![]()
![]()
![]()
QUELS SONT LES POISSONS DES GRANDS FONDS?
Les océans étant un écosystème continu, toute zonation basée sur la profondeur serait arbitraire. Pour certains spécialistes, les habitats d'eau profonde commencent à 500 mètres, alors que pour d'autres, les poissons des grands fonds sont ceux qui vivent en dessous de 400 mètres - il n'existe pas de définition stricte et universellement applicable des poissons des grands fonds. Cependant, les poissons des grands fonds peuvent être classés en plusieurs catégories: mésopélagiques, bathypélagiques et benthopélagiques. Les espèces mésopélagiques et bathypélagiques sont des poissons pélagiques au vrai sens du terme, généralement de petite taille à l'âge adulte et rarement exploités à l'échelle commerciale. Les poissons mésopélagiques, comme les lanternes (Myctophidae) et les Gonostomatidae vivent en dessous de la zone photique à une profondeur d'environ 1000 mètres. Les poissons bathypélagiques vivent en dessous de 1 000 mètres et sont ordinairement très adaptés aux environnement pauvres en nutriments. Les espèces benthopélagiques vivent tout près du fond, dans une zone aujourd'hui connue pour la variété de ses caractéristiques biologiques.
Dans ce document, les poissons des grands fonds sont ceux qui vivent au-delà du plateau continental, dans des eaux plus profondes, c'est-à-dire à proximité et en dessous des talus. Les caractéristiques de comportement de nombreuses espèces dites «des grands fonds» font qu'il est encore plus difficile de trouver une définition simple et applicable de ces espèces. De nombreuses espèces des grands fonds migrent vers la surface la nuit et redescendent vers le fond dans la journée, de sorte qu'il se crée un lien trophique entre les eaux de surface et les poissons benthopélagiques, lorsque ces derniers attaquent les poissons qui redescendent vers le fond pour les manger. D'autres poissons migrent eux-mêmes entre le jour et la nuit, se nourrissant dans les couches superficielles, puis descendant vers le fond, probablement pour éviter de se faire manger. Certaines espèces n'habitent les eaux profondes que lorsqu'elles sont adultes et peuvent être exploitées dans leurs phases peu profonde et profonde, de sorte qu'il est difficile de discerner s'il s'agit de captures d'espèces des grands fonds ou non.
Les premières espèces démersales des grands fonds existantes étaient présentes il y a environ 80millions d'années et, dans l'ordre chronologique d'apparition des fossiles, ces espèces étaient les Holosauridae, Bythitidae, Ophidiidae (abadèches et brotules), Macrouridae (grenadiers et rats), Scorpaenidae (rascasses), Trachichthyidae, et plus récemment, Notocanthidae et Moridae (mores) (Andriyashev, 1953). Comme les poissons démersaux sont distribués en fonction de la profondeur, ceux qui vivent sur le talus et le seuil continental sont répartis le long de régions définies en fonction de la bathymétrie se déroulant en ruban sur le pourtour des mers. Lorsque des espèces pélagiques profondes empiètent sur ces habitats, cela crée des possibilités d'alimentation pour les deux groupes.
La plupart de ces familles de poissons se rencontrent partout dans le monde, mais l'existence de bassins d'eau profonde délimités par les continents et les dorsales océaniques a créé des différences régionales qui découlent probablement de la dérive des continents et de la formation successive des océans. Une autre caractéristique importante des poissons des grands fonds est qu'il s'agit d'un domaine encore mal connu, dans lequel on continue à faire de nouvelles découvertes, comme le requin grande gueule, de 4,5 m de long et de 750 kg et la raie Hexanchidae, qui ont récemment été découverts et représentent de nouvelles familles taxonomiques. De fait, la conservation des élasmobranches d'eau profonde est particulièrement difficile à assurer, même quand ils ne sont pas exploités, étant donné leur faible nombre, leur maturité tardive, leur faible fécondité et leur reproduction intermittente.
Malgré l'existence de quelques pêcheries hauturières traditionnelles (la pêche à la palangrotte du sabre noir (Aphanopus carbo) à Madère et aux Açores est l'une des plus intéressantes), les espèces commerciales les plus importantes sont celles qui sont pêchées au chalut. Beaucoup se rencontrent à proximité des monts sous-marins et des crêtes océaniques, alors que dans le passé, d'autres étaient aussi capturées dans des eaux moins profondes, mais les zones de pêche se sont étendues aux grands fonds au fur et à mesure que les technologies de pêche hauturière se sont améliorées et que les stocks des eaux moins profondes se sont épuisés à force d'être exploités.
Les principales espèces qui forment des rassemblements dans les eaux profondes sont l'hoplosthète orange (Hoplostethus atlanticus) et les oréos (Allocyttus spp., Neocyttus spp. Pseudocyttus spp., etc.), qui sont souvent pêchés ensemble, les béryx (Beryx spp.) dans les pêcheries situées dans les basses latitudes, la légine australe (Dissostichus eleginoides) dans les pêcheries de l'Océan Austral, la tête casquée pélagique (Pseudopentaceros wheeleri) et diverses espèces de Scorpaenidae que l'on trouve près des deux côtes de l'Amérique du Nord.
Loin des monts sous-marins, prédominent des poissons gadiformes, comme les macrouridés. Ces espèces tendent aussi à avoir une croissance lente, sans avoir des caractéristiques aussi «extrêmes» que celles qui sont associées aux pêcheries des monts sous marins. Les autres espèces qui peuvent être incluses dans ce groupe sont la morue charbonnière (Anoplopoma fimbria), le flétan noir (Reinhardtius hippoglossoides), les moridés (Moridae), les abadèches (Brotulidae) et les merlus (Merlucciidae). Certaines de ces espèces étaient généralement considérées comme sans marché, mais avec un système de commercialisation efficace et des précautions particulières lors de la transformation (à défaut de retirer tous les cérides cutanés des filets d'hoplosthète orange lors de la transformation, ce poisson peut donner la diarrhée), même les sceptiques ont été surpris par la percée de nombreuses espèces de grands fonds sur le marché.
Hoplosthète orange
Figure C3.2 - Captures nominales annuelles ('000t) d'Hoplostète orange
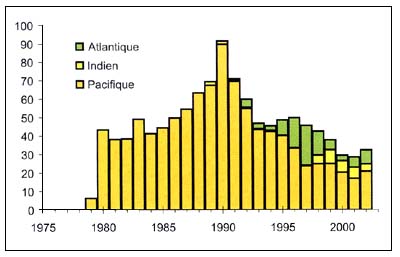
Source FAO
L'hoplosthète orange (Hoplostethus atlanticus) (Figure C3.2), est un Trachichthyidae présent dans le sud et le nord de l'Atlantique, le sud de l'Océan Indien, la mer de Tasmanie, la mer de Nouvelle-Zélande et le sud du Pacifique. Il est capturé à plus de 800 mètres de fond, par des navires qui visent généralement les rassemblements de ponte à proximité des monts sous-marins. Le pourcentage de la ressource présent dans d'autres zones est inconnu.
Du fait de la croissance lente et de la longévité exceptionnelle de cette espèce, la pêche conduit à un épuisement progressif du stock. Le recrutement semble être épisodique et la forte demande du marché stimule l'exploitation. Les taux d'exploitation durable sont de l'ordre de 5 pour cent de la biomasse vierge. Cette espèce est présente dans des stocks nationaux, transfrontaliers, chevauchants et de haute mer. La gestion du stock de haute mer est compromise par l'insuffisance des données disponibles.
Oréos
Figure C3.3 - Captures nominales annuelles ('000t) d'Oréos
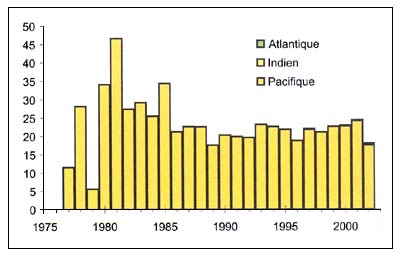
Source FAO
Les oréos (Allocystus spp. Neocystus spp. et Pseudocustus spp.) (Figure C3.3), sont des Oreostomadidae d'eau profonde qui vivent près du fond et forment de grands bancs au-dessus des fonds accidentés, près des monts et des canyons sous-marins. Les juvéniles sont pélagiques et vivent en haute mer. Ils sont généralement dispersés au-dessus des fonds lisses. Leurs œufs flottent près de la surface de la mer, et les larves habitent aussi les eaux superficielles. Dans les eaux australiennes, Neocyttus rhomboidalis est plus abondant à des profondeurs intermédiaires (600–800 m) et Allocyttus verrucosus dans les eaux plus profondes (900–1200 m). Ces deux espèces se nourrissent sur le fond de salpes, de crustacés, de poissons et d'encornets.
La gestion de ces pêcheries de haute mer est entravée par la difficulté d'obtenir les données nécessaires. D'après des données sur la détermination de l'âge provenant d'Australie et de Nouvelle-Zélande l'espérance de vie maximale est de 86 ans pour Pseudocyttus maculatus et de 153 ans pour Allocyttus niger . La mortalité naturelle du premier a été estimée à 0,063 année-1et celle du second à 0,044 année-1. Les estimations provenant de Nouvelle Zélande indiquent que la «PME» est de l'ordre de 1,6 pour cent de B0 si l'on veut éviter que la biomasse du stock tombe à un niveau inférieur à 0,2B0 (probabilité <0,2). Pour des raisons opérationnelles, s'ils font l'objet d'un aménagement, les oréos Pseudocyttus maculatus, Allocyttus niger et Neocyttus rhomboidalis peuvent être gérés comme un stock unique avec les dangers que cela comporte (Annala, Sullivan & O'Brien 1999).
Béryx
Figure C3.4 - Captures nominales annuelles ('000t) de Béryx
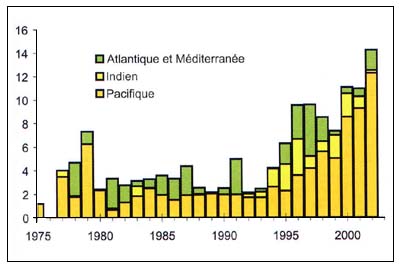
Source FAO
Les béryx longs (Beryx splendens), (Figure C3.4) sont des Bericidae distribués dans tous les océans, même s'ils se rencontrent rarement dans le Pacifique Nord-Est. Ils vivent en dehors du le plateau (180m) et sur le talus jusqu'à une profondeur d'au moins 1300 mètres, et s'éloignent probablement davantage du fond la nuit. Les Beryx splendens sont capturés au chalut pélagique au dessus des monts sous-marins peu profonds, des dorsales sous-marines et du rebord du talus, entre 300 et 500 mètres. Les juvéniles sont pélagiques. D'après une analyse génétique, il y a, ou il y a eu, des échanges génétiques entre les océans et ces poissons sont répartis dans toutes les mers. Il y a sans doute un réservoir de juvéniles dans l'Océan Austral qui approvisionne le recrutement vers le nord, créant ainsi des stocks génétiques uniques d'Allocyttus niger et de Pseudocyttus maculatus, mais formant des stocks écologiques distincts avec dispersion limitée des recrues (Lehodey, Grandperrin & Marchal 1997, Smith 2000).
Ces espèces vivent au maximum 17 ans et atteignent la maturité sexuelle à environ 4 ans. La mortalité naturelle est estimée à 0,23. Des pêcheries viables en eaux profondes peuvent donc très bien être ciblées sur ces espèces. On sait peu de choses sur la structure de ces espèces dans le stock local et il est vraisemblable que la pêcherie de Nouvelle Zélande exploite un stock plus grand dans le Pacifique Sud (Annala, Sullivan & O'Brien 1999).
Légine
Figure C3.5 - Captures nominales annuelles ('000t) de Légine
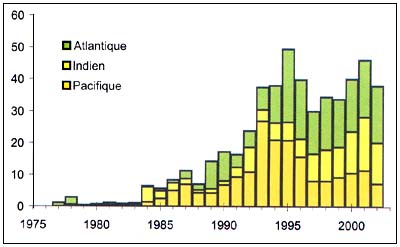
Source FAO
Les légines (Dissostichus spp.), (Figure C3.5) sont des Notothenidae à répartition circumpolaire, dans les eaux de l'Antarctique et de l'Océan Austral. La légine australe (D. eleginoides) se rencontre autour de la zone sud de l'Amérique du Sud alors que la légine antarctique (D. mawsoni) est présente à des latitudes plus élevées, surtout dans la région du Pacifique. Les deux espèces se chevauchent entre 60°S et 65°S et se rencontrent l'une et l'autre à des profondeurs de 3 000 m. La limite septentrionale de la plupart des populations de légines australes est 45°S, sauf le long des côtes du Chili et de l'Argentine où elles peuvent s'étendre au nord dans des eaux plus profondes et plus froides. Il existe d'importantes populations de légines australes dans les eaux de diverses îles sub-antarctiques ou dans les zones adjacentes ainsi que dans les eaux du Chili, de l'Argentine, de l'Uruguay et du Pérou.
L'ampleur des activités de pêche illicite, non déclarée et non réglementée a considérablement diminué durant la campagne de 2003–2004 (6342 tonnes dans la Zone statistique 41 et 3701 tonnes dans la Zone statistique 87), mais reste une préoccupation majeure. En outre, des captures de légines déclarées comme pêchées en dehors de la zone de compétence de la CCAMLR, ont très probablement été prélevées sur des stocks présents à l'intérieur de cette zone. Dans le passé, les stocks de légines ciblés par des flottilles de pêche illégales se sont amenuisés rapidement et de manière sensible. Par exemple, les ressources en légines des îles du Prince Édouard (Afrique du Sud) sont désormais très en deçà du niveau qu'elles avaient avant d'être exploitées.
Tête casquée pélagique
Figure C3.6 - Captures nominales annuelles ('000t) de Tête casquée pélagique
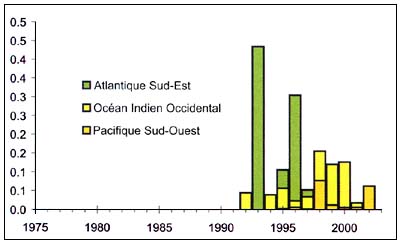
Source FAO
Les têtes casquées pélagiques (Pseudopentaceros wheeleri), (Figure C3.6) sont des Pentacerotidae associés aux monts sous-marins, en particulier dans le Pacifique Nord mais aussi dans d'autres océans, et pêchés au chalut de fond.
