) a,b foram substituidos por c,d no modelo Fox
Folha de exercícios 9.1a
| esforço barco/dias | capturas de equilíbrio (toneladas) | |||||
| f | Schaefer | Fox | ||||
| 5000 | 2021 | 2039 | ||||
| 10000 | 3640 | 3544 | ||||
| 15000 | 4854 | 4620 | ||||
| 20000 | 5666 | 5354 | ||||
| 25000 | 6074 | 5817 | ||||
| 27565 | 6128 | = | MSY | |||
| 30000 | 6080 | 6068 | ||||
| 35000 | 5681 | 6153 | ||||
| 35660 | 6154 | = | MSY | |||
| 40000 | 4880 | 6112 | ||||
| 45000 | 3675 | 5976 | ||||
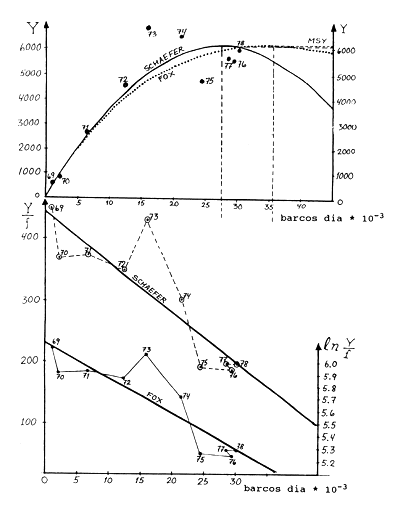
Fig. 18.9.1 Representação combinada dos modelos de Schaefer e Fox para uma pescaria de camarão. Em cima: Capturas contra esforço. Em baixo: CPUE e respectivamente ln CPUE contra esforço. Fonte de dados: Naamin e Noer, 1980. (Ver Folha de exercícios 9.1 e 9.1a)
Exercício 13.8 Método da área varrida, precisão da biomassa estimada, estimação do MSY e repartição óptima dos lances
Folha de exercícios 13.8
ESTRATO I:
| CPUE | BARCO | ARRASTO | CORRENTE | DIST | ÁREA | CPUA | |||
| lance núm. i | Cw/t kg/h | veloc. nós VS | rumo graus dirV | abert. boca(m) R*x2 | veloc. nós CS | dir. graus dirC | nm. D | varrida sq.nm. a | Cw/a = Ca kg/sq.nm. Ca |
| 1 | 7.0 | 2.8 | 220 | 18 | 0.5 | 90 | 2.508 | .02438 | 287.2 |
| 2 | 7.0 | 3.0 | 210 | 16 | 0.5 | 180 | 3.442 | .02974 | 235.4 |
| 3 | 5.0 | 3.0 | 200 | 17 | 0.3 | 135 | 3.139 | .02881 | 173.6 |
| 4 | 4.0 | 3.0 | 180 | 18 | 0.4 | 230 | 3.271 | .03180 | 125.8 |
| 5 | 1.0 | 3.0 | 90 | 17 | 0.5 | 270 | 2.500 | .02295 | 43.6 |
| 6 | 4.0 | 3.0 | 45 | 18 | 0.4 | 160 | 2.854 | .02774 | 144.2 |
| 7 | 9.0 | 3.5 | 25 | 18 | 0.4 | 200 | 3.102 | .03015 | 298.5 |
| 8 | 0.0 | 3.0 | 210 | 18 | 0.3 | 300 | 3.015 | .02930 | 0.0 |
| 9 | 0.0 | 3.5 | 0 | 18 | 0.4 | 0 | 3.900 | .03790 | 0.0 |
| 10 | 14.0 | 2.8 | 45 | 18 | 0.6 | 0 | 3.252 | .03161 | 442.9 |
| 11 | 8.0 | 3.0 | 120 | 18 | 0.3 | 300 | 2.700 | .02624 | 304.9 |
| Ca = 186.9 | |||||||||
ESTRATO II:
| CPUE | BARCO | ARRASTO | CORRENTE | DIST | ÁREA | CPUA | |||
| lance núm. i | Cw/t Kg/h | veloc. nós VS | rumo graus dirV | abert. boca(m) R*x2 | veloc. nós CS | dir. graus dirC | nm. D | varrida sq.nm. a | Cw/a = Ca kg/sq.nm. Ca |
| 12 | 42.0 | 4.0 | 30 | 17 | 0.5 | 160 | 3.698 | .03395 | 1237.1 |
| 13 | 98.0 | 3.3 | 215 | 17 | 0.4 | 90 | 3.088 | .02835 | 3457.3 |
