![]()
![]()
![]()
R.D. ALLEN, C.R.C. SCHROEDER and A.K. FOK
Pacific Biomedical Research Center and Department of Microbiology
University of Hawaii
Honolulu, HI 96822, USA
Membranes Belonging to the Phagosome-Lysosome System
Characteristics of the Cytopharynx and Nascent Phagosome Membrane
Evidence for in Situ Membrane Modification
Fates of the Different Pools of Membrane Incorporated into the Membrane of Phagosomes or Phagolysosomes
Discussion and Conclusions
References
Membranes are in constant flux with their surroundings. They exchange molecules with other membranes and receive or give up additional molecules in tune with the cell's physiological requirements. Some membranes can suddenly grow larger or retract in area. To alter membranes, the cell sometimes replaces an existing membrane with relatively large areas of prefomed membrane while the putative original membrane is being removed as small vesicles. Thus, the cell has the capacity to modify its membranes to fit immediate as well as ongoing requirements.
As a hypothesis, we are proceeding in our research on the assumption that a given membrane can be modified in situ to provide for changing roles. The well-documented receptor clustering into coated pits, and the sorting out and return of the receptors to the plasma membrane following ligand release in some types of receptor-mediated endocytosis,1,2,3 involves modifications of this type, albeit on a more limited scale than we envision possible. To test for broader possibilities of this hypothesis, we are investigating the membranes of the system of the free-living protozoan Paramecium. These membranes are ideally suited to such a study because of the numerous functions they must assume in the life of a vacuole and their well-documented highly plastic nature.
Phagosome-lysosome membranes perform a range of critical functions for the cell. (1) The phagosome membrane encloses entering food and particulate matter and maintains an unbroken hydrophobic barrier between these potentially harmful substances and the cell's cytosol. (2) The mature phagosome membrane encloses a highly acid environment during the process of prey killing and digestion and probably contains the mechanism for the acidification of the phagosome. (3) These membranes protect the cytosol from digestive enzymes. (4) These membranes facilitate the movement of the products of digestion into the cytosol subsequent to the selective entry of acid hydrolases into the phagosome. (5) Finally, the various membranes of the phagosome-lysosome system have the capacity to recognize other membranes in a time-dependent preprogrammed manner.
The intent of this short review will be to illustrate the way the fine structure of one membrane is modified in synchrony with the changing state of the space enclosed by the membrane and to explain how this modification is brought about. Figure 1 provides a guide to the structures and membranes discussed in this paper.
We have recently been able to determine with some certainty the total range of membranous vesicles and structures in Paramecium that can be considered part of the phagosome-lysosome system.4 Using the lectin wheat germ agglutinin (WGA), all membranes previously known to be related to this system were labelled. These included not only those membranes around the discoidal vesicles, digestive vacuoles of undetermined stage, Lysosomes (shown by Lüthe and Plattner to bind to WGA-gold on Lowicryl K4M sections), but also the acidosomes, the trans cisterna of the Golgi stack, a reticulum of membranes answering the description of the trans-Golgi network and a continuum of vesicles ranging from 70 nm to 500 nm or more.4 Using WGA-gold on frozen-thin sections of cells, which had been serially pulse-fed with latex beads of a different size for each pulse and with the pulses interspersed with appropriate chase times allowing the labelled vacuoles to reach predetermined stages, we have shown conclusively that all vacuole stages label with WGA. Because the labelling with WGA is completely inhibited by triacetylchitotriose and because Paramecium seems to lack sialic acid,6 we conclude that these membranes may be distinguished from other membranes by having in common a glycocalyx containing either b -(14) oligomers of N-acetyl-glucosamine (b -(1-4) GlcNAc) or a very high content of GlcNAc. No label was found on the plasma membrane or coated pits at the cell surface. However, the membranes of crystal-containing compartments were heavily labelled, suggesting that these structures are related to the phagosome-lysosome system.
Having determined the full range of membrane-bound compartments associated with the phagosome-lysosome system, we can now look at the characteristics of its subdivisions.
Freeze-fracture images7 as well as deep-etch, rotary shadowing (unpublished observations) show the membrane of the cytopharynx to have a highly particulate E-fracture face unlike the plasma membrane with which it is continuous. Discoidal vesicles have an identical E-fracture face. These two membranes also bear a glycocalyx of identical appearance and thickness (10 nm).8 In addition, their similarities to each other and dissimilarities from other membranes of the phagosome-lysosome system are shown by the existence of a monoclonal antibody9 that binds only to epitopes found on these two membranes.10 These similarities, along with the morphological evidence of discoidal vesicles that have fused with the cytopharynx11, indicate that nascent phagosomes obtain their membrane from the pool of discoidal vesicles.
