![]()
![]()
![]()
Floodplain fish stocks display a characteristic response to increasing fishing effort. As illustrated in Part 1, total catches may be maintained as fishing increases, but species compositions deteriorate as the large and valuable species become over-exploited. This section compares data from the Fisheries Dynamics project to illustrate the effect of increasing fishing effort on floodplain fish catches.
Fisherman densities at the project sites were approximately an order of magnitude greater in Bangladesh than in Indonesia. Fisherman densities in Bangladesh were estimated at 47 and 36/km2 (high water flooded areas) respectively in the Inside and Outside regions, not including subsistence fishermen (MRAG, 1997). Fisherman densities in Indonesia can not be so accurately estimated due to the less well known high water flooded area of the study site. Assuming that flood waters cover approximately 100km2 within the River Lempuing study area (MRAG, 1997), licensed fishermen were present at densities of approximately 3.5 and 3.2/km2 in 1995 and '96 respectively, well below those in Bangladesh.
Reflecting the active nature of their fishing gears (see Section 4.1), fishermen in Bangladesh also work significantly longer hours than those in Indonesia (Table 3.7). For those respondent fishermen actively engaged in fishing in each month, daily fishing hours were almost three times longer in Bangladesh (11.1 hours) than in Indonesia (4.0 hours). Taking non-fishing respondents into account, average fishing hours in Bangladesh (4.7) were still almost twice those Indonesia (2.7).
With ten times as many fishermen in Bangladesh, each of them working significantly longer hours than in Indonesia, the total annual fish catches per individual fishermen in Bangladesh are 6–16 times lower than in Indonesia (Table 3.7). Allowing for the differences in fishing practices in the two countries, and the relative daily hours of active work, fishing in Bangladesh is between 11 and 28 times less productive per hour than in Indonesia. These strong differences in the numbers of fishers and their fishing practices are well reflected by the much increased fishing mortality rates found in Bangladesh (Section 3.2.5). In contrast, however, the two fisheries still produced similar total catches per unit area (51–130kg/ha/yr in Bangladesh, and approximately 72–118kg/ha/yr in Indonesia). As shown in the following sub-section, much more of the Bangladeshi catch is comprised of small fish species.
Table 3.7 Comparison of fishing and catch rates at the R5953 Bangladesh and Indonesian study sites (summarised from MRAG, 1997)
| Bangladesh | Indonesia | |||
| Fisherman density (no / km2) | 36–47 | 3–4 | ||
| Fishing activity (hrs / day) | 11 | 4 | ||
| Catch / fisherman (t / yr) | 0.2–0.4 | 2.2–3.3 | ||
| Catch / area (kg / ha / yr) | 51–130 | 72–118 | ||
Taking a broader perspective, the results presented here for the two R5953 study sites are supported by comparable data from the R4791 project and from other sites around the world. Asian floodplain river fisheries are clearly among the most heavily exploited and the most productive in the world (Figure 3.10). Much lower productivities are found in South America, where population densities and fishing efforts are also low. The high catches in Hail Haor are the result of fishermen densities as much as a hundred times higher than elsewhere. Comparing the project sites, fishermen densities in Hail Haor are four and seven times higher than in the Lempuing and Thale Noi respectively. Reflecting the R5953 results, the catch-per-fisherman declines as fishing effort increases, from a maximum of over 10t per year in the most lightly fished sites, down to as little as 128kg in Bangladesh (MRAG, 1994).
While individual catches are thus low in the Bangladesh Hail Haor site, the total catch remains as high as those elsewhere (Figure 3.10), and it is not certain that it should be classed as overexploited, even with 30 fishermen/km2. Significantly lower catches-per-unit-area were, in contrast, taken from the western Bangladesh beels with the highest fishing efforts of any floodplain fishery. This point suggests that fish yields may begin to decline at very high effort levels, though some catches may still be maintained.
Regardless of the exact shape of the surplus yield curve, it is evident from the data that most floodplain fisheries including the Sumatran and Thai sites are relatively lightly to moderately exploited, and still produce reasonably good catches. The Hail Haor fishery in contrast is heavily exploited, and its high fish catches could almost certainly be taken with far less effort.