Leur pêche illustre bien le sort qui peut être réservé aux pêcheries associées aux monts sous-marins. Des navires japonais et russes ont commencé en 1969 à pêcher au chalut dans les zones de la Chaîne des volcans de l'Empereur et de la dorsale du nord des Hawaï. On ne connaît pas avec précision le volume total des captures des navires russes, mais il est estimé à plus de 133400 tonnes pour la période 1967–77. Les Japonais ont envoyé, entre 1969 et 1977, entre deux et cinq chalutiers par an dans cette zone qui ont capturé en moyenne de 22800 à 35100 tonnes par an, dont 90 pour cent de têtes casquées pélagiques. Par la suite les captures sont tombées à 5800 – 9900 tonnes entre 1977 et 1982. Les têtes casquées pélagiques, jadis dominantes ont été supplantées par le béryx long (Beryx splendens), mais leurs stocks n'ont jamais atteint l'abondance des têtes casquées pélagiques qu'ils ont supplantées et rien n'indique que l'un ou l'autre de ces stocks reviendra à des niveaux commercialement exploitables dans le futur proche.
Hoki
Le hoki (Macruronus novaezelandiae), (Figure C3.7) est un Macruronidae benthopélagique qui vit ordinairement près du fond, mais forme des rassemblements de ponte dans les eaux intermédiaire. Les gros poissons adultes se rencontrent généralement en dessous de 400 m, alors que les juvéniles sont présents plus près de la surface. Les pêcheries opérant au chalut pélagique ciblent les rassemblements de poissons près des canyons, souvent situés près des côtes dans des zones de plateau continental étroit.
Figure C3.7 - Captures nominales annuelles ('000t) de Grenadier bleu
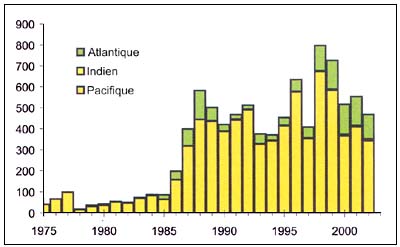
Source FAO
La structure des stocks est souvent mal connue, d'où la difficulté d'attribuer les TAC à des zones de pêche précises; dans certaines zones, les captures accidentelles de phoques et d'autres espèces non visées posent un problème. L'expérience de gestion dans au moins quelques juridictions indique que cette ressource peut être gérée de manière durable, quitte à modifier de temps à autre le TAC, dans la principale pêcherie mondiale de cette espèce, qui est la Nouvelle Zélande, pour l'ajuster à la taille des stocks de hoki, qui a fluctué entre 200000 et 250000 tonnes au cours des quinze dernières années. En2004, le TAC a été abaissé à 100 000 tonnes par suite d'une diminution du stock de reproducteurs, attribuée à des phénomènes océanographiques en rapport avec El Niño.
Autres espèces
Une autre série d'espèces de grands fonds, ou de talus, ont été ciblées par des pêcheries dans de nombreuses régions tropicales. Ces espèces peuvent être ciblées par des pêches artisanales en eaux profondes le long du rebord et du talus du plateau, partout où la plate-forme continentale est relativement étroite et où les fonds de pêche sont accessibles aux pêcheurs opérant à bord de petites embarcations. Les principales espèces appartiennent aux familles Lutjanidae, Serranidae, et Carangidae et les plus importantes sont les vivaneaux flamme et rubis (Etelis coruscans et E.carbunculus) et les lutianidés (Pristomopoides filamemtosus, P. typus et P. multidens). Ces pêcheries sont particulièrement importantes pour les petits états insulaires qui ont souvent peu d'autres ressources démersales, bien qu'elles soient largement distribuées le long des marges de nombreux continents des zones tropicales et subtropicales.
LE CYCLE BIOLOGIQUE DES POISSONS DE GRANDS FONDS
Les modifications particulières de la coloration, de la structure de la mâchoire, de la musculature, de la forme de la nageoire et de la vessie natatoire et des yeux chez les poissons des grands fonds, n'ont rien de surprenant vu les caractéristiques de leur habitat. En l'absence de lumière, la plupart des poissons de grands fonds sont noirs ou rouges car cette longueur d'onde est fortement atténuée dans l'eau de mer et ces poissons ont effectivement une couleur noire. Malgré l'absence de lumière, les poissons de grands fonds ont souvent de grands yeux et des adaptations particulières de la rétine et du cristallin pour détecter la lumière ambiante, qui vient habituellement des organes photophores présents sur les proies, les prédateurs ou les autres poissons de la même espèce.
De nombreuses espèces de grands fonds ont une masse musculaire et osseuse réduite, qui leur donne une flottabilité presque neutre: ceux qui chassent en croisant lentement ont des vessies natatoires qui peuvent excréter du gaz malgré les pressions hydrostatiques ambiantes d'environ 400 atmosphères présentes à de grandes profondeurs (Marshall 1954, 1979).
Les espèces de grands fonds doivent aussi réussir leur ponte, croître et revenir dans la zone qui leur sert d'habitat à l'âge adulte. Les conditions extrêmes des eaux profondes se reflètent dans la variété des stratégies de reproduction. Malgré la faible taille des populations, il y a des cas d'hermaphrodisme, de dimorphisme sexuel poussé à l'extrême et de sex-ratio déséquilibré. Bien que les espèces des grands fonds soient peu nombreuses, elles adoptent diverses stratégies de reproduction. Certaines espèces privilégiant nettement la stratégie k (peu d'œufs pondus et mortalité faible), elle peuvent être semelpares (Coryphaenoides armatus , un macrouridé largement distribué). D'autres étant ovovivipares et ovipares et d'autres encore privilégiant fortement la stratégie r (beaucoup d'œufs pondus et mortalité élevée). On estime que les bonnes classes d'âge peuvent être intermittentes et que de nombreuses espèces profondes pondent périodiquement au fil des années.
Un certain nombre de tactiques ont été adoptées pour réduire la dispersion des œufs. Les œufs flottants des macrouridés des grands fonds, qui ont une vaste aire de répartition, portent des marques en relief qui ralentissent leur ascension, une adaptation que n'ont pas les espèces de cette famille qui vivent dans des eaux moins profondes. Les œufs d'Hoplostethus atlanticus sont dans un premier temps flottants, puis ils coulent et terminent probablement leur cycle de développement sur le fond marin, ce qui facilite leur rétention dans l'habitat des adultes de cette espèce.
PRODUCTIVITÉ
L'un des principaux déterminants de la productivité d'une pêcherie est le taux de croissance et les distributions par taille des populations de poissons, qui sont presque toujours asymétriques et négatives. Toutefois, des distributions bimodales des tailles ont été observées chez certaines espèces des grands fonds. Cela s'expliquerait en partie par l'habitat peu productif, qui fait que les espèces profondes consacrent leur surplus énergétique à la croissance somatique jusqu'à atteidre une taille suffisante, et qu'ensuite seulement elles le consacrent à la reproduction.
Quelle que soit la raison, il est clair que de nombreuses espèces profondes ont une croissance si lente qu'il est difficile de déterminer leur âge réel, et lorsque cela a été fait, les résultats sont sujets à caution. Pour quelques espèces, en particulier l'hoplosthète orange (Hoplostethus atlanticus), la technique de détermination de l'âge appropriée a été violemment contestée et la longévité potentielle des espèces pouvait varier, selon les hypothèses de modélisation, de 21 ans à plus de 100 ans; la thèse aujourd'hui admise est que ces poissons vivent relativement veux, au maximum jusqu'à plus de100ans.
De nombreux biologistes spécialistes des pêches en eaux profondes estiment que cela n'a aucun sens de parler de durabilité à propos des pêches en eaux profondes, en raison de l'extrême lenteur des taux de croissance de beaucoup d'espèces d'eaux profondes et, circonstance aggravante, de leur recrutement apparemment épisodique. Dans ces pêcheries, si la ressource était constituée de classes d'âge bien distinctes qui sont pêchées jusqu'à ce que leur biomasse tombe à des niveaux d'abondance très faibles, la reprise des stocks découlant d'un bon recrutement s'étalerait nécessairement sur plusieurs générations dont la durée est supérieure à celle des hommes.
PÊCHES EN EAUX PROFONDES
Récemment encore, les zones profondes de la haute mer étaient difficiles à exploiter et comme les ressources étaient comparativement plus abondantes dans les mers moins profondes, la pêche hauturière ne présentait guère d'intérêt. Il existe peu de pêcheries en eaux profondes implantées depuis longtemps, mis à part la pêcherie palangrière portugaise (Madère) de sabre noir (Aphanopus carbo) et les pêcheries d'escoliers serpents (Gempylidae) et de ceintures d'argent (Trichiuridae) des îles du Pacifique, qui étaient initialement exploitées à l'échelle artisanale et ne constituaient pas une menace pour la durabilité de la ressource.
Parmi les pêches en eaux profondes qui se sont récemment développées, l'une des plus connues est celle d'hoplosthète orange, une espèce qui vit près des talus, des monts sous-marins et du fond marin, en particulier dans les eaux de la Nouvelle-Zélande et du Sud-Est de l'Australie, où son exploitation commerciale a commencé. On notera que l'hoplosthète orange a été décrit pour la première fois dans les eaux des Açores. Sa zone de pêche s'est par la suite étendue à la Crête de Walvis dans l'Atlantique Sud-Est (Namibie), au sud-ouest de l'Océan Indien et il existe une petite pêcherie dans la Golfe de Gascogne. Ce poisson longève atteint environ 40 cm et 2 kg à l'âge adulte, avec toutefois des variations de la taille maximale suivant les régions. Des techniques de chalutage contrôlé spécialisé ont été mises au point car, avec les techniques antérieures, des volumes considérables étaient prélevés en quelques minutes dans des rassemblements de ponte, si bien que les culs de chalut se cassaient et une partie des captures était perdue. L'hoplosthète orange repère immédiatement les objets qui s'approchent (une adaptation pour éviter la prédation des cachalots), de sorte que les évaluations acoustiques au moyen de transducteurs remorqués se sont avérées difficiles dans certaines zones. Les niveaux maximaux d'exploitation soutenable de l'hoplostète orange sont très faibles, de l'ordre de 5 à 10 pour cent de la biomasse non exploitée (M ~ 0.04). D'après les indices accumulés jusqu'à présent, il existe peu de pêcheries exploitées de manière durable et la continuité des rendements reste incertaine. Les plus petits stocks sont généralement voués à l'épuisement à partir du moment où ils sont ciblés, mais il semblerait, sans que ce soit prouvé, que d'autres stocks se soient avérés plus résistants que prévu à la surexploitation, probablement pour deux raisons: a) la pêche disperse le poisson avant que le stock ne soit épuisé et b) la ponte est épisodique. Dans ce cas, lorsque la pêcherie exploite des rassemblements de ponte, tout le stock ne peut pas être capturé au cours d'une année, car tous les poissons ne pondent pas tous les ans (Butterworth et Brandão 2005).
Une pêcherie de trichiuridés, exploitant Aphanopus carbo , est une des rares pêcheries en eaux profondes qui se soit avérée durable sur une période d'environ 150 ans, car elle reposait sur l'utilisation traditionnelle de lignes avec des hameçons. À l'âge adulte, cette espèce est benthopélagique et vit entre 400 et 1600 mètres de fond. Elle se rencontre du Groënland aux Îles Canaries et de part et d'autre de la dorsale médio-atlantique. Cette espèce d'eaux profondes a une croissance rapide et une longévité d'environ 8 ans. Toutefois, comme pour l'hoplosthète orange, les signaux habituels montrent que cette pêcherie est en danger. Les captures sont passées de 1100 tonnes en 1980 à 3000 tonnes en 1992 et l'efficacité des engins a augmenté grâce à l'introduction de lignes monofilament et à une forte augmentation du nombre d'hameçons par série de lignes (4000–5000 hameçon par ligne).
Les Macrouridae sont un autre groupe dont les membres sont largement distribués et abondants, dans certains endroits. Ces espèces typiquement pélagiques vivent dans la zone comprise entre le milieu et le haut du talus continental. Dans l'Atlantique Nord, des stocks de Macrourus berglax et Coryphaenoides rupestris , sont pêchés au chalut de fond, jadis à des profondeurs comprises entre600et 800 mètres, et plus récemment jusqu'à 1 500 m de fond. Toutefois, les pêcheries situées au large de Terre-Neuve sont caractérisées par un schéma bien connu, avec des totaux admissibles des captures reflétant un déclin général des débarquements déclarés. Les Coryphaenoides rupestris peuvent vivre 70 ans, mais dans l'Atlantique Nord-Est, les poissons sont généralement âgés de 20 à 30 ans. Ainsi, comme de nombreuses autres espèces d'eaux profondes, les macrouridés ont des caractéristiques qui les rendent particulièrement vulnérables à la surexploitation.
Figure C3.8 - Captures nominales annuelles ('000t) d'espèces profondes
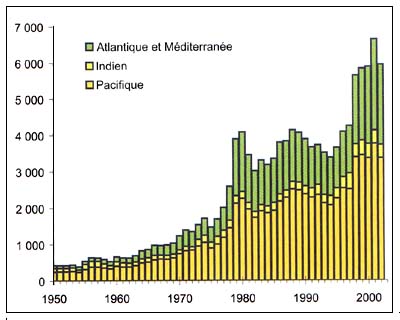
Source FAO
Les Pleuronectidae sont un groupe très évolué qui n'est habituellement pas pêché en eaux profondes, même s'il existe d'importantes pêcheries exploitant certaines espèces de ce groupe dans l'Atlantique Nord et dans le Pacifique Nord. Dans l'Atlantique, les plus connues sont celles de flétan noir (Reinhardtius hippoglosoides) dans les eaux profondes du talus continental. Alors que le poids moyen de ces poissons était de 1 kilo jusqu'au milieu des années 80, il n'était plus que de 200 grammes au début des années 90.
La figure C3.8 illustre l'évolution des captures des espèces de grands fonds recensées dans le Tableau C3.1. Certaines de ces espèces peuvent cependant aussi être capturées dans d'autres zones sans qu'il soit possible de déterminer le pourcentage des captures provenant de la haute mer. Ces statistiques reflètent seulement le volume des captures des espèces profondes que les pays membres déclarent à la FAO. On ne dispose d'aucune information permettant de dire si les changements survenus en 1979 et en 1998 sont attribuables à une augmentation des captures effectives ou à des quantités déclarées plus conformes à la réalité.
LA GESTION DES RESSOURCES
Les caractéristiques biologiques des espèces d'eaux profondes que l'on peut capturer dans une même pêcherie sont variées et requièrent une adaptation des stratégies d'aménagement. Différents groupes d'espèces peuvent être présents dans la même zone mais à des profondeurs différentes. Ainsi, dans le sud de l'Océan Indien, les pêcheries opérant à des profondeurs de 800 mètres, sont ordinairement ciblées sur l'hoplosthète orange (Hoplostethus atlanticus), et sur les oréos Allocyttus niger, Neocyttus rhomboidalis et Pseudocyttus maculatus. À des profondeurs de l'ordre de 400m, les mêmes navires pêchent ordinairement le béryx long (Beryx splendens), la tête casquée pélagique (Pseudopentaceros richardsoni), le poisson cardinal (Epigonus telescopus) et la rouffe antarctique (Hyperoglyphe antartica).
