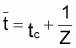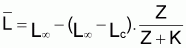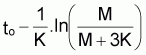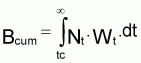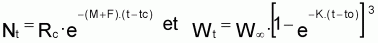![]()
![]()
![]()
On désigne par cohorte ou classe annuelle ou par génération, un groupe de poissons d’un stock nés la même année. Le schéma suivant illustre les différentes phases du cycle de vie d’une cohorte:
Figure 3.1 Cycle de vie d’une cohorte
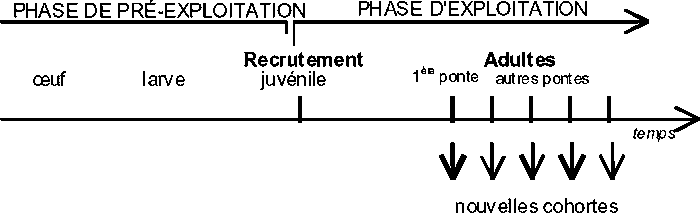
On commence, par exemple, par la phase de l’œuf. Les phases suivantes seront larve, juvénile et adulte.
On désigne par recrutement à la pêche ou à la phase d’exploitation le nombre d’individus d’une cohorte qui entre pour la première fois dans la zone de pêche. Ces individus grandissent, pondent (une ou plusieurs fois) et meurent.
Dès la 1ère ponte les individus de la cohorte sont appelés adultes, et, généralement, ils pondent à nouveau tous les ans, donnant ainsi origine à de nouvelles cohortes.
Les phases de la vie de chaque cohorte qui précèdent le recrutement à la zone de pêche (œuf, larve, pré-recrues), sont des phases importantes de leur cycle de vie, mais, généralement, elles ne sont pas sujettes à l’exploitation, les variations d’abondance étant dues essentiellement à la prédation et aux facteurs de l’environnement (vents, courants, température, salinité...). La mortalité dans ces phases de non-exploitation est normalement très élevée surtout pendant la dernière phase larvaire (Cushing, 1996) résultant, ainsi, en une survie très réduite jusqu’au recrutement, cette mortalité, comme indiqué plus haut, n’est pas causée par la pêche.
Le recrutement à la phase d’exploitation d’une cohorte (le nombre d’individus qui entrent pour la première fois dans la phase exploitée dans une année déterminée), peut survenir pendant plusieurs mois sous les formes schématiques suivantes:
Figure 3.2 Recrutement à la phase d’exploitation
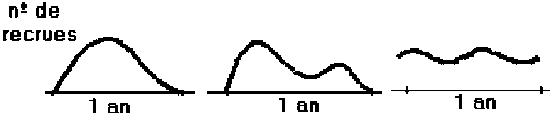
À l'exception de quelques cas, les formes de recrutement peuvent se simplifier si l’on admet que tous les individus recrutent à l’instant, tr, désigné par âge de recrutement à la phase d’exploitation. Par convention, on considère que le recrutement a lieu le 1er janvier (début de l’année civile dans plusieurs pays). La simplification et la convention mentionnées antérieurement n’altèrent pas beaucoup les résultats des analyses, mais simplifient ces dernières et correspondent aux périodes auxquelles les statistiques commerciales s’appliquent.
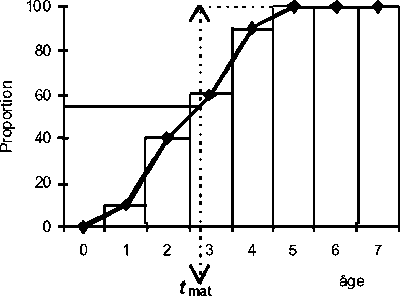
La 1ère ponte ne se réalise pas au même âge pour tous les individus de la cohorte. La proportion des individus qui pondent pour la 1ère fois augmente avec l’âge, de zéro à 100%. À partir de l’âge auquel 100% ont pond la 1ère fois, tous les individus sont adultes. L’histogramme ou la ligne polygonale qui représente ces proportions est nommé ogive de maturation.
Dans quelques cas l’ogive de maturation peut être simplifiée supposant que la 1ère ponte a lieu à l’âge tmat dénommé par âge de 1ère maturation. Cette simplification signifie que les individus d’âge inférieur à tmat sont considérés juvéniles et que ceux d’âge égal ou supérieur sont considérés adultes.
La Figure 3.3 représente une ogive de maturation en forme d’histogramme ou de courbe.