Again, freeze-fracture evidence has shown that the membrane of the early phagosome (also termed DV-I stage) is very quickly and dramatically altered so that by 15 sec to 1 min after pinching off from the cytopharynx its membrane no longer contains the high number of intramembrane particles (IMPs) on its E-fracture face but now is almost entirely devoid of IMPs.7 These particles are not simply transferred to the P-fracture face because the IMP number on this face remains about the same. The phagosomes at this stage are termed DV-II. The glycocalyx of the membrane of these DV-II phagosomes are also altered by being reduced to a slightly uneven 7 to 10 min in thickness.8
This membrane is identical in freeze-fracture12 and glycocalyx8 appearance to a set of fairly large vesicles that bind to the nascent phagosome and travel with the phagosome to the cell's posterior pole. At the pole the vesicles are seen to fuse with the phagosome.13 Studies with weak bases such as neutral red and acridine orange show that these vesicles are acidic and, furthermore, the phagosome becomes acidic after, but not before, these vesicles have fused with the phagosome.14 Prevention of fusion by the application of cytochalasin B inhibits phagosome acidification. Thus, we have termed these vesicles acidosomes.14 Acidosome and acidified-phagosome (DV-II) membranes have equal concentrations of prominent particles on their P-fracture faces 7,12 which is quite different from the P-fracture face of the DV-I.7 The relationship between DV-II and acidosomes is further demonstrated by the unique presence of epitopes for a second monoclonal antibody,9 which reacts with no other DVs.10 These epitopes are also found in the crystal-containing compartments (unpublished observation). We have always been impressed by the fact that the contents of acidosomes resemble the contents of the extracellular space in freeze-fractured replicas. This resemblance is even more pronounced in deep-etched images (unpublished observation). In NH4Cl studies we see that these acidosomes are sometimes labelled with horseradish peroxidase within a very short time, as short as 1 see, following exposure of the cells to this enzyme marker (unpublished observation). Yet we have so far been unable to determine the origin of these acidosomes. Using deep-etch techniques following rapid freezing of living cells, we have recently observed a previously unrecognized set of small vesicles aligned along the same microtubules at the cytopharynx to which discoidal vesicles are attached (unpublished observations). Though not yet studied in detail, their proximity to the site at which acidosomes bind to phagosomes suggests a possible role for these vesicles in the formation of acidosomes should they coalesce to form the larger vesicles.
Our evidence thus suggests that shortly after they are formed phagosomes undergo a radical membrane replacement12 during which the original membrane is removed by vesiculation and the membrane remaining in the DV-II is, for the most part, derived from that of the acidosomes. The timing of this change corresponds to the time when the phagosome becomes acid, beginning at about 1 min and reaching a maximum acidity at about 5 min into the digestive cycle. 12
At this time Lysosomes can approach and bind to the DV-II. Lysosome membranes have a moderate number of very prominent IMPs on their E-fracture faces7 and an extensive 30 nm thick glycocalyx lining their luminal surface.8 Lysosomes also bear epitopes not shared by discoidal vesicles, acidosomes or the phagosomes.10 However, the membranes of phagolysosomes (DV-III) resemble Lysosomes in IMP number and distribution,7 appearance of their glycocalyxes8 and in monoclonal antibody specificity.10 Thus, the vacuole membrane has undergone another substantial modification, which occurs concomitant with fusion of Lysosomes beginning 8 minutes into the cycle.15
The DV-III now contains acid phosphatase activity for the first time.l6 The pH then rises dramatically.15 By the time the pH has returned to 6 or 7, digestion is presumably completed and a final membrane modification begins. Portions of the planar membrane are remolded into long tubules of a diameter of 45 nm, which contain acid phosphatase activity.l7 These tubules expand at their distal ends into vesicles that resemble Lysosomes in all ways, IMP appearance,17 glycocalyx appearance,l7 acid phosphatase activity17 and the presence of epitopes for the lysosome-specific monoclonal antibody (unpublished observation).
Vacuoles become defecation competent, under the axenic growth conditions we routinely use, at around 20 min.15 However, defecation of the labelled vacuoles in a population of cells follows an exponential pattern. The rate of defecation depends to a marked extent on the rate of vacuole formation.18 The membranes of these spent vacuoles (DV-IV), which are now acid phosphatase-negative,16 have not been studied extensively but they are probably quantitatively rather than qualitatively different from membranes of the DV-III phagolysosomes.