Ces pêcheries se caractérisent par la faible importance des captures accessoires (et donc des rejets), et peuvent donc être considérées comme «propres» en ce sens que peu d'espèces non recherchées sont prises. Cependant, les opérateurs non qualifiés peuvent endommager gravement le benthos, en raclant le fond avec les chaluts. En outre la perte de poissons par les culs de chalut cassés, entraîne une mortalité par pêche qui est rarement déclarée.
Dans ces pêcheries, les instruments traditionnels de gestion des pêches, qui se fondent sur des hypothèses de recrutement annuel et sur des relations fonctionnelles stock-recrutement, peuvent aller à l'encontre de l'effet recherché et mettre en péril la durabilité des ressources, au lieu d'assurer leur conservation. Bien que les ressources des eaux profondes soient pêchées au chalut, du point de vue de l'aménagement, les tactiques de pêche s'apparentent davantage à la pêche à la senne. La pêche peut se faire sur les détections du sondeur, en particulier dans le cas de l'hoplostète orange, ou par chalutages prolongés, comme dans les opérations ciblées sur le béryx. Dans la pêche à l'hoplosthète orange, le succès des captures dépend beaucoup plus des capacités de l'équipage et de l'efficacité des tactiques de pêche que d'autres considérations comme les dimensions du navire. Lorsque l'on enregistre l'effort de pêche, il suffit de savoir si le filet est un chalut de fond ou un chalut pélagique et dans ce dernier cas, si le chalut a raclé le fond et si l'engin s'est comporté comme prévu, c'est-à-dire si le trait a été «réussi» même si aucun poisson n'a été pris. Cela nous amène aux tactiques de pêche en présence d'autres navires. Les navires qui ont à leur bord des équipages peu qualifiés suivent souvent des bateaux plus performants sur lesquels ils comptent pour localiser les concentrations de poissons. Dans ces cas-là, il peut être encore plus difficile de se prononcer sur le succès des opérations d'un navire, à cause des stratégies complexes qu'adopte le patron de pêche pour leurrer ses concurrents. L'indice des Captures par unité d'effort (CPUE) reste l'un des instruments les plus utiles pour évaluer l'état d'un stock, mais dans ces pêcheries vulnérables, il pourrait être trompeur, sinon dangereux, de lui accorder trop d'importance.
Tableau C3.1 Espèces de poissons des grands fonds figurant dans la base de données sur les débarquements communiqués à la FAO
| Crustacés | ARISTAEIDAE | Plesiopenaeus edwardsianus | Gambon écarlate |
| GERYONIDAE | Geryon quinquedens | Gériocrabe rouge | |
| LITHODIDAE | Lithodes aequispina Paralomis spinosissima | Crabe royal doré Crabe royal de l'Antarctique | |
| PANDALIDAE | Pandalus spp | Crevettes Pandalus nca | |
| SOLENOCERIDAE | Pleoticus robustus | Salicoque royale rouge | |
| Chondricthyens | CALLORHINCHIDAE | Callorhinchus capensis Callorhinchus milii Callorhinchus spp | Masca du Cap Masca laboureur Mascas nca |
| Chimaeras, etc. | Chimaeriformes | Chimères, etc. nca | |
| CHIMAERIDAE | Chimaera monstrosa Hydrolagus novaezealandiae Hydrolagus spp | Chimère commune Hydrolagus novaezealandiae Chimères nca | |
| HEXANCHIDAE | Hexanchus griseus | Requin griset | |
| RAJIDAE | Bathyraja meridionalis Bathyraja spp | Bathyraja meridionalis Raies bathyraja nca | |
| SQUALIDAE | Centroscymnus coelolepis Centroscymnus crepidater Somniosus microcephalus Somniosus pacificus | Pailona commun Pailona à long nez Laimargue du Groenland Laimargue dormeur | |
| Téléostiens | ARGENTINIDAE | Glossanodon semifasciatus | Argentina du Pacifique |
| BERYCIDAE | Beryx spp Centroberyx affinis | Béryx nca Centroberyx affinis | |
| BRANCHIOSTEGIDAE | Branchiostegidae | Tiles nca | |
| CAPROIDAE | Caproidae | Sangliers nca | |
| CENTROLOPHIDAE | Hyperoglyphe antarctica Seriolella caerulea | Rouffe antarctique Seriolella caerulea | |
| CHLOROPHTHALMIDAE | Chlorophthalmidae | Verdyeux | |
| EMMELICHTHYIDAE | Emmelichthyidae Emmelichthys nitidus | Andorrèves, poissons rubis nca Andorrève du Cap | |
| EPIGONIDAE | Epigonus spp | Poissons-cardinaux nca | |
| GADIDAE | Brosme brosme Micromesistius australis Micromesistius poutassou Molva dypterygia Molva molva | Brosme Merlan bleu austral Merlan bleu Lingue bleue Lingue | |
| GEMPYLIDAE | Lepidocybium flavobrunneum Rexea solandri Ruvettus pretiosus Thyrsitops lepidopoides | Escolier noir Escolier tifiati Rouvet Escolier blanc | |
| GERYONIDAE | Geryon spp | Géryons nca | |
| MACRORAMPHOSIDAE | Macroramphosus scolopax | Bécasse de mer | |
| MACROURIDAE | Coryphaenoides rupestris Lepidorhynchus denticulatus Macrouridae Macrourus berglax | Grenadier de roche Lepidorhynchus denticulatus Macrouridae Grenadier berglax | |
| Macrourus spp | Grenadiers nca | ||
| MERLUCCIIDAE | Macruronus magellanicus Macruronus novaezelandiae Macruronus spp | Grenadier de Patagonie Grenadier bleu Grenadiers bleus nca | |
| MORIDAE | Antimora rostrata Mora moro | Antimora bleu Moro commun | |
| MYCTOPHIDAE | Lampanyctodes hectoris Myctophidae | Lanternule de Hector Lanternules nca | |
| NOTOTHENIIDAE | Dissostichus eleginoides Dissostichus mawsoni | Légine australe Légine antarctique | |
| OPHIDIIDAE | Genypterus blacodes Genypterus capensis Genypterus chilensis Genypterus maculatus Genypterus spp | Abadèche rosé Abadèche du Cap Abadèche rouge Abadèche noir Abadèches nca | |
| OREOSOMATIDAE | Oreosomatidae | Oréos nca | |
| PENTACEROTIDAE | Pseudopentaceros richardsoni | Tête casquée pélagique | |
| PLEURONECTIDAE | Reinhardtius hippoglossoides | Flétan noir | |
| POLYPRIONIDAE | Polyprion americanus Polyprion oxygeneios | Cernier commun Cernier de Nouvelle-Zélande | |
| SCORPAENIDAE | Scorpaena scrofa | Rascasse rouge | |
| STERNOPTYCHIDAE | Maurolicus muelleri | Maurolicus muelleri | |
| SYNODONTIDAE | Harpadon nehereus | Scopelidé | |
| TRACHICHTHYIDAE | Hoplostethus atlanticus Trachichthyidae | Hoplosthète orange Poissons-montres nca | |
| TRICHIURIDAE | Aphanopus carbo Lepidopus caudatus Trichiuridae Trichiurus lepturus | Sabre noir Sabre argenté Poissons-sabres, sabres nca Poisson-sabre commun | |
| TRIGLIDAE | Pterygotrigla picta | Pterygotrigla picta |
Les recherches sur les espèces de grands fonds, en particulier de haute mer, sont aussi entravées par la difficulté d'obtenir des données essentielles pour l'aménagement des ressources (FAO 2001, 2002). Les opérateurs de nombreux navires de pêche hauturière ne sont pas tenus de déclarer à l'État du pavillon leurs captures, leur effort de pêche ou leur zone d'opérations. Si les captures sont débarquées dans des pays étrangers, l'enregistrement des données est encore plus difficile. C'est notamment le cas pour la pêche à la tête casquée pélagique (Pseudopentacerus wheeleri) dans le Pacifique Nord, dont on ne trouve aucune trace dans la Base de données de la FAO sur les captures et les débarquements en valeur nominale, qui est l'archive mondiale la plus importante et la plus complète des captures de poissons. Les captures déclarées de cette espèce atteignent pourtant 200 000 tonnes par an! De même, pour la période prenant fin en 2000, la base de données de la FAO signale des débarquements officiels provenant de trois pays seulement, alors qu'il ressort de contacts informels par l'intermédiaire de groupes de travail qu'au moins sept pays ont exploité cette espèce dans l'Océan Indien. Dans certains cas, ce manque d'informations est une conséquence de la législation nationale sur la confidentialité de l'information. En général, seuls quelques bateaux, voire un seul, peuvent participer à des pêcheries associées à des monts sous-marins à l'intérieur d'une zone d'aménagement, et si cette zone repose sur un stock, elle peut-être constituée d'un seul mont sous-marin. En outre, les lois nationales empêchent souvent les gouvernements de diffuser des informations qui identifient les opérations d'un navire ou d'une compagnie de pêche spécifique. Cela peut compromettre définitivement toute possibilité de gestion efficace des ressources.
OBJECTIFS DE GESTION: LE «RENDEMENT DURABLE» EST-IL UN OBJECTIF RÉALISTE?
Les systèmes d'aménagement axés sur un objectif de «rendement durable» sont de plus en plus critiqués dans les pêcheries conventionnelles depuis près de deux décennies, car de nombreuses administrations des pêches se sont efforcées de mettre en œuvre ce paradigme d'aménagement. Cela a conduit à réorienter la gestion vers l'utilisation de points de référence, considérés comme des outils à la fois plus simples, plus maniables et plus utiles. L'approche de gestion basée sur l'écosystème, préconisée depuis peu, est une évolution complémentaire et plus complexe de cette démarche. Les caractéristiques de nombreux poissons des grands fonds font qu'il est extrêmement difficile, sinon impossible, d'adapter ces modèles au contexte des pêcheries hauturières. Premièrement, la «durabilité» peut être un concept d'aménagement peu pertinent ou du moins inutilisable dans des pêcheries où les ressources ont un recrutement épisodique et imprévisible, quelle que soit la taille du stock de reproducteurs. Deuxièmement, avec des espèces qui ont une croissance lente et une maturité sexuelle tardive, les niveaux de référence limites conventionnels de la biomasse féconde, ne seraient applicables qu'à un petit nombre de stocks, et dans le cas où ils pourraient être utilisés, les taux d'exploitation devraient être de l'ordre de 5pour cent de la biomasse vierge. Troisièmement, les temps de reconstitution des stocks peuvent s'étaler sur plusieurs décennies, ou même au-delà des horizons temporels commerciaux (et politiques). Quatrièmement, même en cas d'accord sur les objectifs d'aménagement et les fonctions de contrôle de l'exploitation, bien souvent, aucun régime d'aménagement ou mécanisme de réglementation n'est en place pour garantir l'application du plan d'aménagement. À défaut de résoudre ces problèmes, on risque de voir disparaître les ressources des grands fonds, qui contribuent au bien être de l'humanité.
GESTION OPÉRATIONNELLE: ÉVALUATION ET GESTION DE L'EXPLOITATION DES PÊCHERIES HAUTURIÈRES
Cette section est extraite du rapport de l'atelier de la FAO sur l'évaluation et la gestion des pêches en eaux profondes, tenu à l'Université d'Otago, à Dunedin, (Nouvelle-Zélande) du 27 au 29 novembre 2003 (Shotton, 2005)
Estimation de l'abondance
L'estimation de l'abondance des ressources halieutiques des eaux profondes a plusieurs points communs avec celle des stocks plus traditionnels de plateau, mais elle présente aussi des particularités spécifiques aux espèces d'eaux profondes. Certaines de ces espèces, comme l'hoplostète orange, ont une faible productivité, une forte tendance à se regrouper et elles se rencontrent souvent, mais pas toujours, à proximité des reliefs sous-marins; en outre leurs réactions aux prospections et aux engins de pêche ne se limitent pas au moment de la capture, mais se prolongent dans le temps. Les raisons du comportement grégaire (ex: conditions propices ou comportement acquis) restent mal connues et il est important de poursuivre les recherches sur le comportement des poissons. Cependant, la faible productivité en eaux profondes n'est peut-être pas universelle. Certaines espèces «d'eaux profondes» (en particulier celles qui ont des phases pélagiques prolongées, des habitudes de migration diurne marquées ou une préférence pour les profondeurs supérieures à 800 mètres) ont des niveaux de productivité modérés.
Les méthodes d'estimation de l'abondance basées sur l'étude des œufs, auxquelles on a souvent recours pour les stocks des eaux peu profondes, peuvent être peu efficaces pour les espèces profondes, pour diverses raisons, dont le comportement de ponte, qui peut être très localisé chez certaines espèces (par exemple l'hoplosthète orange), ou panmictique à l'échelle de la région chez d'autres (certains oréos et le sanglier). Les campagnes de chalutage et les prospections acoustiques sont probablement les méthodes les plus appropriées et elles peuvent toutes les deux être utilisées pour obtenir des indices de l'abondance relative; l'utilisation de méthodes acoustiques à cette fin pose cependant des problèmes lorsque l'espèce visée est associée à d'autres espèces au moment de la prospection. En revanche, les deux méthodes présentent des inconvénients pour estimer l'abondance absolue. Pour les estimations acoustiques, les difficultés consistent à estimer les propriétés réfléchissantes des espèces et à identifier leurs détections. Avec les campagnes de chalutage, c'est l'évaluation de la vulnérabilité à la capture (ou du coefficient de capturabilité) qui pose problème.
On peut utiliser des véhicules télécommandés et des submersibles autonomes pour estimer l'abondance ou dans des expériences visant à estimer la vulnérabilité à la capture par chalut, mais ces méthodes doivent être perfectionnées. L'efficacité des méthodes basées sur des progrès technologiques dépendra cependant d'une meilleure compréhension du comportement des poissons (comportement inné et réactions aux navires de recherche et aux engins de pêche). L'étude des facteurs qui influencent le comportement grégaire doit notamment être approfondie et ces technologies sous-marines peuvent être utiles à cet égard.
