![]()
![]()
![]()
Ana María Caramelo y Merete Tandstad *
INTRODUCCIÓN
El Atlántico centro-oriental, situado frente a la costa occidental de África, desde el Estrecho de Gibraltar hasta la desembocadura del río Zaire (Figura B4.1), abarca un total de 14,2millones de km2. La plataforma continental es en general estrecha, con sólo 0,65millones de km2 para toda la zona. El área34 comprende aguas templadas, tropicales y ecuatoriales, lagunas y manglares, así como peculiaridades oceanográficas, como importantes corrientes, afloramientos y la convergencia ecuatorial. Durante el período 1950 a 2002 se recibieron informaciones sobre unas 190especies o grupos de especies, presentadas por 22Estados ribereños y 47países que pescan en aguas distantes.
Figura B4.1 - Atlántico centro - oriental (Área 34)
En las pesquerías de esta región predominan las pequeñas especies pelágicas, en particular la sardina europea (Sardina pilchardus) y otros clupeidos (Grupo35 de la CEIUAPA), que representaban más del 52por ciento del total de las capturas nominales en 2002.
En los últimos años del decenio de 1980, los cambios ocurridos en Europa oriental dieron lugar inicialmente a la aparición de varias naciones con flotas dedicadas a la peaca en aguas distantes. Posteriormente, el mayor hincapié en las fuerzas del mercado ha favorecido una reducción de las actividades de algunas de esas flotas especializadas en pequeñas especies pelágicas. Durante los cinco últimos años, la intensidad de la pesca de pequeñas especies pelágicas por las flotas de la Unión Europea ha aumentado en la parte noroccidental de esta zona.
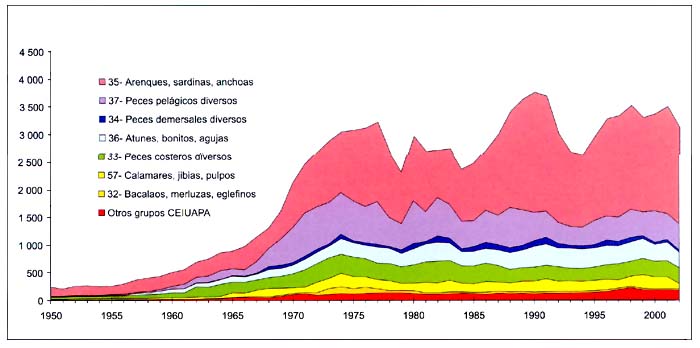
Figura B4.2 - Capturas nominales anuales ('000t) por grupos de especies CEIUAPA en el Atlántico centro-oriental (Área 34)
Fuente FAO
PERFIL DE LAS CAPTURAS
El total nominal de las capturas registrado en el área34 se multiplicó casi por 12, pasando de unas 300000 toneladas en 1950 a casi 3,6millones de toneladas en 1977. Desde entonces, las capturas han oscilado de forma relativamente suave, entre 2,5millones de toneladas en 1979 y 4,1millones de toneladas en 1990, debido a los cambios ocurridos en los mercados y en el esfuerzo de pesca de las flotas que faenan en aguas distantes, y a cambios inducidos por el clima en la productividad de las poblaciones, factores todos ellos que han tenido repercusiones sobre todo en las capturas del grupo35 de la CEIUAPA. Las capturas se han mantenido relativamente estables, con un promedio de 3,6millones de toneladas desembarcadas desde 1995 (Figura B4.2 y CuadroD4), y 3,4millones de toneladas en 2002.
Destacan por su importancia en las capturas las especies del grupo35 de la CEIUAPA (arenques, sardinas, anchoas, etc.), que representan casi la mitad del total (Figura B4.2). Los desembarques de este grupo descendieron ligeramente a 1,7millones de toneladas en 2002, frente a 2,2millones en 1990.
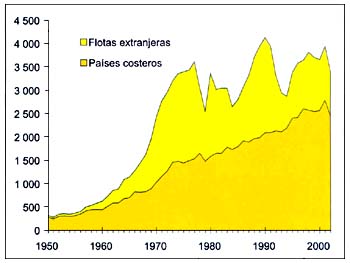
Los Estados ribereños desarrollaron de forma constante sus pesquerías nacionales, pasando de representar el 43por ciento del total de las capturas en esta zona a un 72por ciento entre 1977 y 2002 (Figura B4.3). Las flotas que faenan en aguas distantes, dedicadas sobre toda a la captura de pequeñas especies pelágicas y túnidos, han realizado contribuciones importantes pero irregulares desde comienzos del decenio de 1970.
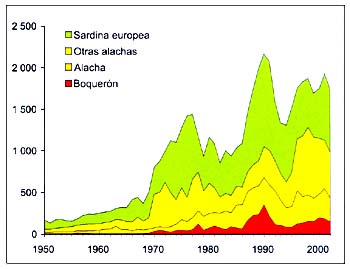
Cuatro categorías de especies representan más del 80por ciento de las capturas en el grupo35 de la CEIUAPA (arenques, sardinas, anchoas, etc.): la sardina europea (Sardina pilchardus) constituye la mayor contribución, seguida de otros arenques, la alacha (Sardinella aurita) y el boquerón (Engraulis encrasicolus) (Figura B4.4). Sardinella spp. representa también una parte considerable del total del grupo y de la zona.
En la figura B4.5 pueden verse las capturas del grupo37 de la CEIUAPA (peces pelágicos diversos). Las capturas de Trachurus spp., la especie predominante, tuvieron un fuerte aumento al final del decenio de 1960, se mantuvieron altas durante la mayor parte del decenio de 1970, y descendieron durante casi todo el decenio de 1980 y el de 1990. Las capturas de estornino (Scomber japonicus) tuvieron dos años excepcionales en 1988 y 1989, seguidos de una caída vertical de las capturas en 1993, una recuperación en 1997 y una evolución fluctuante desde entonces.
La merluza del Senegal (Merluccius senegalensis) ha representado la contribución más importante en el grupo32 de la CEIUAPA (bacalaos, merluzas, eglefinos), con capturas de más de 100000 toneladas en el decenio de 1970 y relativamente estables, pero a un nivel mucho más bajo desde entonces (CuadroD4).
Figura B4.3 - Capturas nominales anuales ('000t) por países costeros y flotas extranjeras, Atlántico centro-oriental (Área 34)
Fuente FAO
Figura B4.4 - Capturas nominales anuales ('000t) de especies seleccionadas del grupo CEIUAPA 35, Atlántico centro-oriental (Área 34)
Fuente FAO
Figura B4.5 - Capturas nominales anuales ('000t) de especies seleccionadas del grupo CEIUAPA 37, Atlántico centro-oriental (Área 34)
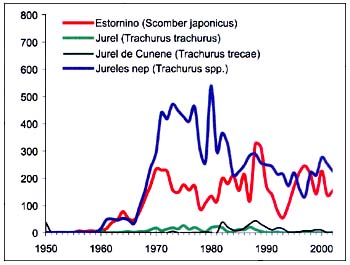
Fuente FAO
Las capturas del grupo36 de la CEIUAPA (atunes, bonitos, agujas, etc.), reflejadas en la Figura B4.6, presentan tendencias semejantes a lo largo del tiempo debido al comportamiento de las flotas y a la influencia de los fenómenos climáticos en la productividad de los recursos. El listado (Katsuwonus pelamis) se ha convertido en el elemento más importante, superando en general al rabil (Thunnus albacares) desde 1991 en adelante. En 2002, las capturas de rabil fueron ligeramente más abundantes.
Figura B4.6 - Capturas nominales anuales ('000t) de especies seleccionadas del grupo CEIUAPA 36, Atlántico centro-oriental (Área 34)
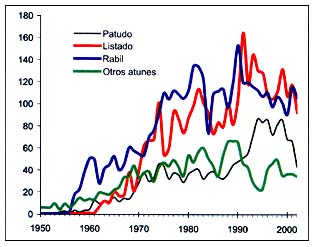
Fuente FAO
Figura B4.7 - Capturas nominales anuales ('000t) de especies seleccionadas del grupo CEIUAPA 45, Atlántico centro-oriental (Área 34)
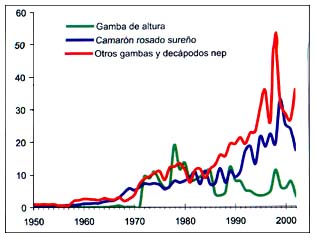
Fuente FAO
Figura B4.8 - Capturas nominales anuales ('000t) de especies seleccionadas del grupo CEIUAPA 57, Atlántico centro-oriental (Área 34)
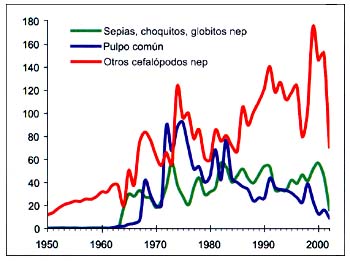
Fuente FAO
En la Figura B4.7 pueden verse las principales capturas de especies incluidas en el grupo45 de la CEIUAPA (camarones, langostinos, etc.). Las capturas de camarón rosado sureño (Penaeus notialis) comenzaron a ser significativas en el decenio de 1960 y posteriormente han aumentado, con algunas oscilaciones y con un máximo muy pronunciado, de 33000 toneladas en 1999, y 17000 toneladas en 2002. La pesquería del camarón de altura (Parapenaeus longirostris) comenzó en 1972 y las capturas han sido variables desde entonces, con un máximo de 19000 toneladas en 1978. En 1998, se registraron en la región capturas de 11000 toneladas de esta especíe. En 2000 fueron de 6000 toneladas y en 2002 de 3000 toneladas.
La pesquería del pulpo común (Octopus vulgaris) del grupo 57 de la CEIUAPA (Calamares, sepias, pulpos) comenzó en 1962 y alcanzó las 93000 toneladas en 1975. Luego las capturas han disminuido periódicamente hasta situarse en 9000 toneladas en 2002 (Figura B4.8). Las capturas de sepia han fluctuado, con un promedio de 44000 toneladas desde 1990 hasta 2001, pero descendieron a 16000 toneladas en 2002. Las capturas de otros cefalópodos nep. han presentado en general un aumento, habiendo subido de 60000 toneladas en 1992 a 152000 toneladas en 2001, seguidas de un fuerte descenso en 2002 (71000 toneladas).
SITUACIÓN DE LOS RECURSOS Y ORDENACIÓN PESQUERA
Este examen de la situación de las poblaciones está basado en evaluaciones llevadas a cabo por recientes grupos de trabajo sobre la evaluación de los recursos de las grandes pesquerías de la región y en las conclusiones del Subcomité Científico para el Atlántico Centro-Oriental (Lomé, Togo, 24–26 de febrero de 2004) y del Comité de Pesca para el Atlántico Centro-Oriental (CPACO), Dakar, Senegal 24–27de mayo de 2004. La situación de muchas poblaciones se ha evaluado durante las reuniones de los grupos de trabajo celebradas entre 1998 y 2004.
Dieciocho de las poblaciones demersales que fueron evaluadas estaban plenamente explotadas o sobreexplotadas. Una de las poblaciones, Epinephelus aeneus , que se encuentra fundamentalmente en Mauritania, Senegal y Gambia, estaba sobreexplotada de tal forma que se ha considerado cercana a la extinción. La Sardinella aurita en la región norte del CPACO muestra signos de sobreexplotación (FAO, 2004b).
Una conclusión general es que, como medida precautoria, el actual esfuerzo de pesca debería mantenerse o incluso reducirse para todas las especies demersales y pelágicas de la región, incluido el camarón.
Zonas septentrionales
En general, la mayoría de los recursos demersales situados frente a la costa del África noroccidental se consideran de plenamente explotados a sobreexplotados (FAO, 2003c; FAO, 2004a, b).
Las merluzas (Merluccius spp.) de la plataforma continental de Mauritania se consideran plenamente explotadas. Entre los otros recursos de peces de escama evaluados, la población de Epinephelus aeneus corre especial peligro y sería necesario tomar medidas inmediatas (FAO, 2003c).
Tres poblaciones de pulpo común (Octopus vulgaris) son capturadas por pesquerías de cefalópodos del África noroccidental, a saber, las poblaciones de Dakhla, Cabo Blanco, y la denominada Senegambia. Tanto la septentrional, frente a la costa de Dakhla, como la central, frente a Cabo Blanco, se consideran sobreexplotadas, con capturas recientes de aproximadamente 50000 toneladas en lo que respecta a la población de Dakhla y de 19000 toneladas para la de Cabo Blanco. El estado de la población que se encuentra más al sur, frente a las costas del Senegal y de Gambia, es incierto. Las capturas de esta población son inferiores a las de las otras dos (FAO, 2003c).
