![]()
![]()
![]()
by/par
W. Nellen & D. Sohnack
Institut für Meereskunde an der Universität Kiel
Federal Republic of Germany/République fédérale d'Allemagne
ABSTRACT
This paper lists and considers details of the various methods that are being used for sampling the eggs and larvae of fish. Special attention is given to surveys of these stages of freshwater species. A great diversity of methods is necessary because the eggs and fry of different species are found in a greater range of habitats and as a result fish larvae surveys in freshwaters may contribute to a general knowledge of larval ecology.
RESUME
Ce document expose les diverses méthodes utilisées pour échantillonner les oeufs et larves de poisson. Une attention spéciale a été prêtée aux enquêtes de ces stades ohez les espèces d'eau douce. Une grande diversité de méthodes sont nécessaires du fait que les habitats des oeufs et des alevins des différentes espèces se trouvent dans des zones variées. Il résulte que les études des larves de poisson en eaux douces peuvent oontribuer à la connaissanoe générale de l'écologie du stade larvaire.
CONTENTS
1. GENERAL ASPECTS OF SAMPLING FISH EGGS AND LARVAE
2. METHODS USED IN MARINE FISH EGGS AND LARVAE SURVEY
3. SOME NOTES ON ACCURACY AND PRECISION OF SAMPLING RESULTS
4. FISH EGGS AND LARVAE SAMPLING IN INLAND WATERS
4.1 Sampling of Eggs
4.2 Sampling of Larvae
5. CONCLUSIONS
REFERENCES
In marine fisheries egg and larval surveys are well established research which have contributed to several fields of marine research and have broadened our knowledge of fish ecology, early life history, systematics, marine resources, and biological oceanography in general.
Examples of major aspects of fisheries research in which surveys of eggs and larvae play a part are species distribution (Ahlstrom, 1965); occurence, size and changes of spawning stocks as an appraisal of fishery resources (Saville, 1964); larval survival and stock recruitment as an aspect of population dynamics (Gulland, 1965); and response of fish fry to environmental conditions as an indicator of changes in the environment due to climate, hydrography or pollution (De Sylva, 1970).
To achieve reasonable results within the different fields of eggs and larval surveys, investigations have to be carried out on the assumption that fry are distributed evenly in the water, i.e., that eggs and larvae do not stick to niches which are difficult to sample by gears.
Eggs and larvae of most of the more important marine fish species are entirely pelagic throughout a certain period of their life and can be found within the upper 200 or less meters of the water column. There is no evidence that fish larvae move through the thermocline while making diurnal vertical migration. Most kinds of fish larvae are confined in their vertical distribution to the upper mixed layer (Ahlstrom 1959). Hence they may be caught fairly easily with plankton sampling gear.
With exception of the “distribution of species” aspect, fish egg and larval surveys have to be done quantitatively, because fishery biologists intend to make quantitative statements from results gained by this kind of research.
The necessity has induced many methodologists to search for the best gear and the following obstacles have to be overcome to get representative samples which can be used for valid interpretation of the fry and fish stocks; clogging of net gauze and changes in the filtration efficiency; avoidance of the gear by the fry; and extrusion of the fry through the meshes of sampling nets.
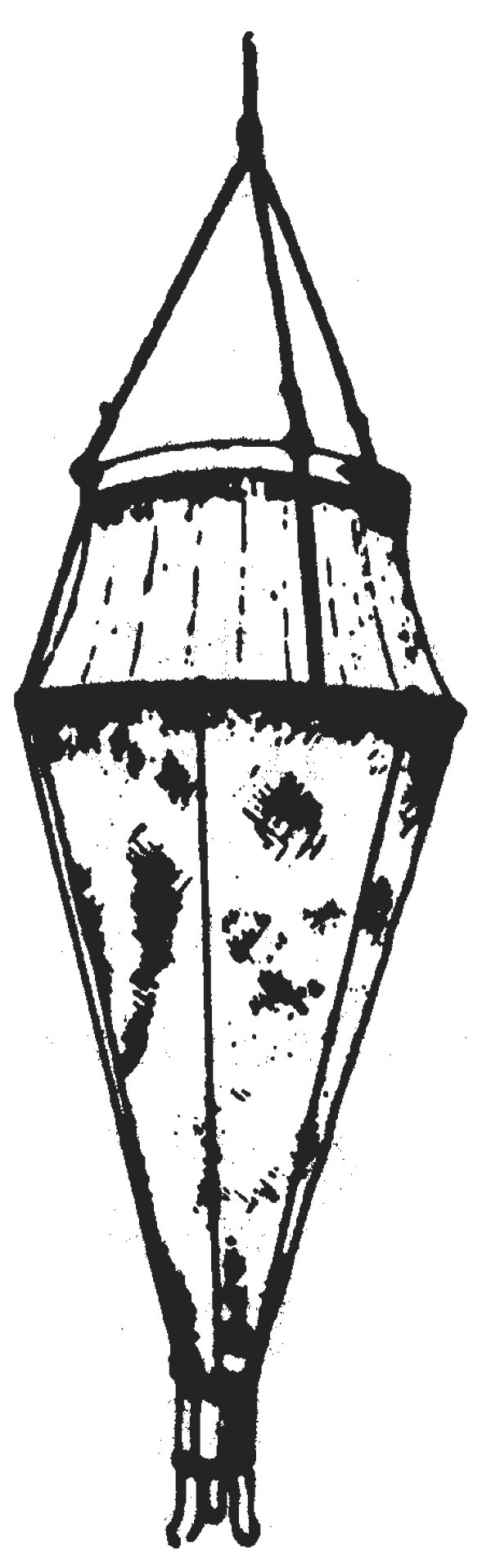
Many different gears have been used for quantitative collections of fish eggs and larvae on survey cruises since scientists started to study plankton (Unesco, 1968). Initially samples were collected from the stopped ship and nets were towed through the water column vertically from a certain depth to the surface. Two well known gears which were used in this way are the Hansen Egg Net and the Heligoland Larvae Net (Fig. 1).