D'autres problèmes sont liés à la détermination de la structure, de la distribution et des déplacements des stocks. L'abondance relative ou absolue peut être estimée d'une manière ou d'une autre, mais la validité des estimations repose sur des plans de prospections appropriés qui tiennent compte des distributions et des mouvements des espèces. Pour l'instant, il n'existe pas de moyen optimal unique pour estimer l'abondance, et la combinaison d'un ensemble de méthodes semble être la meilleure démarche. Dans certaines zones, des campagnes de chalutage et des prospections acoustiques sont effectuées régulièrement et les estimations et les indices qui en résultent sont utilisés pour évaluer les stocks avec une efficacité variable, mais qui dépend souvent de la disponibilité et de la qualité des autres sources de données.
Les enquêtes sur les ressources halieutiques des eaux profondes coûtent cher et leurs coûts-avantages est systématiquement pris en considération dans le choix des méthodes les plus appropriées dans une circonstance particulière. Les campagnes de prospection spécifiques sont le moyen le plus efficace pour obtenir des estimations de l'abondance et on peut souvent en profiter pour étudier d'autres aspects des stocks. Cependant, les campagnes de prospections acoustiques fournissent des données utiles pour les évaluations des stocks et des progrès considérables sont faits dans cette direction.
Biologie, âge et croissance
La gestion des espèces d'eaux profondes est complexe car leurs caractéristiques biologiques sont très diverses. De nombreuses espèces ont une grande longévité, une croissance lente, une maturité tardive et une faible fécondité et beaucoup, mais pas toutes, ont une productivité annuelle relativement faible par rapport à leur biomasse. Cela signifie que ces stocks ont des niveaux de rendement durable relativement bas et que la reconstitution des stocks épuisés peut être lente.
La collecte et l'analyse de données doivent être faites de façon régulière et pas seulement à l'occasion des évaluations des stocks, pour les faciliter. Les caractéristiques biologiques (telles que la taille à la maturité) peuvent changer au fur et à mesure que la taille du stock évolue, d'où la nécessité de suivre leur évolution, qui peut fournir des renseignements utiles pour évaluer l'état d'un stock. Les effets dépendant de la densité doivent aussi être pris en considération, car les estimations de la capturabilité, de la maturité, des taux de croissance, de la réussite de la ponte et d'autres facteurs peuvent changer. Les structures des stocks de la plupart des espèces profondes sont généralement mal connues et les méthodes de suivi direct de leur distribution et de leurs déplacements (notamment par marquage) sont rarement applicables dans des habitats aussi profonds.
Les paramètres de population des poissons sont souvent mal connus et variables. En outre, l'utilisation des modèles de ponte pose des problèmes car plusieurs espèces d'eaux profondes (comme l'hoplosthète orange et le hoki) ne pondent pas tous les ans. Les données sur l'état des gonades ne sont donc pas suffisamment fiables pour déterminer l'âge de maturité ou l'ogive de maturation. Dans le cas de l'hoplosthète orange, une zone de transition dans les otolithes semble correspondre au début de la ponte. Cependant, la fréquence de la ponte est importante pour déterminer chaque année la proportion de la population, ou de la biomasse, disponible pour la pêche, surtout si cette proportion varie d'une année sur l'autre. On connaît mal les relations stock-recrutement et les niveaux de recrutement de la quasi-totalité des espèces des grands fonds. Ces informations sont pourtant importantes pour comprendre les variations de la taille des stocks et gérer les pêches de manière durable.
Grâce aux méthodes chimiques et radiométriques, l'interprétation des anneaux de croissance des otolithes des espèces d'eaux profondes et les méthodes basées sur l'âge sont devenues plus fiables. À défaut de données initiales, on peut utiliser des estimations générales de l'âge et de la croissance provenant d'autres zones ou pêcheries. La détermination de l'âge reste cependant nécessaire pour estimer la production dans les grandes pêcheries.
Il est absolument indispensable d'essayer de comprendre les processus écologiques influençant la variabilité biologique si l'on veut obtenir des modèles prévisionnels fiables. Il convient à tout le moins d'étudier les interactions trophiques et les liens entre les poissons démersaux des grands fonds et les sources d'énergie mésopélagiques.
Évaluation des pêches hauturières
Un certain nombre de données essentielles manquent et sont nécessaires pour faire des évaluations fiables des espèces des grands fonds. Les trois principales sont, dans l'ordre, les suivantes:
- Données sur les captures: il faut, au moins connaître le volume et la localisation des captures. Ces données doivent être recueillies à une échelle spatiale appropriée compte tenu des rassemblements localisés que forment de nombreuses espèces d'eaux profondes. Il serait également utile de réunir des données sur les distributions de fréquence de longueur des espèces cibles et des espèces des prises accessoires.
- Indices valides de l'abondance relative - les indices indépendants des pêches sont préférables mais il s'agira plus vraisemblablement des taux de captures commerciales d'une pêcherie. Il est extrêmement difficile de mettre au point des interprétations «valides» des indices à l'ouverture d'une pêcherie, mais des programmes devraient être mis au point pour recueillir des données sur des facteurs pertinents comme la relation entre les CPUE commerciales et l'abondance, la sélectivité de la pêche, les limites géographiques d'un stock, les habitudes de migration et la dynamique du comportement grégaire.
- Estimations de l'abondance absolue - ces données sont indispensables pour estimer les rendements soutenables à long terme. Elles peuvent provenir d'une enquête indépendante des pêches qui fournit des estimations de l'abondance absolue (par exemple une prospection acoustique) ou d'un modèle d'évaluation estimant l'abondance absolue sur la base des captures et de l'abondance relative.
D'autres données sont cruciales pour certains stocks et presque toujours utiles pour améliorer l'exactitude et la précision des éventuelles évaluations disponibles, notamment:
- Les estimations du recrutement relatif ou absolu;
- Les données sur la relation entre la taille du stock et le recrutement;
- es données sur l'âge.
S'il est essentiel d'avoir des informations sur l'âge pour estimer la productivité de la population, il est moins utile de consacrer des ressources à la détermination de l'âge des stocks, si l'on dispose déjà de données sur l'âge et la croissance d'autres stocks de la même espèce. La connaissance de l'âge des poissons (qui permet d'estimer les taux de croissance, la mortalité naturelle et parfois le recrutement) a été cruciale pour déterminer les rendements soutenables des pêcheries d'hoplosthète orange, au moment de leur établissement. Mais il a fallu attendre qu'un système de détermination de l'âge fiable soit mis au point pour que l'on sache de façon certaine que l'hoplosthète orange avait des taux de croissance anormalement faibles, une mortalité naturelle basse, une maturité tardive et une grande longévité. La détermination de l'âge d'autres populations d'hoplostète orange a mis en évidence quelques différences dans ces paramètres du cycle biologique, mais les écarts n'étaient pas suffisamment marqués pour justifier des études plus approfondies sur l'âge, surtout en présence de moyen limités. Il peut être préférable de privilégier d'autres besoins d'informations, comme les indices de l'abondance, et de se contenter d'adopter les clés d'identification âge-longueur, les paramètres de croissance et les estimations de la mortalité naturelle d'autres stocks similaires.
Enfin, pour atteindre des objectifs qui sont souvent imposés par des accords internationaux ou des politiques nationales, il peut être nécessaire de renforcer les programmes existants ou d'en mettre sur pied de nouveaux, pour fournir des données à l'échelle de l'écosystème, comme les espèces des prises accessoires, d'autres espèces associées et les effets de la pêche sur l'habitat. Dans les pêcheries multispécifiques, dans lesquelles il est extrêmement difficile d'obtenir des données sur les captures et l'abondance des espèces individuelles, les espèces peuvent être évaluées et gérées comme des assemblages d'espèces, en appliquant l'approche de précaution pour déterminer les limites des captures ou de l'effort.
Problèmes de gestion des pêches hauturières
Points de référence biologiques
Les diverses possibilités d'utilisation des points de référence reposent sur l'adoption de réglementations visant à établir des objectifs ou des limites et des procédures d'aménagement combinant des systèmes d'évaluation et d'aménagement et diverses mesures des performances pour évaluer les résultats. En général, il est recommandé de recourir à des méthodes d'évaluation pour en déduire des procédures d'aménagement, mais quand il existe de nombreuses pêcheries sur lesquelles on dispose de peu d'informations, ce n'est pas toujours possible. Pour de nombreux stocks peu productifs, les niveaux cibles de la biomasse utilisés aux fins de l'aménagement varient entre 30 et 55 pour cent ou plus du niveau de la biomasse vierge et, plutôt que de fixer des objectifs, il est préférable de chercher à éviter une probabilité élevée (en général 90 pour cent) que la biomasse ne tombe à un niveau inférieur (en principe 20 pour cent de la biomasse vierge). En fonction de l'incertitude inhérente à toute évaluation et projection, la biomasse cible variera, mais elle devrait rester dans la fourchette des objectifs normalement adoptés. À cet égard, certains scientifiques donnent cependant la préférence à une approche basée sur des points de référence de la mortalité par pêche.
Approche de la gestion des pêches axée sur l'écosystème
Même dans le domaine de la pêche en eaux profondes, la première étape fondamentale pour progresser vers une approche écosystémique de l'aménagement des pêcheries consiste à contrôler la mortalité par pêche de chaque espèce pour la maintenir en dessous d'un certain niveau. Cela nécessite de mieux intégrer l'évaluation et la gestion des ressources marines avec des cadres d'aménagement appropriés propres à garantir la bonne gestion du stock, tout en tenant compte des problèmes de l'ensemble de l'environnement ou de l'écosystème.
Cas où les données sont insuffisantes
Dans de nombreuses pêcheries en eaux profondes, il n'y a pas assez de données disponibles pour définir les captures initiales, évaluer l'état des stocks ou estimer les points de référence. Les mesures d'aménagement adaptatives peuvent s'avérer utiles mais il y a souvent des antagonismes entre d'une part ces mesures fondées sur l'observation des données en situation, qui nécessitent de tester les variations des niveaux de capture, et d'autre part l'approche de précaution qui préconise une réduction des captures des stocks à faible productivité. Les procédures de gestion adaptative «éprouvées» peuvent aussi être remises en question si l'information qui en dérive montre que les hypothèses sur lesquelles se fondent la gestion et la modélisation des ressources sont insoutenables et doivent être reconsidérées. Dans ces situations, des méta-analyses peuvent permettre d'exploiter au maximum les données mondiales dispersées sur les stocks de grands fonds, les pêches en eaux profondes et les habitats associés. Une telle analyse pourrait permettre d'établir des limites de captures initiales pour les nouvelles pêcheries, qui serviraient de base pour une approche de gestion adaptative et/ou de précaution.
Approche de précaution
L'approche consistant à avancer prudemment et à limiter les captures dès les premiers stades d'une pêcherie en eaux profondes a beaucoup de partisans. En Nouvelle-Zélande, la pêche de l'hoplostète orange est volontairement plafonnée à 500 tonnes pendant tout le temps où l'on réunit des informations. La Commission pour la conservation de la faune et la flore marines de l'Antarctique (CCAMLR) ne permet pas d'établir de nouvelles pêcheries à moins qu'elles ne soient expressément autorisées ou que leur autorisation ne soit subordonnée à des conditions précises, telles que les procédures de collecte de données obligatoires. Malgré le soutien croissant dont bénéficient ces approches, on note un déclin rapide de la biomasse des stocks qui ont une faible productivité car leurs biomasses initiales ont presque toujours été surestimées.
CONSIDÉRATIONS RELATIVES À L'ÉCOSYSTÈME DE LA HAUTE MER
Étant donné sa profondeur, son étendue et, jusqu'à une période très récente, sa connaissance faible ou nulle, les problèmes de l'environnement de la haute mer n'ont pas retenu l'attention, ce qui n'a rien de surprenant. Cependant, il est aujourd'hui tout à fait évident que l'activité humaine dégrade le fond marin, et dans le même temps, l'habitat des poissons de grands fonds. Ces dégâts sont principalement causés par le passage des chaluts sur le benthos (Koslow et al. 2000) et par le déversement dans les océans des déchets et boues; en outre, les activités de prospection pétrolière suscitent des préoccupations croissantes, et ce sera probablement bientôt aussi le cas pour les activités d'extraction de minéraux.
Les prospections de haute mer ont montré qu'il existait des coraux tout au fond des mers dans l'Atlantique Nord, le Pacifique Nord, les eaux du sud de l'Australie et de la Nouvelle-Zélande et il y en a presque certainement ailleurs aussi. Dans l'Atlantique Nord, le genre dominant Lophelia se rencontre à des profondeurs comprises entre 200 et 1000 mètres et croît au rythme de 1 mm par an. Les récifs les plus hauts ont atteint 35 mètres, ce qui correspond à un âge de 8500 ans et les plus grands récifs de Lophelia font plus de 13 km de long et jusqu'à 400 mètres de large. Au large de la Nouvelle-Écosse on trouve des coraux loin du rivage à plus de 200 mètres de fond. Là, les espèces connues sont Paragorgia arborea (Gorgonacea), qui peut atteindre cinq mètres de haut et Primnoa resedaeformis (Paragorgia), qui, d'après les rapports des pêcheurs, forme des blocs denses, dans les endroits où ils n'ont pas été détruits par le chalutage démersal.
Ces récifs fournissent un abri aux poissons marins qui dans l'Atlantique Nord Est, comprennent des espèces commerciales comme le sébaste, le lieu noir, la morue, la lingue et le brosme. Les récifs ont toujours été de riches fonds de pêche exploités à la palangre et aux filets maillants et des campagnes de pêche expérimentale ont démontré la présence de fonds de pêche productifs à proximité des coraux profonds. La conservation de la structure de ces habitats coralliens profonds est considérée comme vitale pour maintenir la production halieutique et, au moins dans les eaux moins profondes, on réalise que la dégradation des habitats par les chaluts de fond peut avoir des effets pervers sur la succession des espèces de poissons, au profit d'espèces de moindre valeur marchande.
Les coraux profonds sont fragiles et très sensibles au chalutage de fond; dans les eaux norvégiennes on estime notamment qu'entre 30 et 50 pour cent des récifs de Lophelia ont été détruits ou abîmés par le passage des chaluts. Des dégâts ont aussi été signalés dans d'autres régions de l'Atlantique Nord Ouest, notamment au Nord Ouest de l'Écosse et dans la plaine porcupine, dans les eaux irlandaises, ainsi que près de la côte de la Nouvelle-Écosse où les récifs coralliens profonds seraient volontairement «défrichés». En Australie et en Nouvelle-Zélande, les chaluts démersaux auraient cassé les coraux des crêtes rocheuses des monts-sous-marins en les raclant, provoquant la disparition de l'abondante faune ichtyologique qui y proliférait.