Figure 3.3 Ogive de maturation
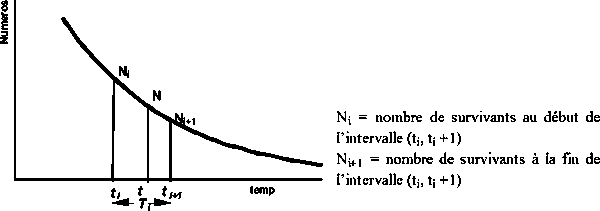
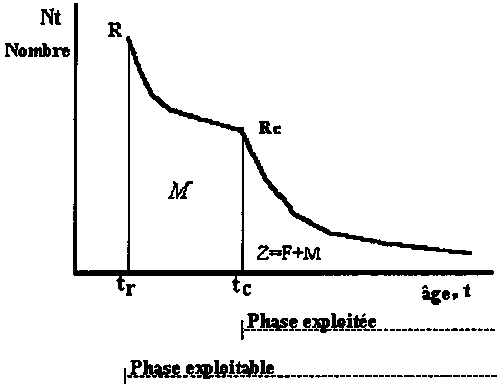
Soit l’intervalle (ti, ti+1) de taille Ti = ti+1 - ti de l’évolution d’une cohorte avec le temps et Nt le nombre de survivants de la cohorte à l’instant t de l’intervalle Ti (voir Figure 3.4).
L’information existante suggère que les taux moyens de variation en pourcentage de Nt peuvent être considérés à peu près constants, soit, tmr(Nt) » constante.
Supposition de base
Le taux instantané relatif de variation de Nt , dans l’intervalle Ti est:
tir (Nt) = constante négative = - Zi
Figure 3.4 Évolution de Ni dans l’intervalle Ti
Le modèle de l’évolution de Nt, dans l’intervalle Ti, est un modèle exponentiel (car tir(Nt) est constant). Ce modèle a les propriétés suivantes:
Propriétés
1. Expression générale. De la supposition de base
tir(Nt) = -Zi
et avec la condition initiale que pour t = ti on aura Nt = Ni , alors:
2. Nombre de survivants, Ni+1 , à la fin de l’intervalle Ti
3. Nombre de morts, Di , pendant l’intervalle Ti
(notez que Di est positif mais la variation DNi = Ni+1- Ni est négative)
4. Nombre accumulé de survivants, Ncumi , pendant l’intervalle Ti
Valeur centrale rapprochée, Ncentrali , dans l’intervalle Ti
6. Nombre moyen,
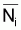 , de survivants pendant
l’intervalle Ti
, de survivants pendant
l’intervalle Ti
(Ricker)
quand
est petit (Zi.Ti < 1)
Commentaires
1. La supposition de base est parfois présentée en termes de taux instantanés absolus, soit,
tia (lnNt) = -Zi ou
tia (Nt) = - Zi.Nt { tia (Nt) proportionnel à Nt}
Zi = coefficient de mortalité total, supposé constant dans l’intervalle Ti
Notez que:
+Zi = tir de mortalité total de Nt
-Zi = tir de variation de Nt
2. Unité de Zi
De la définition on déduit que Zi est formulé en unités de [temps]-1. Par convention on adopte l’unité "an-1" même s’il s’agit d’un intervalle inférieur ou supérieur à un an.
Les expressions suivantes montrent de façon simplifiée le calcul de l’unité de Zi, avec les règles et symboles [..] de dimension habituelle dans la détermination d’unités physiques.
3. Taux annuel de survie, Si
Quand Ti =1 an:
et aussi
Si = taux annuel de survie en l’an i
(ou % du nombre initial d’individus qui survivent à la fin de l’année).
1 - Si = taux annuel de mortalité en l’an i
Par définition c’est le % du nombre initial qui meurt pendant l’année.
Soit le taux relatif moyen tmr(Nt) de mortalité de Nt, pendant un an, par rapport au nombre initial, Ni
4. Taux absolut moyen
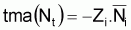
5. Taux relatif moyen
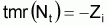 relatif à
relatif à
6. Notez que Si est compris entre 0 et 1, soit,
0 £ Si £ 1 mais Zi peut être > 1
7. Si les limites de l’intervalle Ti
étaient (ti,  ) alors on
aurait:
) alors on
aurait:
Ti =
Ni+1 = 0
Di = Ni
et
Les causes de mort dues à la pêche seront séparées des autres causes de mort des individus de la cohorte. Ces dernières sont groupées en une cause unique désignée par mortalité naturelle. Ainsi, des propriétés du modèle exponentiel, on aura
tir(Nt) total = tir(Nt) naturel + tir(Nt) pêche
Si l’on considère l’intervalle Ti et en supposant que, dans cet intervalle, les taux instantanés de mortalité par causes naturelles et par pêche sont constants et égaux à Mi et à Fi, respectivement, on aura:
Zi = Fi + Mi
En multipliant les deux termes de l’égalité
précédente par
 on aura:
on aura:
comme 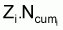 est
le nombre, Di , de morts dus à la mortalité
totale, alors,
est
le nombre, Di , de morts dus à la mortalité
totale, alors,
De même
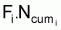 sera le nombre
d’individus morts par la pêche, soit, la capture, Ci, en nombre et,
donc,
sera le nombre
d’individus morts par la pêche, soit, la capture, Ci, en nombre et,
donc,
Notez aussi que
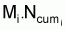 sera le nombre
d’individus morts par des causes naturelles.