La explotación de otras especies de cefalópodos, como calamares (Loligo vulgaris) y sepias (Sepia officinalis), es menos intensa que la del pulpo: sus capturas representan el 30por ciento de los cefalópodos desembarcados en 2002. Los desembarques de las capturas de pulpo y las correspondientes capturas incidentales de las pesquerías de cefalópodos del África noroccidental se han desplazado durante los últimos años de Las Palmas, en las islas Canarias, a otros puertos del África noroccidental. Las medidas de ordenación adoptadas en la zona económica exclusiva (ZEE) de Marruecos incluyen una veda de dos meses y una reducción del número de embarcaciones no marroquíes que faenan en la zona.
Las capturas de camarón de altura (Parapenaeus longirostris) y de camarón rosado sureño (Penaeus notialis) realizadas en la región norte del CPACO son de aproximadamente 18000 toneladas y parece ser que las poblaciones están fuertemente explotadas o incluso sobreexplotadas (FAO, 2003c).
En la zona de Mauritania, las poblaciones de langosta mora (Palinures mauritanicus) continúan estando fuertemente explotadas. Las poblaciones septentrionales y meridionales de langosta real (Panulirus regius) están probablemente sobreexplotadas (FAO, 2003b). Las poblaciones de gerión de Guinea (Chaceon maritae) parecen estar fuertemente explotadas.
La biomasa de las grandes poblaciones de pequeñas especies pelágicas en la región septentrional del CPACO - sardina (Sardina pilchardus), sardinela (Sardinella aurita y Sardinella maderensis), estornino (Scomber japonicus) y jurel (Trachurus spp.)- es muy variable.
Las prospecciones acústicas realizadas frente a las costas del África noroccidental en noviembre–diciembre del período 1995–2002 revelaron que la densidad por cardumen de las pequeñas especies pelágicas (sobre todo sardinela, con exclusión de la sardina) era muy elevada, en particular frente a las costas de Mauritania, con una biomasa estimada en aproximadamente 3,5millones de toneladas. Además, la biomasa estimada mediante prospecciones acústicas en los casos del estornino, el jurel y otras pequeñas especies pelágicas de la zona comprendida entre Marruecos y el Senegal era de aproximadamente dos millones de toneladas en las campañas de prospección de noviembre–diciembre realizadas durante el período 1995–2002 (FAO, 2003a).
La biomasa de la sardina estimada con métodos acústicos al norte de Cabo Blanco acusaba un descenso del 80por ciento entre 1996 y 1997. La biomasa de la sardina al norte de Cabo Blanco y en aguas de Mauritania se estimó en 3–5millones de toneladas durante el período 1986–1996. Desde 1999, la biomasa en esta región ha aumentado gradualmente, y se estimaba aproximadamente en 4,5millones de toneladas en diciembre de 2002. Las flotas de España y Rusia dejaron de pescar en aguas de Marruecos en 1998, al no renovarse los acuerdos de pesca entre Marruecos y Rusia y entre Marruecos y la UE. En términos generales, se considera que la tasa de explotación ha disminuido como consecuencia de ello, pero la sardina se explota también en aguas de Mauritania (FAO, 2003a).
Las capturas combinadas de pequeñas especies pelágicas en el África noroccidental no deberían aumentarse más allá del promedio de los cinco últimos años, con exclusión de la sardina (Sardina pilchardus) en la zona C (FAO, 2004a). Los diferentes grupos de especies de sardinelas, jureles y estorninos se explotan ya en forma intensa (FAO, 2004a).
En aguas de Cabo Verde, las pesquerías más importantes son las de túnidos, con capturas medias estimadas de 4000 toneladas en los 10últimos años. Las poblaciones de langosta de Cabo Verde (Palinurus charlestoni) se consideran ahora sobreexplotadas y se ha recomendado que no siga aumentando el esfuerzo de pesca. En 2001, la captura de especies demersales representó un 21por ciento del total de capturas realizado por la pesca artesanal. Los potenciales estimados de las especies demersales son de 3000 a 5000 toneladas y los de las pequeñas especies pelágicas de 4500 a 6500 toneladas (Carvalho, Morais y Nascimento, 1999).
Zonas meridionales
Las plataformas continentales de Guinea Bissau, Guinea y Sierra Leona se caracterizan por las congregaciones de corvinones que se encuentran principalmente en zonas costeras y de estuarios ricos en nutrientes.
La tasa de explotación de las poblaciones de sepia en Guinea ha aumentado desde los años noventa; y en 2001, con capturas de 5800 toneladas, las poblaciones se consideraban sobreexplotadas (FAO, 2003c).
La producción anual actual de camarón rosado sureño en Guinea y Sierra Leona es de aproximadamente 2000 toneladas. Se considera que esta especie está moderadamente explotada en Sierra Leona, mientras que en Guinea se considera sobreexplotads (FAO, 2003c).
Los recursos marinos del Golfo de Guinea son explotados fundamentalmente por Côte d'Ivoire, Ghana, Togo, Benin, Nigeria, Camerún y Guinea Ecuatorial, entre otros países. Las pesquerías de especies múltiples son comunes en el Golfo de Guinea. Los pequeños recursos pelágicos son explotados sobre todo mediante redes de cerco de jareta artesanales y semiindustriales en Côte d'Ivoire, Ghana, Togo y Benin, y exclusivamente por pesquerías a pequeña escala en Nigeria y el Camerún. Los recursos demersales costeros están formados por esciénidos (explotados mediante pesquerías a pequeña escala y semiindustriales en Nigeria, Benin, Togo y el Camerún), meros y pargos (pescados en Togo y Ghana con sedal y anzuelo en las zonas donde no es posible la pesca de arrastre) y espáridos (Côte d'Ivoire y Ghana). Frente a las costas de Nigeria y el Camerún los recursos de camarón blanco son explotados exclusivamente mediante pesquerías artesanales, mientras que el camarón rosado se explota con arrastreros de la pesquería semiindustrial. Los camarones peneidos de Togo y Benin y de Côte d'Ivoire se capturan en las pesquerías de laguna. Los recursos demersales de altura de Ghana y Côte d'Ivoire están integrados por espáridos junto con la comunidad del talud, mientras que los recursos demersales de altura de Nigeria y el Camerún son fundamentalmente la arioma (Arioma spp.) y las gallinetas (Scorpaenidae).
Los sondeos mediante arrastre realizados en la plataforma continental del Golfo de Guinea por el buque de investigación Dr. Fridtjof Nansen han revelado que entre 1999 y 2001 la biomasa estimada de recursos demersales osciló a lo largo del período entre 17500 toneladas y 39500 toneladas. En el Golfo de Guinea, desde Côte d'Ivoire hasta el Camerún, el potencial estimado de camarones fue de 10000 toneladas, y se consideró que las poblaciones estaban fuertemente explotadas (FAO, 1999).
Los recursos demersales estan plenamente explotados, o sobreexplotados (FAO, 2004a, b). En consideración de las incongruencias existentes en la consignación de los datos, el Grupo de trabajo sobre recursos demersales del CPACO recomendó que se adoptara un enfoque precautorio, evitando todo aumento del esfuerzo de pesca, para todas las especies demersales del Golfo de Guinea (FAO 2003c, 2004a, b).
Las pequeñas especies pelágicas (sardinelas, estorninos o caballas y anchoas) son recursos importantes pero inestables en el Golfo de Guinea occidental, compartidos por Côte d'Ivoire, Ghana, Togo y Benin, lo que complica todavía más la ordenación de las pesquerías que las explotan. Los potenciales de las pequeñas especies pelágicas en el Golfo de Guinea occidental y central se han estimado en 330000 toneladas y las poblaciones parecen estar plenamente explotadas. Muchos países no han desarollado estructuras de investigación ni bases de datos adecuadas para evaluar la situación de las poblaciones y su estado de explotación.
REFERENCIAS
Carvalho, M.E.M., Morais J. y Nascimento J. (eds.) 1999. Investigação e gestão haliêuticas em Cabo Verde. Actas da reunião realizada em Mindelo, 10e 11de Dezembro de 1996). Mindelo, Cabo Verde. Instituto Nacional de Desenvolvimento das Pescas. 252págs.
FAO. 1997. Rapport du Groupe the Travail ad hoc sur les Céphalopodes. Tenerife,19–26de mayo de 1997. M.Lamboeuf (ed.) COPACE/PACE/ Ser. , №97/63. Roma, FAO. 1997. 103págs.
FAO. 1998a. Report of the Workshop on the Review of the Pelagic Surveys off Northwest Africa in the 1990'fs. Bergen, Noruega, 28de septiembre– 2de octubre de1998. FAO Informe de Pesca. №592. 63págs.
FAO. 1999. Report of the Workshop on the Assessment and Management of Shrimps and Crabs in Southwest Africa. Luanda, Angola 8–12de marzo de1999. Proyecto GCP/RAF/302/EEC, Improvement of the legal framework for fisheries cooperation, management and development of coastal States of West Africa. 107págs. Documento № 62.
FAO.2003a. Report of the FAO Working Group on the Assessment of Small Pelagic Fish off Northwest Africa. Agadir, Marruecos, 31de marzo – 10de abril de2003. FAO Fisheries report. №723:152págs.
FAO.2003b. Evaluation des stocks et aménagement des pêcheries de la ZEE mauritanienne. Rapport du 5ème groupe de travail IMROP. Nouadhibou, Mauritania, 9–17de diciembre de 2002. En prensa.
FAO. 2003c. Report of the CECAF Working Group on the assessment of demersal resources. Conakry, Guinea, 19–29de septiembre de2003. En prensa.
FAO 2004a. Comité de Pesca para el Atlántico Centro-Oriental. Informe de la tercera reunión del subcomité científico. Lomé, Togo, 24–26de febrero de2004. FAO Informe de Pesca. № 750. Accra, FAO. 2004. 35págs.
FAO. 2004b. Informe de la 17a reunión del Comité de Pesca para el Atlántico Centro-Oriental. Dakar, Senegal, 24–27de mayo de2004. FAO Informes de pesca. №754. 57pags.
* FAO, Servicio de Recursos Marinos, Dirección de Recursos Pesqueros.
Jordi Lleonart *
INTRODUCCIÓN
El Mediterráneo (Figura B5.1) es un mar semicerrado con una superficie de aproximadamente 3,3millones de km2, que representan el 0,8por ciento del total de la superficie marina mundial. Debido a su situación geográfica, dentro de un margen de latitud relativamente estrecho (de 30°N a 46°N) de la zona templada del hemisferio norte, el Mar Mediterráneo tiene un pronunciado ciclo estacional. Las masas de aguas están estratificadas en verano, pero la temperatura de las aguas profundas (por debajo de 400m) es de 13°C ± 0,3°C durante todo el año. La escasez de precipitaciones durante el verano es la característica principal del clima mediterráneo. Este mar tiene un balance hídrico negativo: la pérdida de agua mediante la evaporación es mayor que las aportaciones debidas a las precipitaciones y la escorrentía de los ríos, pero se compensa con la aportación de unos 1700km3/año de agua del Atlántico a través del Estrecho de Gibraltar (Oliver, 2003).
Figura B5.1 - Mediterráneo y Mar Negro (Área 37)
El sistema de cuencas incluye aguas que van desde las de origen atlántico en el Mediterráneo occidental hasta el agua hipersalina templada de la cuenca del Levante, las aguas hiposalinas del Mar Negro, y las aguas frías-templadas de estuario en el Mar de Azov. El agua superficial que entra desde el Atlántico a través del estrecho de Gibraltar avanza hacia el Mediterráneo oriental, perdiendo progresivamente nutrientes y aumentando su salinidad debido a la evaporación. Con el tiempo, vuelve al Atlántico para formar el componente altamente salino de la circulación atlántica. Ello permite un gran número de hábitats con muchas poblaciones locales de pequeño tamaño y especies cuyas tendencias no pueden describirse por separado aquí pero presentan diferencias en las diversas subzonas (Garibaldi y Caddy, 1998).
Figura B5.2 - Capturas nominales anuales ('000t) por grupos de especies CEIUAPA en el Mar Mediterráneo y el Mar Negro (Área 37)
Fuente FAO
El Mediterráneo se ha considerado globalmente como un mar oligotrófico (Margalef, 1985; Estrada, 1996; Stergiou et al. , 1997b). Normalmente, la disminución gradual de los nutrientes daría lugar a un descenso de productividad de oeste a este, pero hay excepciones locales resultantes de un gradiente de productividad norte-sur, que se hace patente también en las imágenes de la clorofila A obtenidas mediante satélite, debido a los nutrientes procedentes de los ríos septentrionales (en particular el Ródano, el Po y la entrada de agua del Mar Negro en el Egeo). Estos efectos antropogénicos en la producción pesquera han sido descritos por Caddy, Refk y Do Chi (1995).
La plataforma continental es fundamentalmente una estrecha franja costera, con excepción del Mar Adriático, el Golfo de Gabés, el norte del Mar Negro, el sur de Sicilia y el Golfo de León, y representa sólo el 23por ciento de la superficie total.
