![]()
![]()
![]()
by J. P. PERRY, Jr., Administrative Assistant, Rockefeller Foundation Agricultural Program in Mexico
Available records show that the death of pine timber over extensive areas in Central Mexico was first attributed to bark beetles in 1903. In that year Dr. Sylvio J. Bonansea was appointed by the Government of Mexico to make investigations in the States of Mexico, Morelos and Michoacán, where the inhabitants were concerned over the dying pine forests.
Dr. Bonansea found that bark beetles were the primary cause of the trouble. His report La Plaga de Ocotes which was not published until 1921, described the aspects of the dead trees and the insects and their work. He also included many observations on the general features of the infestation and suggested various control measures.
Dr. Bonansea sent specimens of the insects collected in these areas to Dr. A. D. Hopkins of the Bureau of Entomology, U. S. Department of Agriculture. From this material Hopkins described Dendroctonus mexicanus Hopk. and Ips bonanseai Hopk. as new species. In his publication entitled The Genus Dendroctonus, he states that Dendroctonus mexicanus "evidently attacks Pinus teocotl Schl. et Cham., P. leiophylla Sch. et Cham., and P. ayacahuite Ehr." Another species, Dendroctonus parallelocollis Hopk., is listed as occurring in Michoacán but specific host trees are not mentioned. A 1936 publication by Donald De Leon lists, with descriptions and host references, a number of the bark beetle species that attack pines in Mexico. In all the literature that is available there is very little information about the life history and habits of the insects, their host relationships or the causes of the aggressive outbreaks that are characteristic of the infestations.
At the present time a highly epidemic infestation of bark beetles is in progress in stands of Pinus leiophylla around the valley of Mexico, similar in all its characteristics to the outbreaks of half a century ago which were described by Prof. Bonansea. 1 Recent observations by the author indicate that widespread bark beetle outbreaks have occurred at periodic intervals in Central Mexico over a very long period and that their effects have at times accelerated the transition from forest cover to unwise agricultural use of the land.
1 An FAO entomologist, J. M. Miller, is now working in Mexico under the Expanded Technical Assistance Program, and two other FAO experts, G. Becker and F. Schwerdtfeger, have been investigating similar bark beetle epidemics in Guatemala.
The purpose of this article is to put on record the results of studies made by the author during the years 1949 and 1950 in Central Mexico. In the course of intermittent field trips from Mexico City, it has been possible to acquire an understanding of the habits of three species of bark beetles, their interrelationships, the manner in which they attack and kill living trees, and the extent to which they may destroy certain stands of pine.
Areas and character of pine stands affected by recent epidemics
The infested areas studied by the author are located near the towns of Tlalmanalco, San Rafael and Amecameca in the State of Mexico. Dr. Sylvio Bonansea refers to large infestations of D. mexicanus in this same locality and undoubtedly there have been intermittent epidemics in the area as a whole during the past 45 years. Judging from the size of the present beetle-killed areas, the timber losses during these years in this locality must have been tremendous.
Information obtained from the local population dates the beginning of the present epidemic et about 1947 and the "peones" questioned were unanimous in locating the first outbreak as occurring on the slopes of a small mountain a few miles west of the town of Amecameca. Ing. Humberto Ortega reports that he observed the first indications of increasing bark beetle activity in this area in 1945. The writer has observed the final stages of this kill and found that the forest was composed of an even-staged stand of Pinus leiophylla ² averaging 10 in. (25 cm.) d.b.h. with an understory of scattered scrub oak (Quercus sp.). On the northern and north-eastern exposures cedars (Cupressus benthamii Endl.) also formed a small part of the overstory. With the exception of a few trees on the south-western fringe, this pine forest had been completely wiped out by 1949 leaving only scrub oak and cedar. The area of this kill is estimated to be 4,000 acres (1,600 ha.). D. mexicanus ³ apparently spread in 1949 from this infested area across the surrounding countryside, attacking many of the small isolated pine stands in the vicinity of Tlalmanalco. It was in a number of these stands that the writer was able to follow in considerable detail the progress and development of the insect.
² Specimens of the trees mentioned in this article are deposited in the author's herbarium.