sera le nombre
d’individus morts par des causes naturelles.
Le taux d’exploitation, Ei , pendant l’intervalle Ti a été défini par Beverton et Holt (1956) comme étant:
et donc,
ou encore
La capture en nombre, Ci, dans l’intervalle Ti, peut être formulée sous les différentes formes suivantes:
Commentaires
1. Ricker (1975) définit le taux d’exploitation, Ei*, comme le pourcentage du nombre initial qui est capturé dans l’intervalle Ti, soit, Ei* = Ci / Ni.
a) La définition de Ricker peut être plus naturelle mais mathématiquement la définition de Beverton & Holt est plus utile.
b) Il est facile de vérifier que
2. Le taux d’exploitation, Ei , n’a pas d’unité, c’est un nombre abstrait.
3. Les valeurs possibles de Ei sont comprises entre 0 et 1, 0 représentant l’absence d’exploitation et 1 la capture Ci égale au nombre de morts Di, soit, Mi = 0.
Pour étudier l’évolution de la biomasse d’une cohorte, on peut utiliser le modèle de l’évolution d’une cohorte en nombre et la combiner avec un modèle d’évolution du poids moyen d’un individu de la cohorte. En effet, la biomasse Bt est égale à Nt.Wt où Wt est le poids moyen individuel à l’instant t. D’un autre côté, il y a aussi intérêt, plus généralement, de disposer d’un modèle pour l’évolution du poids individuel, Wt, et d’un modèle pour l’évolution de la longueur individuelle, Lt.
Pour définir un modèle pour la croissance en
poids individuel, 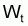 , on
dispose de plusieurs alternatives.
, on
dispose de plusieurs alternatives.
1ère ALTERNATIVE:
A) Définir un modèle pour la croissance individuelle moyenne en longueur,
B) Définir la relation Longueur-Poids.
C) Combiner A) avec B) et obtenir un modèle pour la croissance individuelle moyenne en poids,
2ème ALTERNATIVE:
D) Définir un modèle directement pour
et Lt.
1ère ALTERNATIVE
A) Modèle pour la croissance individuelle en longueur
Les modèles à utiliser en biologie marine sont valides pour la phase d’exploitation de la ressource. Le plus connu est le Modèle de von Bertalanffy (1938). Les observations suggèrent qu’il existe une longueur asymptotique, c’est-à-dire qu’il existe une longueur vers laquelle la longueur individuelle tend.
Figure 3.5 Modèle de von Bertalanffy
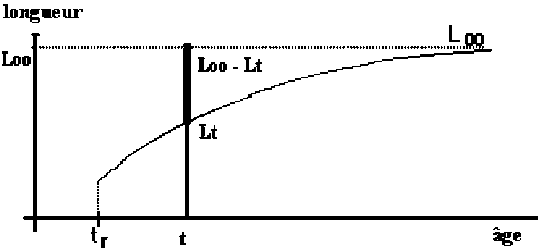
t - âge
Lt - longueur moyenne individuelle à l’âge t
L
tr - début de la phase d’exploitation
Ainsi, Lt présente une évolution où l’on peut vérifier que:
Le tia(Lt) n’est pas constant (car la croissance n’est pas linéaire)
Le tir(Lt) n’est pas constant (car la croissance n’est pas exponentielle)
On observe, pourtant, que la variation de la quantité
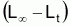 (que l’on pourrait
appeler “ce qu’il reste à grandir”), présente un
taux relatif constant et pourra être décrite par un modèle
exponentiel. Ainsi on peut adopter la:
(que l’on pourrait
appeler “ce qu’il reste à grandir”), présente un
taux relatif constant et pourra être décrite par un modèle
exponentiel. Ainsi on peut adopter la:
Supposition de base
tir (L
où K est le coefficient de croissance (attention: le coefficient de croissance K n’est pas la vitesse de croissance mais la vitesse relative de ce qu’il “reste” à grandir !!).