Las pesquerías del Mediterráneo han mostrado una sorprendente capacidad de recuperación en comparación con algunas zonas del Atlántico. Ello resulta especialmente notable si se tiene en cuenta que en la mayor parte de los países mediterráneos no se ha adoptado prácticamente ninguna medida oficial y coordinada de ordenación de la pesca, aunque desde 2000 se intenta corregir esa situación con actividades más frecuentes y una revisión del mandato de la Comisión General de Pesca del Mediterráneo (CGPM) y de su Comité Científico Asesor (SAC). Estas pesquerías han sido estudiadas, entre otros, por Farrugio et al. (1993), Farrugio (1996), Anón. (2001), Bas (2002), Oliver (2003) y Lleonart y Maynou (2003).
PERFIL DE LAS CAPTURAS
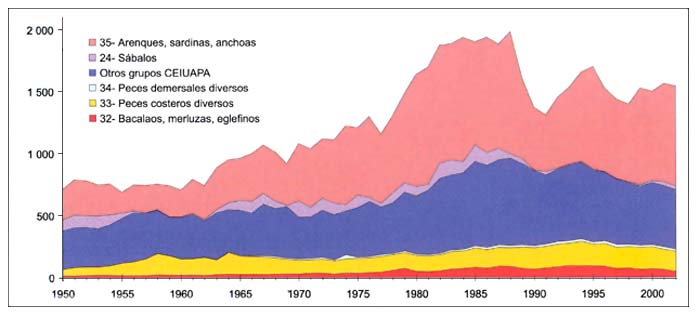
Las capturas nominales en el área37 aumentaron de poco más de 0,7millones de toneladas en 1950 a casi dos millones de toneladas entre 1982 y 1988 (Figura B5.2 y CuadroD5). Posteriormente, las capturas disminuyeron con rapidez llegando a 1,3millones de toneladas como consecuencia del colapso de la pesquería del espadín y el boquerón en el Mar Negro (Figura B5.3). El total de las capturas ha aumentado posteriormente a 1,5millones de toneladas en 2001 y 2002, después de algunas pequeñas fluctuaciones.
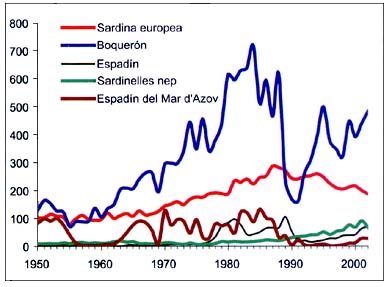
Las pequeñas especies pelágicas representan aproximadamente el 50por ciento del total de las capturas mediterráneas. El boquerón (Engraulis encrasicolus), con el 59por ciento de las capturas de pequeñas especies pelágicas, y la sardina (Sardina pilchardus), con el 16por ciento, son las más abundantes. Otras pequeñas especies pelágicas son el espadín (Sprattus sprattus), la alacha (Sardinella aurita) y el espadín del Mar de Azov (Clupeonella cultriventris).
El considerable aumento de las capturas de boquerón desde 1960 hasta mediados del decenio de 1980 (Figura B5.3) se debe en parte al aumento del esfuerzo de Turquía en el Mar Negro, que se sumó al de la flota de la antigua Unión Soviética que predominaba anteriormente en esta pesquería. Como en el caso del espadín, ello podría ser consecuencia de la eutrofización del Mar Negro por los ríos que desembocan en él (Mee, 1992; Zaitsev, 1993). El colapso del boquerón, el espadín y el espadín del Mar de Azov en 1990 se debió a la introducción de una especie de ctenóforo, Mnemiopsis leidy, en el Mar Negro. El aumento posterior de las capturas no alcanzó los niveles precedentes. La serie de la sardina presenta una leve tendencia ascendente hasta finales del decenio de 1980 (282000 toneladas en 1988) y un descenso moderado posterior, situándose en unas 187000 toneladas en 2002.
Figura B5.3 - Capturas nominales anuales ('000t) de especies seleccionadas, de los grupos CEIUAPA 24 y 35, Mar Mediterráneo y Mar Negro (Área 37)
Fuente FAO
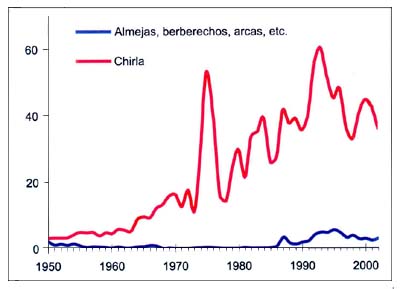
Las pesquerías mixtas de fondo basadas en la utilización de arrastres de malla pequeña, redes de enmalle, trasmallos, trampas, nasas y rastras capturan una gran variedad de peces demersales invertebrados de alto valor para el mercado de pescado fresco, pero ninguna de esas especies representa más del 10por ciento del total de las demersales. No obstante, en las zonas en que las flotas de arrastre faenan habitualmente, a pesar de la inherente complejidad de las capturas de especies múltiples, hay una serie identificable de especies primarias que en términos de biomasa o económicos constituyen una base importante de producción. Las especies consideradas como demersales (más de 100, aunque algunas muestran un comportamiento pelágico) representan aproximadamente el 40por ciento del total de las capturas registradas en el Mediterráneo y el Mar Negro. La merluza (Merluccius merluccius), los salmonetes (Mullus spp.), la bacaladilla (Micromesistius poutassou), el plegonero (Merlangius merlangus), los rapes (Lophius spp.), las brecas (Pagellus spp.), la boga (Boops boops), las chuclas (Spicara spp.), la chirla (Chamelea gallina), el pulpo (Octopus spp.), la sepia (Sepia officinalis), la gamba rosada (Aristeus antennatus), la cigala (Nephrops norvegicus) y el camarón de altura (Parapenaeus longirostris) son las principales especies demersales.
Figura B5.4 - Capturas nominales anuales ('000t) de especies seleccionadas, del grupo CEIUAPA 56, Mar Mediterráneo y Mar Negro (Área 37)
Fuente FAO
Figura B5.5 - Capturas nominales anuales ('000t) de especies seleccionadas, de los grupos CEIUAPA 45 y 57, Mar Mediterráneo y Mar Negro (Área 37)
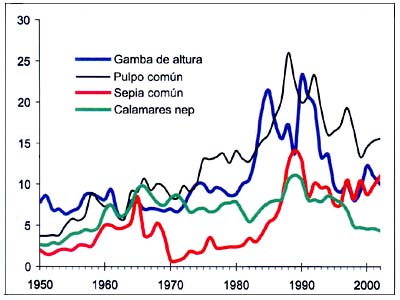
Fuente FAO
Figura B5.6 - Capturas nominales anuales ('000t) de especies seleccionadas, de los grupos CEIUAPA 32 y 33, Mar Mediterráneo y Mar Negro (Área 37)
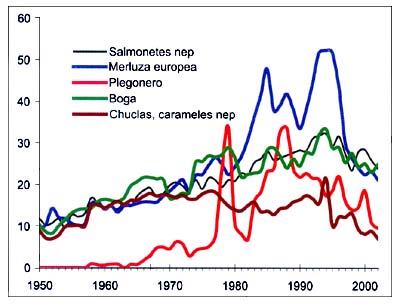
Fuente FAO
Entre las especies capturadas incidentalmente se incluyen especies no registradas individualmente en las capturas oficiales o que no siempre están presentes, pero que en conjunto pueden representar una proporción significativa de las capturas, como los tiburones pequeños (Carbonell et al. , 2003).
Hay también otras especies locales y/o de precio elevado que se capturan incidentalmente en cantidad relativamente pequeña, como algunas especies de peces planos, escorpaenidos, espáridos, tríglidos, mujílidos, serránicos, y algunos invertebrados. El último grupo de capturas incidentales está formado por especies que a veces son también relativamente abundantes pero cuyo precio es elevado, como algunos escómbridos y carángidos, o especies que tienen poca aceptación en el mercado, como la boga o las chuclas (Spicara spp.). También los descartes son importantes, y diversas las razones de este fenómeno: tamaños ilegales, especies o tamaños no comerciales, o razones de mercado (Carbonell et al. , 1998; Lleonart y Maynou, 2003).
Conviene señalar el espectacular aumento de las capturas de chirla (Chamelea gallina), sobre todo en el Adriático, que alcanzó un máximo en 1993 (Figura B5.4). Ello podría ser resultado de la interacción de la pesca con la transición de condiciones oligotróficas en el período inicial a situaciones casi mesotróficas.
Las capturas de cefalópodos (Figura B5.5) presentan un máximo en torno a 1990, quizás debido a la menor abundancia de grandes peces depredadores, como opinan Caddy y Rodhouse (1998), pero en la mayor parte de los casos éstos se trata de componentes de una pesquería mixta de peces de fondo. El comienzo de las actividades de arrastre en aguas profundas en las zonas de talud a mediados del decenio de 1980 parece explicar el fuerte aumento de las capturas de camarón de altura, que se habrían beneficiado de una reducción de la biomasa de las grandes merluzas en las zonas de talud.
La mayoría de los gádidos y de otras especies semipelágicas, como la boga y la chucla (Spicara spp.), así como los salmonetes (Mullus spp.), han presentado una tendencia ascendente constante durante todo el período, con excepción del descenso de varias especies en los últimos años. Es más notable el caso de la merluza cuyas capturas, después de alcanzar máximos históricos a mediados del decenio de 1990 (más de 52000 toneladas), han descendido a menos de la mitad en 2002. El plegonero, sólo presente en el Mediterráneo oriental, sigue también esa pauta general, con un máximo en torno a 1990 (Figura B5.6).
Las principales especies pelágicas de gran tamaño son el atún rojo (Thunnus thynnus) y el pez espada (Xiphias gladius). Representan el 3por ciento del total de las capturas, pero su valor económico es mucho mayor. Las capturas de atún rojo aumentaron entre mediados del decenio de 1960 y de 1990, pero luego descendieron de forma significativa (Figura B5.7). La pesquería del atún rojo en el Mediterráneo es motivo de preocupación debido a la gran expansión de las actividades de cría/engorde, de ejemplares capturados, que contribuyen al aumento de la presión pesquera sobre este recurso abriendo nuevos mercados, y es posible que no todas las capturas se registren. Las capturas de pez espada aumentaron de forma constante desde 1950 hasta comienzos del decenio de 1980, y su volumen se duplicó con creces entre 1983 (7000 toneladas) y 1989 (18000 toneladas), en parte como consecuencia de la mejora de los sistemas nacionales de recopilación de estadísticas. Las capturas volvieron a situarse entre 12000 y 16000 toneladas desde el decenio de 1990. Las capturas de atún blanco (Thunnus alalunga) siguen una pauta semejante, aunque con menores cantidades. En el caso del bonito (Sarda sarda), las capturas son muy variables de un año a otro, y presentan una ligera tendencia descendente a largo plazo.
Las capturas de especies pelágicas de tamaño mediano representan el siete por ciento del total de las capturas, y comprenden caballas y jureles (Trachurus spp.), el jurel mediterráneo (Trachurus mediterraneus), el jurel del Atlántico (Trachurus trachurus), el estornino (Scomber japonicus) y la caballa del Atlántico (Scomber scombrus). Las capturas del jurel mediterráneo son también muy variables, y no se observa una tendencia definida. El estornino alcanzó cifras máximas de mediados del decenio de 1980 a mediados del decenio de 1990. Las especies de laguna y costeras, como las lisas y los pejerreyes, presentan tendencias semejantes, aunque en menores cantidades (Figura B5.8).
Figura B5.7 - Capturas nominales anuales ('000t) de especies seleccionadas, del grupo CEIUAPA 36, Mar Mediterráneo y Mar Negro (Área 37)
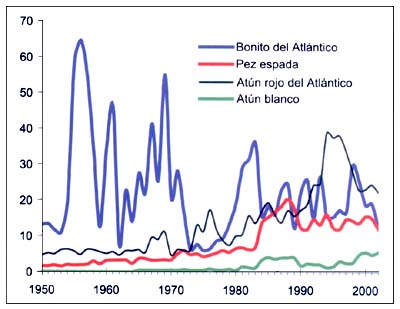
Fuente FAO
Figura B5.8 - Capturas nominales anuales ('000t) de especies seleccionadas, de los grupos CEIUAPA 33 y 37, Mar Mediterráneo y Mar Negro (Área 37)
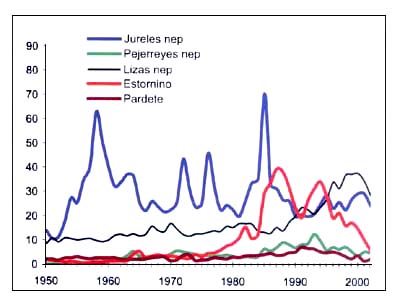
Fuente FAO
La disminución de las capturas en los cinco últimos años es una tendencia general de las especies no pelágicas. Después de 25años de constante aumento, alcanzaron un máximo de aproximadamente 700000 toneladas en 1994, habiendo disminuido lentamente desde entonces, hasta situarse en menos de 400000 toneladas en 2002.