³ Duplicate specimens of the insects mentioned in this article are deposited in the U. S. National Museum.
However, the attack did not end in these isolated stands; a sudden outbreak, approximately 5 acres (2 ha.) in extent, was first observed in 1950 in the foothills of the Sierra Nevada range, near the town of San Rafael, approximately 5 miles (8 km.) from Amecameca. The beetle is now spreading up the pine forested slopes of this mountain range and only a rough estimate can be made of the number of acres of trees that will be destroyed. The predominant pine on the lower slopes is Pinus leiophylla with an intermingling of P. montezumae and P. montezumae var. macrocarpa. Martinez. At higher elevations the two latter pines become more evident and fir (Abies religiosa) begins to appear and increases in numbers with the increase in elevation. As the forest gradually changes with increasing preight from predominantly P. leiophylla to P. montezumae var. macrocarpa - A. religiosa, the epidemic apparently loses force and vigor and gradually dies out.
Life history of Dendroctonus mexicanus
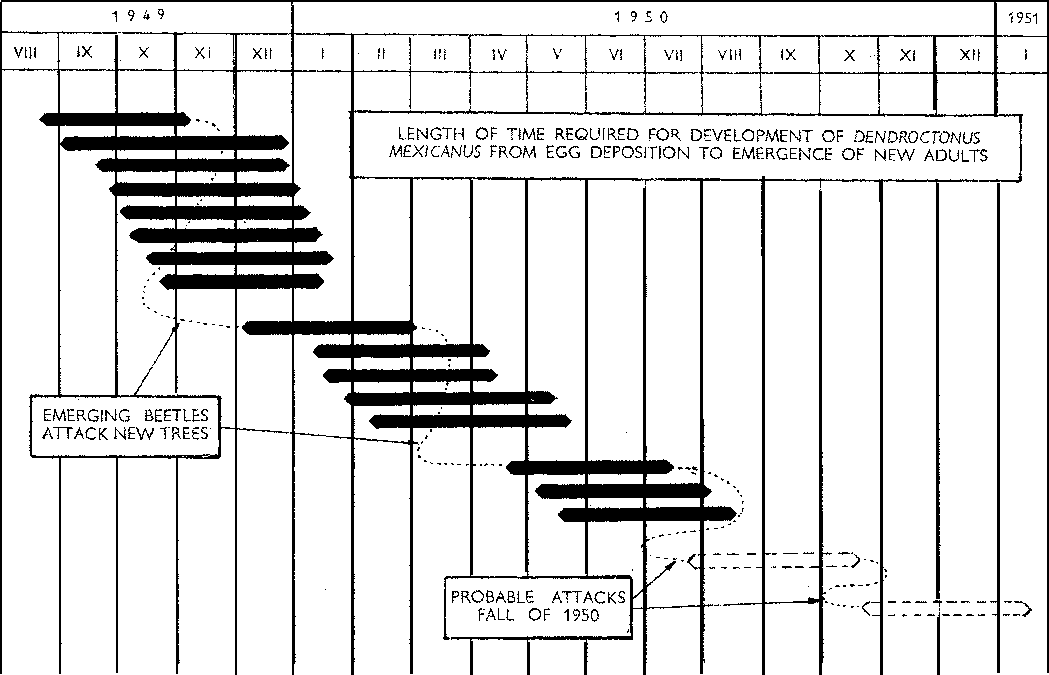
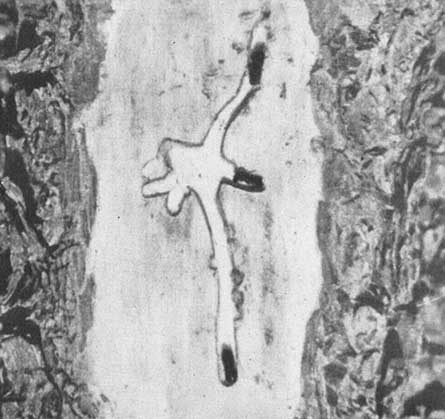
The bark beetle primarly responsible for the extensive kills in these areas is Dendroctonus mexicanus. The habits of this species have not been fully described in previous publications, so are covered in some detail. In 1906 Hopkins described the adults of this species as averaging 3.8 mm. in length and closely resembling D. frontalis though somewhat smaller. The adults attack the living tree in force but each individual entrance in the bark represents the work of a pair of beetles, generally with the female actually excavating the tunnel entrance. From the entrance the tunnel is excavated vertically up the trunk for a short distance, allowing the pitch to flow freely from the entrance hole. During this stage the activities of the male are limited to pushing out the resin as it flows into the tunnel. When the resin flow ceases, the tunnel is curved sharply and extended down and around the stem in a winding, elongated "S" shaped pattern. The writer has recorded tunnel length of 13 to 16 in (33-41 cm.) with the adults still actively elongating the tunnel and laying eggs.