Les propriétés de ce modèle (puisqu’il s’agit d’un modèle exponentiel) peuvent être obtenues directement à partir des propriétés générales du modèle exponentiel avec la condition initiale:
Propriétés du modèle de croissance
individuelle en taille de Beverton & Holt (1957) déduites du
modèle exponentiel de (L-Lt).
|
Propriétés du modèle exponentiel
pour (L |
Modèle de von Bertalanffy pour Lt |
|
|
1. |
Expression générale |
|
|
|
Paramètres L
pour
|
Paramètres L
pour
|
|
2 |
Valeur à la fin de l’intervalle Ti |
|
|
|
|
expression de Ford-Walford (1933-1946) |
|
3. |
Variation pendant l’intervalle Ti |
|
|
|
|
comme
sera:
|
|
4. |
Valeur accumulée pendant l’intervalle Ti |
|
|
|
|
car: |
|
5. |
Valeur moyenne pendant l’intervalle Ti |
|
|
|
|
expression de Gulland et Holt (1959) car:
|
|
6. |
Valeur centrale de l’intervalle Ti |
|
|
|
|
et
|
B) Relation Longueur-Poids
Il est commun d’utiliser la fonction puissance pour mettre en rapport le poids individuel avec la longueur totale (ou autre). Ainsi:
où la constante a est désignée par facteur de condition ou indice pondéral et la constante b par constante de morphisme. Cette relation peut être justifiée en acceptant grosso-modo que “le pourcentage de croissance en poids” est proportionnel “au pourcentage de croissance en longueur” (dans le cas contraire l’individu serait “disproportionné”) et, en adoptant la supposition basique:
où b est la constante de proportionnalité.
|
C) Combinaison de A) et B) et commentaires: |
|
|
1. |
La combinaison de
Cette relation de croissance en poids est désignée par équation de Richards (1959). Quand b=3 l’équation est l’équation de croissance de von Bertalanffy (1938). |
|
2. |
De |
|
3. |
Soit Comme
On aura,
dans la pratique on
utilise |
|
4. |
Les modèles de Richards et de von Bertalanffy ne sont pas les seuls modèles utilisés pour l’évolution de Wt. D’autres modèles aussi utilisés en évaluation, et à discuter plus tard sont: Le modèle de Gompertz (1825) et le modèle de Ricker (1969) |
|
5. |
Historiquement, le modèle de von Bertalanffy a été développé à partir de la supposition basique
Où Cte1 et Cte2 ont été désignées par von Bertalanffy comme la constante d’anabolisme et de catabolisme, respectivement. En adoptant la relation
(où Cte1 et Cte2 sont d’autres constantes). Équation différentielle dont la solution nous donne l’équation de von Bertalanffy |
2ème ALTERNATIVE
D) Modèle directement pour Wt et Lt
Dans la 1ère alternative, on a développé un modèle pour la croissance en longueur, et ensuite un modèle pour la croissance en poids, en utilisant la relation entre le poids et la longueur.
La supposition de base adoptée par la
1ère alternative a utilisé la caractéristique
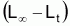 au lieu du poids afin
d’avoir une caractéristique où tir est constant,
c’est-à-dire un modèle exponentiel, et considérer la
relation W=a.Lb pour dériver le modèle de croissance en
poids. Notez que l’on peut dire que L a été
considéré comme étant fonction de W, soit,
L=(W/a)1/b. Il sera, donc, possible préférer à
cette fonction de W, une autre fonction du poids
au lieu du poids afin
d’avoir une caractéristique où tir est constant,
c’est-à-dire un modèle exponentiel, et considérer la
relation W=a.Lb pour dériver le modèle de croissance en
poids. Notez que l’on peut dire que L a été
considéré comme étant fonction de W, soit,
L=(W/a)1/b. Il sera, donc, possible préférer à
cette fonction de W, une autre fonction du poids
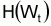 , de façon a que
l’on puisse formuler directement la supposition de base:
, de façon a que
l’on puisse formuler directement la supposition de base:
avec la condition initiale
Propriétés
Les propriétés de ce modèle
(puisqu’il s’agit d’un modèle exponentiel) peuvent
êtres obtenues directement a partir des propriétés du
modèle exponentiel. Il est particulièrement intéressant de
dériver l’expression générale pour
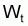 résultante de
différents choix de la fonction H.
résultante de
différents choix de la fonction H.
