![]()
![]()
![]()
T. ABOAGYE-KWARTENG, O.K. ole-MOIYOI and
J.D. LONSDALE-ECCLES
International Laboratory for Research on Animal Diseases
P.O. Box 30709, Nairobi, Kenya
Methods
Results
Discussion
References
In the life cycle of African trypanosomes, the parasites differentiate through several morphologically and biochemically distinct forms. In the bloodstream of the mammalian host, Trypanosoma brucei differentiates from a rapidly dividing slender form to a non-dividing stumpy form. While it is clear that this differentiation is important to the life cycle and survival of the parasite, the factors that control the processes have not been identified.
The role of protein phosphorylation in both differentiation and transformation of eukaryotic cells is well established. In Saccharomyces cerevisiae, cAMP-dependent protein phosphorylation is required for cell cycle initiation.¹ In the fission yeast Schizosaccharomyces pombe, the cdc2 gene, whose transcription is required at the start of the cell cycle and probably later in mitosis, encodes a phosphoprotein with protein kinase activity.² Our objective in this study is to identify differentially phosphorylated proteins in bloodstream Trypanosoma brucei and to determine their roles in the differentiation from slender to stumpy form parasites. The basis of an experimental approach with a phosphoprotein of unknown function was nicely formulated by Robinson, who said that: "One enters the story in the middle by first targetting a phosphoprotein as a 'band on a gel' and then proceeding both backwards (what protein kinase is responsible for its labelling?) and forwards (what is the protein's function?)". This report describes the identification of two differentially phosphorylated proteins and a partial characterization of the protein kinase(s) that phosphorylate them.
Preparation of trypanosome cytosolic fraction
Trypanosoma brucei brucei ILTat 1.1 was used in this study. The infection produced in laboratory rodents is sufficiently synchronous to allow 100% slender and greater than 90% stumpy trypomastigotes to be isolated on days 4 and 7, respectively, after infection.
Trypanosomes were grown in lethally irradiated rats and the percentage of stumpy forms on different days after infection determined according to Ormerod.4 When the desired stage of parasitaemia was attained, the trypanosomes were purified from blood elements by isopycnic Percoll gradient centrifugation5 and DEAE-cellulose chromatography.6 The purified trypanosomes (5 x 1010) were suspended in 25 mM Hepes, pH 7.0, 250 mM sucrose, 2 mM EDTA, 10 mM EGTA and 2 mM DTT with 40 m g/mL each of the protease inhibitors leupeptin, antipain and E-64 and disrupted in a French Press at a pressure of 2,500 psi. The homogenate was centrifuged at 100,000 g for 60 minutes to give a cytosolic fraction (HSS) and pellet (HSP).
Protein kinase assay
Assays were carried out in a total volume of 150 m l with the following components: 50 mM Tris-HCl pH 7.2, 150 mM KCl, 10 mM MgCl2, 5 m M ATP, 10, m Ci [32P]ATP (5000 Ci/mmol), 150 m g of protein and enzyme source (usually stumpy HSS). The reaction was carried out on ice for 10 minutes and stopped by the addition of 50 m l 4 x SDS sample buffer (200 mM Tris-HCl pH 6.8,8% SDS, 40% glycerol and 4% 2-mercaptoethanol) and boiling for 5 minutes.
Electrophoresis
The phosphorylated proteins were separated by SDS polyacrylamide gel electrophoresis (SDS-PAGE) according to Laemmli,7 using 7.5-15% polyacrylamide gradient gels. The gels were stained with Coomassie blue, destained and dried. Radioactive protein bands were detected by autoradiography. Quantitative measurement of phosphate incorporation into individual bands were made by counting, in aquasol, the respective regions excised from the gel.