Male Dendroctonus mexicanus working the pitch flow in order to keep tunnel and entrance open.
Full grown larva and young adult in cells in outer hark of Pinus leiophylla.
Egg laying proceeds as follows: the female first excavates a small niche in the tunnel wall, then the egg is deposited in this niche and frass packed round it. The eggs are placed in niches alternately along both sides of the tunnel. It was found from a number of egg counts that the female deposits an average of 6 eggs per inch (2.5 cm.) of tunnel. The male working approximately two in. (5 cm.) behind the female, packs the tunnel solidly with frass. As the eggs hatch, the minute larvae mine outward into the phloem at right angles to the main tunnel. As they increase in size, they gradually feed toward the outer, dry bark, attaining their maximum size as they reach the outer bark. When they enter the bark, all feeding ceases and a cell is constructed wherein the changes from larvae to pupae to adult take place. The new adults mine directly outward from the pupal cell through the bark and emerge completely developed and capable of immediately initiating a new attack on living pines.
The following table gives data on the average number of days required for the development of D. mexicanus in Pinus leiophylla:
TABLE 1. - NUMBER OF DAYS REQUIRED FOR DEVELOPMENT FROM EGG TO NEW ADULT, D. mexicanus IN Pinus leiophylla
|
Area No.* |
No Trees in which Brood Development was followed |
Average No Days Egg to Adult |
|
4 |
36 |
79.9 |
|
5 |
2 |
76.0 |
|
1 |
19 |
78.6 |
|
3 |
57 |
79.4 |
* The area numbers are selected stands representing slightly different conditions in elevation and exposure.
Characteristic form of gallery made by adult beetles in Pinus leiophylla.
The biotic potential. It can be seen from the data on the developmental period that it is possible to obtain four generations of insects per year. This gives the insect a tremendous biotic potential - particularly since no true period of dormancy was observed during its life cycle. Even during the coldest months of the year - November, December and January - temperatures during the middle of the day were high enough to insure some feeding and development of larvae. The writer has also recorded numerous successful attacks as occurring during these coldest months.
Time was not available in these studies to determine whether any percentage of the parent adults re-emerge, after constructing egg-galleries in the first tree attacked, to start new attacks in other trees. This habit has been studied in a closely related species in California, D. brevicomis, where it was found that at times as high as 50 percent of the parent adults re-emerge to start new attacks. However, observations of the egg-galleries constructed by D. mexicanus indicate that most of the parent adults die near the ends of these galleries, and re-emergence of the parent adults apparently is not an important factor in the biotic potential of this bark beetle.
Dendroctonus valens. After the initial attack by D. mexicanus the infested pine is then invaded by D. valens. This insect is well known throughout the United States, and its habits have been recorded in bulletins of the U. S. Department of Agriculture 4.
4 Hopkins, A. D., The Genus Dendroctonus, U.S.D.A. Bulletin No. 83, 1909.
Beal, J. A., and Massey, C. L., Bark Beetles and Ambrosia Beetles, School of Forestry, Duke University, Durban, N. Carolina, Bulletin 10,1945.
Keen, F. P., Insect Enemies of Western Forests, U.S.D.A. Miscellaneous publication No. 273, 1938.
It is interesting to note, however, that there is apparently a minimum diameter limit under which it does not attack P. leiophylla in Central Mexico. Numerous records by the author place this minimum diameter limit at approximately 6 in. (15 cm.) outside the bark, measured at the ground line. There is also apparently no true period of dormancy in its life cycle in this area, since attacks were commonly found occurring during all months of the year.