Las especies pelágicas muestran una tendencia diferente, marcada por los extraordinarios acontecimientos del Mar Negro. Después de un fuerte aumento de 700000 toneladas a 1,3millones de toneladas en seis años, se mantuvieron en esa cifra desde 1983 hasta 1988 y luego han oscilado, siguiendo una tendencia descendente, hasta situarse en 990000 toneladas en 2002.
Las capturas del Mar Negro representaron el 39por ciento del total de las capturas mediterráneas en 2002, y un 57por ciento del total de las capturas de pequeñas especies pelágicas.
SITUACIÓN DE LOS RECURSOS Y ORDENACIÓN PESQUERA
El grado de exactitud de las series de datos cronológicos sobre capturas en el Mediterráneo es variable, pero los datos sobre el esfuerzo son prácticamente inexistentes. No obstante, dada la complejidad y diversidad de las pesquerías mediterráneas, probablemente los datos disponibles no son suficientes para realizar evaluaciones periódicas y fiables de la mayoría de las especies. La estructura misma de las pesquerías mediterráneas (dispersión de las flotas, número enorme de puntos de desembarque, capturas multiespecíficas y falta de grandes industrias como las existentes en otros mares) hace difícil y costosa la obtención de datos para evaluar las poblaciones.
Los parámetros biológicos de las principales especies (merluza, boquerón, sardina, salmonetes, algunos espáridos y peces planos, atún rojo, pez espada, algunos crustáceos de valor elevado) se conocen bastante bien. No obstante, la mayor parte de los conocimientos se ha obtenido por conducto de proyectos científicos específicos, sin continuidad en el tiempo. Es difícil mantener y actualizar series de datos biológicos, de distribución sobre la frecuencia de los tamaños, etc. sin contar con proyectos de seguimiento a largo plazo.
La evaluación tiene los mismos inconvenientes. Son pocas las evaluaciones oficiales de poblaciones y pesquerías llevadas a cabo en el Mediterráneo, en buena parte porque la mayoría de las poblaciones no se comparten entre países y, por consiguiente, no se ejerce presión alguna para evaluar los recursos y no se ha puesto en práctica ningún sistema de ordenación adaptativa. Con excepción de las especies de grandes pelágicos y de algunas pesquerías muy concretas (como las de chirla en el Adriático) la ordenación pesquera no comporta la utilización de cuotas sino que se basa en la limitación del esfuerzo.
Cada vez se dispone de mayor información sobre algunos aspectos de la dinámica de las poblaciones de las principales especies, y se han reunido parámetros poblacionales en las bases de datos y sistemas de información de la FAO, como POPDYN (Coppola, Garibaldi y Die, 1994) o FIGIS (http://www.fao.org/fi/figis/). Hay numerosos documentos sobre evaluaciones y estimación de parámetros, pero la mayoría de ellos tienen una distribución limitada u oficiosa (Lleonart y Maynou, 2003). En una lista bibliográfica de documentos relacionados con la evaluación de poblaciones de peces del Mediterráneo (en sentido amplio) reunidos recientemente por el Subcomité de Evaluación de Poblaciones del Comité Científico Asesor de la CGPM figuran más de 700títulos (ftp://cucafera.icm.csic.es/pub/scsa/biblio_MED.pdf).
La variedad de especies de la mayor parte de las pesquerías de arrastre, así como la diversidad geográfica del medio ambiente, parecen indicar que en muchos casos las unidades de población pueden ser pequeñas y locales (FAO/GFCM, 1998), pero también que debería contemplarse la posibilidad de adoptar planteamientos multiespecíficos y ecológicos de evaluación y ordenación.
En el Mar Negro se han producido cambios ecológicos drásticos debido a la introducción de una serie de especies exóticas nocivas, tanto de fitoplancton como de animales, a través del agua de lastre de los buques. De la misma manera, un número creciente de especies del Mar Rojo está penetrando en el Mediterráneo oriental a través del Canal de Suez (migrantes “lessepsianos”), dirigiéndose luego hacia el norte y el oeste. En concreto, las especies procedentes del Atlántico Norte se encuentran sobre todo en las áreas septentrionales del Mediterráneo, las especies saharianas y subtropicales fundamentalmente en el sector oriental, mientras que las especies ponto-caspians se ven con mayor frecuencia en el Mar Egeo y el Mar Adriático septentrional (Quignard y Tomasini, 2000). La introducción accidental y difusión de una especie de alga (Caulerpa taxifolia) en el Mediterráneo occidental probablemente también tendrá repercusiones en las cadenas alimentarias de las especies demersales de esa zona en formas no fácilmente previsibles (Zaitsev y Öztürk, 2001; Galil et al. , 2002; Golani et al. , 2002).
El Mediterráneo
En general, los datos acerca de las tendencias del esfuerzo pesquero y de la mortalidad por pesca se basan en informaciones fragmentarias sobre el tamaño de la flota y las capturas por unidad de esfuerzo. En el Mediterráneo occidental (por ejemplo, Baino, Auteri y Serena, 1985; Abella, Caddy y Serena, 1997) probablemente se han aplicado tasas elevadas de explotación a las poblaciones demersales durante los últimos decenios, a juzgar por los informes del Subcomité de Evaluación de Poblaciones de la CGPM. Al mismo tiempo, la comparación de las poblaciones locales (por ejemplo, Abella et al. , 1999) presenta una gran variedad de tasas de explotación, según las flotas y oportunidades de pesca locales. Así pues, la clasificación general de la situación de los recursos demersales del Cuadro D-5, entre plenamente explotados y sobreexplotados no sólo refleja la incertidumbre de la evaluación, sino también las diversas tasas de explotación aplicadas a las poblaciones locales de una especie en todo el Mediterráneo.
En el Golfo de León, un modelo de producción multiespecífico reveló que ya a finales del decenio de 1980 la pesquería estaba totalmente desarrollada, y estudios semejantes ponen de manifiesto que la situación era semejante en las aguas meridionales de Túnez (FAO/GFCM, 1995), aunque en los fondos rocosos septentrionales de este país parece haber cierto margen para una mayor explotación. En el Mar Egeo, y probablemente también en el Mediterráneo oriental y meridional, parece que el esfuerzo y las capturas han aumentado de forma constante al menos desde comienzos del decenio de 1970 (Stergiou et al. , 1997a). En Aldebert et al. (1993) y en informes presentados al Subcomité de Evaluación de Poblaciones de la CGPM en 2001 y 2002, por medio de un análisis de población virtual y de un análisis de rendimiento por recluta en que se supone un reclutamiento constante, se estima la interacción entre tipos de artes (dos arrastres, red de enmalle y palangre) que explotan la población de merluza del Golfo de León, y se llega a la conclusión de que la población está claramentesobreexplotada y de que los arrastreros, que pescan sobre todo ejemplares pequeños, tienen una gran impacto en las capturas de los otros artes (redes de enmalle y palangres). Este análisis confirma los realizados en otras zonas, y que muestran que las flotas de arrastre capturan las clases de edad más jóvenes y, por lo tanto, pueden interferir en la pesca antes de que accesibles a otros artes.
Recursos demersales de la plataforma continental
En la reunión del Comité Científico Asesor, celebrada en 2004, se presentaron 22evaluaciones, relativas a siete subáreas geográficas, nueve especies y dos poblaciones compartidas. El proyecto SAMED de evaluación de poblaciones del Mediterráneo (UE, proyecto № 99/047) basado en las campañas de arrastre MEDITS (1994–1999) (Bertrand et al. , 1998; Bertrand y Relini, 1998) presentó al Subcomité de Evaluación de Poblaciones en 2002 la evaluación de tres especies: Merluccius merluccius , Mullus barbatus y Nephrops norvegicus . Estas evaluaciones se refieren a los países de la UE (con exclusión de las Islas Baleares y del Mar de Alborán) y a Malta .
Merluza europea
Se trata de la especie que más se ha estudiado. En Lleonart y Maynou (2003) se mencionan más de 20referencias sobre evaluaciones de la merluza. Es el principal objetivo de pesca de muchas flotas de arrastre (utilizando mallas de pequeñas dimensiones para los ejemplares jóvenes), de palangre y de redes de enmalle, que faenan en fondos rocosos o mar adentro para la captura de ejemplares de mayor tamaño. Aunque las altas tasas de mortalidad juveniles han llevado a suponer una situación de sobreexplotación, no se ha observado ningún descenso evidente del reclutamiento (Oliver 1993), y de hecho las capturas han aumentado en el Mediterráneo tanto oriental como occidental hasta 1995 (Fiorentini, Caddy y De Leiva, 1997). No obstante, desde 1995 hasta 2002 las capturas registradas han disminuido bruscamente, pasando de 52000 toneladas a 21000 toneladas, y en la actualidad su nivel es parecido al de 1980 (Figura B5.6). Ello apunta a la probable reducción de la población reproductora que sobrevive en algunas zonas menos explotadas (llamadas refugios de reproductores, - Caddy, 1990) de quienes depende el reclutamiento. En cualquier caso, las evaluaciones presentadas al Subcomité de Evaluación de Poblaciones de la CGPM demuestran una clara sobreexplotación de esta especie. Dos evaluaciones sobre la merluza (Golfo de León, y Mar de Liguria y Tirreno septentrional), basadas en métodos analíticos ponían de manifiesto una evidente situación de sobreexplotación y se recomendaba una disminución del esfuerzo. En el caso del Golfo de León se recomendaba también un aumento de la dimensión de las mallas en las capturas de arrastre. Con el proyecto SAMED (basado en los inventarios de arrastre MEDITS) se comprobó una intensa explotación, con tendencias a una disminución en muchas subáreas geográficas. En términos generales, cabe suponer una situación de sobrepesca de crecimiento. Se recomendaron vedas temporales en las zonas de cría y la limitación del esfuerzo.
La determinación de la importancia relativa de una mayor posibilidad de escape de ejemplares juveniles y de reproductores continúa siendo un tema importante para ésta y otras especies demersales del Mediterráneo, en particular si se ha de tener en cuenta la introducción de zonas de cría y de zonas marinas protejidas.
Salmonete de roca (Mullus surmuletus) y salmonete de fango (Mullus barbatus)
Estas especies son capturadas por arrastreros y algunas otras pequeñas embarcaciones de pesca con una gran variedad de artes prácticamente en todo el Mediterráneo. La pesquería comienza con un grupo de edad cero, y en muchas zonas los ejemplares pequeños (capturados en verano y a comienzos del otoño) tienen un precio más elevado. En el Adriático, los supervivientes emigran más tarde hacia el este, donde suelen quedarse como adultos, lo que significa que se trata de una población compartida. Se han registrado tasas de explotación muy elevadas en aguas italianas (FAO/GFCM, 1995; Baino, Auteri y Serena, 1985; Abella et al. , 1999), con excepción de Cerdeña y algunas zonas del Mar Tirreno; la especie está sobreexplotada en el Adriático (Ungaro, Rizzi y Marzano, 1994) y en el Mediterráneo occidental (Martin et al., 1999). Mientras que el salmonete de roca está totalmente explotado en aguas meridionales de Túnez y en el Mar Jónico (Tursi et al. , 1995), al parecer el salmonete de fango está infraexplotado en la plataforma tunecina septentrional (FAO/GFCM, 1995) y sobreexplotado en Chipre (Hadjistephanou, 1992). Los cierres estacionales de las actividades de arrastre en aguas italianas han tenido como objetivo proteger a los juveniles de rápido crecimiento de ésta y otras especies durante los limitados períodos de reclutamiento hasta el fondo. Los resultados han sido un aumento muy positivo de la supervivencia hasta la edad de 1+ años, aunque luego se han registrado descensos estacionales de los precios debido a la saturación del mercado cuando se abría la veda (Froglia, 1989). Al mismo tiempo, la pesca en las zonas de cría situadas a menos de tres millas de la costa ha dado lugar a abundantes capturas de ejemplares jóvenes (Baino, Auteri y Serena, 1985). En las series cronológicas (Figura B5.6) se observa una tendencia ligeramente descendente en los 10últimos años. Las evaluaciones presentadas al Subcomité de Evaluación de Poblaciones se referían principalmente a Mullus barbatus , que es la especie principal. En el Mar de Liguria y en el Tirreno septentrional la especie parece estar plenamente explotada o sobreexplotada. Se recomendaron vedas estacionales y espaciales con el fin de proteger a los ejemplares jóvenes. De acuerdo con el proyecto SAMED, cabe suponer una situación de plena explotación y de sobrepesca de crecimiento en la mayor parte de la subáreas geográficas, y se ha recomendado el cierre de la pesca en aguas poco profundas para proteger el reclutamiento.