The phosphorylated amino acids recovered from protein bands of approximate Mr 42 and 37, which had been electroeluted from SDS-PAGE gels, were determined using electrophoresis on cellulose thin-layer plates.8
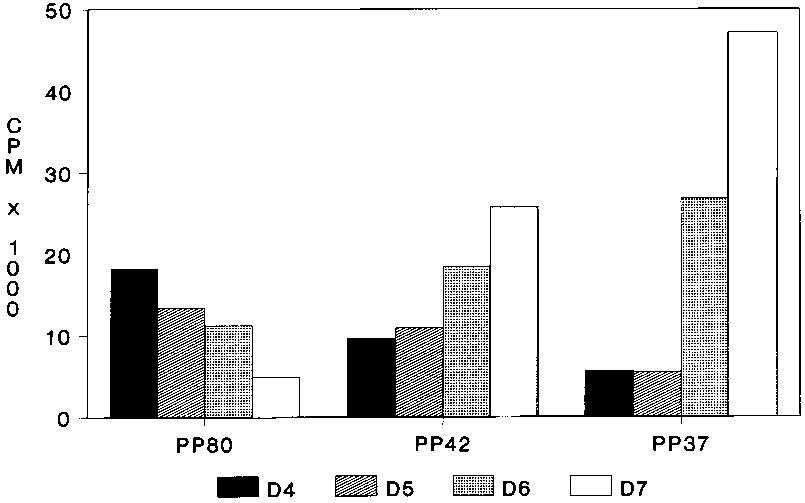
Clear differences were observed in the profiles of the cytosoluble (HSS) phosphoproteins of trypanosomes isolated between 4 and 7 days after infection (Figure 1). There is decreased phosphorylation of a protein with Mr 80 kDa (pp80) with the change from the slender to stumpy form parasite. However, all the work described in this paper is on the two proteins, with Mr of about 42 kDa (pp42) and 37 kDa (pp37), which show increased phosphorylation with the differentiation of the slender to the stumpy parasite. Protein kinase assays of the nonsoluble fractions (HSP) show a much lower level of phosphorylation of both pp42 and pp37, which suggests that both proteins are predominantly cytosolic.
Trypanosome protein kinases are able to use both ATP and GTP as phosphoryl donors. Heparin, which inhibits casein kinase II but not casein kinase I,9 inhibits the phosphorylation of pp42 but has little effect on pp37 phosphorylation. Suramin, a trypanocidal polysulphated naphthylamine, was also found to inhibit the activity of trypanosome protein kinases (50% inhibition at about 25 m M).
The metal ion requirements for the phosphorylation of both proteins suggest the presence of more than one protein kinase in T. b. brucei.10 The optimum Mg2+ concentration for the phosphorylation of pp42 is 10 mM. For pp37, however, the optimal Mg2+ concentration is 100 mM. The trypanosome protein kinases can use Mn2+ at approximately five-fold lower concentrations than those required for Mg2+.
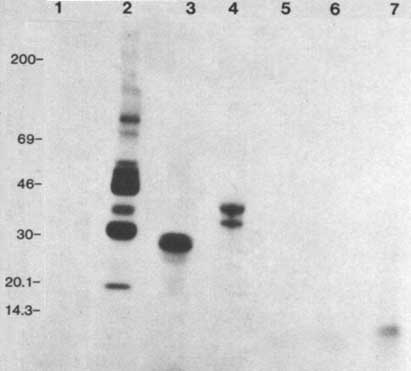
Assays with enzymatic amounts of D7 HSS and a variety of substrates showed that the trypanosome protein kinases can phosphorylate the acidic proteins casein and phosvitin but not the basic histones or protamin (Figure 2). Analysis showed that both pp37 and pp42 are phosphorylated on serine and threonine residues.
Protein phosphorylation has emerged as a mechanism involved in the hormonal control of metabolism in mature cells11 and in the changes in cellular activity associated with cellular proliferation and differentiation.12 The work presented above demonstrates the coincidental increased phosphorylation of two proteins (pp42 and pp37) with the differentiation of slender bloodstream forms of T. b. brucei into the stumpy forms. Previous studies of T. b. gambiense have shown the presence of multiple cAMP-independent protein kinases, which can phosphorylate phosvitin, protamine and histones However, no endogenous protein substrates of these kinases were determined.13 Our work has identified two differentially phosphorylated endogenous proteins in T. b. brucei.