Later it became obvious that catches were more effective when tows were performed from a moving ship. For standard hauls nowadays nets are usually lowered at a ship's speed between 2 and 8 kn (depending on the type of gear) down to a depth at which fish larvae can be expected, then the net is retrieved. Oblique tows which are done in such a way, have the advantage that much more water is strained within each depth layer than in vertical tows.
Almost all of the extensive nationaland international fish eggs and larval surveys of the past ten to twenty years chose oblique tows for sampling fish fry. The following three types of nets have mainly been used; the CalCoFi net (Fig. 2); the Gulf III-sampler (Fig.3); the Bongo-net (Fig.4).
The CalCoFi net is handled at a low ship's speed of two kn. Its mouth opening is 1 m in diameter, so it is relatively large compared to other gears. The net bag is cylindrical shape in front and becomes conical toward the rear. The cylindrical part has a self-cleaning effect which is of great advantage regarding the clogging problem. A bridle precedes the net when towed.
The Gulf III was designed as a high speed sampler. A mouth cone reduces the mouth opening of the net from 45 cm to 20 cm. The net is encased by a solid metal pipe but unencased versions of the Gulf III are used as well and in the same way. The gear is suspended from the towing wire in a way that no bridle precedes the net when in action. Towing speed is 4 to 6 knots.
The Bongo is a paired net. Two rings or cylinders of 60 cm diameter each are combined by an axis which also holds the towing wire. This assembly may be towed at low speeds as well as up to 6 knots.
There exist many similar gears and several modifications of the three types just described, which more of less differ in their sampling efficiency. Beside these gears which may sample the whole water column the neuston net came into use several years ago (David, 1965; Hempel and Weikert, 1972). This sampler strains the upper 10 cm of the water column only. The net is mounted on a catamaran-like frame to which a second net, fishing the depth layer 10 to 25 cm below the surface, may be fixed (Fig. 5). Neuston nets are useful for getting large numbers of those fish larvae and eggs which aggregate at the surface and which are hardly caught by ordinary gear.
For international survey programmes on fish fry Gulf III and Bongo are recommended as sampling gears. The Gulf III is used for instance within the ICES herring larval survey in the North Sea. The Bongo is used for the same purpose in the ICNAF area and is becoming generally accepted for all kinds of fish eggs and larval surveys. The reliability of abundance estimates for plankton organisms based on sampling with these gears is, however, still under discussion.
The accuracy of the results depends on knowledge of the sampling efficiency of the gear. In this respect not only filtration efficiency, avoidance and extrusion, but also handling of the gear has to be considered.
Filtration performance in simple plankton nets without encasement is fairly well understood. If the ratio of effective straining (aperture) area to mouth area is not less than 5 to 1 the initial filtration efficiency approaches closely to 100 percent. This is the fact for all Bongo models. In encased samplers with a mouth reducing cone, on the other hand, filtration performance is much more complex, depending primarily on the shape of encasement rather than on the filtering surface (Tranter and Smith, 1968 et al.). According to channel tests, initial filtration efficiency of the Lowestoft model of Gulf III is less than 100 percent (Harding, 1969) whereas for the German “Hai”, which is equivalent to the Dutch type of Gulf III, it is slightly more than 100 percent. Efficiency has to be tested for any model of different shape separately. Moreover differences in filtering capacity, depending on the degree to which the nets are affected by clogging, have to be considered. In this respect, cylinder cone nets have proved better than simple conical nets (Smith, Counts and Clutter, 1968) and the unencased Gulf III model seems to be less quickly affected by clogging than the encased version (Shnack, 1974). Clogging is of special importance as reduction in filtration efficiency during an oblique tow may affect the accuracy of sampling results in several ways:
It may lead to unequal amounts of water being filtered at different depths and hence organisms, not homogeneously distributed over the total depth range, will not be sampled representatively for that water column.