Espáridos
Los espáridos desempeñan un papel importante en las pesquerías demersales del Mediterráneo y en general están fuertemente explotados, con tasas máximas de mortalidad en aguas tunecinas que generalmente alcanzan o superan el límite de mortalidad por pesca que da lugar al rendimiento máximo sostenible (FMSY). El análisis de los datos correspondientes a Sparus aurata en el Golfo de León (Farrugio y Le Corre, 1994) indicaba que el nivel de pesca es superior a FMSY, y que una parte elevada de la producción de esta especie procede ahora de la acuicultura. Las brecas (Pagellus spp.), como en el caso de los meros, parecen ser una de las especies demersales menos resistentes a una explotación intensa, y las tasas de captura han disminuido en varias zonas. Se consideran infraexplotadas frente a las zonas rocosas del norte de Túnez (Ben Meriem, Garbi y Ezzedine-Najai, 1996) pero sobreexplotadas en el Golfo de Gabes. Las poblaciones de brecas están fuertemente sobrepescadas en aguas griegas (Papaconstantinou et al. ,1988b), y agotadas en Chipre, lo mismo que la boga y las poblaciones de salmonete de roca (Hadjistephanou, 1992), aunque las chuclas (Spicara spp.) están explotadas en niveles próximos al rendimiento máximo sostenible.
Lenguados
El análisis de los datos sobre el Golfo de León (Farrugio y Le Corre, 1994) indicaba que el nivel de explotación del lenguado está a nivel del rendimiento máximo sostenible (FMSY), pero aunque se encontraron pruebas de que las tasas de explotación habían aumentado a comienzos del decenio de 1990, no había una indicación clara de que el reclutamiento hubiera disminuido. En aguas tunecinas parece que el nivel de explotación del lenguado es casi óptimo.
Cefalópodos
Los cefalópodos figuran como capturas incidentales importantes de las pesquerías de arrastre, y en algunas zonas existe una pesquería específica de pulpo. Una elevada proporción de especies de vida breve (calamares, sepias y pulpos) en las capturas demersales de las zonas intensamente explotadas, como el Adriático (FAO/GFCM, 1995), parece indicar que la explotación es elevada, y que las bajas tasas de captura se compensan en parte con los altos precios por unidad. Los análisis disponibles, aunque fragmentarios (por ejemplo, Baino, Auteri y Serena, 1985 para el Mar Tirreno), indican una explotación casi óptima de estos recursos de cefalópodos, mientras que las especies ícticas están algo sobreexplotadas. No se ha presentado ninguna evaluación de cefalópodos al Subcomité de Evaluación de Poblaciones.
Jureles
Si bien existen pocos datos sobre las especies Trachurus trachurus y T.mediterraneus , aparentemente éstas no son objeto de pesca intensa, y su biomasa es variable, quizás en respuesta a los cambios ambientales.
Lisas (mujílidos)
Se alimentan fundamentalmente de plantas y detritus y al parecer han encontrado condiciones favorables en los estuarios eutróficos. La nueva tendencia hacia la explotación de pequeños peces costeros, como los pejerreyes (aterínidos), demuestra la búsqueda de otras alternativas a los recursos próximos a la costa e intensamente explotados.
Cigala
Se trata de una especie que se ha estudiado bastante bien. En 1998 se publicó un volumen monográfico sobre ella (Sardà, 1998). En él se recogen los conocimientos biológicos sobre la especie, en particular las pesquerías y explotación, distribución, dinámica de población y selectividad. El proyecto SAMED presentó una evaluación al Subcomité de evaluación de poblaciones, en la que se llegaba a la conclusión de que esta especie está ligeramente sobreexplotada o plenamente explotada en la mayor parte de las subzonas geográficas, y se recomendaban mejoras técnicas de los artes para evitar la captura de ejemplares pequeños. Las conocidas características de comportamiento de esta especie (los individuos permanecen largos períodos enterrados y de los que salen periódicamente dando lugar a fluctuaciones estacionales y circadianas en las capturas) hacen que sea menos vulnerable que otras especies a la presión pesquera. No obstante, el carácter discontinuo de la estructura de población y los fenómenos densodependientes podrían estar relacionados con casos de sobreexplotación de poblaciones locales. Debido al período de enterramiento mencionado, se considera importante reducir los efectos de las puertas del arte de arrastre sobre el fondo para proteger esos hábitats.
Recursos del talud continental
El aumento de las pesquerías de talud, especialmente en lo que se refiere a la gamba rosada y la merluza, durante los últimos decenios suscita cierta preocupación. Las pesquerías tradicionales de merluza han utilizado arrastres de malla pequeña en las zonas costeras para la captura de los grupos de edad comprendidos entre cero y dos años. Las pesquerías de merluza con otros artes en aguas más profundas se centran ahora en concentraciones de peces maduros. También el aumento de las pesquerías de altura de peces maduros es motivo de preocupación debido a que la productividad continuada de las poblaciones demersales se debe posiblemente al hecho de que una pequeña proporción de los grupos de edad entre cero y dos años huye de las zonas costeras y encuentra “refugio” para desovar en aguas de altura. Esta hipótesis parece encontrar confirmación en algunos estudios de selectividad, de los que se deduce que los peces de mayor tamaño, como las grandes merluzas, son menos accesibles que los ejemplares menores a la captura con arrastres de malla pequeña.
Gamba rosada (Aristeus antennatus)
Esta especie está intensamente explotada en el Mediterráneo occidental por arrastreros de fondo que la pescan en el talud continental junto con significativas capturas de langostino moruno (Aristaeomorpha foliacea). Es probable que las dos especies no siempre estén debidamente separadas en las estadísticas, lo que permite pensar en un total de capturas de aproximadamente 5000 toneladas al año (Oliver, 1996). Las evaluaciones que aplican análisis de cohortes de tallas y análisis de rendimiento por recluta indican que la actual mortalidad por pesca (F) es muy próxima a la que permite obtener el rendimiento máximo por recluta (FMAX), pero los valores de la mortalidad natural (M) continúan siendo inciertos (Demestre y Lleonart, 1993; Fiorentino et al. , 1998; Ragonese y Bianchini, 1996). El nivel de explotación actual parece ser sostenible pero debería controlarse el aumento constante del esfuerzo de pesca. En las Islas Baleares la aplicación de los análisis mencionados y un modelo de producción de Fox en el supuesto de una situación estable muestra la misma condición de sobreexplotación y la necesidad de reducir el esfuerzo actual (Carbonell y Lauronce, 2000). En cuanto a Grecia e Italia, un volumen considerable de capturas de gamba en aguas profundas se notifica probablemente a la FAO como decápodos Natantia sin especificar. (20000–32000 toneladas), y también es posible que se incluya una cantidad significativa de Aristeus y Aristaeomorpha . De acuerdo con una evaluación presentada al Subcomité de evaluación de poblaciones, la gamba rosada parece estar sobrepescada en el Mediterráneo occidental, y se recomienda una reducción del esfuerzo.
Recursos de pequeñas especies pelágicas
Sardina y boquerón
El seguimiento de las poblaciones de sardina y boquerón desde hace más de un decenio en zonas como el Mar de Alborán y la zona noroccidental, el Adriático, el Egeo y el Mar Negro, se ha realizado con prospecciones acústicas (Abad et al. , 1996; Abad y Giráldez, 2000; Azzali y Luna, 1988; Patti et al. , 2000; Guennegan et al. , 2000). Los métodos basados en la producción diaria de huevos (DEPM) se han utilizado varias veces en el Mediterráneo pero no suelen emplearse de forma sistemática año tras año. Estos métodos se han aplicado sobre todo al boquerón, pero también a la sardina en algunos casos (Chavance, 1980; Regner, 1990; Palomera y Pertierra, 1993; García y Palomera, 1996; Somarakis y Tsimenides, 1997; Casavola et al. , 1998; Casavola, 1999; Quintanilla et al. , 2000). La comparación entre los resultados de los DEPM, prospecciones y modelos de dinámica de población mostraron que los resultados eran muy semejantes (Pertierra y Lleonart, 1996).
Estos análisis presentan una pauta de fluctuaciones que no está relacionada de forma evidente con la intensidad de la pesca. En el Adriático, las capturas de boquerón alcanzaron un máximo en 1980 y disminuyeron posteriormente, mientras que las poblaciones de sardinas locales alcanzaron el máximo a mediados del decenio de 1980. La biomasa de las poblaciones de boquerón pasó de 18000 toneladas en 1989 a unas 340000 toneladas en 1991, cifra que es, no obstante, más baja que las de los decenios de 1970 y 1980 (Arneri, 1994). En Cingolani et al. (2004a, 2004b) se analizaron la dinámica del boquerón y de la sardina en el Mar Adriático utilizando un análisis de población virtual (serie cronológica de 1975 a 2003), en el que se observaba una disminución constante de la población de sardinas. En el Mar de Alborán las capturas alcanzaron un máximo en 1982, con 27878 toneladas, para descender posteriormente a 1016 toneladas en 1988 (Abad y Giráldez, 1990). En el Golfo de León y en la parte septentrional de España las capturas aumentaron en 1989, probablemente debido a que las flotas se trasladaron de la parte meridional de España, donde las capturas de boquerón habían disminuido.
En el Mar Egeo, como ocurrió con la mayoría de los recursos, las tasas medias de captura de pequeñas especies pelágicas disminuyeron hasta mediados del decenio de 1970 antes de estabilizarse, mientras que el total de capturas, especialmente en el Mediterráneo oriental, ha continuado aumentando, lo que permite pensar en un crecimiento de la productividad en aguas que anteriormente tenían un contenido de nutrientes muy bajo. Esta explicación adquiere mayor credibilidad si se tiene en cuenta el aumento paralelo de la abundancia de fitoplancton y zooplancton, así como las estimaciones acústicas de la biomasa de pequeñas especies pelágicas a lo largo de una línea que va desde el Egeo septentrional hasta el Mediterráneo sudoriental (Stergiou et al. ,1997b). Las biomasas pelágicas son también generalmente bajas en aguas libias (FAO/GFCM, 1995), excepto por lo que se refiere a una corriente estacional local ascendente en el Golfo de Sirte. Se observó también un fuerte descenso de las capturas de especies pelágicas en la región de Levante después de la construcción de la presa de Asuán (Halim et al. , 1995). Se estima que en aguas de Túnez se explota cada año el 40por ciento de las poblaciones de sardinas, quizás por encima del nivel óptimo, y las biomasas de sardina y boquerón tienden a fluctuar de forma no sincronizada.
En la reunión del Subcomité de Evaluación de Poblaciones de 2004 se presentaron evaluaciones sobre el boquerón y la sardina en el Mediterráneo occidental, Mar de Alborán y Mar Adriático en las que no se observaba ninguna tendencia definida, excepto en el caso de la sardina del Adriático, que presentaba una disminución de la biomasa y un aumento de la mortalidad por pesca durante los últimos 15 años. Las conclusiones generales fueron las siguientes: la abundancia depende en gran medida del reclutamiento, y la biomasa de las poblaciones acusa una elevada variabilidad interanual; cuando se dectecta el riesgo de sobrepesca de reclutamiento, es aconsejable evitar la captura de peces que no han alcanzado todavía la talla de primera madurez; las estimaciones sobre la biomasa deberían darse con suficiente antelación para permitir que los administradores puedan adaptar la explotación en tiempo real.
Sardinelas y pejerreyes
Se observan fluctuaciones semejantes a largo plazo en todos los lugares del Mediterráneo donde se capturan pequeñas especies pelágicas, lo que hace suponer que quizás estén produciéndose fluctuaciones periódicas a largo plazo en las condiciones ambientales, que se superpondrían a los efectos antropogénicos. La recuperación de la producción pelágica en las aguas situadas frente al delta del Nilo después del mencionado descenso parece referirse en buena medida a la alacha (Sardinella aurita), especie de pelágico pequeño predominante en el Mediterráneo sudoriental, cuyas capturas presentan una clara tendencia ascendente. Este aumento continuo de la alacha (tropical) y el aparente descenso del espadín (boreal) puede ser indicativo de un proceso de calentamiento global. En otras partes del Mediterráneo oriental se han desarrollado nuevas pesquerías costeras para especies pelágicas muy pequeñas, como los pejerreyes.
Caballas, jureles y chicharros
Las caballas presentan una gran variabilidad en las capturas. No se ha llevado a cabo ninguna evaluación específica sobre estas especies, y aunque las capturas de caballa en aguas griegas han aumentado hay también pruebas indirectas de que en los últimos años han dejado de penetrar en gran número en el Mar Negro.