Les monts sous-marins sont un autre habitat particulier et important pour les poissons. Ces reliefs se rencontrent généralement au fond des océans et sont des vestiges de volcans éteints. Leur altitude est comprise entre 500 et 1000 mètres au-dessus du fond marin; certains montent jusqu'à quelques dizaines de mètres de la surface alors que d'autres ont leur sommet à plus d'un kilomètre de profondeur. La faune associée aux monts sous-marins est mal connue et très diversifiée et elle peut comprendre des coraux, des plumes de mer, des éponges, des araignées de mer et des langoustes. Bon nombre de ces espèces sont endémiques à des monts sous-marins spécifiques qui forment des habitats isolés. Lorsque ces habitats ont été endommagés par des engins de pêche, il leur faut plusieurs décennies, ou même plusieurs siècles pour se reconstituer.
RÉFÉRENCES
Andriyashev, A.P. 1953. Ancient deepwater and secondary deepwater fishes and their importance in a zoogeographical analysis. In «Notes on Special Problems in Icthyology. Akademiya Nauk SSSR, Ikhtiologicheskaya Kommossiya, Moscou: 58–64. (traduction anglaise par A.R. Gosline, Bureau of Commercial Fisheries, USNM, Washington)
Annala, J. H., Sullivan, K.J., et O'Brien, C.J. 1999. Report from the Fishery Assessment Plenary, April 1000: stock assessments and yield estimates. Ministry of Fisheries. 430p.
Butterworth, D.S. et Brandão, A. 2005. Experiences in Southern Africa in the Management of Deepsea Fisheries.In Proceedings of the Conference on Management and Governance of Deepwater Fisheries 1–4 décembre 2003, Queenstown, Nouvelle-Zélande. Sous presse.
FAO. 2001. Report of the Ad Hoc Meeting on Management of Deepwater Fisheries Resources of the Southern Indian Ocean, Swakopmund, Namibie, 30mai–1 juin 2001. FAO Fisheries Report, No. 652: 61p.
FAO. 2002. Report of the Second Ad Hoc Meeting on Management of Deepwater Fisheries Resources of the Southern Indian Ocean, Fremantle, Australie de l'Ouest, 20–22 mai 2002. FAO Fisheries Report, No. 672: 106p.
Shotton, R. (éd), 2005. Report on DeepSea 2003; Conference on the Governance and Management of Deep-sea Fisheries. Part 1: Queenstown, Nouvelle-Zélande , 1–5 décembre 2003 FAO Fisheries Proceedings, No. 3/1. Rome , FAO. 2005. 718p.
Garibaldi, L et Limongelli L. 2002. Trends in oceanic captures and clustering of large marine ecosystems: two studies based on the FAO capture database. FAO Fisheries Technical Paper, No. 435: 71p.
Koslow, A.J., Boehlert, G.W., Gordon, J.D.M., Haedrich, R.L., Lorance, p. et Parin, N. 2000. Continental slope and deepsea fisheries: implications for a fragile ecosystem. ICES J. Mar. Sci 57:548–557.
Lehodey, P., Grandperrin, R. et Marchal, p. 1997. Reproductive biology and ecology of a deep-demersal fish, alfonsino Beryx splendens, over the seamounts off New Caledonia. Mar. Biol . 128(1):17–27.
Marshall, N.B. 1954. Aspects of Deepsea Biology. Hutchinson Press, Londres. 380p.
Marshall, N.B. 1979. Developments in Deepsea Biology. Blandford Press, Poole. 566p.
Merrett, N.R. et Haedrich, R.L. 1997. Deepsea Demersal Fish and Fisheries . Chapman et Hall. Londres. 282p.
Smith, P.J. 2000. Managing biodiversity: Invertebrate by-catch in seamount fisheries in the New Zealand Exclusive Economic Zone. NIWA, Wellington, Nouvelle-Zélande. 30p.
Jorge Csirke et Marcelo Vasconcellos *
INTRODUCTION
L'extrême variabilité de l'abondance et des captures de la plupart des stocks de poissons marins sauvages suscite de grandes inquiétudes parmi les pêcheurs, les scientifiques et les administrateurs des pêches qui s'en occupent. Cette instabilité rend la planification difficile, même à court terme, car dans ces conditions, même de bonnes statistiques des captures passées et présentes ne sont pas des indicateurs valables pour estimer la production potentielle future. L'expérience a montré que nos connaissances sur les fluctuations naturelles de l'abondance des stocks halieutiques, leurs causes et leur évolution, ainsi que sur les moyens d'y faire face, étaient très incomplètes.
On s'est continuellement efforcé de maîtriser ou d'atténuer les fluctuations interannuelles des captures de poissons et de l'abondance des stocks. Pendant plusieurs décennies, les scientifiques et les administrateurs des pêches avaient principalement porté leurs efforts sur le contrôle de la pression de pêche et la résolution du problème de la surpêche, en partant du principe que les facteurs environnementaux et les causes naturelles des variations de l'abondance des stocks pouvaient être considérés comme des paramètres aléatoires ou constants. Toutefois, la proportion relativement élevée de stocks de poissons qui, au fil des années, ont été surexploités et épuisés laisse penser que l'approche consistant à prévenir la surpêche et à limiter les fluctuations des ressources, sur la base d'un contrôle de la pression de pêche, comme l'ont préconisé Russell (1931), Beverton et Holt (1957), Ricker (1958) Gulland (1969) et d'autres, n'a pas toujours donné les résultats attendus.
Lorsque l'on ne parvient pas à maintenir les stocks halieutiques exploités en bonne santé et à prévenir ou à éviter leur surexploitation et leur épuisement, cela est dû au moins à deux causes: les caprices de la Nature et des erreurs au niveau de la gestion des pêches et de la prise de décision. Bien souvent, des variations naturelles aussi importantes qu'imprévisibles de l'abondance des stocks ont été responsables de la baisse des populations de poissons et de l'effondrement de pêcheries importantes. Cependant la cupidité et la mauvaise gestion, notamment le défaut d'application ou de mise en exécution des directives et des réglementations convenues, ont parfois été responsables, entièrement ou en partie, du déclin catastrophique d'importantes ressources halieutiques et de la faillite de leurs pêcheries. Malgré leur importance, nous accorderons ici peu de place à ces derniers facteurs, et aux causes naturelles à court terme des fluctuations des stocks, afin de centrer notre attention sur quelques aspects pertinents des changements climatiques naturels prolongés qui peuvent avoir une incidence sur les pêches, en provoquant des fluctuations parfois importantes des stocks de poissons, au fil du temps.
FLUCTUATIONS À LONG TERME DES PÊCHERIES
La FAO a organisé en 1983 une importante réunion sur les changements ayant une incidence sur les stocks halieutiques et les pêcheries, en vue d'examiner les différentes causes de changement, les types de fluctuations, leurs causes et leur relation possible avec des facteurs environnementaux, ainsi que les stratégies de recherche et de gestion proposées pour y faire face, et de prendre les mesures qui s'imposent. (Sharp et Csirke, 1983; Csirke et Sharp, 1984). Après avoir d'abord attiré surtout l'attention de la communauté scientifique, avec le développement des pêches et l'accès à des données plus nombreuses et portant sur des périodes plus longues, les changements à long terme qui influent sur les pêches se sont mis à attirer l'attention des pêcheurs, des responsables de l'aménagement des pêches, des responsables politiques, des décideurs et du grand public.
Le premier rapport scientifique sur les fluctuations à long terme de l'abondance des harengs a été publié en 1879, sur la base d'observations faites depuis le XVIe siècle. Ljungman (1879, cité dans Lindquist, 1983) décrivait les fameux «cycles du hareng» qui duraient entre 30 et 60 ans, dans l'archipel suédois de Bohuslän. Depuis, de nombreux autres rapports sur les fluctuations à long terme des pêches maritimes ont vu le jour, en particulier celui de Kawasaki qui tentait de trouver une explication aux amples fluctuations enregistrées par certaines populations de poissons pélagiques (Kawasaki, 1983). En mettant en relief l'évolution synchronisée des captures des trois principales espèces de pilchards du Pacifique, du genre Sardinops depuis le début du XXe siècle, son rapport a convaincu la communauté scientifique que les explosions, les pics et les retours à de très bas niveaux que connaissaient ces ressources (Csirke et Sharp, 1984) étaient induits par des causes naturelles. Les théories de Kawasaki ont trouvé confirmation au fur et à mesure que les pêches mondiales se sont développées et que sont apparus de nouveaux indices des fluctuations prolongées de l'abondance des poissons. Cela est illustré par exemple, par la mise à jour des principales captures de pilchards (genre Sardinops) à la figure C4.1, où l'on note cependant que depuis le début des années 90, le pilchard de Californie ne suit pas la tendance observée chez d'autres espèces du même genre dans le Pacifique. Les captures de pilchard de Californie ont culminé en 2002, dernière année pour laquelle on dispose de statistiques officielles, alors que celles de pilchards sud-américain et de pilchards du Japon ont accusé une brusque baisse après les pics respectivement atteints en 1985 et en 1988.
Figure C4.1 - Variabilité observée à long terme des captures de pilchard du Japon (Sardinops melanostictus) dans le Pacifique Nord-Ouest, de pilchard de Californie (S. caeruleus) dans le Pacifique Centre-Est et de pilchard sud-américain (S. sagax) dans le Pacifique Sud-Est (mise à jour d'après Kawasaki, 1983; Csirke, 1995 et FAO, 1997)
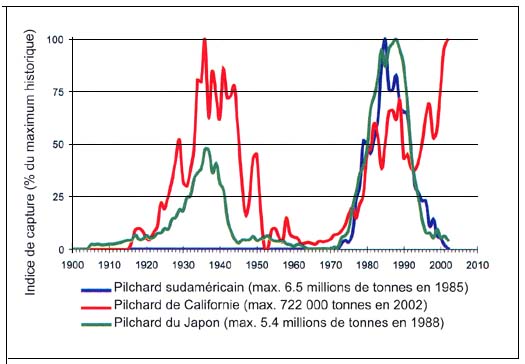
Deux séries de facteurs au moins semblent pouvoir raisonnablement expliquer la tendance haussière continue des captures de pilchards de Californie. Tout d'abord une succession de bonnes classes annuelles, en particulier en 1998, et une biomasse du stock élevée dans les eaux californiennes qui s'est stabilisée à environ 1 million de tonnes depuis 1999 (Conser et al., 2002), peut-être favorisées par le phénomène «El Niño» de 1997–98. En second lieu, les activités intenses de la flottille de pêche mexicaine à l'intérieur du Golfe de Californie (dont les prises ont représenté ces dernières années environ 60 pour cent des captures totales), alors que dans les eaux d'Ensenada, les prises sont très inférieures et au même niveau qu'au large de la côte Pacifique des États-Unis, où la pêche est sévèrement réglementée par un système de contingents visant à préserver le stock et à prévenir sa surexploitation. Ces deux séries de facteurs peuvent contribuer à retarder la phase de déclin de ce récent cycle d'abondance élevée des pilchards de Californie.
Comme on le verra plus loin, des fluctuations à long terme de l'abondance et/ou des captures de poissons ont été signalées pour plusieurs autres stocks importants à l'échelle mondiale, dont les morues, les saumons et les thons.
Mécanismes déclencheurs possibles
On cherche de plus en plus à établir des corrélations entre les cycles halieutiques et les indicateurs disponibles concernant la variabilité climatique à long terme, pour identifier les mécanismes auxquels pourraient être dues ces fluctuations Au cours des deux dernières décennies, les chercheurs se sont surtout efforcés de décrire et d'analyser les fluctuations à long terme de l'abondance des espèces commerciales et les rapports pouvant exister entre le climat océanique et la taille des stocks de poissons. L'un des cas les mieux documentés dans la littérature est celui, mentionné plus haut, des poussées synchrones périodiques des populations de pilchards et d'anchois dans les hémisphères Nord et Sud qui laissent penser que les populations de poissons sont gouvernées par des phénomènes climatiques similaires à l'échelle mondiale. Poursuivant plus avant ses travaux, Kawasaki (1994) a été l'un des premiers à étudier plus à fond le concept des changements cycliques du climat pour expliquer les cycles réguliers des captures de pilchards du Japon, au cours des 350 dernières années.
L'analyse des séries chronologiques reconstituées des captures de morues dans l'Atlantique Nord a aussi mis en évidence des oscillations synchrones du stock occidental et du stock oriental, probablement causées par les mêmes mécanismes climatiques (Garrod et Schumacher 1994; Jonsson 1994). D'autres auteurs ont analysé les liens possibles entre le réchauffement de la planète et les fluctuations des stocks de poissons (Francis, 1990; Glantz, 1990; Regier et Meisner, 1990). Selon Kawasaki (1992) et Lluch-Belda et al., (1992), les captures de pilchard de Californie et du Japon seraient influencées par les variations de la température de l'air et de l'eau en surface dans l'hémisphère Nord. Beamish et Bouillon (1993) ont montré que les fluctuations des captures de saumon du Pacifique suivaient la dynamique de l'Indice régional de variation de la cellule de basse pression aléoutienne, qui est reconnu comme le facteur climatique prépondérant dans le Pacifique Nord. Bakun (1990; 1996) a relié les fluctuations à long terme des petits poissons pélagiques à des variations de l'intensité des upwellings , affirmant que les changements climatiques mondiaux sont accompagnés de variations significatives dans la circulation atmosphérique et dans l'intensité des upwellings . Comme les populations les plus abondantes de petits pélagiques se trouvent à proximité des épicentres d'upwellings intenses, toute variation de l'intensité des remontées d'eau froide a une influence considérable sur la production primaire océanique et sur les captures de poissons dans ces zones. De même, après avoir analysé des séries chronologiques à long terme s'étalant sur plus de 300ans, des captures de thons rouges pêchés à la madrague selon des méthodes traditionnelles, dans l'Atlantique et en Méditerranée, Ravier and Fromentin (2001) ont montré que le stock oriental de thons rouges de l'Atlantique présentait deux types de variations cycliques, l'une avec une période de 100–200 ans et l'autre de l'ordre de 20 ans. Il existait une forte corrélation négative entre les fluctuations à long terme et l'évolution de la température. La relation étroite entre les captures et la température semble surtout due à des modifications des habitudes de migration des thons rouges résultant de modifications des conditions océanographiques des zones de ponte, qui ont fait que les ressources présentes là où se trouvent les engins de pêche fixes n'étaient plus les mêmes (Ravier et Fromentin, 2004).