|
1. |
Expression générale |
|
|
|
Ou |
|
|
2a. |
Équation de Richards en poids |
En adoptant la fonction suivante H(Wt) = Wt1/b on obtiendra l’expression générale: |
|
|
|
|
|
|
L’équation de Richards, quand b=3, est l’équation de von Bertalanffy, ainsi: |
|
|
2b. |
L’équation de von Bertalanffy, en poids, sera: |
|
|
3. |
Équation de Gompertz en poids |
En choisissant la fonction H(Wt) = ln Wt on obtiendra l’expression générale:
|
|
4. |
Les équations en longueur respectives pourront être obtenues en choisissant d’autres fonctions de H(Wt). Ainsi: |
|
|
4a. |
Équation de von Bertalanffy, en longueur |
Choisissant H(Wt) = Lt sera: Lt = L |
|
4b. |
Équation de Gompertz, en longueur |
Choisissant H(Wt) = ln Lt sera: lnLt = lnL |
|
5. |
Équations simplifiées Les équations de croissance individuelle, tant en longueur qu’en poids, sont simplifiées quand on sélectionne |
|
|
|
|
H(Wa) = 0 pour ta = t* |
|
|
Ainsi l’expression générale est réduite à: |
|
|
5a. |
L’équation de Richards, en poids simplifiée sera: |
|
|
|
où t* est représenté par t0 car H(Wa) = 0. Dans le modèle de Richards cela signifie que Wa sera aussi zéro. |
|
|
5b. |
L’équation de Gompertz, en poids simplifiée sera: |
Dans ce cas H(Wa) = 0 correspond à Wa = 1 |
|
5c. |
L’équation de Richards, en longueur simplifiée sera: |
(avec La = 0 pour ta = t0) |
|
5d. |
L’équation de Gompertz, en longueur simplifiée sera: |
(avec La = 1 pour ta = t*) |
Commentaires
1. L’équation de Gompertz, en poids est semblable à l’équation de Gompertz, en longueur, mais dans ces formes simplifiées t* sont des âges différents, puisqu’ils correspondent respectivement à Wa = 1 et à La = 1.
2. L’équation de Gompertz en longueur est semblable à celle de von Bertalanffy avec Lt substitué par lnLt. Dans les cas pratiques cela permet l’utilisation des mêmes méthodes particulières d’estimation des paramètres dans les deux équations, utilisant L dans l’expression de von Bertalanffy et lnL dans l’expression de Gompertz. (voir Section 7.4)
3. On rappelle à nouveau que, dans la pratique, on
utilise Lcentrali et Wcentrali au lieu des valeurs
moyennes, 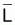 et
et
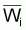
4. La courbe de croissance en longueur de Gompertz infléchit au point (tinfl, Linfl) avec:
tinfl = ta+(1/K).ln(ln(L
5. La courbe de croissance en poids de Gompertz infléchit au point (tinfl, Winfl) avec:
tinfl = ta+(1/K).ln(ln(W
6. La courbe de croissance en longueur de Richards n’a pas de point d’inflexion mais la courbe de croissance en poids a une inflexion au point (tinfl, Winfl):
Dans le cas particulier de l’équation de von Bertalanffy sera:
Winfl = (8/27).W et tinfl =
t0+(1/k).ln3
7. Quelques auteurs se réfèrent à
l’équation de Gompertz sous d’autres formes, en utilisant comme
paramètres ta et Wa le point d’inflexion
tinfl et le poids asymptotique W.
Donc, on aura
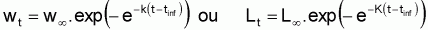
Parfois l’expression est présentée dans sa
forme générale
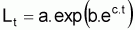
Les paramètres du modèle seront L = a; k = -c et tinfl =
(1/c).ln(-b)
8. En aquaculture, où l’étude de la croissance comprend des âges très tendres et où une inflexion dans la croissance en longueur est vérifiée, on a l’habitude d’utiliser l’équation de Gompertz. Dans la pêche on utilise traditionnellement l’équation de von Bertalanffy.
9. Un modèle, qui peut s’avérer utile, est celui de Ricker (1975). Ce modèle est valide pour un intervalle de temps Ti donné et non pas seulement pour toute la vie exploitée de la ressource. En effet le modèle est basé sur la supposition basique que la croissance individuelle est exponentielle dans l’intervalle Ti.
Il en sera ainsi, par exemple dans
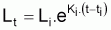 où Ki
peut différer d’intervalle en intervalle.
où Ki
peut différer d’intervalle en intervalle.