Recursos de grandes especies pelágicas
En la sección C1 se facilita información de alcance mundial sobre las grandes especies pelágicas. No obstante, cabe formular algunas observaciones específicas sobre el Área37:
Atún rojo y pez espada
El atún rojo (Thunnus thynnus) y el pez espada (Xyphias gladius) son las especies de grandes pelágicos más importantes del Mediterráneo. Representan aproximadamente el tres por ciento del total de las capturas registradas, pero su importancia económica es mucho mayor. La Comisión Internacional para la Conservación del Atún del Atlántico (CICAA) considera una sola población de atún rojo en el Atlántico oriental y el Mediterráneo, correspondiendo a éste último la principal zona de desove. Las redes de deriva, los palangres y las redes de cerco con que se explota el atún rojo representan la única flota industrial real en el Mediterráneo. Después de alcanzar desembarques de 39000 toneladas en 1994, el volumen descendió a 22000 toneladas en 2002. Se ha manifestado una justa preocupación por la situación de ambas especies. En los dos casos el crecimiento (aparentemente incontrolado) de las pesquerías durante el último decenio ha agravado la vulnerabilidad de estas poblaciones, como lo ha hecho la pesca habitual de los Estados no ribereños con respecto a las poblaciones de atún rojo. En el caso de esta especie, la CGPM ha adoptado los reglamentos sobre las pesquerías del atún rojo de la CICAA, por lo que se deben aplicar a todos los países miembros de la CGPM. Recientemente ha habido un aumento sustancial del número de jaulas costeras para el engorde o engrase de atún rojo procedente de la pesca,, con el fin de aumentar la calidad e independizar el mercado de la temporada de capturas. Se cree que esta situación también ha dado lugar a un aumento de la presión pesquera, debido a que se han abierto nuevos mercados y porque es posible que no se notifiquen todas las capturas. Estas actividades influyen en muchos aspectos del proceso de evaluación-ordenación, como la recopilación de datos y, por consiguiente, en la evaluación de poblaciones. Tienen efectos reales (en las estadísticas, la biología y la ordenación) y potenciales (en el medio ambiente, en la situación socioeconómica y en la ordenación) (Anón., 2002; Oray y Karakulak, 2003). La preocupación causada por el rápido desarrollo de esta actividad, basada en las pesquerías de captura, ha dado lugar a la organización de un grupo de trabajo intersectorial sobre prácticas sostenibles de cría y engorde del atún en el Mediterráneo para ofrecer orientaciones comunes que permitan hacer de esta actividad una práctica sostenible.
El pez espada es la segunda gran especie pelágica por orden de importancia. La CICAA considera la existencia de una única población mediterránea. El pez espada se captura con palangres y redes de deriva. En el caso del pez espada, la elevada tasa de explotación se refleja también en el descenso progresivo de la media del tamaño y la edad en el momento de la captura.
Bonito y atún blanco
Las pesquerías de estas especies no se han evaluado específicamente, pero hay pruebas indirectas de que el bonito (Sarda sarda) ha reducido su zona de distribución, y que ahora son muy pocas las ocasiones en que llega al Mar Negro en gran número. El bonito no presenta ninguna tendencia a largo plazo, y sus capturas se caracterizan por una gran variabilidad. Las capturas de atún blanco (Thunnus alalunga) han aumentado desde 1980, con fluctuaciones.
Mar de Azov y Mar Negro
La ligera mejoría de las condiciones ambientales registrada en el pasado decenio en el Mar Negro y el Mar de Azov crea un medio favorable para la recuperación de la biodiversidad y de los recursos marinos vivos. A pesar de esta mejoría, en las pesquerías demersales persisten todavía situaciones de sobrepesca, degradación de hábitats vitales, incluidas las zonas de puesta y de cría, y perturbación del balance estructural en la biota.
A diferencia de lo que ha ocurrido en el resto del Mediterráneo, en el Mar de Azov y el Mar Negro todos los países ribereños han declarado ZEE. El marco legislativo y regulador establecido permite la ordenación de los recursos vivos en el plano nacional, pero hace falta una mayor armonización.
De acuerdo con las conclusiones de un reciente taller sobre la pesca responsable y la conservación del ecosistema del Mar Negro (http://www.blacksea–environment.org/text/default.htm), las actividades de evaluación y seguimiento de las poblaciones son fragmentarias e irregulares mientras que la pesca ilegal continúa siendo una de las cuestiones pendientes más importantes para todos los Estados ribereños del mar Negro.
En el Mar de Azov en el decenio de 1960 se produjeron varios episodios de colapso de las poblaciones, por ejemplo, de lucio (Esox lucius), brema (Abramis brama) y rutilo (Rutilus rutilus) (Ivanov y Beverton, 1985). Estas caídas estaban asociadas con la salinización progresiva debida a la construcción de presas y la extracción de agua en los principales afluentes, lo que ha aumentado la salinidad en varias unidades. Ello, más los efectos en las rutas de desove y zonas de cría en los principales ríos, ha afectado gravemente a las poblaciones naturales de esturiones, cuyas capturas ahora se mantienen en parte mediante la producción en criadero. Se registraron descensos en las poblaciones de peces demersales, en particular el rodaballo (Psetta maxima), los crustáceos bentónicos y zoobentos y fitobentos. Éstos últimos desempeñaban antes un papel importante, por ejemplo, los mejillones (filtración) y las algas rojas (oxigenación del agua de la plataforma en el Mar Negro noroccidental). Su desaparición se debió en gran parte a acontecimientos anóxicos causados por una producción de plancton mucho mayor que, a su vez, estaba asociada a una mayor aportación de nutrientes de la gran cuenca de captación del Mar Negro. Las fuertes caídas de las poblaciones de espadín del Mar de Azov, y de boquerón en el Mar Negro, y la disminución del chicharro del Mediterráneo y de las poblaciones de espadín al final del decenio de 1980, casi ciertamente estuvieron vinculadas con la introducción en el Mar Negro y el Mar de Azov del ctenóforo Mnemiopsis leidyi del Atlántico occidental (Zaitsev, 1993). Mientras que una especie de lisa (Mugil soiuy), introducida desde la costa del Pacífico de Rusia, que se alimenta de detritus y crece en aguas poco profundas, parece inmune a la depredación de M.leidyi en su fase de larva, y se ha producido un aumento de la población. Varias otras especies introducidas parecen estar preadaptadas a condiciones eutróficas, como la almeja del género Mya y el caracol marino del género Rapana ; éste último representa ahora un importante producto de exportación en algunos países, y puede considerarse como plenamente explotado.
La recuperación parcial de la población de boquerón, que era anteriormente objeto de pesca directa en el Mar Negro septentrional pero que ahora se pesca sobre todo en el Mar Negro meridional, es quizás el acontecimiento más importante registrado en la pesca en esta zona. Recientemente se ha visto beneficiada por la aparición de una especie de ctenóforo (Beroe cucumis, o B.ovata, según las fuentes), depredador de M. leidyi - al que se debía en gran parte la caída de la población de boquerón (Zaitsev y Öztürk, 2001). A pesar de su colapso y parcial recuperación, sería un error considerar que la población de boquerón está infraexplotada. Con excepción de las flotas de los antiguos países socialistas, en proceso de reconstrucción y con escasez de fondos para reparaciones e inversiones, continúa todavía vigente en el Mar Negro meridional el exceso de capitalización de la industria de la pesca que se mencionó en la revisión anterior. Por el contrario, es posible que en la actualidad las poblaciones de espadín y de varias otras especies del Mar Negro septentrional no estén sobreexplotadas. Cuando las condiciones ambientales lo permitan, debería favorecerse la recuperación de los restantes recursos demersales (Prodanov et al. , 1997).
Los principales predadores del Mar Negro, como los delfines, son ahora mucho menos abundantes. Los peces predadores, incluida la caballa (Scomber scombrus), la anjova (Pomatomus saltatrix) y el bonito, que solían entrar estacionalmente desde el Mar de Mármara (sometido también a una fuerte contaminación e intensa pesca), ahora penetran raramente en aguas situadas al norte y oeste del Mar Negro. Las poblaciones de estas especies pueden considerarse agotadas, aunque no sólo necesariamente por efecto de la pesca.
Cinco especies pueden servir como indicadores útiles de los últimos cambios drásticos de los ecosistemas pesqueros del Mar Negro (Prodanov et al. , 1997; Daskalov, 1998 y 1999).
Sábalos
La biomasa de estas especies anádromas (como las del esturión, más importante) ha disminuido en un 75por ciento o más con respecto al decenio de 1970. Las tasas de explotación han sido altas, pero no han mostrado una tendencia ascendente continuada y es posible que el deterioro, de los últimos decenios, de las condiciones de los ríos que desembocan en el Mar Negro haya contribuido en buena medida al agotamiento de las poblaciones.
Espadín del Mar de Azov
La población de espadín, con una biomasa muy variable, presenta amplias fluctuaciones pero fue objeto de una pesca intensiva por parte de la antigua Unión Soviética durante los decenios de 1950 y 1970. Los aumentos más recientes de la tasa de explotación están asociados con descensos de las capturas, vinculados a la explosión del ctenóforo depredador Mnemiopsis leydei , que ha ocupado un lugar predominante en el ecosistema pelágico desde el final del decenio de 1980.
Boquerón
Esta especie constituye el recurso más importante del Mar Negro. Su biomasa aumentó al final del decenio de 1970 y comienzos del decenio de 1980, en un momento en que las capturas también estaban aumentando, al parecer como consecuencia de las entradas de nutrientes en el Mar Negro. Sufrió una fuerte caída al final del decenio de 1980, debido en gran parte a la depredación y a la competencia alimentaria con el ctenóforo Mnemiopsis leydei . Posteriormente hubo una disminución del esfuerzo de pesca, que hizo posible que la especie se recuperara algo, pero de todas formas la biomasa y las capturas de boquerón no han vuelto a sus valores anteriores.
Plegonero
Esta especie, principal depredador del espadín del Mar Negro, se ha visto afectada por el descenso de las poblaciones de espadín debido a la plaga de Mnemiopsis . El plegonero es fundamentalmente una captura incidental de la pesca del espadín, cuya población está intensamente explotada, lo que repercute en la disminución de la población de este depredador (Prodanov et al. , 1997).
Otras especies demersales
De acuerdo con las conclusiones del taller sobre recursos demersales en el Mar Negro y el Mar de Azov (Öztürk y Karakulak [Eds.], 2003) las pesquerías demersales continúan representando una importante actividad económica con efectos significativos en la población costera y en el ecosistema marino. Aunque hay problemas de sobrepesca y de degradación de hábitats de gran importancia, los recursos demersales todavía tienen posibilidades de recuperación. Las especies más importantes, además del plegonero, son las siguientes: rodaballo del Mar Negro (Psetta maxima maeotica), mielga (Squalus acanthias), Raya de clavos (Raja clavata) y lisas (mujílidos). Las capturas de la mayoría de ellas han disminuido en los últimos años.
Esturiones
Con relación a las especies más comunes - Acipenser gueldenstaedtii , Acipenser stellatus y Huso huso - las cifras revelan una tendencia descendente durante los últimos años (con oscilaciones y evoluciones diversas, por especies y países). Acipenser sturio y Acipenser nudiventris prácticamente han desaparecido. A.sturio y A.brevirostrum figuran en el ApéndiceI de la CITES (especies amenazadas de extinción), y el resto de las especies en el ApéndiceII (especies que pueden estar en peligro de extinción a menos que se controle estrictamente su comercio). A.guendelstaedtii , A.nudiventris, A.persicus, A.stellatus, A.sturio y Huso huso están incluidas en la Lista roja de animales en peligro de la Unión Internacional para la Conservación de la Naturaleza y de los Recursos Naturales (IUCN).
Evaluación de poblaciones
El Comité Científico Asesor de la Comisión General de Pesca del Mediterráneo (CGPM) analizó, en mayo de 2000, la información científica disponible sobre 1985–1999 acerca de las ocho especies demersales y pequeñas especies pelágicas compartidas más importantes: merluza, salmonetes, bacaladilla, gamba rosada, boquerón, sardina y alacha (Fiorentino, 2000). Se determinaron y analizaron más de 100evaluaciones (FAO/GFCM, 2000a). Se tuvieron también en cuenta tres grandes especies pelágicas (atún rojo, pez espada y atún blanco) (CICAA, 1999).