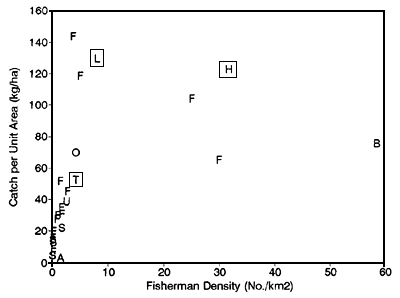
| Code | Location | Source |
| H | Hail Haor, Bangladesh | R4791 Project study site |
| L | Lempuing River, Indonesia | R4791 Project study site |
| T | Thale Noi, Thailand | R4791 Project study site |
| B | Western Bangladesh Beels | BCAS, 1992 |
| U | Lubuk Lampam, Indonesia | RIFF, Palembang, Sumatra |
| O | OKI regency, Indonesia | Fisheries Service, Palembang, Sumatra |
| A | Sepik River, Papua New Guinea | MRAG, 1993 |
| S | South American Rivers | MRAG, 1993 |
| F | African Rivers | Bayley, 1988 |
Figure 3.10 Estimated total catches scaled by high water floodplain area, plotted against fisherman densities for the three study sites and other tropical floodplain fisheries in Asia, Africa and South America (from Hoggarth and Kirkwood, 1996).
The impact of fishing on the structure and composition of fish stocks was also investigated at R4791 study sites in Bangladesh, Thailand and Indonesia. All three fish stocks are exploited by a wide range of fishing gears including seine nets, gill nets, individual fish traps, barrier traps, hooks and other less familiar approaches. The relative importance of the different fishing gears at each site depends on their morphological and hydrological characteristics. At the most riverine Indonesian site, exploitation is notable for the extensive use of efficient barrier traps (Figure 3.11), catching fish as they migrate around the floodplain and river system. In the more lacustrine Thai site, gill nets take more than 50% of the total catch, being most effective in the calm waters of the lake environment. In the dry seasons, fish in Bangladesh and Thailand are caught by de-watering isolated ponds with diesel pumps. In Indonesia, such fish are caught by seining enclosed ponds or river sections up to 1km in length with nets and sections of bamboo fences.
Similar mesh sizes are used in both the Indonesian and Thai fisheries, but all gears at the Bangladesh site are particularly small meshed (Figure 3.11). Reflecting the high fisherman densities and fishing activity levels in Bangladesh (previous section), fishermen have been forced to reduce mesh sizes to maintain their catches. As a result of these gear characteristics, the catches in Bangladesh are now comprised of many more small fish (Figure 3.11) with even the largest species often being caught as fry of only 2–3cm in length (see Figure 3.6 for the large catfish Wallago attu).
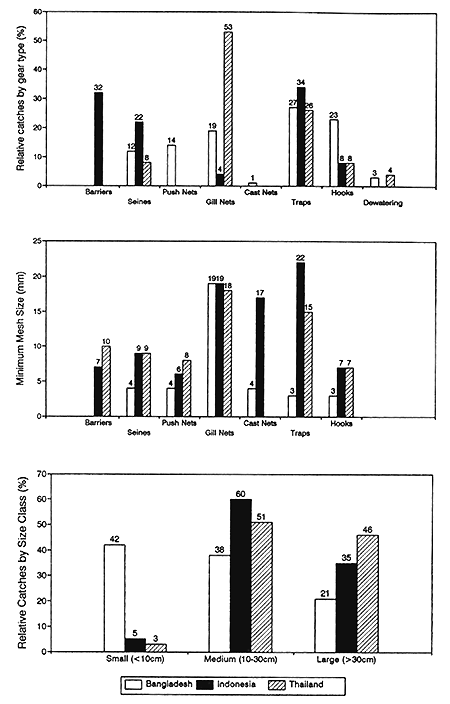
Figure 3.11 Between site comparisons of the relative catches and mesh sizes of the most important gear types, and the catches of fish species with maximum lengths in three different size classes (from Hoggarth and Kirkwood, 1996).
The PIRDP flood control, drainage and irrigation scheme in Bangladesh delays and smoothens high water flooding (Figure 2.8, Table 2.3), thereby providing benefits for both floodplain communities and agricultural production (Section 2.3.2). Against these benefits, the reduction in flooding and the compartmentalisation of the floodplain has raised concerns (e.g. IUCN, 1991; Majumder and Durante, 1993) that flood control schemes may have significant negative impacts on floodplain fisheries, on which many of Bangladesh's landless poor depend. These concerns illustrate the need for an integrated catchment management approach for floodplain fisheries, as promoted in Part 1.