| 14 | 223.0 | 3.9 | 30 | 17 | 0.0 | 0 | 3.900 | .03580 | 6229.2 |
| 15 | 59.0 | 3.8 | 35 | 17 | 0.3 | 180 | 3.558 | .03266 | 1806.3 |
| 16 | 32.0 | 3.5 | 210 | 17 | 0.5 | 270 | 3.775 | .03465 | 923.5 |
| 17 | 6.0 | 2.8 | 210 | 17 | 0.5 | 330 | 2.587 | .02374 | 252.7 |
| 18 | 66.0 | 3.8 | 45 | 17 | 0.5 | 30 | 4.285 | .03933 | 1678.0 |
| 19 | 60.0 | 4.0 | 30 | 18 | 0.5 | 180 | 3.576 | .03475 | 1726.5 |
| 20 | 48.0 | 4.0 | 210 | 18 | 0.5 | 180 | 4.440 | .04315 | 1112.3 |
| 21 | 52.0 | 3.8 | 20 | 18 | 0.4 | 180 | 3.427 | .03331 | 1561.3 |
| 22 | 48.0 | 4.0 | 30 | 18 | 0.5 | 190 | 3.534 | .03435 | 1397.4 |
| 23 | 18.0 | 3.0 | 210 | 18 | 0.3 | 190 | 3.284 | .03192 | 563.9 |
| Ca = 1828.8 | |||||||||
| limites de confiança de Ca: | ||||||
| estrato | número de lances n | Ca | s | s/√ n | distr. Student t(n-1) | limites de confiança de Ca |
| 1 | 11 | 186.9 | 141.6 | 42.7 | 2.23 | [ 92, 282] |
| 2 | 12 | 1828.8 | 1597.5 | 461.2 | 2.20 | [814, 2843] |
Biomassa média da área total:
Área dos estratos 1 e 2 combinados A = A1+A2 = 24+53 = 77 sq. nm.
Ca(A) = (24*186.9+53*1828.8)/77 = 1317.0 kg/sq.nm. (cf. Eq. 13.7.5)
Biomassa total para toda a área: B(A) = 1317.0*77/0.5 = 202818 kg,
digamos 203 toneladas
Da Eq. 9.2.1: MSY = 0.5*0.6*203 = 61 toneladas/ano.
Folha de exercícios 13.8a (para traçar o gráfico de erro máximo relativo)
| número de lances n | t(n-1) | estrato I ε *) | estrato II ε **) |
| 5 | 2.78 | 0.94 | 1.09 |
| 10 | 2.26 | 0.54 | 0.62 |
| 20 | 2.09 | 0.36 | 0.41 |
| 50 | 2.01 | 0.22 | 0.25 |
| 100 | 1.98 | 0.15 | 0.17 |
| 200 | 1.97 | 0.11 | 0.12 |
a) 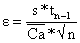 com s = 141.6 e Ca= 186.9
com s = 141.6 e Ca= 186.9
b) com s = 1597.5 e Ca = 1828.8
Folha de exercícios 13.8b (repartição óptima)
| estrato | s de Ca s | A | A*s | A*s/∑A*s | 200*A*s/∑A*s |
| 1 | 141.6 | 24 | 3398 | 0.039 | 8 lances |
| 2 | 1597.5 | 53 | 84670 | 0.961 | 192 lances |
| Total | 88068 | 200 lances |
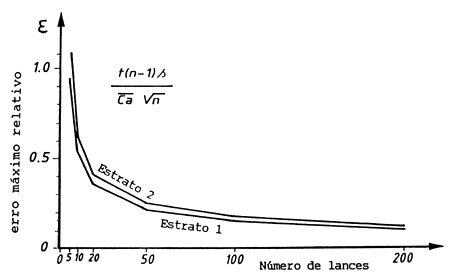
Fig. 18.13.8 Erro máximo relativo na captura média por área, do roncador de manchas pequenas, contra o número de lances de arrasto. Linha superior: estrato 2; linha em baixo: estrato 1. Fonte de dados: Project KEN/74/023. (ver Folha de exercícios 13.8a)

![]()
![]()