CuadroB5.1 Evaluaciones presentadas en la reunión del Comité Científico Asesor (2001–2004
| Especies | Evaluaciones | Observaciones y recomendaciones | |||
| 2001 | 2002 | 2003 | 2004 | ||
| Merluccius merluccius | 4 | 2 | 3 | 3 | O, sobrepesca de crecimiento, riesgo de sobrepesca de reclutamiento.Reducir el esfuerzo. Mejorar la selectividad de los arrastres. Cierres estacionales de las zonas de cría. |
| Mullus barbatus | 3 | 1 | 2 | 4 | O - F, Cierre de zonas costeras para p roteger los ejemplares jóvenes. Uso de arrecifes artificiales Cierre estacional. Reducción del esfuerzo. Aplicación de las normas vigentes de ordenación. |
| Mullus surmuletus | 1 | ||||
| Pagellus erythrinus | 3 | F | |||
| Diplodus annularis | 3 | F | |||
| Pomatomus saltatrix | 1 | ||||
| Nephrops norvegicus | 1 | 1 | F, Mejora técnica de los artes para evitar la captura de ejemplares pequeños. Reducir los efectos de las puertas de los arrastres en el fondo. | ||
| Aristeus antennatus | 2 | 3 | 3 | 3 | O - F |
| Aristaeomorpha foliaciea | 1 | 1 | F | ||
| Parapenaeus longirostris | 1 | U - F | |||
| Sardina pilchardus | 3 | 5 | 4 | 5 | O - F, no aumentar el esfuerzo. |
| Engraulis encrasicolus | 3 | 6 | 3 | 3 | Riesgo de sobrepesca de reclutamiento. Fijar tamaño mínimo legal de acuerdo con el de la primera madurez. |
| Trachurus trachurus | 1 | 1 | F, Mantener el esfuerzo de pesca en los niveles actuales. | ||
| Boops boops | 1 | F, Mantener el esfuerzo de pesca en losniveles actuales. | |||
| Total | 24 | 19 | 17 | 22 | |
O- Sobreexplotada;
F- Totalmente explotada;
U- Insuficientemente explotada.
La mayor parte de las evaluaciones corresponden a poblaciones de la parte septentrional y occidental de la región. Las evaluaciones estaban basadas en la aplicación de análisis de población virtual o de cohortes en función de la talla, junto con análisis de rendimiento por recluta y, algunas veces, métodos de producción excedente, basados en series breves de datos y en los resultados de campañas científicas. Desafortunadamente, en la reunión no se pudo disponer de ciertas informaciones existentes que no estaban publicadas.
Las evaluaciones presentadas al Comité Científico Asesor entre 2001 y 2004 (con inclusión de SAMED) se resumen en el CuadroB5-1 (no se presentaron evaluaciones sobre las grandes especies pelágicas).
Se necesita con urgencia un esfuerzo considerable en el Mediterráneo para mejorar el asesoramiento científico así como su aplicación por los organismos de ordenación. La inversión actual en evaluación de poblaciones y, en términos más generales, en investigación pesquera para fines de evaluación no es suficiente ni adecuada para poder asesorar en forma pertinente y eficiente a los responsables de la ordenación de la pesca y al sector privado. Con el fin de alentar los esfuerzos para ofrecer el asesoramiento científico necesario, el Comité Científico Asesor recomendó en 2000, entre otras cosas, que se actualizara y mejorara la calidad y cobertura de los datos y de las estadísticas sobre pesca, que se aumentara el número de evaluaciones en las zonas meridional y oriental y que se hiciera lo posible para que todas las evaluaciones se llevaran a cabo de forma periódica. El Comité llamó también la atención sobre la importancia de la relación existente entre medio ambiente y recursos. Además, insistió en la gran importancia de definir las unidades de ordenación geográfica para notificar los indicadores y en la necesidad de elaborar indicadores socioeconómicos homogéneos en cada una de las unidades de ordenación.
Ordenación pesquera
La plataforma y el talud continental, sobre todo en el Mediterráneo septentrional, se explotan hasta una profundidad de 800m con una gran variedad de artes para distintos peces e invertebrados demersales, entre los que predominan la merluza, la bacaladilla, el salmonete de fango, el plegonero, los cefalópodos, la cigala y las gambas. La combinación varía de acuerdo con la situación geográfica, y la mayoría de las capturas están integradas por peces de edades comprendidas entre 0+ y 2+ años (FAO/GFCM, 1995). En lo que respecta al tamaño en el momento de la primera captura, continúa utilizándose una luz de malla estirada en el copo de 40mm o menos. Ello significa que la pesca de arrastre de especies de mayor tamaño, como la merluza, es sobre todo una pesca de ejemplares jóvenes, a pesar de que, como han observado Dremière (1979) y Bertrand (1990), el tamaño óptimo de la primera captura de especies más pequeñas económicamente importantes en la pesca multiespecífica (gambas y pequeños cefalópodos) ha representado un obstáculo para aumentar la luz de malla. Además, no se ha evaluado la supervivencia de ejemplares que escapan a través de las mallas. En Chipre, las chuclas representan la mayoría de las capturas que, al parecer, se verían gravemente afectadas si se aumentara la luz de malla (Hadjistephanou, 1992). En algunas zonas se han decretado cierres estacionales durante el período de reclutamiento al fondo, y en algunos países existen reglamentos que exigen el cierre permanente de la pesca de arrastre en aguas poco profundas y parques marinos.
Es difícil evaluar las tendencias del esfuerzo de pesca en el Mediterráneo, especialmente en el caso de la pesca de arrastre, pero se han adquirido nuevas flotas, por ejemplo, en Croacia y Libia, y se han puesto en práctica planes de sustitución de las flotas en otras zonas, lo que indica que el esfuerzo real quizás haya aumentado. En términos generales, en la mayoría de los países hay pocas medidas de ordenación que limiten el esfuerzo de pesca o el total de las capturas. Al mismo tiempo, son sorprendentemente pocos los indicadores de colapso de las poblaciones, siendo más las especies que revelan un aumento que una disminución de las capturas (Fiorentini et al. , 1997). En los últimos años, con financiamiento de la Comisión Europea, se han realizado en aguas de sus países miembros campañas de prospección de arrastre para determinar la situación de los recursos demersales, y este conjunto de datos está comenzando a permitir una cierta evaluación de poblaciones (por ejemplo, Papaconstantinou et al. , 1988a, b; Farrugio, 1994; Oliver, 1996).
En Abella et al. (1999) se atribuye la capacidad de recuperación de la merluza del Mediterráneo, a pesar de la intensa pesca de juveniles, al resultado neto de la distribución espacial desigual del esfuerzo de la pesca de arrastre, en particular la de ejemplares maduros fuera de las principales zonas de pesca de arrastre y la vulnerabilidad relativamente baja de los grandes ejemplares a los arrastres de malla pequeña (Caddy, 1990). Estos efectos no se habían previsto en el pasado pero deben tenerse en cuenta al planificar medidas futuras de ordenación. Debería evaluarse, especialmente, el efecto de los posibles aumentos del esfuerzo de pesca en aguas profundas sobre el reclutamiento de la merluza.
La CGPM, en su 25° período de sesiones de septiembre de 2000, después de examinar el asesoramiento ofrecido por el Comité Científico Asesor, recomendó, entre otras cosas, “la elaboración y aplicación de las medidas de ordenación necesarias para corregir estos problemas de sobrepesca” (FAO/GFCM 2000b).
La CGPM recomendaba también “que se hiciera lo posible para aumentar el número de evaluaciones llevadas a cabo en las zonas meridional y oriental” y reconocía “que era fundamental la participación activa de los países miembros para proporcionar a los grupos de trabajo toda la información que tuvieran a disposición”.
REFERENCIAS
Abad, R. y Giráldez, A. 1990. Descripción de la pesca de cerco en la región surmediterránea. Informes Técnicos del Instituto Español de Oceanografía. No. 86. 48pp.
Abad, R., Miquel, J. y Iglesias, M. 1996. Campañas de evaluación por métodos acústicos de sardina, boquerón y ochavo en el Mediterráneo Occidental. FAO Fisheries Report, No. 537: 191–193.
Abad, R. y Giraldez, A. 2000. Acoustic estimation of aboundance of sardine, anchovy and sardinelle in the NW Mediterranean. FAO/GFCM - SAC Stock Assessment Working Group on small pelagics, Fuengirola (Spain) 1–3 March 2000. Mimeo.
Abella, A.J., Caddy, J.F. y Serena, F. 1997. Do natural mortality and availability decline with age? An alternative yield paradigm for juvenile fisheries, illustrated by the hake Merluccius merluccius fishery in the Mediterranean. Aquat. Living Resour. , 10: 257–69.
Abella, A.J., Belluscio, A., Bertrand, J., Carbonara, P.L., Giordano, D., Sbrana, M. y Zamboni, A. 1999. Use of MEDITS trawl survey data and commercial fleet information for the assessment of some Mediterranean demersal resources. Aquat. Living Resour . 12 (3): 155–166.
Aldebert, Y., Recasens, L. y Lleonart, J. 1993. Analysis of gear interactions in a hake fishery: The case of the Gulf of Lions (NW Mediterranean). Sci. Mar. , 57(2–3):207–217.
Anon. 2001. Towards holistic fisheries management: a Mediterranean perspective . Report of a workshop held in Heraklion, Crete, March 2001, under the auspices of the European Union. Accompanying Measures Programme, Contract Q5AM-2000-00002. 22 pp.
Anon. 2002. Report of the sixth GFCM-ICCAT meeting on stocks of large pelagic fishes in the Mediterranean, Sliema, Malta, April 2002. 36 pp.
Arneri, E. 1994. Fisheries resources assessment and management in the Adriatic and Ionian Seas: GFCM Document TC/CM/III/94 14 pp. Mimeo.
Azzali, M. y Luna, M. 1988. A preliminary note on the pelagic resources in Italian Seas, assessed through acoustic techniques.Seminari delle unita operative responsabili dei progetti di ricerca promossi nell'ambito dello schema preliminare di piano per la pesca e l'acquacoltura.Atti del Ministero della Marina Mercantile, Vol 1, Roma. pp. 101–171.
Baino, R. Auteri, R. y Serena, F.1985.Groundfish stocks in the Northern Tyrrhenian Sea management with Gulland-Fox production model. FAO GFCM Report, 336, Annex (E) R. pp. 125–130.
Bas, C. 2002.El Mar Mediterráneo: Recursos Vivos y Explotación . Ariel Ciencia, Barcelona. 518pp.
Ben Meriem, S., Gharbi, H. y Ezzeddine-Najai, S. 1996.Le rouget de roche (Mullus surmuletus) en Tunisie: Evaluation des stocks et aménagement des pêcheries.FAO Fisheries Report, No. 533 (Suppl.): 269–284.
Bertrand, J. 1990. Typologie des ressources halieutiques du golfe du Lion et application au problème du maillage des chaluts de fond.FAO Rapports sur les pêches No. 447: 151–159.
Bertrand, J., Gil de Sola, L., Papaconstantinou, C., Relini, G. y Souplet, A. 1998.An international bottom trawl survey in the Mediterranean: the MEDITS programme. In Demersal Resources in the Mediterranean. (Coordinated by J. A. Bertrand y G. Relini). Actes de Colloques IFREMER , 26: 76–93.
Bertrand, J. y Relini, G. (Coordinators) 1998. Demersal Resources in the Mediterranean. Actes de Colloques IFREMER, 26.
Caddy, J.F. 1990. Options for the regulation of Mediterranean demersal fisheries. Natural Resource Modeling , 4: 427–475.
Caddy, J.F. y Rodhouse, P.G. 1998. Cephalopod and groundfish catches: evidence for ecological change in global fisheries? Rev. Fish Biol. Fisheries, 8: 431–444.
Caddy, J.F., Refk, R. y Do Chi, T. 1995. Productivity estimates for the Mediterranean: evidence of accelerating ecological change. Ocean and Coastal Management , 26(1): 1–18.
Carbonell, A. y Lauronce, V. 2000. Application of VPA model and Fox production model to short time data series of the red shrimp (Aristeus antennatus) of SW Majorca waters (Balearic Sea, western Mediterranean). GFCM - SAC Stock Assessment Working Group on demersals, Sète (France) 21– 23 March 2000. 15pp. Mimeo.
Carbonell, A., Martín, P., De Ranieri, S. y WEDIS team. 1998. Discards of the Western Mediterranean trawl fleets. Rapp. Comm. int. Mer Médit. , 35: 392–393.
Carbonell, A., Alemany, F., Merella, P., Quetglas, A. y Román, E. 2003.The by-catch of sharks in the western Mediterranean (Balearic Islands) trawl fishery. Fisheries Research, 61: 7–18.
Casavola, N. 1999. Valutazione della biomassa di alici mediante la stima della produzione giornaliera di uova lungo le coste adriatiche pugliesi nel 1995.Biol. Mar. Medit ., 6 (1): 553–555.
Casavola, N., De Ruggieri, P., Rizzi, E. y LoCaputo, S. 1998. Daily egg production method for spawning biomass estimates of Sardine in the South-Western Adriatic Sea. Rapp. Comm. int. Mer Médit ., 35 (2): 396.
Coppola, S.R., Garibaldi, L. y Die, D. 1994.Popdyn: population dynamic database. user's manual. FAO Computerized Information Series (Fisheries). No. 7. Rome, FAO: 30 pp.
Chavance, P. 1980.Production des aires de ponte, survie larvaire et biomasse adulte de la sardine et de l'anchois dans l'est du golfe du Lion (Méditerranée occidentale).Tethys , 9(4): 399–413.