To demonstrate the potential interactions between fisheries and other sectors, this section investigates the impact of the Bangladesh PIRDP flood control scheme on its fish stocks and fisheries. These investigations were carried out under project R5953 (MRAG, 1997; Halls, 1998), and are presented in four sections:
Firstly, the impact of the PIRDP on the potential productivity of fish stocks is investigated by comparing growth, reproduction and mortality parameters inside and outside the flood control scheme. It is shown that the productive potential of individual fish inside the PIRDP is at least as high as those outside.
Secondly, the accessibility of the PIRDP is investigated from the tagging studies and from records of fish catches taken at sluice gates. This work showed that some fish are able to migrate both in and out of the PIRDP through the sluice gates during four distinct migration seasons.
Thirdly, the fish species compositions found inside and outside the PIRDP and other flood control schemes in Bangladesh are investigated. It is shown that fish species compositions are indeed different inside and outside flood control schemes, with inside stocks comprising fewer fish species, and having less of the large, valuable, migratory ‘whitefish’ species.
Finally, the overall productivity of the inside and outside regions is compared, measured as their fish catches per unit area. Catch rates outside the PIRDP are shown to be 60–104% greater than those inside. Since the production potential of individual fish is not reduced by the PIRDP, but species compositions are depleted, it is concluded that the reduction in catches is due to the partial restriction on fish movements imposed by the flood control embankment.
The impact of floodplain modification on the potential productivity of individual fish was investigated by comparisons of growth rates, reproductive parameters and mortality rates inside and outside the PIRDP. The combination of these parameters determine the mean weight achieved by a given fish in its lifetime and its potential contribution to spawning activities. In all cases, these indicators of productivity were either not significantly different inside and outside the scheme, or were better inside.
Comparison of growth performance
Due to the previously mentioned difficulties in fitting growth curves, relative growth performance was compared using the observed mean length of the ‘0+ aged’ cohort in November (LNov). November samples were selected for two reasons: firstly, most of the year's growth was achieved by this month (see e.g. the length frequency for W. attu in Figure 3.6), and secondly, two hydrological years (95/96 and 96/97) could be compared for which flood index and abundance data were also available to assist interpretation (Halls, 1998). The estimates of LNov for the two sampling locations and for the two hydrological years were compared using two-way analysis of variance (ANOVA). Length-weight relationship parameters were also estimated for each species and sampling location to enable comparisons of ‘condition factors’ (Wootton 1990).
These growth comparisons are presented in detail in Halls, 1998; MRAG, 1997 and Halls et al, in press (a). In summary, LNov was, with the exception of G. giuris, found to be significantly higher during the 95/96 flood year compared to the 96/97 year. Mean lengths ranged from 4% larger for P.sophore to 23% larger for W. attu. During this flood year, growth performance of four of the six species was also significantly higher inside the PIRDP scheme than outside. These differences in performance were comparable with those between the two flood years, ranging from 7% for P.sophore to 18% for W. attu. In contrast, Catla catla grew 6% larger outside the scheme compared to inside and A.testudineus showed no significant difference in growth in the two locations.
The condition (weight at length) of C. catla, G.giuris, P.sophore and W.attu was also found to be significantly higher inside the scheme compared to outside, for the 24 month sampling period. No significant differences in condition were found for either A.testudineus and C.striatus (Halls, 1998).
In an attempt to explain these observed differences, it should first be noted that none of the key species showed any significant differences in diet composition in the two sampling locations. Feeding intensity, measured as the mean monthly stomach fullness index (SFI), was also found to be not significantly different inside and outside the PIRDP FCDI scheme for any of the key species. There is some qualitative evidence that suggests that feeding intensity was, however, greater during the 95/96 flood year compared with 96/97, eg for A.testudineus and G.giurus, though W. attu fed better in 96/97 (MRAG, 1997; Halls, 1998).
Relative in/out abundances were also examined for the key species between the two flood seasons (MRAG, 1997; Halls, 1998), and found to be higher outside in the 95/96 flood year for C.catla, G. giurus, P.sophore and W. attu and higher inside in the 96/97 flood year for A. testudineus. Abundances were not significantly different for other species/year combinations.