Ips bonanseai. Following the successful invasion of the tree by D. mexicanus and D. valens, there occurs another invasion by a third scolytid, Ips bonanseai Hopk. The adult insect as described by Hopkins averages 3.3 mm. in length, is elongated, cylindrical and of a dark reddish brown color. The declivity of the elytra is excavated and each side is armed with four short acute teeth. This species only invades the tree after the attacks of D. mexicanus have completely stopped the flow of resin, and then only the upper part of the stem and larger branches are attacked. The entrance hole is bored through the bark and phloem to the xylem. A nuptial chamber is excavated in the phloem and from this chamber three or four winding egg-galleries are constructed. The females deposit their eggs in niches spaced alternately along the sides of the galleries. Frass is packed around each egg in its niche but the egg-galleries and nuptial chamber are kept open at all times. As the eggs hatch, the larvae feed outward from the galleries into the phloem. As the larvae mature, a pupal cell is constructed in the phloem and the pupal period is passed in this cell. The new adults work their way through the old frass that was once phloem tissue, to the old galleries and emerge through the entrance and ventilation holes. No effort was made to determine the average length of time required to complete the life cycle of this insect. However, it was noted that infestations occurred during the coldest months of the year which suggests the absence of a dormancy period in the life cycle.
Sequence of attack on bark areas of Pinus Leiophylla
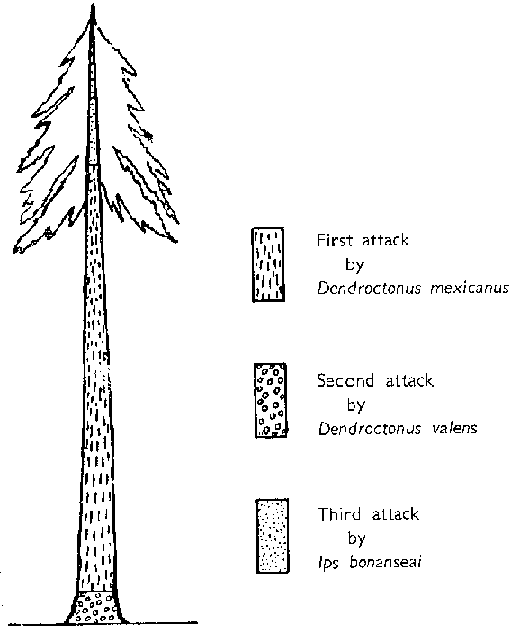
Significance of the sequence of attack in individual trees
There is thus a sequence of invasions of the living tree by these three species of scolytids. The primary attack by D. mexicanus is always made on the stem of the tree, generally at a height of 18 to 20 ft. (5-6 m.) from the ground if the bole is fairly clean. In smaller tees or trees with many low branches, the attack is generally started in the area where the first branche occur. From this point the infestation extends down the stem to a distance approximately 18 in. (46 cm.) from the ground and at the same time up the stem to a top diameter of 4 in. (10 cm.). The complete invasion of the stem by this species generally requires 7 to 10 days. At this point D. valens successfully invades the tree from the ground line up the stem for a distance of approximately 18 in. (46 cm.)
Following the primary and secondary invasions, Ips bonanseai attacks the top portion of the tree. In every instance the attack was observed to occur only when the foliage of the pine had started to wilt or approximately 20 days after the invasion of the stem by D. mexicanus, when the latter had reached the minute larval stage. The Ips infestation extended down the stem to a diameter of 6 in. (15 cm.) and was also found in the bases of the larger branches. In no instance was there observed a reversal of this sequence of attack. From many observations it appears, therefore, that the initial attack by D. mexicanus is absolutely necessary for subsequent successful invasions by D. valens and Ips bonanseai.
Group attacks
Group behavior of D. mexicanus in attacking stands of Pinus leiophylla appears to be quite similar to that of other species of Dendroctonus attacking coniferous stands in the western U.S.A. The spread of a number of infestations in small isolated stands was followed in considerable detail and in every instance the pattern was the same. At first a small group of trees was heavily attacked, then the adjoining trees were rapidly invaded until a center of infestation including 10 to 20 or more trees was established. The surrounding trees wore generally attacked by the new beetles emerging from the first group, the infestation slowly spreading through the stand in ever widening bands. A characteristic feature of these small killed areas is the few isolated surviving trees left on the fringes.