Our studies on the effects of metal ions on the phosphorylation of pp42 and pp37 suggest that the two proteins are phosphorylated by different enzymes. This is in agreement with studies of T. b. gambiense that have demonstrated the presence of multiple protein kinases.11
It has been shown that phosphorylation on both serine and threonine residues differentiates casein kinase II activity from casein kinase I, which only phosphorylates serine residues. Casein kinase II is also distinguished from casein kinase I in its ability to use both ATP and GTP as phosphoryl donors in phosphotransferase reactions.9 Our studies with effectors are independent of cyclic-nucleotides, calcium/calmodulin, and calcium/phospholipid. Phosphoamino acid studies on pp42 and pp37 showed that the amino acid residues phosphorylated were serine and threonine, thus implicating the involvement of casein kinase II-like enzymes in these phosphorylations and eliminating the involvement of tyrosine kinases. Further studies established the trypanosome protein kinases as casein kinase II-like because of their ability to phosphorylate casein and phosvitin and their stimulation by KCl.
Sensitivity to heparin inhibition has been used to differentiate casein kinase II from casein kinase I.9 We observed that while the phosphorylation of pp42 was sensitive to inhibition by heparin, the phosphorylation of pp35 was relatively unaffected. However, recent studies in Xenopus laevis oocytes10 and yeast14 have identified heparin-insensitive casein kinase II enzymes. Thus the validity of using susceptibility to heparin inhibition as a criterion for distinguishing of casein kinase II from the type I enzyme is questionable.
The polyamine spermine has been shown to activate casein kinase II.9 In contrast, we found that the phosphorylation of pp42 was inhibited by spermine while pp37 phosphorylation was largely unaffected. This further suggests that the two proteins are phosphorylated by different enzymes. Another difference between the casein kinase II-like activity of trypanosomes and the enzyme described from mammalian sources is the effect of polylysine and polyarginine. While the classical mammalian enzymes are activated by these effectors, the low-density-lipoprotein-receptor kinase15 and the trypanosome enzymes are inhibited by them. Thus the inhibition studies show that the protein phosphorylation in trypanosomes is inhibited by large negatively and positively charged molecules.
In conclusion, we have identified two differentially phosphorylated proteins in T. b. brucei. The proteins with Mr 42 and 37 kDa on SDS-PAGE are more heavily phosphorylated in the stumpy parasite than in the slender. The activities of the enzymes that phosphorylate the proteins are independent of cyclic-nucleotides, calcium/calmodulin or calcium/phospholipids. The trypanosome protein kinases are casein kinase II-like but show some significant differences from the classical mammalian casein kinases.
1. MATSUMOTO, K., I. UNO, Y. OSHIMA and T. ISHIKAWA. 1982. Proc. Natl. Acad. Sci. USA 79: 2355-2359.
2. SIMANIS, V. and P. NURSE. 1986. Cell 45: 261-268.
3. ROBINSON, P.J. 1985. Cell 42: 703-704.
4. ORMEROD, W.E. 1963. Exp. Parasitol. 13: 374-385.
5. GRAB, D.J. and J.J. BWAYO. 1983. Acta Trop. 39: 363-366.
6. LANHAM, S.M. and D.G. GODFREY. 1970. Exp. Parasitol. 28: 521-534.
7. LAEMMLI, U.K. 1970. Nature 227: 680-685.
8. MANAI, M. and J. COZZONE. 1982. Anal. Biochem. 124: 12-18.
9. HATHAWAY, G.M. and J.A. TRAUGH. 1982. Curr. Top. Cell. Regul. 21: 101.
10. TAYLOR, A., C.C. ALLENDE, R. WEINMANN and J.E. ALLENDE. 1987. FEBS Lett. 226: 109-114.
11. COHEN, P. 1985. Eur. J. Biochem 151: 439-448.
12. HELDIN, C-H. and B. WESTMARK. 1984. Cell 37: 9-20.
13. WALTER, R.D. 1978. Hoppe Seyler's Z. Physiol. Chem. 359: 601-606.
14. STERNBACH, H. and H. KUNTZEL. 1987. Biochem. 26: 4207-4212.
15. KISHIMOTO, A., BROWN, M.S., C.A. SLAUGHTER and I.L. GOLDSTEIN. 1987. J. Biol. Chem. 262: 1344-1351.
![]()
![]()
![]()
{kind=link}
{kind=link}