Dans le cadre du rôle qui lui revient, en tant que principale institution spécialisée des Nations Unies en matière de pêche, la FAO a appuyé plusieurs études visant à mieux comprendre les relations possibles entre les changements climatiques et les variations de la production halieutique, leurs causes et leurs mécanismes déclencheurs, ainsi que les utilisations et les applications potentielles de ces nouvelles connaissances dans le domaine de la planification de la conservation et du développement des pêcheries de la planète. La Consultation d'experts de la FAO qui a analysé ces changements (Sharp et Csirke 1983; Csirke et Sharp 1984) est une initiative importante dans cette direction. D'autres tentatives ont suivi, notamment les études de Klyashtorin (2001) et Sharp (2003) qui sont parmi les plus récentes.
Figure C4.2 - Fluctuations cycliques des températures et périodes d'abondance du pilchard du Japon (S.melanosticus), entre 1600 et 2000 (extrait de Klyashtorin, 2001)
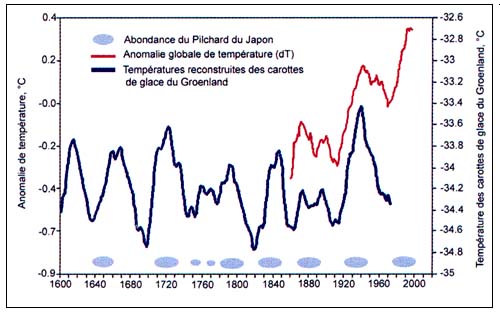
L'existence d'une corrélation entre les indices climatiques et l'abondance de diverses espèces qui connaissent des fluctuations sur de longues périodes a été démontrée. Des observations à long terme portant sur les augmentations soudaines de populations de pilchard du Japon et les indices de température atmosphérique (Figure C4.2) ont donné lieu à des hypothèses selon lesquelles les variations régulières des captures de pilchard du Japon observées sur de longues périodes pourraient s'expliquer par des changements climatiques cycliques (Kawasaki, 1994). Plus récemment, des ensembles de données sur les débarquements mondiaux disponibles auprès de la FAO et d'autres organismes ont été analysés par Klyasthorin (2001) pour tenter d'examiner les rapports pouvant exister entre divers indices climatiques et les captures de certains groupes de poisson. Cet auteur a également élaboré un modèle chronologique basé sur des cycles climatiques connus pour déterminer d'éventuelles tendances concernant les principaux types de capture commerciale pour les5à 15 prochaines années. De telles prévisions exigent une grande prudence, mais les résultats donnent suffisamment à réfléchir et suscitent assez d'intérêt pour mériter une attention plus grande et être analysés plus en profondeur. Les récentes théories et conclusions de Klyashtorin, reposent sur l'analyse de la relation possible entre les indices d'abondance du poisson, reflétés par les captures annuelles, et les changements climatiques. Dans ce contexte, l'expression «changements climatiques» désigne les effets de grande ampleur et de longue durée - ou les passages d'un état climatique à un autre - qui semblent obéir à des cycles déterminés, plutôt que les phénomènes climatiques individuels comme El Niño ou des tendances à long terme comme le réchauffement de la planète.
On ne sait toujours pas exactement quels mécanismes sont à l'origine de la plupart des fluctuations périodiques de longue durée de l'abondance de poisson analysées par Klyashtorin, et certaines de ses conclusions restent des hypothèses de travail. Cependant, les indications et les tendances concernant les indices climatiques et l'évolution des débarquements de poisson qui ressortent de ses travaux sont du plus haut intérêt et méritent d'être étudiés de près afin de mieux comprendre les mécanismes gouvernant le changement climatique et la variabilité de la production de poisson sur de longues périodes et de les utiliser aux fins de l'aménagement des pêches.
Figure C4.3 - Périodicité de l'indice de circulation atmosphérique (ACI), des valeurs - tendances ôtées- de l'anomalie des températures globales (dT) et de l'opposé de l'index de longueur du jour (-LOD) avec décalage temporel non modifié (A, haut) et avec ACI et - LOD décalé de 4 et de 6 ans en avant (B, bas) (modifié d'après Klyashtorin, 2001)
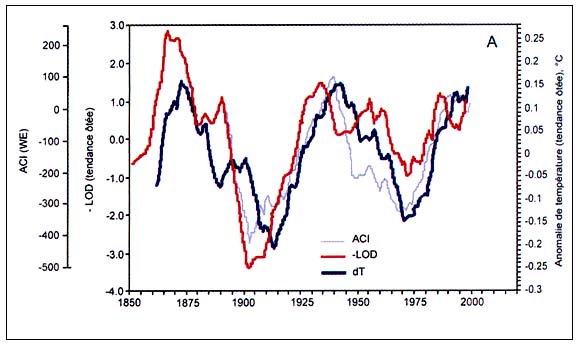
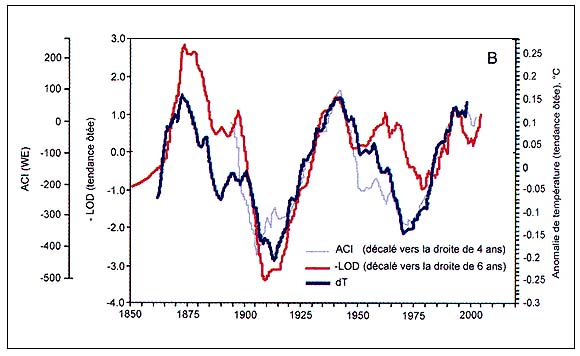
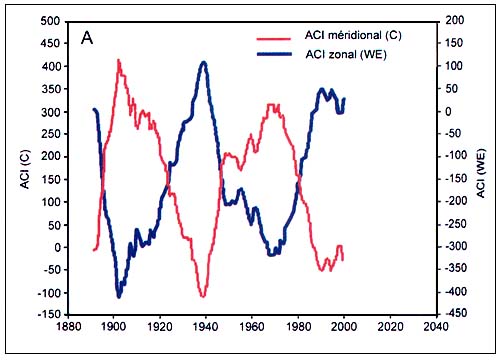
Figure C4.4 - Dynamique des formes méridionale (C) et zonale (WE) de l'indice de circulation atmosphérique (ACI) (A, haut) et alternance des époques de circulation méridionale et zonale correspondantes (B, bas) (extrait de Klyashtorin, 2001)
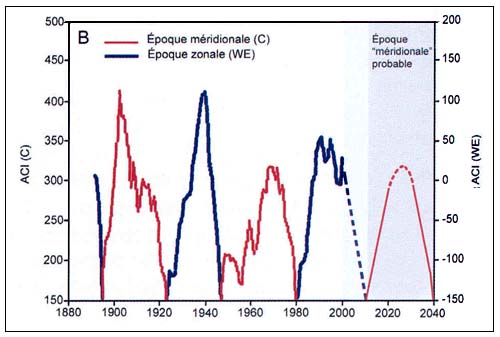
L'argument avancé dans la plupart des cas examinés par Klyasthorin est que la biomasse et les captures dépendent en fin de compte des fluctuations climatiques, ce qui va à l'encontre du principe communément accepté en matière d'aménagement des pêches, selon lequel la biomasse et les captures sont principalement influencées par l'effort de pêche, et les effets de l'environnement sur les stocks de poissons sont le plus souvent aléatoires ou stochastiques. En fait, les études sur le climat et les pêches ont démontré que les effets de l'environnement sur les stocks de poissons sont caractérisés par une longue séquence de phénomènes et de processus qui tendent à devenir cycliques ou à donner naissance à de nouveaux régimes (Sharp, 2003). Avec des observations suffisantes sur les liens de cause à effet, il devrait être possible de prévoir le passage d'un régime à l'autre.
On a déjà émis l'hypothèse (Bakun 1996) que l'intensité des upwellings était liée aux effets climatiques de forte ampleur qui influent finalement sur le taux de transport des nutriments dans la couche eutrophique supérieure de l'océan, modifiant ainsi la production primaire et, ensuite, la production de poisson. Or, les hypothèses de l'existence d'un lien entre le climat et la disponibilité de nutriments sont peut-être exactes, mais il n'existe aucune preuve directe d'un tel mécanisme, et il n'a pas encore été possible d'établir un modèle probant de ce rapport de cause à effet.
Les travaux de Klyashtorin ont mis en évidence une corrélation constante entre les indices climatiques et géophysiques et la manifestation de processus importants liés aux pêches. Ces indices sont l'anomalie de la température de l'air à la surface (dT), principal indice du changement climatique mondial; la longueur du jour (LOD), un indice géophysique qui caractérise les variations de la vitesse de rotation de la terre; et l'Indice de circulation atmosphérique (ACI), qui caractérise les périodes de relative dominance du transport directionnel des masses d'air à l'échelle d'un hémisphère. L'indice LOD représente l'écart entre la longueur astronomique et la longueur standard du jour. Les échelles des temps servant à calculer l'indice LOD sont disponibles dans les archives du Bureau international des poids et mesures (http://www.bipm.org), et les séries chronologiques de données s'étalent sur plus de 350 ans.
L'indice ACI se calcule sur la base d'un système de classification mis au point par les climatologistes russes. Dans ce système, il existe trois grandes catégories de variations observables de la circulation atmosphérique, qui sont fonction de la direction du transfert des masses d'air, à savoir les variations méridionales, zonales occidentales, et zonales orientales). Les transferts des masses d'air dans chaque direction sont calculés sur la base des graphiques de la pression atmosphérique quotidienne pour la région de l'Atlantique Nord et l'Eurasie. L'indice ACI est défini par le nombre de jours avec le type de circulation atmosphérique dominant, exprimé en tant qu'anomalie par rapport à la moyenne sur une longue période.
L'analyse spectrale des séries chronologiques de dT, ACI et LOD estimées sur la base des observations directes disponibles (110 à 150 ans) révèle une périodicité commune de 55 à 65 ans (Figure C4.3). L'analyse spectrale des séries chronologiques reconstituées des températures de l'air en surface au cours des 1 500 dernières années semble révéler une périodicité analogue (55 à 60 ans). De plus, les observations de l'indice ACI révèlent une alternance entre deux phases climatiques durant environ 30 ans chacune selon les modes dominants de transport des masses d'air à l'échelle de l'hémisphère (Figure C4.4).
Chaque période de 30 ans correspond à une «époque de circulation» des transferts des masses d'air de type méridional (C) et zonal (occidental + oriental). Les époques zonales correspondent à des périodes de réchauffement de la planète et les époques méridionales à des périodes de refroidissement de la planète (Sharp, 2003). Les variations de longue durée des indices à l'échelle planétaire (LOD) ont aussi été associées à des modifications de processus régionaux dans le Pacifique, notamment de l'intensité des upwellings locaux et de la pression au niveau de la mer à l'échelle du bassin (Sharp, 2003). Ainsi, il existe une force directe résultant de l'action due au vent synchrone avec la LOD qui détermine les régimes de température (froid/chaud) dans l'Océan Pacifique. Ces régimes sont connus pour influencer la faune marine du Pacifique Nord, notamment les populations nombreuses de petits poissons pélagiques. Des études sur les écosystèmes de la bordure orientale montrent par exemple que les anchois prospèrent durant les périodes d'upwellings côtiers plus froids. Les pilchards préfèrent les eaux plus chaudes et semblent résister aux périodes de fortes remontées d'eau froide en se maintenant, par petites colonies, à distance des influences des upwellings (Sharp, 2003). Klyashtorin a estimé que les indices de circulation atmosphérique et de la longueur du jour étaient des indices climatiques appropriés pour étudier de façon plus approfondie les variations régulières de longue durée des débarquements des principaux stocks de poisson commerciaux.
CORRÉLATION ENTRE LES PÊches et le climat
Il existe deux sources principales de données révélant un lien entre le climat et les débarquements de poisson: quelques indices concernant le climat et la taille des stocks de poisson couvrant une période prolongée allant jusqu'à 1700 ans, qui révèlent des modalités cycliques similaires ainsi qu'une corrélation entre les séries; et les fluctuations des captures des 12 principales espèces commerciales de l'Atlantique et du Pacifique, qui se sont synchronisées depuis 1900 et correspondent aux indices climatiques de la même période. Les séries de longue durée et celles de brève durée semblent avoir un cycle commun. La périodicité la plus prononcée des fluctuations à long terme des captures pour toutes les séries chronologiques (sauf pour l'anchois) varie entre environ 54 et 58 ans. Les cycles climatiques correspondants (mesurés ou reconstruits) varient entre 50 et 65 ans (avec une moyenne de 56 ans). D'autres cycles moins importants (les fluctuations sur 13 et 20 ans des températures estivales) peuvent aussi présenter un intérêt, mais on n'a encore constaté aucune corrélation fiable entre ces cycles et les fluctuations des captures commerciales.
En fonction des particularités de leur dynamique à long terme, Klyashtorin a divisé les 12 espèces examinées en deux groupes. Le premier comprenait le pilchard du Japon, le pilchard de Californie et le pilchard sud-américain, le saumon du pacifique, le lieu d'Alaska, le chinchard du Chili et la sardine européenne. Les séries chronologiques indiquent que les captures de ces espèces ont été à leur plus haut niveau à la fin des années 30 et au début des années 90 et à leur plus bas niveau dans les années 60. Le deuxième groupe, qui comprend la morue de l'Atlantique, les harengs de l'Atlantique et du Pacifique, le pilchard d'Afrique du Sud et l'anchois du Pérou, affiche un cycle de production qui a évolué en sens contraire, avec des captures maximales dans les années 60 et minimales dans les années30 et 90. Alors que les captures du premier groupe (Figure C4.5) sont très corrélées avec la courbe de l'indice de circulation atmosphérique zonal, celles du deuxième groupe (Figure C4.6) sont en rapport étroit avec la courbe de l'indice de circulation atmosphérique méridional.
En ce qui concerne les cycles à long terme, les chroniques japonaises contiennent des données historiques sur l'abondance du pilchard du Japon au cours des 400 dernières années (Figure C4.2). Des variations de la disponibilité et de l'abondance des pilchards au large du Japon ont favorisé l'expansion de plusieurs villages côtiers de pêcheurs aussi bien que la disparition d'autres villages. La durée moyenne d'un cycle est d'environ 60 ans, et les périodes de forte abondance coïncident généralement avec des périodes marquées par une température atmosphérique plus élevée.