The growth performance measures of the key species and the potential explanatory variables that were found to be significantly different in relation to the sampling location and hydrological year are summarised in Table 3.8 below. Inter-annual and spatial variation in flooding (see bottom of table) explains the larger mean lengths (LNov) found in 95/96 for five of the six key species, but fails to explain, with the exception of C.catla, the larger mean lengths found inside the FCDI scheme. The latter are much better explained by CPUE (a reflection of fish density), which also explains all the observed differences in condition. The stomach fullness indices (SFIs) also correspond with the larger flood of 95/96, further explaining four of the larger mean lengths for this year.
These results imply that both the magnitude of the flood and fish density are important factors in determining the growth performance of the key species, while stomach fullness is more likely to reflect the effects of these two factors (Halls et al, in press (a)).
Table 3.8 Summary table indicating where the growth performance measures and explanatory variables were found to be significantly greater in relation to the sampling location and hydrological year (NS = no significant difference; Halls et al, in press (a)).
| Growth performance variables | Explanatory variables* | |||||
|---|---|---|---|---|---|---|
| LNov | Condition | CPUE | SFI | |||
| Species | Year | Location | IN/OUT | 95/96 | 96/97 | Year |
| A.testudineus | 95/96 | NS | NS | NS | IN | 95/96 |
| C.catla | 95/96 | OUT | IN | OUT | NS | 95/96 |
| C.striatus | 95/96 | IN | NS | NS | NS | 95/96 |
| G.giuris | NS | IN | IN | OUT | NS | 95/96 |
| P.sophore | 95/96 | IN | IN | OUT | NS | NS |
| W.attu | 95/96 | IN | IN | OUT | NS | 96/97 |
* Flood index : 95/96 > 96/97 IN < OUT in both years
Contrary to what might be expected, then, given the reduced flooding inside the PIRDP, the growth performance of the key species has been shown to be significantly higher in this location. This greater growth performance appears to be due to the lower levels of abundance inside the scheme, implying that intraspecific competition is an important component of floodplain fish growth (Halls, 1998).
Comparison of reproductive parameters
None of the reproductive parameters investigated showed any strong differences inside and outside the PIRDP scheme. Firstly, in contrast to the large differences in size at maturity between the sexes (Section 3.2.4), there was no strong evidence to suggest that Lm50 was significantly different inside the PIRDP scheme for any of the key species (Figure 3.7, Table 3.1, Halls et al, in press (a)).
Secondly, the coefficients (slopes) of the fecundity at length relationships were also not significantly different between the two sampling locations for P.sophore, C.striatus or G.giuris: common regression models were thus fitted for these species (Halls et al, in press (a)). For A.testudineus, the slope was significantly higher, implying that individuals were more fecund at length inside the PIRDP.
Thirdly, seasonal changes in mean monthly GSI for each key species and sampling location were given in Figure 3.8. Again, there was no evidence to suggest that spawning time was significantly different inside and outside the PIRDP for any of the key species (Halls et al, in press (a)).
Comparison of mortality rates
As mentioned in Section 3.2.5, mean total mortality rate (Z) estimates, for each species were slightly higher outside the PIRDP scheme compared to inside, though not significantly so in any case (Table 3.2). These results further support the conclusions of the growth rate studies that conditions for fish production inside the PIRDP are more positively influenced by the low fish densities, than they are negatively influenced by the reduced flooding.
The previous section showed that the ‘inside’ PIRDP floodplains are potentially as productive for individual fish as those outside. The overall productivity of the PIRDP modified floodplains are thus dependent on the actual numbers of fish present inside the scheme, and its accessibility from outside. This section firstly investigates the attempts of fish to migrate in and out of the PIRDP, from records of catches at its flood control sluice gates, and secondly confirms that some fish are indeed able to pass through the gates. The following section shows that fish species compositions are nevertheless different between the two regions, indicating that the PIRDP embankment does reduce the accessibility of its impounded floodplains.
Studies of fish catches at PIRDP sluice gates
The potential movements of fish into the PIRDP from outside were examined by investigating the catches of fishing gears at the PIRDP sluice gates. For this purpose, those ‘interceptory’ fishing gears operating within a 100m radius of the sluice gates were monitored during the five months (July to November) of the 1996 flood season. These fishing gears include lift nets (veshal or khora jal); bag nets (suti jal) and jump nets (urani) deliberately positioned to take advantage of the movements of fish towards or through the sluice gates. The orientation or placement of such gears was used to determine the directional behaviour of the fish migrations.