At this stage in our story, solid evidence becomes scarce. Discoidal vesicle membrane seems for the most part to be retrieved soon after phagosomes are formed. Only one or two phagosomes per cell react with monoclonal antibodies specific for the discoidal vesicle and nascent phagosome membranes.9 We presume this membrane can be recycled to the cytopharynx for new phagosome formation but we have no hard evidence for this. The second group of monoclonal antibodies that are specific for acidosomes also bind strongly to a few vacuoles near the cytopharynx,9 but these epitopes seem to be diluted out of the vacuole membrane with time. However, even spent DV-IV contain some epitopes for the latter group of monoclonal antibodies.9 Lysosome membrane is retrieved from the older DV-III. Whether this retrieval is in bulk or is composed of only certain components of the Lysosomes is not known. Thus the spent vacuole membrane may be composed of a mixture of phospholipids derived from the three vesicle populations and may contain an assortment of glycoproteins and proteins retained in the vacuole membrane that were not sorted out and retrieved at earlier stages. Spent vacuole membrane apparently does not bear epitopes for discoidal vesicles, only a low level of epitopes for acidosomes, and few, if any, epitopes for Lysosomes. Spent vacuole membrane like other vacuole membranes is, however, highly reactive for WGA.4
Once the vacuole is defecated, the DV-IV membrane is retrieved as tubules and vesicles of different shapes.19 At least some of these vesicles become discoidal, attach to microtubular ribbons and move toward the cytopharynx.20 This portion of the spent vacuole membrane is recycled as discoidal vesicles. With our new evidence for another set of small spherical vesicles, we will need to see if these arise also from the cytoproct. Horseradish peroxidase is inactivated below pH 5,21 so we will need to use another marker to be certain we do not miss vesicles that might have a very acid lumen.
Morphological and monoclonal antibody studies provide conclusive evidence for the extensive in situ modification of an existing membrane that follows the changing physiology of the space that it surrounds. These changes can be explained for the most part by the insertion and retrieval of vesicular membrane of different vesicle populations. Whether slower membrane changes can occur by intercalation of individual molecules of proteins and phospholipids is not known.
Membrane modifications such as those reported here can be very fast and can involve large amounts of membrane surface area. To carry out such a rapid change, the cell sequesters preformed membrane around the membrane to be modified. This sequestered membrane is poised to fuse with only a limited subset of the vacuole population of the cell. Specific recognition factors must be present on the cytosolic side of the membrane of this subset of vacuoles to provide binding sites for only one specific set of vesicles. The nature of these recognition factors is completely unknown. Nor is anything known about the triggers that cause the vesicles to fuse.
The putative recognition factors presented on the cytosolic vacuole surface must be replaced with time since acidosomes and, later, Lysosomes bind to the same vacuole. This replacement of recognition factors may be brought about by the addition to the vacuole membrane of vesicular membrane bearing the required recognition sites. Yet the answer is probably not this simple. In the case of lysosome binding, recognition sites for Lysosomes could be added to the vacuole during acidosome fusion, but Lysosomes do not bind directly to vesicular acidosomes free in the cytoplasm. Other factors must be active in the proper presentation of the putative recognition factors in a form that can be recognized by the vesicles.
Those aspects of membrane traffic in the phagosome-lysosome system in Paramecium remaining to be worked out include the origins of acidosomes and the identification of all sources of discoidal vesicles, that is, do these vesicles have any origin than that from the membrane itself. The ultimate origin of the lysosome membrane appears to be from the trans-Golgi network and the Golgi stacks.4 This membrane may represent the source of new membrane entering the system. Of immediate concern is the determination of the amount of intermixing of membrane components from separate sources and the determination if the cell has the capacity to sort and retrieve, as a package, those components that comprise uniquely acidosome, discoidal vesicle and lysosome membrane components. These questions are being studied using both morphological and biochemical techniques.