Figure C4.5 - Corrélation entre les captures annuelles de pilchard du Japon (A, haut) et de saumon du Pacifique (B, bas) et l'indice de circulation atmosphérique (ACI) zonal (extrait de Klyashtorin, 2001)
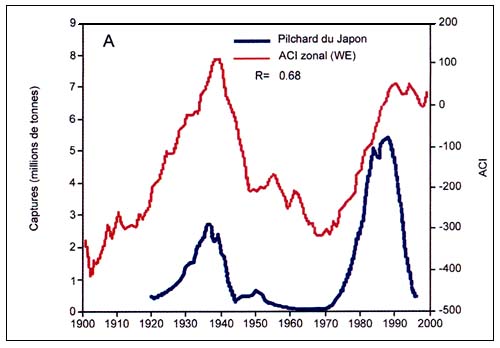
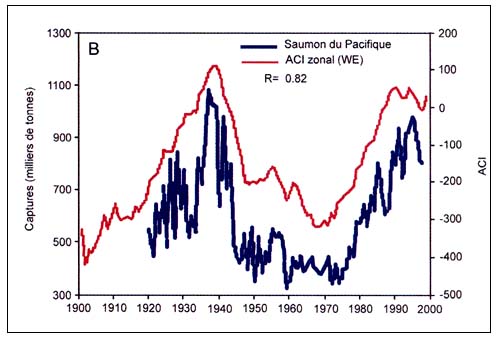
Figure C4.6 - Corrélation entre les captures annuelles de morue de l'Atlantique (A, haut) et de hareng du Pacifique (B, bas) et l'indice de circulation atmosphérique (ACI) méridional (extrait de Klyashtorin 2001)
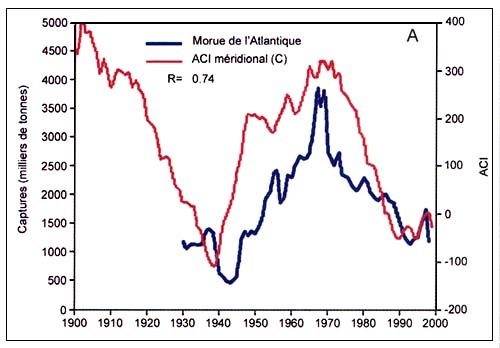
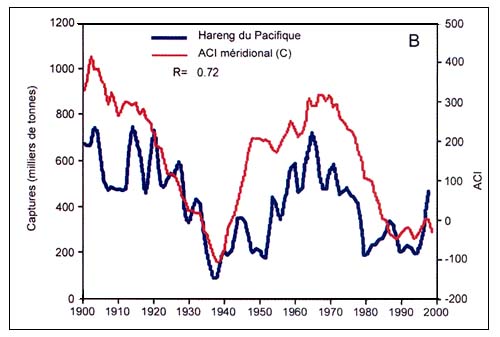
Au large de la côte de la Californie, les conditions anaérobies régnant dans les sédiments déposés au fil des saisons ont préservé les écailles de populations de petits poissons pélagiques, ce qui a permis de reconstruire deux séries chronologiques de l'indice d'abondance pour les stocks de pilchards et d'anchois au cours des 1700 dernières années (Baumgartner, Soutar et Ferreira-Bartrina, 1992). Même si elles révèlent de fortes fluctuations, il est intéressant de noter que ces séries chronologiques ne font ressortir aucune différence nette entre la période plus ancienne, où la pêche était un facteur négligeable, et la période plus récente au cours de laquelle l'exploitation a pris beaucoup d'ampleur.
L'analyse des séries chronologiques longues des écailles des sédiments a révélé deux oscillations principales dans l'abondance de pilchards, l'une se produisant tous les 54 à 57 ans, et l'autre tous les 223 à 273 ans. La première est semblable à celles observées pour la température atmosphérique mesurée à partir des carottes de glace fossile et pour la biomasse de pilchards, ce qui la rend particulièrement importante pour les prévisions en matière de pêche. Pour l'anchois, les périodes de fluctuation dominantes ont une durée d'environ 100, 70 et 55 ans. Les augmentations de l'abondance de pilchards et d'anchois semblent être liées respectivement aux deux régimes atmosphériques (phases zonale et méridionale) déjà mentionnés, ce qui donne à penser que ces deux espèces peuvent être favorisées par des conditions climatiques différentes.
Toutefois, la dynamique normale de la dépendance climatique de l'anchois du Pérou est différente de celle d'autres espèces pélagiques commerciales; elle est fortement perturbée tous les 10 à 15 ans par les manifestations violentes du phénomène El Niño, si bien qu'une courbe lisse des valeurs moyennes ne peut donner une bonne idée approximative de la future dynamique des captures de cette espèce. Dans les années normales (sans El Niño), l'anchois du Pérou représente environ 70 à 90 pour cent des captures totales d'anchois dans le Pacifique.
On pourrait logiquement s'attendre à ce que les captures de poisson présentent une corrélation plus grande avec les indices climatiques régionaux correspondants qu'avec ceux qui ont une portée plus mondiale. Toutefois, d'après les résultats obtenus par Klyasthorin (2001) la dynamique des captures des principales espèces commerciales du Pacifique (saumon du Pacifique, pilchard du Japon, pilchard de Californie et pilchard sud-américain, lieu de l'Alaska et chinchard du Chili) semble plus étroitement reliée aux indices climatiques mondiaux d'anomalie thermique et de circulation atmosphérique qu'aux indices régionaux correspondants, mais cela n'a pas encore été expliqué de façon satisfaisante.
La faible corrélation avec les indices régionaux pourrait s'expliquer par le fait que, dans les analyses de la relation entre le climat et les pêches, on tend à amalgamer des données provenant de vastes régions et de stocks géographiquement distincts. Cela semble être le cas pour les populations de saumon du Pacifique. Analysant les taux de recrutement de multiples stocks de saumon provenant de différents bassins hydrographiques de la Colombie-Britannique et de l'Alaska, Peterman et al. (1998) ont conclu que les facteurs environnementaux qui influencent les taux de survie étaient principalement ressentis à des échelles spatiales régionales, démontrant notamment qu'une variation décennale de la productivité dans les années 70, selon la thèse de Klyashtorin, ne se produisait que dans les stocks de saumon provenant de bassins hydrographiques déterminés. Cela indique que d'importants processus environnementaux ayant une incidence sur la survie des saumons agissent à l'échelle régionale et non à celle du bassin océanique. Dans le cas des stocks de saumon du Pacifique, les variations de la productivité résultent d'une combinaison de processus se produisant dans les eaux douces et marines, qui ne sont pas encore parfaitement compris
POSSIBILITÉS DE PRÉVISION
Ce qui frappe le plus lorsque l'on examine les corrélations entre les indices dt, LOD et ACI, c'est que, bien que les courbes aient une forme très similaire, les variations de dt sont retardées d'environ 6 ans par rapport a LOD, et de 4 ans par rapport à l'ACI (Figure C4.3). Cela permet de prévoir la dynamique de dt au moins six ans à l'avance. Le décalage dans le temps observé entre les indices climatiques, leur récurrence dans le passé, et leur oscillation synchrone avec la production halieutique ont incité Klyashtorin à prévoir les variations du climat et des populations marines.
Partant du principe que la corrélation observée dans le passé entre les captures de poisson et les régimes atmosphériques se maintiendrait à l'avenir, Klyashtorin tente de prévoir les captures totales d'un groupe déterminé d'espèces commerciales importantes en fixant les durées des cycles à 55, 60 et 65 ans (sur la base du cycle climatique) plutôt qu'en se fondant sur les estimations de la durée des cycles établies à partir des séries chronologiques relativement courtes des captures. D'après ces analyses, il y aurait bien un passage de l'un à l'autre des deux régimes atmosphériques potentiels. De ce fait, si la synchronisation observée entre les phases d'abondance du poisson et les cycles climatiques persiste, et si l'effort de pêche ne varie pas dans une mesure significative, on pourrait s'attendre à ce que la production de ces importantes espèces commerciales diminue dans le Pacifique Nord et augmente dans l'Atlantique Nord au cours de la prochaine décennie (Figure C4.7). Les prévisions concernant les captures des principaux poissons commerciaux durant les 30 à 40 prochaines années ne sont, de façon générale, pas influencées par le choix de la périodicité dans une gamme située entre 55 et 65 ans. Des espèces comme les harengs de l'Atlantique et du Pacifique, la morue de l'Atlantique, le pilchard d'Afrique du Sud et les anchois du Pérou et du Japon devraient augmenter entre 2000 et 2015 et diminuer ensuite. Pendant la même période, les captures totales d'espèces telles que le pilchard du Japon, le pilchard de Californie, le pilchard sud-américain, la sardine européenne, le saumon du Pacifique, le lieu de l'Alaska et le chinchard du Chili devraient diminuer, puis remonter. Dans l'ensemble, les captures totales des principales espèces commerciales mondiales prises en considération dans ces analyses, qui représentent environ un tiers des débarquements mondiaux d'espèces marines, devraient augmenter de 5,6 millions de tonnes d'ici 2015, puis diminuer de 2,8 millions de tonnes d'ici 2030, uniquement à cause du climat.
MISES EN GARDE ET LIMITES
Klyashtorin a conclu que les variations cycliques du climat pouvaient servir de base pour prédire la production et les captures des principales espèces commerciales de poissons. Il avertit cependant que son modèle n'est applicable qu'aux espèces abondantes (captures commerciales >1–1,5 million de tonnes) distribuées sur de vastes zones, comme l'ensemble du Pacifique Nord ou de l'Atlantique Nord. Une autre limite mise en évidence par l'auteur est que les prévisions ne sont valables qu'à condition que l'effort de pêche commerciale ne varie pas de façon significative par rapport à la moyenne des 20à 25 dernières années.
Comme l'a suggéré Sharp (2003) lorsque l'on étudie des systèmes complexes, tels que les interactions entre le climat et les pêches, il est indispensable de disposer de systèmes de suivi acceptables, gérés sur des périodes suffisamment longues pour pouvoir capter la dynamique intégrale du système. Or, dans le domaine des pêches, il est très rare que des séries chronologiques couvrent plus d'un ou deux cycles d'évolution complets (expansion et contraction) des populations concernées. Les cas accompagnés d'observations du (des) système(s) de forçage qui influence(nt) directement ou indirectement ces populations sont encore moins nombreux. La pénurie de données est, selon Sharp (2003), l'un des principaux obstacles qui empêchent de prévoir l'effet du climat sur les pêches. En outre, la difficulté est aggravée par le fait que la majorité des données sur les pêches utilisées dans des analyses climatiques portent sur les 50 dernières années, une période caractérisée, selon l'auteur, par son absence de dynamique et sa relative stagnation par rapport aux données enregistrées sur un siècle ou sur des périodes plus longues.
Figure C4.7 - Variations du volume des captures observées (1920–1998) et prévues (2000–2040) pour certaines des principales espèces commerciales de l'Atlantique Nord (modifié d'après Klyashtorin, 2001)
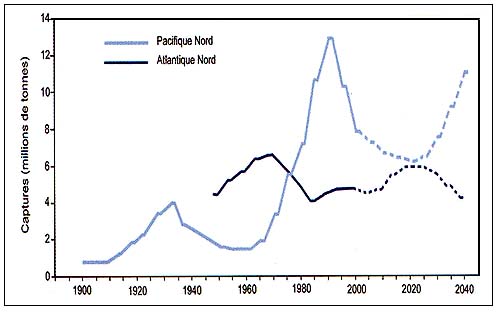
Il ne faut pas non plus oublier que les séries chronologiques sur 50 ans disponibles pour la plupart des stocks examinés par Klyasthorin(2001) concernent une période marquée par les changements les plus drastiques de l'effort de pêche et des technologies de localisation et de capture des poissons. Ces changements ont permis à la pêche de se développer vers des zones jusque là inexploitées et de devenir beaucoup plus efficace. L'analyse des tendances des captures halieutiques durant cette période doit tenir compte de ces effets. Enfin, en tant qu'éléments des écosystèmes, les stocks de poissons sont liés à d'autres espèces, par des interactions prédateur-proie ou par des relations d'une autre nature, de sorte que l'on doit s'attendre à ce que les importants prélèvements effectués par les pêches dans la biomasse des poissons aient des répercussions positives et négatives sur la production d'autres ressources, éventuellement aussi ciblées par les pêcheries commerciales. La littérature regorge d'exemples de modifications de la dominance des espèces, découlant probablement de la compétition ou de la prédation. Par exemple, Fogarty et Murawski (1998) ont démontré qu'une réduction due à la pêche de plus de 50 pour cent de la biomasse de gadidés et de plies dans le Banc Georges avait conduit à une augmentation de l'abondance de squales et de raies. On a également tenté d'établir un lien entre les alternances observées entre les espèces de pilchard et d'anchois, la surexploitation et les mécanismes trophiques. Les hypothèses les plus fréquemment émises dans la littérature sont la compétition alimentaire et la prédation de pilchards ou d'anchois adultes sur des espèces concurrentes, aux premiers stades de leur vie. (Vasconcellos, 2000).
En résumé, toutes les tendances des captures halieutiques observées au cours des 50 dernières années devraient être considérées comme résultant de l'interaction de processus humains (pêche), écologiques et climatiques sur les populations de poissons, dont on ignore cependant encore l'importance relative. Ces processus sont confondus dans les séries chronologiques sur les captures, de sorte que les prévisions et corrélations établies sur la base de ces données sont sujettes à caution.
IMPLICATIONS EN MATIÈRE DE POLITIQUES
Les changements climatiques peuvent avoir une influence sur la pêche, aussi bien à court terme qu'à long terme. Alors que les pêcheries ont mis au point des mécanismes pour faire face aux variations environnementales à court terme (calendriers d'activité saisonniers), c'est la composante à long terme de la variabilité (sur plusieurs années ou décennies) qui est la moins bien comprise, la moins prévisible, et celle qui a le plus d'influence sur la viabilité d'une pêcherie.
La possibilité de prévoir les changements à long terme du volume mondial des captures de poisson en se basant sur les indices observables de la variabilité climatique à long terme suscite des attentes et des préoccupations justifiées aux plans scientifique, économique et politique. Les organismes d'aménagement considèrent généralement que c'est l'effort de pêche qui a la principale influence sur la productivité et la taille à long terme des stocks de poissons. Une idée communément acceptée est qu'un aménagement approprié pourrait maintenir la taille des stocks à des niveaux permettant de réaliser durablement des captures élevées et que les erreurs d'aménagement ont habituellement pour conséquence la diminution - voire l'épuisement - des stocks et la baisse des rendements. La prise de conscience du fait que, pour quelques espèces clés, les effets déterministes à long terme du climat sur l'abondance des stocks sont, ou pourraient être, aussi importants qu'on l'a dit incite à examiner les stratégies et les objectifs de la recherche et de l'aménagement pour les pêches des espèces concernées ainsi que pour les espèces apparentées. Les mesures d'aménagement prises pour réagir face aux fluctuations à court terme devraient tenir compte de l'existence possible de fluctuations déterministes de longue durée influant sur les premières. La prise en compte des effets climatiques à long terme favoriserait aussi l'atteinte des objectifs à long terme de l'aménagement, qui devraient en général inclure des immobilisations ainsi que des mesures sociales et le développement des infrastructures.