Catches were monitored at two sluice gates: the PIRDP's largest, 6-panel, 94.6m2-aperture gate on the Badai river at Talimnagar, and the smaller, 3-panel, 8.1m2 gate on the Natuabari canal at Baulikhola, 8km northwards along the embankment (Figure 2.2). At each sluice gate, daily recordings were made of both hydrological information (in / out water heights, sluice apertures and flow directions) and fishing effort and catches by gear type and species (see further details in MRAG, 1997 and Hoggarth et al, in press (b)).
The daily operation of the sluice gates, and their impacts on water levels inside the PIRDP was described in Section 2.3.2. The height of the sluice gate sills means that water may only flow in or out of the PIRDP during the high water season, approximately May to November (Figure 2.8). During the dry season months, both gates completely dry out. In 1996, the flood started early on 23 March, but did not reach the floodplain height until much later at the end of June. Outside water levels then rose until mid/late July and gradually fell until the end of November (Figure 2.8). Due to the low water levels, the sluice gates were partly or fully open on all days up until mid August, thereby allowing water to pass into the PIRDP scheme. The gates were closed (though leaking) over the higher water period, and then opened again to allow water out from early/mid September.
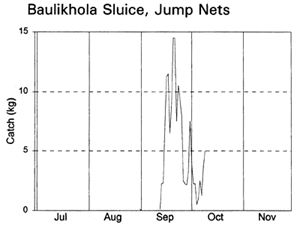
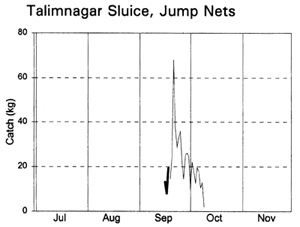
Fishing activities at the two sluice gates depended on their size and fishing opportunities. Lift nets and bag nets were fully utilised at the small Baulikhola gate, but not at the larger Talimnagar gate due to the strong River Badai currents. Jumping traps caught fish attracted towards the sluice gates by the outflowing water at the start of the ebb season, while the expensive bag net gear caught fish emigrating from the PIRDP at the end of the ebb (MRAG, 1997; Hoggarth et al, in press (b)). Lift nets were set in various positions on either side of the sluice gates to catch fish moving in both directions in different seasons.
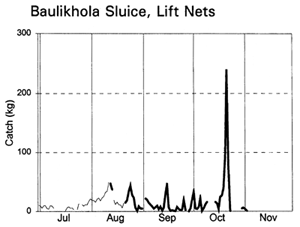
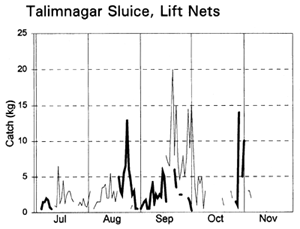
Migrating fish displayed four different migration phases, including both passive and active responses (with and against the currents) in both inward and outward directions (Table 3.9, based on data in Figure 3.12). During the early flood, small fish, such as Puntius sophore, Glossogobius giurus and shrimps drifted in passively with the current. Towards the end of the flood season, when the gates were closed but leaking, some of the same species were then attracted towards the gates and moved back out of the system. During the early ebb season, the major carps Labeo rohita, Catla catla, Cirrhinus reba, and Labeo calbasu, in addition to the predatory catfish Wallago attu were attracted in to the PIRDP, while other small species migrated out. The largest migration, including virtually all the species previously recorded, was a passive drift out with the ebb, particularly coinciding with the final fall in water levels below the floodplain height.
The largest catches of migrating fish were taken during the late ebb passive emigrations at the small Baulikhola gate (Table 3.10). Surprisingly, more fish were caught trying to migrate against the flows in to the PIRDP after high water, than passively at the beginning of the flood. Due to the difficulty of fishing the large Talimnagar gate, though, it was concluded that many fish both entered and escaped from the PIRDP waters through this route, without capture. The weight of young fish migrating in to the PIRDP during the early flood was roughly estimated between 0.9 and 4 tonnes, mostly through the Talimnagar gate, compared to the 0.6t catch from the existing fishing gears (Hoggarth et al, in press (b)).