1. Biomasses
Théoriquement, on pourrait dire que la biomasse à l’instant t de l’intervalle Ti est donnée par:
Bt = Nt. Wt
et, donc, la biomasse accumulée dans l’intervalle Ti serait:
et la biomasse moyenne dans l’intervalle Ti serait:
Le poids moyen de la cohorte,
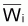 , dans l’intervalle
Ti serait:
, dans l’intervalle
Ti serait:
Et, en divisant les deux termes de la fraction par Ti
sera 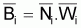
2. Capture, en poids
La capture en poids, Yi , pendant l’intervalle T sera exprimée comme le produit de la capture, Ci , fois le poids moyen individuel pendant l’intervalle Ti:
Commentaires
Dans la pratique
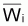 est considéré
à peu près égal à
est considéré
à peu près égal à
 dans
l’intervalle Ti.
dans
l’intervalle Ti.
L’expression antérieure de Yi peut aussi être formulée comme:
On considère l’évolution d’une cohorte pendant toute la vie exploitable, débutant à l’âge tr. On considère des intervalles, Ti, de temps, où i varie du premier au dernier intervalle, couvrant toute la phase d’exploitation (fréquemment les intervalles sont d’un an...).
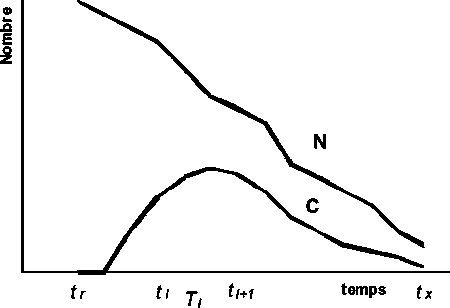
La Figure 3.6 simule l’évolution du nombre de survivants de la cohorte, N , et l’évolution des captures en nombre, C, qui sont obtenues pendant les successifs intervalles de temps Ti.
Figure 3.6 Évolution du nombre de survivants de la cohorte, N, et des captures en nombre, C
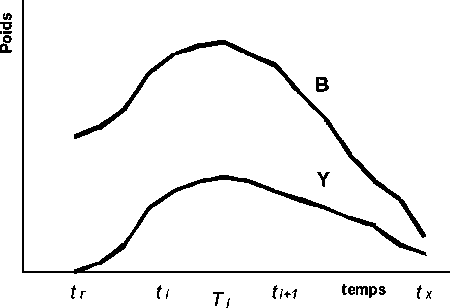
La Figure 3.7 simule l’évolution de la biomasse de la cohorte, B, et l’évolution des captures en poids, Y, qui sont obtenues pendant les intervalles de temps successifs Ti.
Figure 3.7 Évolution de la biomasse de la cohorte, B, et des captures en poids, Y
Valeurs des caractéristiques plus importantes de la cohorte, pendant toute la phase d’exploitation
|
Durée de vie de la cohorte |
l = S Ti |
|
Nombre total de morts |
D =S Di (= R (recrutement) quand tous les individus sont morts) |
|
Nombre accumulé de survivants |
|
|
Nombre moyen de survivants |
|
|
Biomasse accumulée de survivants |
|
|
Biomasse moyenne de survivants |
|
|
Capture en nombre |
C = S Ci |
|
Capture en poids |
Y = S Yi |
|
Poids moyen de la cohorte |
|
|
Poids moyen de la capture |
|
|
Âge critique |
tcritique = âge où B est maximum, quand la cohorte n’est pas exploitée |
Commentaires
1. À première vue, il semblerait que les valeurs des caractéristiques d’une cohorte pendant toute la phase d’exploitation ne présentent pas grand intérêt, car, dans la pratique, la pêche s’exerce rarement sur une cohorte isolée. À chaque instant, les survivants de plusieurs cohortes sont présents et disponibles à la pêche simultanément.
2. Malgré cela et pour des raisons que l’on verra plus tard, il importe d’analyser les caractéristiques d’une cohorte pendant toute sa vie exploitable. La connaissance de l’évolution d’une cohorte, en nombre et en biomasse, particulièrement l’âge critique, a une grande importance pour le succès des activités de l’aquaculture. Comme Bt = Nt. Wt l’âge critique, tcritique , sera l’âge t dans l’intervalle Ti où tir (Wt) = - tir(Nt)=M car étant l’âge critique correspondant à la biomasse maximale non exploitée, la dérivée de B à l’ordre de t sera égale à zéro.