Dans l'ensemble, les augmentations et les diminutions de la production de poisson dues au climat ne semblent pas avoir une grande importance au plan mondial, étant donné que les augmentations d'un groupe de stocks dans une région sont généralement compensées par des diminutions touchant un autre groupe dans une autre région. Toutefois, il convient d'examiner sérieusement le fait que les changements climatiques à long terme pourraient entraîner d'importantes augmentations périodiques de la production de poissons appartenant à certains stocks dans certaines zones et des réductions équivalentes d'autres stocks dans d'autres zones, parce que cela aura des répercussions nécessairement beaucoup plus marquées aux niveaux local et régional. Par exemple, à l'heure actuelle, les grands flux commerciaux internationaux vont des pays en développement vers les pays développés (c'est-à-dire du Sud vers le Nord), mais les oscillations de la production entre l'Atlantique Nord et l'Océan Pacifique risquent de modifier ces flux, ce qui aurait de fortes répercussions sur les marchés nationaux et régionaux, même si les approvisionnements totaux mondiaux restaient stables. On s'attend également à ce que les investissements et les mouvements des flottilles de pêche (ainsi que les accords de pêche) changent davantage que ne le donnerait à penser le montant cumulatif des débarquements mondiaux.
La présente analyse ne tient pas compte des changements climatiques anthropogéniques et de leurs effets possibles sur la production halieutique. Toutefois, comme les données disponibles semblent montrer l'existence d'un lien entre la production de poisson et le climat, il est clair que les effets du réchauffement planétaire doivent être inclus dans les éventuelles projections. Au vu des résultats mentionnés, toute modification du climat pourrait avoir des effets positifs ou négatifs notables sur certains des principaux stocks de poissons commerciaux, sinon sur la plupart d'entre eux.
L'incertitude est un élément important de l'interaction entre les pêches et le climat. Dans ce contexte, il est indispensable de se demander comment les stratégies d'aménagement devraient être formulées pour faire face aux incertitudes sur les changements climatiques futurs ayant une influence sur les stocks de poissons. Il semble être universellement reconnu que la viabilité dans le temps des systèmes de gestion des ressources dans un environnement en évolution, repose essentiellement sur l'adaptation (GIEC, 2001). L'adaptation de la gestion des pêches aux changements climatiques peut reposer sur des arrangements officiels et officieux. Les politiques officielles sont les mesures prises par les gouvernements, notamment les lois, les règlements et les incitations, pour provoquer ou faciliter des réformes des systèmes socio-économiques, en vue de réduire la vulnérabilité aux variations climatiques. Les communautés de pêcheurs ont aussi trouvé des moyens informels d'accroître leurs chances de succès et de survie dans un environnement qui change. Les stratégies d'adaptation des communautés comprennent l'adoption de modes de pêche saisonniers flexibles, la mise en place de réseaux pour le partage des ressources alimentaires et autres, et le commerce entre communautés (Berkes and Jolly, 2001). L'objectif des politiques et des stratégies d'aménagement adaptatives est de réduire la vulnérabilité, et une étude de la vulnérabilité des pêches aux variations présentes et futures du climat peut assurément contribuer à améliorer les politiques d'adaptation.
La gestion des pêches dans des conditions climatiques incertaines peut aussi être facilitée par des procédures d'évaluation et de gestion des risques et par l'adoption d'une approche de précaution (Cochrane, 2002). Dans ce cas précis, le risque qu'il convient d'évaluer et de gérer est la probabilité que les stocks tombent en dessous de certains seuils convenus, par suite du climat et des activités de pêche. Une stratégie propre à renforcer la protection de la ressource dans les phases de faible abondance et/ou lorsqu'elle tombe en dessous d'un certain seuil, et à permettre un accroissement de l'effort de pêche dans les phases d'abondance élevée, a été proposée pour les ressources et les pêcheries sujettes à de grandes variations (MacCall, 1980; Csirke, 1984, 1987). Une telle stratégie devrait garantir une protection suffisante contre l'épuisement des ressources dû à la pêche, et permettre d'intensifier les opérations de pêche pour exploiter pleinement les stocks durant les périodes d'abondance élevée. Toutefois cette approche suppose que l'on dispose d'informations à jour et fiables sur ce qui se produira ensuite et que les responsables de l'aménagement et les pêcheurs soient capables de s'adapter rapidement à ces changements, à défaut de quoi l'incertitude ne ferait qu'augmenter. Mais la gestion du risque repose largement sur l'identification et l'adoption de mesures plus solides pour pallier aux incertitudes des régimes climatiques futurs et au manque d'informations. Une stratégie robuste minimise le risque qu'une décision ait des conséquences très négatives et laisse une place à l'inconnu. À cet égard, on a fait valoir qu'une stratégie robuste pour faire face aux incertitudes dérivant des effets climatiques sur les populations halieutiques marines, consisterait souvent plutôt à adopter une règle basée sur un taux de captures constant, qui autoriserait à prélever chaque année un pourcentage fixe du stock (Walters et Parma, 1996).
Les stratégies de gestion devraient aussi permettre une adaptation rapide n'engendrant pas de perturbations économiques et sociales inutiles dans le cas où le système ne réagirait pas comme le laissait présager l'évaluation (Cochrane, 2002). Par ailleurs, l'approche de précaution définit les principes généraux à adopter pour tenir compte des incertitudes dans les décisions concernant la gestion des pêches. L'approche de précaution suggère de donner la priorité à la conservation en cas d'incertitude sur la productivité des stocks. L'approche de précaution prévoit par exemple, pour les stocks de poissons caractérisés par des régimes de productivité de longue durée, d'adopter des mesures de gestion prudentes et d'ajuster les plans d'investissement en prenant pour limite le niveau de productivité élevée des poissons dans un régime antérieur. La précaution implique aussi d'éviter des changements potentiellement irréversibles et de déterminer à l'avance les effets indésirables et les mesures permettant de les prévenir ou de les corriger rapidement.
Enfin, le succès de toute stratégie de gestion visant à faire face aux variations du climat, repose sur l'information. L'absence de données à long terme est l'un des principaux obstacles qui empêchent de comprendre les causes des variations régionales de la production halieutique et de prévoir les régimes futurs. Les informations sur les changements et les impacts passés sont aussi fondamentales pour doter les institutions de gestion de la flexibilité adaptative requise car les attentes concernant les changements futurs tendent à être mieux comprises si elles peuvent être mises en parallèle avec l'expérience passée (GIEC, 2001). Les améliorations qualitatives et quantitatives de l'information sur les interactions entre le climat et la pêche reposent sur une amélioration des systèmes de surveillance des pêches, sur la mise au point de systèmes d'échantillonnage intégrés pour les propriétés physiques et biologiques de l'océan, y compris l'abondance des poissons aux premiers stades de leur vie, des progrès dans les modèles de circulation et les modèles des écosystèmes, et dans la collecte de donnée sur les répercussions sociales et économiques de la variabilité du climat et des conditions climatiques extrêmes.
RÉFÉRENCES
Bakun, A. 1990. Global climate change and intensification of coastal ocean upwelling. Science, 247: 198–200.
Bakun, A. 1996. Ocean processes and marine population dynamics. California Sea Grant and Centro de Investigaciones Biologicas, La Paz, Mexique. 323p.
Baumgartner, T.R., A. Soutar et Ferreira-Bartrina, V. 1992. Reconstruction of the history of Pacific sardine and northern anchovy populations over the past two millennia from sediments of the Santa Barbara Basin, California. CalCOFI Report, 33: 24–40.
Beamish, R.J. et Bouillon, D.R. 1993. Pacific salmon production trends in relation to climate. Can. J. Fish. Aquat. Sci., 50: 1002–1016.
Berkes, F. et Jolly, D. 2001. Adapting to climate change: Social-ecological resilience in a Canadian western Arctic community. Conservation Ecology 5(2): 18.
Beverton, R.J.H. et Holt, S.J. 1957. On the dynamics of exploited fish populations. Fishery Invest., Lond., Series 2, 19: 533p.
Conser, R.J. Hill, K.T. Crone, p.R. Lo, N.C.H. et Bergen, D. 2002. Stock assessment of Pacific sardine with management recommendations for 2003. Executive Summary. Pacific Fishery Management Council, octobre 2002. 11p.
Cochrane, K.L. 2002. A fishery manager's guidebook. Management measures and their application. FAO Fisheries Technical Paper, No. 424: 231p.
Csirke, J. 1984. Report of the working group on fisheries management, implications and interactions. In G.D. Sharp et J. Csirke, eds. Reports of the Expert Consultation to examine changes in abundance and species composition of neritic fish resources. San José, Costa Rica, 18–29 avril 1983. pp. 67–90. FAO Fisheries Report, No. 291 Vol.1: 102p.
Csirke, J. 1987. The patagonian fishery resources and the off-shore fisheries in the South-West Atlantic. FAO Fisheries Technical Paper, No. 286: 75p.
Csirke, J. 1995. Fluctuations in abundance of small and midsize pelagics. In C. Bas, J.J.Castro et J.M. Lorenzo éds. International Symposium on Middle-Sized Pelagic Fish. Sci. Mar. , 59: 481–492 (voir aussi erratum: Sci. Mar. 60: 431).
Csirke, J. et Sharp, G.D. (éds). 1984. Reports of the Expert Consultation to examine changes in abundance and species composition of neritic fish resources. San José, Costa Rica, 18–29 avril 1983. FAO Fisheries Report, No. 291 Vol.1: 102p. (http://www.fao;org/fi)
Fogarty, M.J. et Murawski, S.A. 1998. Large-scale disturbance and the structure of marine systems: fishery impacts on Georges Bank. Ecological Applications , 8(1): 6–22.
Francis, R.C. 1990. Climate change and marine fisheries. Fisheries (Bull. Amer. Fish. Soc.) , 15(6): 7–10.
Garrod, D.J. et Schumacher, A. 1994. North Atlantic cod: the broad canvas. ICES Mar. Sci. Symposium, 198: 59–76.
GIEC. 2001. Climate Change 2001: Synthesis Report. Summary for Policymakers. 34p.
Glantz, M.H. 1990. Does history have a future? Forecasting climate change effects on fisheries by analogy. Fisheries (Bull. Amer. Fish. Soc.) , 15(6): 39–45.
Gulland, J.A. 1969. Manual of methods for fish stock assessment. Part 1. Fish population analysis. FAO Manual in Fisheries Science. (4): 154p.
Jonsson, J. 1994. Fisheries off Iceland, 1600–1900. ICES Mar. Sci. Symposium, 198: 3–16.
Kawasaki, T. 1983. Why do some pelagic fishes have wide fluctuations in their numbers? Biological basis of fluctuation from the viewpoint of evolutionary ecology. In G.D.Sharp et J. Csirke, éds. Proceedings of the Expert Consultation to examine changes in abundance and species composition of neritic fish resources. San José, Costa Rica, 18–29 avril 1983. pp. 1065–1080. FAO Fisheries Report, No. 291 Vol. 3: 557–1224.
Kawasaki, T. 1992. Climate-dependent fluctuations in far eastern sardine population and their impact on fisheries and society. In M. Glantz, éd. Climate variability, climate change and fisheries , pp. 325–355. Cambridge Univ. Press.
Kawasaki, T. 1994. A decade of the regime shift of small pelagics. FAO Expert Consultation (1983) to the PICES III (1994). Bull. Japanese Soc. Fish. Ocean, 58: 321–333.
Klyashtorin, L.B. 2001. Climate change and long-term fluctuations of commercial catches: the possibility of forecasting. Rome, FAO Fisheries Technical Paper, No. 410: 86p.
Lindquist, A.V. 1983. Herring and sprat: fishery independent variations in abundance. In G.D.Sharp et J. Csirke, eds. Proceedings of the Expert Consultation to examine changes in abundance and species composition of neritic fish resources. San José, Costa Rica, 18–29 avril 1983. pp. 813–821. FAO Fisheries Report, No. 291 Vol. 3: 557–1224.
Ljungman, A.V. 1879. Bidrag till lösningen of fragan om de stora sillfiskenas sekulära periodicitet. Nord. Tidskr. f. Fiskeri : pp. 257–268.
Lluch-Belda, D. Hernandez-Vazquez, S., Salinas-Zavala, C. et Shwartzlose, R. 1992. The recovery of the California sardine as related to global change. Calif. Coop. Oceanic Fish. Invest. Rep., 33: 50–59.
MacCall, A.D. 1980. Population models for the northern anchovy (Engraulis mordax). Rapp.p.-V. Réun. CIEM, 177: 282–306.
Peterman, R.M., Pyper, B.J., Lapointe, M.F., Adkison, M.D. et Walters, C.J. 1998. Patterns of covariation in survival rates of British Columbian and Alaskan sockeye salmon (Oncorhynchus nerka) stocks. Can. J. Fish. Aquat. Sci., 55: 2503–2517.
Ravier, C. et Fromentin, J.M. 2001. Long-term fluctuations in the eastern Atlantic and Mediterranean bluefin tuna populations. ICES Journal of Marine Science 58: 1299–1317.
Ravier, C. et J-M. Fromentin, J.M. 2004. Are the long-term fluctuations in Atlantic bluefin tuna (Thunnus thynnus) population related to environmental changes. Fisheries Oceanography , 13(3):145–160.
Regier, H.A. et Meisner, J.D. 1990. Anticipated effects of climate change on freshwater fishes and their habitats. Fisheries (Bull. Amer. Fish. Soc.) , 15(6): 10–15.
Ricker, W.E. 1958. Handbook of computations for biological statistics of fish populations. Bull. Fish. Res. Bd. Can., 119: 300p.
Russell, E.S. 1931. Some theoretical considerations on the overfishing problem. J.Cons. Perm. Int. Explor. Mer., 6(1): 3–27.
Sharp, G.D. 2003. Future climate change, and regional fisheries: a collaborative analysis. FAO Fisheries Technical Paper, No. 452: 75p.
Sharp, G.D. et Csirke, J. (éds) 1983. Proceedings of the Expert Consultation to examine changes in abundance and species composition of neritic fish resources. San José, Costa Rica, 18–29 avril 1983. FAO Fisheries Report, No. 291 Vols. 2–3: 1224p.
Vasconcellos, M.C. 2000. Ecosystem impacts of fishing forage fishes. An analysis of harvesting strategies for the Brazilian sardine. PhD Thesis. Université de Colombie Britannique. Canada. 190p.
Walters, C.J. et Parma, A.M. 1996. Fixed exploitation rate strategies for coping with effects of climate changes. Can. J. Fish. Aquat. Sci., 53: 148–158.
* FAO, Service des ressources marines, Division des ressources halieutiques.
![]()
![]()
![]()