Table 3.9 Seasonal fish migration phases at the PIRDP's Talimnagar and Baulikhola sluice gates during the 1996 flood season, summarising catch data in Figure 3.12 (from Hoggarth et al, in press (b)).
| Season | Migration Response | Migration Direction | Gate Aperture | Fishing Gears1 | |
|---|---|---|---|---|---|
| Early Flood | Passive | In | Partly/fully open | Out-facing LNs | |
| July – mid Aug. | (with flow) | ||||
| Late Flood | Active | Out | Closed, leaking | In-facing LNs | |
| mid Aug. – mid Sep. | (against flow) | ||||
| Early Ebb | Active | In | Partly open | Out-facing LNs in side eddies and JTs | |
| mid Sep. – mid Oct. | (against flow) | ||||
| Late Ebb | Passive | Out | Partly/fully open | In-facing LNs, BN and JTs | |
| mid Oct. – Nov. | (with flow) | ||||
1 LN = lift net, JT = jumping trap, BN = bag net.
To promote high catches inside the PIRDP, it was recommended that these flood control sluice gates should always be opened as long as possible in the early flood season, whenever inside water levels are low enough to allow water into the PIRDP without threatening crop production or livelihoods. Encouraging restraints on fishing activities around sluice gates at this time also has potential for increasing productivity.
 |  |
 |  |
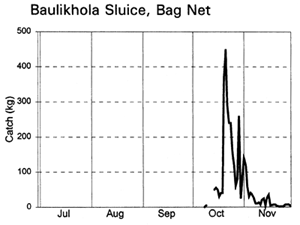 | 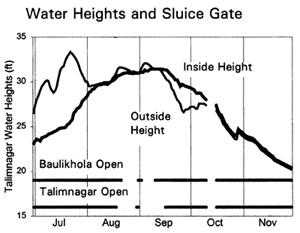 |
Figure 3.12 Total daily catches (kg) of inwardly migrating fish (thin lines) and outwardly migrating fish (thick lines), by the three interceptory gear types at the PIRDP's Baulikhola and Talimnagar sluice gates during the 1996 flood season. Lower right graph shows daily water heights and periods of sluice gate opening, for comparison (data missing in mid-October; from Hoggarth et al, in press (b)).
Table 3.10 Total catches (kg) at the Baulikhola and Talimnagar sluice gates, subdivided by migration phase and gear type (from Hoggarth et al, in press (b)).
| Migration Phase | Baulikhola sluice gate (small) | Talimnagar sluice gate(large) | Total catch (kg) | ||||
|---|---|---|---|---|---|---|---|
| Lift Nets | Jump Traps | Bag Nets | Lift Nets | Jump Traps | Bag Nets | ||
| Early Flood Passive Immigration | 582 | 79 | 661 | ||||
| Late Flood Active Emigration | 371 | 76 | 446 | ||||
| Early Ebb Active Immigration | 16 | 154 | 206 | 479 | 855 | ||
| Late Ebb Passive Emigration | 824 | 2 | 3307 | 71 | 41 | 4244 | |
Tagging studies of migrations in and out of the PIRDP
Of the 1,389 tagged fish recaptured, 14 C. catla, 6 C. striatus and 15 W. attu were released and then recaptured on opposite sides of the PIRDP embankment. Of these fish, 23 migrated into the PIRDP while 12 migrated outwards (Table 3.11, Figure 3.2). Though the numbers of fish are small, these data confirm the ability of these species to actually penetrate the sluice gates in both directions. The other three key species, A. testudineus, G. giuris and P. sophore were not observed to cross the embankment, though they were caught in the catches at the sluice gates.
Based on the release and recapture locations, it is likely that both C. catla and C. striatus passed through both the main 94.6m2 aperture Talimnagar sluice gate on the 50m wide Badai River, and also through the smaller 8.1m2 Baulikhola sluice gate on the narrow Natuabari Canal (Figure 2.2). Migrations across the embankment by W. attu were only seen in the vicinity of the Talimnagar gate.
Table 3.11 Numbers of tagged fish released and recaptured, by species, and the numbers which penetrated the PIRDP sluice gates, during 1995/96 (from Halls et al, in press (b).