1. WILLINGHAM, M. C. and I. PASTAN. 1984. Int. Rev. Cytol. 92: 51-92.
2. WILEMAN, T., C. HARDING and P. STAHL. 1985. Biochem. J. 232: 1-14.
3. HOPKINS, C.R. 1986. Trends Biochem. Sci. 11: 473 - 477.
4. ALLEN, R.D., C.C. SCHROEDER and A.K. FOK. In press. Histochem. Cytochem.
5. LUTHE, N. and H. PLATTNER. 1986. Histochem. 85: 377 - 388.
6. PAPE, R., B. HAACKE-BELL, N. LUTHE and H. PLATTNER. 1988. J. Cell Sci. 90: 37-49.
7. ALLEN, R.D. and L.A. STAEHELIN. 1981. J. Cell Biol. 89: 9-20.
8. ALLEN, R.D. and A.K. FOK. 1984. Eur. J. Cell Biol. 35: 149-155.
9. FOK, A.K., M.S. UENO and R.D. ALLEN. 1986. Eur. J. Cell Biol. 40: 1-8.
10. FOK, A.K. and R.D. ALLEN. 1988. In Paramecium. H.D. Gortz, Ed.: 301-324. Springer-Verlag, Berlin.
11. ALLEN, R.D. 1974. J. Cell Biol. 63: 904 - 922.
12. ALLEN, R.D. and A.K. FOK. 1983. Eur. J. Cell Biol. 29: 159-165.
13. ALLEN, R.D. and A.K. FOK. 1983. Eur. J. Cell Biol. 29: 150-158.
14. ALLEN, R.D. and A.K. FOK. 1983. J. Cell Biol. 97: 566-570.
15. FOK, A.K., Y. LEE and R.D. ALLEN. 1982. J. Protozool. 29: 409-414.
16. FOK, A.K., J.H. MURAOKA and R.D. ALLEN. 1984. J. Protozool. 31: 216-220.
17. ALLEN, R.D. and A.K. FOK. 1984. J. Cell Biol. 99: 1955-1959.
18. FOK, A.K., B.C. SISON, Jr., M.S. UENO and R.D. ALLEN. 1988. J. Cell Sci. 90: 517-524.
19. ALLEN, R.D. and R.W. WOLF. 1974. J. Cell Sci. 14: 611-631.
20. ALLEN, R.D. and A.K. FOK. 1980. J. Cell Sci. 45: 131-145.
21. FOK, A.K., M.S. UENO and E.A. AZADA. 1985. Eur. J. Cell Biol. 38: 306-311.
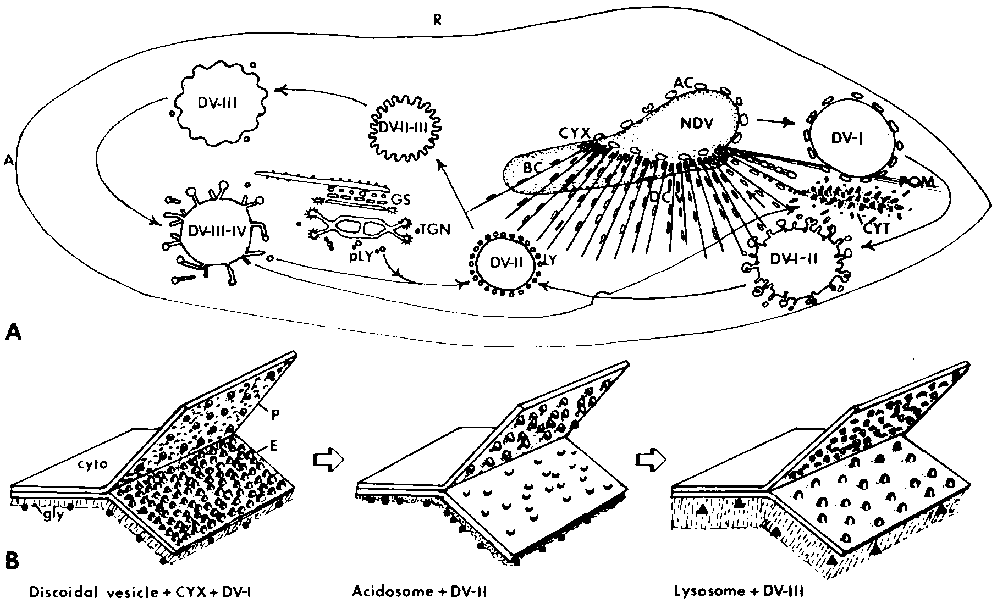
Figure 1. Schematic drawings of membrane flow and modifications in the phagosome-lysosome system of Paramecium. A. View of Paramecium from the dorsal surface. The anterior end (A) is to the left and the right side (R) is at the top. The buccal cavity (BC) is in a mid-ventral location and is continuous with the cytopharynx (CYX), where nascent phagosomes (NDV) form. Ribbons of microtubules fan out from the cytopharynx, directing discoidal vesicles (DCV) to this region. Acidosomes (AC) bind to the NDV as it forms and travel with the phagosome (DV-I) as it moves along the postoral bundles of microtubules (POM) toward the cell's posterior pole. Fusion of the acidosomes and retrieval of DV-I membrane results in a smaller and acidified DV-II which binds to lysosomes (LY). Fusion of the lysosomes with the DV-II leads to a DV-III or phagolysosome. After digestion, lysosome components are retrieved, mix with primary lysosomes (pLY), which bud from the Golgi stack (GS) and/or the trans-Golgi network (TGN), and eventually bind to another DV-II. The spent vacuole (DV-IV), from which lysosome membrane components and acid phosphatase are removed, passes to the cytoproct (CYT), where it is defecated. Spent vacuole membrane is retrieved as tubular vesicles, some of which become discoidal vesicles. B. Membrane especially on the E-fracture face (E) but also on the P- fracture face (P). Another modification occurs in the appearance of the glycocalyx (gly) lining the luminal side of vesicle and vacuole membranes. The cytosolic (cyto) side of the membranes has not been studied but must bear recognition factors. In addition, different populations of vesicles have different antigenic binding sites, which are indicated here as circles, squares and rectangles on the luminal sides of the membranes. The precise location of these antigenic sites is not known.
![]()
![]()
![]()
{kind=link}