Cingolani, N., Santojanni, A., Arneri, E., Berlardinelli, A., Colella, S., Donato, F., Giannetti, G., Sinovčić, G. y Zorica, B. 2004a.Sardine (Sardina pilchardus , Walb.) stock assessment in the Adriatic Sea: 1975–2003. AdriaMed Occasional Papers, 13:09 pp.
Cingolani, N., Santojanni, A., Arneri, E., Berlardinelli, A., Colella, S., Donato, F., Giannetti, G., Sinovčić, G. y Zorica, B. 2004b.Anchovy (Engraulis encrasicolus , L.) stock assessment in the Adriatic Sea: 1975–2003. AdriaMed Occasional Papers, 14:10 pp.
Daskalov, G. 1998.Pêcheries et changement environnemental à long terme en Mer Noire . Thèse de doctorat, Université de la Méditerranée (Aix-Marseille II).
Daskalov, G. 1999. Relating fish recruitment to stock biomass and physical environment in the Black Sea using generalized additive models. Fisheries Research . 41, pp.1–23.
Demestre, M. y Lleonart, J. 1993. Population dynamics of Aristeus antennatus (Decapoda: Dendrobranchiata) in the northwestern Mediterranean. Sci. Mar . 57 (2–3): 183–189.
Dremière, P.Y. 1979. Paramètres biologiques et dynamiques disponibles sur les principaux stocks halieutiques du golfe du Lion: sous-zone 37-2 du CGPM. FAO Rapport sur les pêches No. 227: 115–127.
Estrada, M. 1996. Primary production in the northwestern Mediterranean. Sci. Mar. 60 (supl. 2): 55–64.
FAO/GFCM. 1995. Report of the third technical consultation on stock assessment in the central Mediterranean. Tunis, 8–12 November 1994. 19 pp.
FAO/GFCM. 1998. Issues in Mediterranean fisheries management: geographic units and effort control: GFCM Studies and Reviews, No. 70, 56 pp.
FAO/GFCM . 2000a. Report of the third session of the Scientific Advisory Committee. Madrid, Spain, 2–5 May 2000.Rapport de la troisième session du Comité scientifique consultatif. Madrid, Espagne, 2–5 mai 2000.FAO Fisheries Report/FAO Rapport sur les pêches .No. 632. Rome, FAO. 2001. 40p.
FAO/GFCM. 2000b. Report of the twenty-fifth session. Sliema, Malta, 12–15 September 2000.GFCM Report .No. 25. Rome, FAO. 27 p .
Farrugio, H. y Le Corre, G . 1994. Diagnostics à l'équilibre de quelques stocks halieutiques démersaux euryhalins du Golfe de Lion: Analyses de pseudocohortes et rendements par recrue.GFCM Consultation Sète, 19–23 Septembre 1994. Mimeo.
Farrugio, H. 1994. FARWEST: An international research programme (1990–93) for the north-western Mediterranean demersal fisheries. 8th technical consultation of GFCM for the Baleares and Gulf of Lions Division. 15 pp. Mimeo.
Farrugio, H. 1996. Mediterranean fisheries status and management. Evolution of the research and improvement of regional cooperation. Diplomatic Conference on Fisheries Management in the Mediterranean, Venice, November 1996. 18 pp.
Farrugio, H., Oliver, P. y Biagi, F. 1993. An overview of the history, knowledge, recent and future research trends in Mediterranean fisheries. In J. Lleonart (Ed) Northwestern Mediterranean Fisheries . Sci. Mar. , 57: 105–119.
Fiorentini, L., Caddy, J.F. y DeLeiva, J.I. 1997. Long and short term trends of Mediterranan fishery resources. FAO GFCM Studies and Reviews, No. 69, 72 pp.
Fiorentino, F., Orsi Relini, L., Zamboni, A. y Relini, G. 1998. Remarks about the optimal harvest strategy for red shrimps (Aristeus antennatus , Risso 1816) on the basis of the Ligurian experience. Cah. Options Méditerr . 35: 323–333.
Fiorentino, F . 2000. A compilation of information on stock assessment in the GFCM areas presented in standard forms Report to the SCSA, 109 pp. (ftp://cucafera.icm.csic.es/pub/ scsa/Subcommittee_2000/).
Froglia, C. 1989. Clam fisheries with hydraulic dredges in the Adriatic Sea. In Caddy, J.F. (Ed) Marine Invertebrate Fisheries: Their Assessment and Management . pp. 507–524.
Galil, B., Froglia, C. y Noël, P. 2002. CIESM Atlas of Exotic Species - Vol. 2. Crustaceans. Frederic Briand, editor. CIESM Publishers, 192pp., 35 original color drawings.
Garcia, A. y Palomera, I. 1996. Anchovy early life history an its relation to its surrounding environment in the Western Mediterranean basin. In I. Palomera y P. Rubiés, Eds., The European Anchovy and its Environment. Scient. Mar. , 60 (supl.2): 155–166.
Garibaldi, L. y Caddy, J.F. 1998. Biogeographic characterization of Mediterranean and Black Sea faunal provinces using GIS procedures. Ocean and Coastal Management , 39: 211–227.
Golani, D., Orsi-Relini, L., Massutí, E. y Quignard, J.-P. 2002.CIESM Atlas of Exotic Species in the Mediterranean - Vol. 1. Fishes Frederic Briand, editor. CIESM Publishers, 256p.
Guennegan, Y., Liorzou, B. y Bigot, J.L. 2000. Exploitation des petits pelagiques dans le Golf du Lion et suivi de l'évolution des stocks par echo-integration de 1999 a 2000 .Paper presented at WG on small Pelagics. Fuengirola (Spain) 1–3March 2000. 27p.
Hadjistephanou, N.A. 1992. Assessment of five demersal marine fish stocks of Cyprus for the year 1989. FAO Fisheries Report, No.477: 23–34.
Halim, Y., Morcos, S.A., Rizkalla, S. y El-Sayed, M. Kh. 1995. The impact of the Nile and the Suez Canal on the living marine resources of the Egyptian Mediterranean waters (1958–1986). FAO Fisheries Technical Paper, No. 349: 19–58.
ICCAT . 1999. Report of the meeting of the ICCAT ad hoc working group on precautionary approach. Dublin, Ireland, 17–21 May 17 1999.
Ivanov, L. y Beverton, R.J.H . 1985. The fisheries resources of the Mediterranean. Part 2: Black Sea. GFCM Studies and Reviews , 60: 135pp.
Lleonart, J. y Maynou F. 2003. Fish stock assessment in the Mediterranean, state of the art. Sci. Mar. , 67 (suppl. 1): 37–49.
Margalef, R . 1985. Introduction to the Mediterranean. In R. Margalef (Ed). Western Mediterranean. Series Key Environments . Pergamon Press, 363 pp. 1–16.
Martin, P., Sartor, P. y García-Rodriguez, M. 1999. Comparative analisis of the exploitation strategy of the European hake (Merluccius merluccius), Red mullet (Mullus barbatus) and striped red mullet (Mullus surmulletus) in the Western Mediterranean . Journal of Applied Ichthyology , 15: 24–28.
Mee, L.D. 1992. The Black Sea in crisis: A need for concerted international action.AMBIO , 21 (4): 278–286.
Oliver, P. 1993. Analysis of fluctuations observed in the trawl fleet landings of the Balearic Islands. Sci. Mar. , 57 (2–3): 219–227.
Oliver, P. 1996.Ressources vivantes de la Méditerranée occidentale: Evaluation des stocks et recommandations scientifiques aux fins de l'aménagement.FAO Fisheries Report, No. 537: 23–56.
Oliver, P. 2003. Mediterranean fisheries. In Development and agri-food policies in the Mediterranean region. Annual report 2002. CIHEAM . Part III: 151–214.
Oray, I. y Karakulak, F.S. (eds) 2003. Workshop on Farming Management and Conservation of Bluefin Tuna . 5–7 April 2003. Published by the Turkish Marine Research Foundation, Istanbul, Turkey, 13: 145 pp.
Öztürk, B. y Karakulak, F.S. (eds). 2003. Workshop on demersal resources in the Black Sea y Azov Sea. Published by the Turkish Marine Research Foundation , Publication No. 14. Istanbul, Turkey, 129 pp.
Palomera, I. y Pertierra, J.P. 1993.Anchovy biomass estimate by the daily egg production method in 1990 in the Western Mediterranean. In J. Lleonart (ed.) Northwestern Mediterranean Fisheries. Sci. Mar., 57(2–3): 243–251.
Papaconstantinou, C., Caragitsou, E., Mytilineou, C., Petrakis, G. y Vasilopoulou, V. 1988a. Population dynamics of demersal fish stock in Corinthiakos, Patraikos Gulfs and Ionian Sea. National Center for Marine Research, Athens, Special publication,16: 275 pp.
Papaconstantinou, C., Mytilineou, C. y Panos, T. 1988b. Aspects of the life history and fishery of Red Pandora, Pagellus erythrinus (Sparidae) off Western Greece. Cybium , 12(4): 267–280.
Patti, B., Mazzola, S., Calise, L., Bonanno, A., Buscaino, G. y Cosimi, G. 2000.Echosurveys estimates and distribution of small pelagics Fish concentrations in the Strait of Sicily during June 1998. Paper presented at WG on small Pelagics. Fuengirola (Spain) 1–3 March 2000.11p.
Pertierra, J.P. y Lleonart, J. 1996. NW Mediterranean anchovy fisheries.In I. Palomera y P.Rubiés (eds). The European Anchovy and its Environment. Scient. Mar. , 60 (Supl.2): 257–267.
Prodanov, K, Mikhailov, K., Daskalov, G., Maxim, C., Chashchin, A., Arkhipov, A., Shlyakhov, V. y Ozdamar, E. 1997. Environmental management of fish resources in the Black Sea and their rational exploitation. GFCM Stud. Rev. , 69: 178 pp.
Quignard, J.P. y Tomasini, J.A. 2000. Mediterranean fish biodiversity.Biol. Marin. Med , 7(3):1–66.
Quintanilla, L.F., Garcia, A., Giraldez, A. y Cuttitta, A. 2000.Daily Egg Production estimate of the spawning biomass of the Sicilian Channel Anchovy during July 1998. Paper presented at WG on small Pelagics. Fuengirola (Spain) 1–3March 2000: 43p.
Ragonese, S. y Bianchini, M.L. 1996.Growth, Mortality and Yield per Recruit of the deep-water shrimp Aristeus antennatus (Crustacea-Aristeidae) of the Strait of Sicily (Mediterranean Sea).Fish. Res. , 26: 125–137.
Regner, S. 1990. Stock assessment of the Adriatic sardine and anchovy using egg surveys. Atti di seminario: Reproductive biology of small pelagics and stock assessment through ichthyoplanktonic methods. ICRAP Quaderno Pesca , 4: 17–31.
Sardà, F. (Ed) 1998. Nephrops norvegicus : comparative biology and fishery in the Mediterranean sea. Sci. Mar. , 62 (supl. 1), 1998 143p.
Somarakis, S. y Tsimenides, N. 1997. A daily egg production method biomass estimate of the northern Aegean sea anchovy stock. Ozeanografika , 2:133–148.
Stergiou, K.I., Christou, E. y Petrakis, G. 1997a.Modelling and forecasting monthly fisheries catches: comparison of regression, univariate and multivariate time series methods. Fish. Res., 29: 55–95.
Stergiou, K.I., Christou, E.D., Georgopoulos, D., Zenetos, A. y Souvermezoglou, C. 1997b. The Hellenic Seas: physics, chemistry, biology and fisheries. Oceanog. Mar. Biol. Ann. Rev. , 35: 415–538.
Tursi, A., Matarrese, A., Donghia, G., Sion, L. y Maiorano, P. 1995. The yield per recruit assessment of hake (Merluccius merluccius L., 1758) and red mullet (Mullus barbatus L. 1758) in the Ionian Sea. FAO Fisheries Report, No. 533 (Suppl): 127–141.
Ungaro, N., Rizzi, E. y Marzano, M.C. 1994. Utilizzo del modello di Beverton e Holt, “Rendimento per recluta (Y/R)”, per la risorsa ” Mullus barbatus L.” nell'Adriatico Pugliese.Biol. Mar. Medit. , 1(1): 317–318.
Zaitsev, Yu. 1993. Impacts of eutrophication on the Black Sea fauna. GFCM Stud. Rev. , 64: 6–85.
Zaitsev, Yu. y Öztürk, B. (eds). 2001. Exotic Species in the Aegean, Marmara, Black, Azov and Caspian Seas . Published by Turkish Marine Research Foundation, Publication No. 8. Istanbul, Turkey, 267 pp.
* FAO, Servicio de Recursos Marinos, Dirección de Recursos Pesqueros.
![]()
![]()
![]()