| Species | Number released | Number recaptured | Number entering PIRDP | Number exiting PIRDP | % of recaptures moving in or out of PIRDP |
|---|---|---|---|---|---|
| A. testudineus | 902 | 356 | 0 | 0 | 0.0% |
| C. catla | 809 | 408 | 12 | 2 | 3.4% |
| C. striatus | 604 | 255 | 2 | 4 | 2.4% |
| G. giuris | 709 | 88 | 0 | 0 | 0.0% |
| P. sophore | 1013 | 72 | 0 | 0 | 0.0% |
| W. attu | 581 | 210 | 9 | 6 | 7.1% |
The impact of flood control, drainage and irrigation (FCDI) schemes on species assemblages was examined using species abundance data (small meshed seine net catch per unit effort during 1993) sampled by FAP17 (1995) at 74 sites located inside and adjacent to seven FCDI schemes in the following four districts of Bangladesh; Pabna, north-west (NW, including the PIRDP region), Tangail, north-central (NC), Sylhet, north-east (NE) and Faridpur, south-west (SW) (Halls et al, in press (b)). These FAP17 study sites covered a broad range of habitat types including main and secondary rivers, canals and floodplains/beels (natural depressions or lakes upon the floodplain).
Non-parametric multidimensional scaling (MDS) was used to analyse the species abundance data (Clarke, 1993). The approach aims to construct a map or ordination of sites (samples) such that their placement reflects the rank similarity of their species assemblages. Sites positioned in close proximity to each other in the ordination have very similar species assemblages whilst sites that are far apart share few common species or have the same species but at very different levels of abundance. The null hypothesis (Ho: there are no differences in species assemblages at sites inside and outside the FCDI scheme (s)) was tested using a non-parametric ‘permutation’ (analysis of similarity or ANOSIM) test (see Halls et al, in press (b)). The species most responsible for site groupings were then determined by computing the average contribution of each species to the overall average dissimilarity between all pairs of inter-group sites using the ‘indicator species analysis’ approach (Clarke and Warwick, 1994).
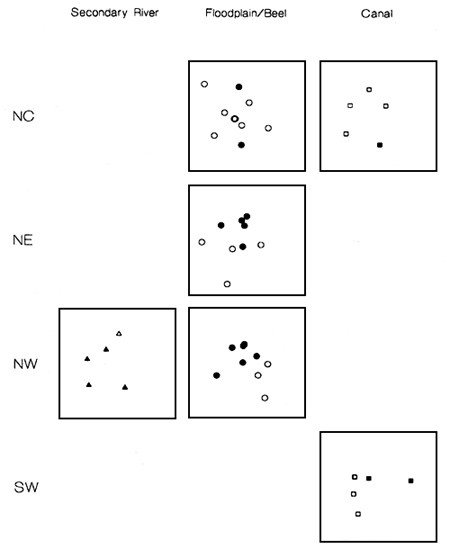
Figure 3.13 MDS ordinations comparing species assemblages inside (black symbols) and outside (open symbols) FCDI schemes in three habitats and four regions in Bangladesh (from Halls et al, in press (b)).
Differences among species assemblages sampled from sites inside and outside FCDI schemes were examined separately for each combination of region and habitat (Figure 3.13). Sufficient replicates were only available to test the null hypothesis (Ho: there are no differences in species assemblages at sites inside and outside the FCDI schemes) for the floodplain/beel habitat in the NE and NW regions (Halls et al, in press (b)). In both cases the null hypothesis was rejected (P<5%). Examination of the remaining ordinations reveals statistically untestable, but visually convincing evidence that species assemblages from canal and secondary river habitat are also different inside and outside FCDI schemes in those regions which could be considered.
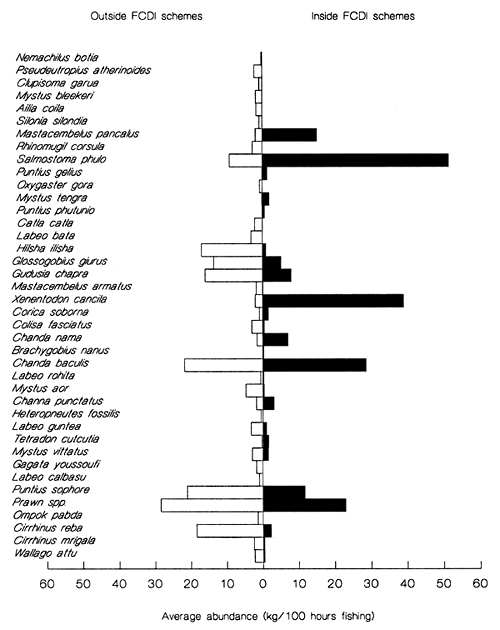
Figure 3.14 Average abundance (seine net CPUE) of species sampled from inside (solid bars) and outside (open bars) FCDI schemes in the NW region. Species are arranged from top to bottom in descending order of their contribution to the average dissimilarity between the two groups of sites (from Halls et al, in press (b)).