In other instances, particularly in large stands of pine, group infestations were observed on a much larger scale. A number of these larger infestations covered from 5 to 10 or more acres (2-4 ha.) and thousands of pine trees were attacked simultaneously. As the first generation brood emerged, they immediately attacked the surrounding living trees and in a comparatively short period of time a forest covering several hundred acres was completely infested. In some cases groups of large size appeared suddenly in areas where no previous attacks had occurred. It is believed that these large spontaneous infestations are the result of mass flights of insects from an infested area where the number of living trees is no longer sufficient to afford hosts for the tremendous population of emerging beetles. The large group outbreak in 1950 near San Rafael is believed to have occurred in this manner when the emerging beetles in the infested forest near Amecameca no longer found enough living host trees nearby. This mass flight covered a distance of approximately 5 miles (8 km.), but on smaller stands where the population pressure never becomes so tremendous, movement of the newly emerged beetles is generally limited to the nearest living pine trees. As long as there are sufficient living pines in the larger infestation centers, the insects apparently merely spread out from the original group kill. This spread of the beetles through a pine forest can be easily followed by the changes that occur in the color of the foliage of the infested pines.
Table 2 indicates the length of time required for the most important color changes:
TABLE 2
|
Color of Foliage and State of Tree |
Approx. no. days from date of attack |
Average Development of Dendroctonus mexicanus |
|
Fading |
30 |
Small to medium larvae in phloem |
|
Brown |
60 |
Pupae in outer bark |
|
Red |
90 |
Adults emerging |
|
Black (needles gone) |
120 |
Emergence complete |
|
Fruiting bodies of fungus Polyporus volvatus on stem |
150 |
|
|
Destruction of wood by fungi proceeding rapidly. Tree now fallen to ground. |
300 |
|
Thus complete destruction of a pine tree killed by D. mexicanus occurs in less than one year. The author has photographed patches of wheat growing among the stumps of pines that had been killed by D. mexicanus for less than a year.
Comparative resistance of Pinus leiophylla and p. Montezumae var. macrocarpa
In the areas that were studied it was noted that P. leiophylla was quickly overcome by the attacks of D. mexicanus and that in the heavy epidemic centers, few trees survived.
The only other pines on which the attack was observed were determined by Prof. Martinez as Pinus montezumae and P. montezumae var. macrocarpa. In the latter variety, which is often present in mixed stands with P. leiophylla, the beetles apparently encountered difficulties, and many trees were found which had survived strong attacks. Very few attacks have been found in P. montezumae.
Differences between P. leiophylla and P. montezumae var. macrocarpa in their resistance to D. mexicanus were noted in the character and abundance of the pitch tubes, the success of the beetles-in producing progeny and the ability of epidemic infestations to sustain their momentum.
Differences in Character of Pitch Tubes. In P. leiophylla the entrance holes of the beetles were marked by a copious flow of the oleoresins. As a rule, well defined pitch tubes do not form, but the pitch tends to drip down the bark surface forming long, glistening streamers. In P. montezumae var. macrocarpa the entrance holes are more numerous and the resins harden, quickly forming distinct turret-like pitch tubes. Many of the attacking adults are trapped by rapid drying of the pitch and die in the pitch tube without gaining entrance to the tree. This difference in the character of the pitch tubes is due to differences between the chemical components of the oleoresins produced by the two species of pines. A report by Dr. N. T. Mirov shows that the oleoresin from P. montezumae is composed almost exclusively of alpha-pine dextrose, while that from P. leiophylla contains only 90 percent alpha-pine dextrose, 8 percent of delta-3-carene dextrose and 1 percent of sesquiterpine dextrose.
Differences in brood potentials of D. mexicanus. Some trees of Pinus montezumae var. macrocarpa are killed by D. mexicanus, but it was noted that in these cases there was a heavier attack of beetles per unit area of bark than in P. leiophylla. A comparison of the number of entrance holes, each of which represents the attack of a pair of parent beetles, and of the exit holes which represent the emergence of their progeny, will illustrate this difference. Table 3 is based on counts of the entrance and exit holes in bark areas, selected at random upon two killed trees representing both species of pine.