3. Notez que Ncum peut être exprimé en fonction du recrutement comme
Ncum = R.{...}
où {...} représente une fonction des paramètres biologiques et des coefficients de mortalité Fi pendant les années de la vie de la cohorte. Bcum peut aussi être exprimé comme:
Bcum = R.{...}
où {...} a le même sens et inclut les paramètres de croissance.
Beverton et Holt (1957) ont déduit des expressions algébriques pour les caractéristiques d’une cohorte pendant la vie exploitée en adoptant quelques suppositions simples:
1. La phase exploitée de la cohorte débute à l’âge tc et s’étend jusqu’à l’infini.
2. Le coefficient de mortalité naturelle, M, est constant pendant toute la phase d’exploitation.
Figure 3.8 Vie d’une cohorte pendant la phase d’exploitation
3. Le coefficient de mortalité par pêche, F, est constant pendant toute la phase exploitée, soit, la phase de la vie de la cohorte qui débute à tc
4. La croissance suit l’équation de von Bertalanffy avec La = 0 pour ta = t0
Équations de base relatives à la phase d’exploitation
|
1. |
c (quotient entre la longueur à l’âge
|
|
|
2. |
Recrutement |
|
|
3. |
Nombre accumulé |
|
|
4. |
Capture en nombre |
|
|
5. |
Biomasse accumulée |
|
|
|
qui peut s’écrire symboliquement |
|
|
6. |
Poids moyen à la capture |
|
|
7. |
Capture en poids |
|
|
8. |
Âge moyen à la capture |
|
|
9. |
Longueur moyenne à la capture |
|
|
10. |
Âge critique |
tcritique =
|
Commentaires
1. La simplification de Beverton et Holt permet le calcul de quelques caractéristiques de la cohorte pendant sa vie avec des expressions algébriques n’étant pas nécessaire la somme des valeurs de la caractéristique dans les successifs intervalles Ti. Ce facteur est utile quand on ne dispose pas d’ordinateur, et c'est pour cela que la simplification a été très utilisée pour effectuer des calculs dans les années 60-70. Elle est aussi utile quand on dispose seulement, pour l’évaluation des ressources, de données de mortalité naturelle, M, et de paramètres de croissance.
2. La simplification est aussi beaucoup utilisée actuellement pour étudier les effets des changements dans le coefficient de mortalité par pêche, F, et aussi des changements de l’âge de première capture, tc , sur la biomasse de la cohorte pendant toute la vie exploitée, sur la capture en poids correspondante et sur le poids moyen à la capture. Ces analyses sont normalement illustrées par des figures. Par exemple la Figure 3.9 illustre l’analyse des biomasses et des captures en poids obtenues d’une cohorte pendant la vie exploitée, sujette à différents coefficients de mortalité par pêche supposant un âge tc fixe.
Figure 3.9 Évolution des biomasses et des captures en poids d’une cohorte sujette à différents coefficients de mortalité par pêche et tc fixe (notez que la Figure illustre seulement l’analyse et ne prends pas en compte les échelles des axes).
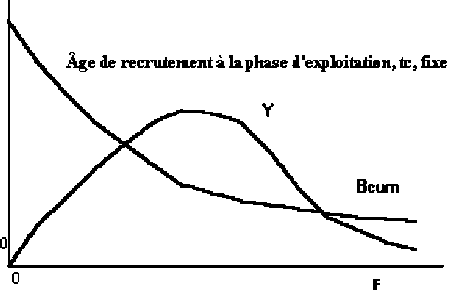
3. Notez que les formes des courbes antérieures, Y et Bcum contre F, ne dépendent pas de la valeur du recrutement et, sont communément désignées par courbes de biomasse et capture par recrue, B/R et Y/R, respectivement. Pour simplifier les calculs sont réalisés en admettant R=1000.
4. De toutes les caractéristiques de la cohorte pendant la vie, le poids moyen à la capture (ainsi comme l’âge et la longueur moyens à la capture) ne dépendent pas de la valeur du recrutement. Ces courbes des caractéristiques d’une cohorte pendant la vie contre le niveau de pêche, F, ou contre l’âge de la 1ère capture, tc, méritent une étude plus soignée pour des raisons exposées dans le chapitre sur les prévisions du stock à long terme.
5. Bcum a été calculé comme:
où
Les calculs peuvent aussi être réalisés en utilisant une valeur de la constante b, de la relation W-L, différente du coefficient d’isomorphisme, b=3.
Pour cela, on peut recourir à la fonction mathématique Beta incomplète (Jones, 1957).