For floodplain/beel habitats in the NW region, including the PIRDP scheme, the indicator species analysis revealed that 25 species were absent or less abundant inside FCDI schemes compared to outside (Figure 3.14). The majority of these species are large predators or conspicuous members of the highly prized migratory ‘whitefish’ category, such as silurid catfish and the carps Labeo rohita, Labeo calbasu, Cirrhinus reba, Cirrhinus mrigala and C. catla.
In the absence of these species, assemblages inside FCDI schemes are dominated by much smaller resident ‘blackfish’ species including the spiny eel Mastacembelus pancalus, the garfish Xenentodon cancila, and the glass perches Chanda nama and Chanda baculis (Figure 3.14). As well as also being less species rich, assemblages inhabiting floodplain/beel habitat inside FCDI schemes have a 25% lower unit value (mean price/kg) based upon average market prices quoted by FAP17 (1995) for the same sampling year. Very similar differences were observed for the other statistically significant comparison of floodplain / beel habitat in the NE (See Halls, 1998 for further details).
The overall impact of the PIRDP flood control scheme is due to the combined effect of the accessibility of the PIRDP to immigrant fish (Section 3.4.2.), and the production potential of individual fish inside the scheme (Section 3.4.1). Section 3.4.3 indicated that the species compositions inside the PIRDP were reduced compared to outside. This section further confirms that the actual catch rates inside the scheme were also lower inside than out.
Fish catches and fishing effort were estimated separately in the two sampling regions using a village-based frame survey, followed by twice-monthly interviewing of 40 randomly-selected respondent fishermen (MRAG 1997). To allow for the mobility of fishermen, respondents were selected both in the two main sampling regions, and in an ‘adjacent’ region (Halls et al, in press (a)). Subsistence fishermen were not included in the surveys. Each respondent provided data on the catches and fishing efforts of each type of fishing gear used in each sampling region during the preceding 1–5 days. Mean monthly catch rates were raised to the total catches in each region using the estimated numbers of fishermen from the frame survey.
Total annual fish productivity, measured as catch per unit area (CPUA), was consistently greater (60–104%) in the outside region than the inside region, and higher in 1996 than in 1995 (Table 3.12). Fishing efforts are more difficult to compare due to the mixture of different gears used, but the differences for comparable gear types were insufficient to explain these differences in CPUA.
Table 3.12 Estimated total annual productivity (kg ha-1 y-1) inside and outside the PIRDP (from MRAG, 1997).
| Region | Estimated Productivity (kg ha-1 y -1) | |
|---|---|---|
| 1995 | 1996 | |
| Inside | 51 | 81 |
| Outside | 104 | 130 |
Detailed examinations of these catch data also provided further evidence that the reduction in catches is likely to be at least partly due to the impact of the flood control embankment. In 1995/96 the flood was both higher and longer than in '96/97 (Figure 2.8). Reflecting this variation in hydrological conditions, the main Talimnagar sluice gate was then mostly closed during the 1995 flood, but opened far more often in 1996 (Figure 2.8). Fish catches in the inside region were then only 20% of the outside region catches in the 1995/96 flood year (June '95 to May '96), but rose to 50% in the 1996/97 flood year (Figure 3.15, based on available data for June to December 1996), which had the smaller flood but more frequently opened sluice gates.
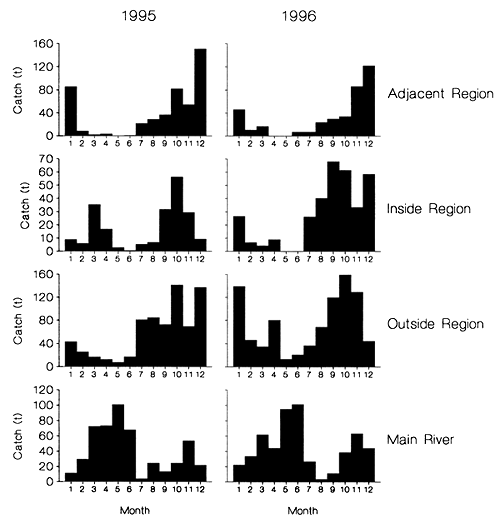
Figure 3.15 Total monthly catches (t) in the four study regions of the PIRDP site in 1995 and '96 (from Hoggarth et al, in press (b)).
![]()
![]()
![]()