TABLE 3. - COMPARISON OF NUMBER OF ENTRANCE AND EXIT HOLES OF D. mexicanus IN P. Ieiophylla AND P. montezumae var. macrocarpa
|
|
P. leiophylla |
P montezumae var. macrocarpa |
||
|
Sq. ft. |
m² |
Sq. ft. |
m² |
|
|
Total bark surface counted |
6.8 |
0.6 |
5.5 |
0.5 |
|
Average No. of entrance holes per sq. ft. (m²) |
8.0 |
0.7 |
16.8 |
1.6 |
|
Average No. of exit holes per sq. ft. (m²) |
104.0 |
9.7 |
39.0 |
3.6 |
This table indicates that more than twice as many beetles attacked the bark surface of P. montezumae var. macrocarpa than attacked P. leiophylla, while more than two and a half times as many progeny emerged from P. leiophylla as from var. macrocarpa. In fact, if we allow two parent adults to each entrance hole, the number of beetles attacking per square foot in var. macrocarpa is 33.6 (362 per m²), the number emerging 39 (420 per m²), which permits only a negligible increase per generation. When mortality of the new beetles during the flight period and attack on new trees is taken into account, a decrease of beetle population is to be expected.
The pitch flow produced by Dendroctonus mexicanus attacks on Pinus leiophylla
The pitch flow produced by Dendroctonus mexicanus attacks on P. montezumal var. macrocarpa
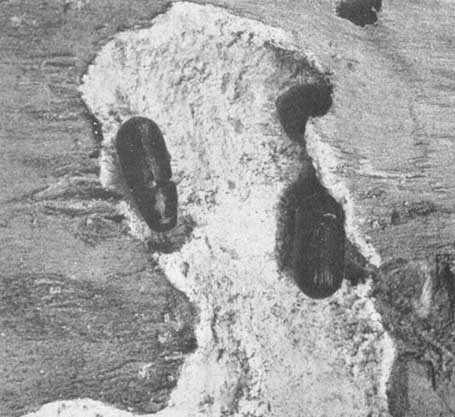
Adult Ips bonanseai (x 3 approx.) in partially constructed egg galleries
Egg gallery and adult (x 5) of Dendroctonus valens
Effect of host species upon populations of D. mexicanus. - The results shown in Table 3 are entirely in line with field observations. In pure or nearly pure even-aged stands of P. leiophylla, D. mexicanus becomes epidemic and spreads rapidly until practically all of the stand has been destroyed. Epidemics are less aggressive where P. montezumae var. macrocarpa makes up an appreciable percentage of the stand, and where a high percent of this tree species is found, no epidemics have yet been observed.
Control experiments were not attempted in these studies as time was available only for collecting biological and ecological data. However, some information on both natural and artificial control of the beetle is available and will therefore he included here.
The sequence of the attacks indicates that Ips and D. valens cannot maintain epidemic infestations except in a secondary capacity to D. mexicanus. For this reason control efforts can be concentrated on D. mexicanus to prevent the virulence and spread of epidemics like those now in progress.
Biological control
Birds. The feeding of woodpeckers on broods of Dendroctonus beetles in the bark of infested trees is very common in the pine forests of the United States, and at times is considered to be an important factor in the natural control of the beetles. Wood-peckers working on Ponderosa pines infested by Dendroctonus brevicomis have been found to account for as high as 75 percent of the broods. However, no woodpecker work has been found in connection with the infestations of D. mexicanus. This may very well be due to the great scarcity of the birds, which through lack of protection seem to be approaching extermination in these areas.
On the other hand Dr. Bonansea mentioned that the reduction of predatory and parasitic insect populations by birds was one of the reasons for the epidemic bark beetle outbreaks. There is no evidence, however, that birds have played any such role in recent outbreaks.
Insect Predators. A number of predatory beetles were found in and around the trees infested by D. mexicanus. Most of these are the same species as those found in connection with D. brevicomis and are undoubtedly important in the biological control of the former species.