6. Pour le calcul de
 et
et
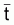 on procède de mode
identique au calcul de
on procède de mode
identique au calcul de 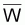 ,
soit, on calcule
,
soit, on calcule
Ces moyennes sont désignées par moyennes pondérées, étant les facteurs de pondération le nombre de survivants Nt à chaque âge t.
![]()
![]()
![]()
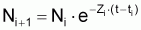
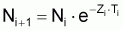
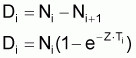
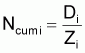
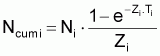
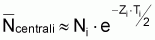
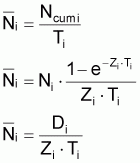
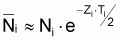
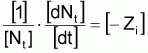
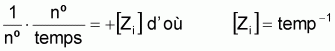
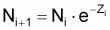
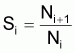
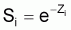
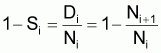
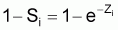
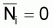
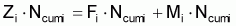
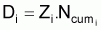
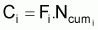

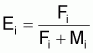
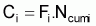
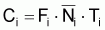
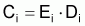
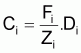
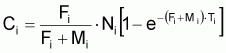
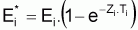
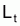
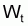
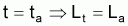
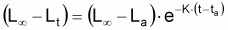

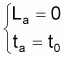
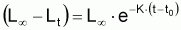
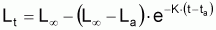

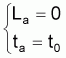
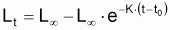
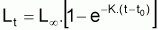
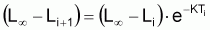
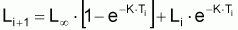
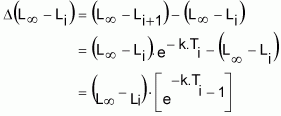
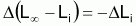
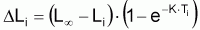
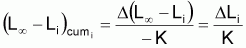


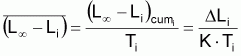
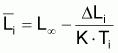
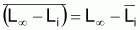
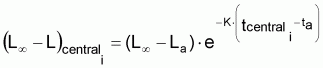

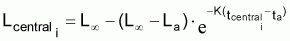

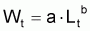
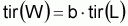
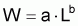 avec
avec
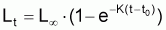 résulte
en
résulte
en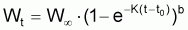 avec
avec
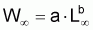
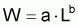 on
aura par définition,
on
aura par définition,
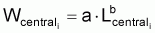 , où
, où
 est la valeur correspondante
à
est la valeur correspondante
à 
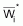 le
poids correspondant à
le
poids correspondant à
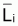 , c’est à dire,
, c’est à dire,
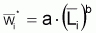


 et
et
 au lieu des points
moyens
au lieu des points
moyens
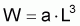 on aura tia(Lt)
= Cte1 - Cte2. L
on aura tia(Lt)
= Cte1 - Cte2. L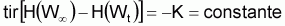
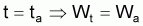

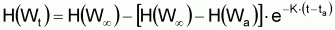
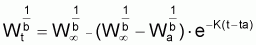
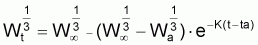
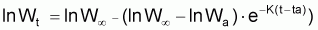
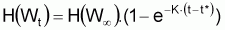
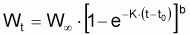
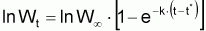
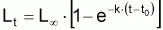
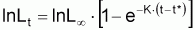
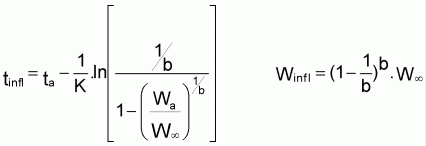
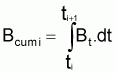
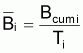
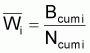
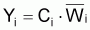
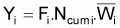
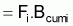
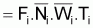
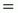
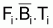
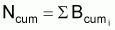
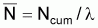
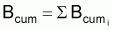

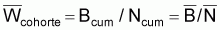
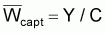
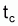 et la longueur
asymptotique)
et la longueur
asymptotique)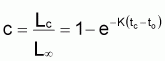
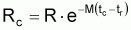
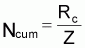
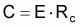
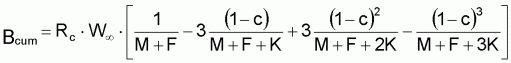
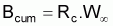 .[...]
.[...]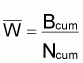 =
Z. W
=
Z. W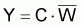 =
F. Bcum = F. Rc. W
=
F. Bcum = F. Rc. W