Enoclerus sphegens F. (F.), Thanasimus nigriventris and Themnochila virascens, all determined by E. A. Chapin.Cymatodera sp. near Morosa (Lee.) determined by O. L. Cartwright.
Adults of the above species were observed feeding on the adults of D. mexicanus and the larval forms were found in the bark with the larval broods of the bark beetle. Although these predators are present in numbers, their control effect has obviously been insufficient to check the rapid increase of the bark beetle in recent epidemics.
Insect Parasites. Adults of small chalcids were found in rearings of D. mexicanus from infested bark collected from P. leiophylla. These were determined by Dr. B. D. Burks of the U. S. National Museum as Pachyceras sp. and Tomicobia sp. The nature of the rearings did not definitely establish these hymenopterous insects as parasites of D. mexicanus, but it is probable that they are because of the known parasitic habits of many species belonging to this group. A very small number of the adult chalcids were reared, however, and their relative scarcity indicates that they are not an important factor in the biological control of the beetle.
Direct Control
The methods used for the direct control of D. brevicomis should be equally effective with D. mexicanus. These consist of burning the infested bark to destroy the insects, or of spraying the bark with penetrative toxic oils until it is well saturated. In either case felling the tree before treatment of the bark is of advantage, and where burning is used, peeling the bark from the log is desirable.
Ing. Humberto Ortega, Chief of the Department of Forest Protection of the Government of Mexico, reports that direct control work was carried on in 1950 against the infestation in parts of the Unidad Forestal of San Rafael. Where the land was privately owned, it was possible to get the owners to provide the labor to cut, limb and peel the infested trees. The forester of the Unidad marked the trees to be cut and supervised the burning of the infested limbs and tops. The owners were permitted to sell the wood which paid for costs of the operation. Several large infestation centers were treated in this way.
When these same areas were examined one year later, no new attacks were found around the areas that had been treated. These results indicate that eradication work on D. mexicanus has a definite local control effect and that good results are possible if the work is throughly done.
In the "ejidos" 5 which contain the communal land of the Unidad Forestal, the dead trees are given to the communal owners or "ejidatarios" under the law, and it has not been possible to raise the funds to carry on direct control work. Because of this, infestations have run unchecked in the forested areas of ejidal lands. The Unidad Forestal, however, maintains a forest nursery and is replanting some of the devasted areas.
5 Lands which have been restored or granted to agricultural communities under the land reform initiated in 1915.
Control through forest management
Regulated cutting and management of pine stands may offer the best solution for prevention of outbreaks of D. mexicanus. In stands of Pinus leiophylla that had been killed by the recent epidemics, growth of the older trees had been rapid for the first 35 to 40 years; but beyond this age there was a very abrupt slowing down of growth as shown by suppression; of the annual rings. It was during this period of suppressed growth that the present bark beetle epidemics occurred. This evidence suggests that in the even-aged stands of P. leiophylla which have passed the period of rapid growth, clear cutting at about the time when growth begins to stagnate, would partially eliminate the hazard of beetle epidemics and allow for full utilization of the wood before it deteriorates. Replanting of these clear cut areas with more resistant species, such as P. montezumae var. macrocarpa, should offer a sound method of silvicultural control of the beetle in those areas where sustained production of pine is desired.
The writer wishes to express his gratitude to the many people who aided in the preparation of this study; to John M. Miller, Entomologist of the Berkeley Forest Insect Laboratory Bureau of Entomology and Plant Quarantine, U.S.D.A., for his assistance in the field and for his many helpful suggestions and criticisms during the preparation of the manuscript; to Dr. J. A. Beal, Chief U.S.D.A., Bureau Ent. & Plant Quar., Div. of Forest Insect Investigations and Dr. Dorothy Parker, Botanist for Rockefeller Foundation Agricultural Program in Mexico for reading the manuscript and offering many helpful suggestions; to Dr. N. T. Mirov, U.S.D.A. Chemist, California Forest & Range Expt. Sta., Berkeley, Cal., for information on the chemical analyses of pine resins; to Ing. Humberto Ortega, Chief of the Mexican Department of Forest Protection, for information on insect control measures now used in Mexico. All photographs were taken by the author; Byron MacLellan, Photographer for the Rockefeller Foundation, made the necessary prints and enlargements.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}