Selectivity of the net will change during a tow as larger fish larvae may avoid even high speed samplers with reduced filtration efficiency and the proportion of small organisms extruded through the meshes will be reduced when clogging occurs (Schnack, 1974; Bridger, 1958; Saville and MoKay, 1970).
Thus, it is necessary to measure the total amount of water filtered during a tow by means of a flow meter mounted in the mouth of the sampler. The filtering capacity of the sampler model must also be sufficient for the particular situation under study and the desired duration of tow. This may be checked by comparing flow meter readings inside and outside the sampler (Tranter and Smith, 1968).
At high filtration efficiency avoidance by fish larvae is apparent only when sampling for the older more mobile larvae with low towing speeds (about 2 kn) whereas good evidence is given that sampling results will not be biased by avoidance when fishing at high speeds (about 5 kn) (Saville and McKay 1970; Posgay and Marak, 1967). In the latter case, however, extrusion may affect accuracy of results by reducing the proportion of very small larvae in the samples. This extrusion effect is obviously more vigorous in the encased Gulf III than in the unencased version of the Bongo (Schnack, 1974; Sherman and Honey, 1968).
In any case mesh size and towing speed have to be adjusted according to the size and mobility of organisms sampled for, and take into account the specific filtration pressure of the gear used. With regard to sorting effort mesh size should, of course, not be smaller than necessary for retaining the desired organisms quantitatively. At low towing speed a mesh size of 0.5 mm will retain most fish larvae (Ahlstrom, Sherman and Smith, 1973). At high speeds, on the other hand, several small forms of larvae will be pressed through meshes of this size as even organisms which are somewhat larger than mesh width may be extruded in this case (Schnack, 1974; Vannucci, 1968). To allow for all possibilities it is recommended that the Bongo should be used with its nets having different mesh sizes (0.5 and 0.3 mm) and be towed at two different speeds (1.5 to 2 and 4 kn) (Ahlstrom, Sherman and Smith, 1973).
Sampling results may also be biased by the handling of gear. Towing distance has been mentioned already in connexion with clogging. Towing profile and direction are other important factors when there is a non-homogeneous vertical distribution of organisms and differences in current speed with depth. Both play a part in conflict of result in comparative tests of samplers. To ensure a linear tow profile, and hence an equal amount of water strained per depth stratum, an effective depressor is essential. At low towing speeds a simple dead weight will be sufficient; at high speeds mostly a V-fin depressor is used which mainly works due to shearing strength. Possible differences in currents may be adjusted by successive hauls in opposite towing directions. In the CalCoFi programme this problem has been met by regulating the ship's speed in such a way that the wire angle remained constant.
In general, all factors affecting the accuracy of sampling results are open to control. Precision, on the other hand, is much more difficult to deal with, as it depends on the distribution of organisms. There are major difficulties in estimating the sampling error (Cassie, 1968). However, it is obvious from statistical theory that precision may be improved by the number of hauls within a certain area and also by the size of samples, as long as the mean number of organisms considered, is still less than 100 per haul. The variation between hauls - and hence the degree of improvement by increasing the number of hauls - depends on horizontal changes in abundance, due to patches larger than the distance of horizontal integration by the sampler (Schnack, 1974).
Battle and Sprules (1960) give a comprehensive review on the investigation of several authors about characteristics of eggs from marine and fresh-water species. It is apparent from their review that most fresh-water fishes produce heavy eggs which sink to the bottom immediately after extrusion. Many fresh-water fish produce adhesive eggs which are fastened to underwater weeds or to the gravel on the bottom. This is the case with almost all of our cyprinids, with perch-like fishes and with the pike. Others, salmon, trout and grayling bury their eggs in the sediment so that they are not exposed to free water.
The only truly pelagic egg we have been able to trace is that of Aplodinotus grunnieus, a fresh water sciaenid from Lake Erie and other North American lakes which is described by Davis (1959). The eggs of the “sheeps head” are able to float because of a large ventral oil droplet.
Semi-bouyant Eggs:
Most fish eggs which are described in the literature as pelagic (Schäperolaus, 1963) are in fact either free floating at the bottom from whence they are sucked up into the open water by currents, or they are at the water surface, having been spawned in floating foam nests built by the adults. The first category is sometimes called semi-buoyant. This can be accepted as a reasonable term for the situation in natural surroundings. It means that the specific gravity is slightly greater than the water. Fish species having such eggs are for instance the burbot (Lota lota), the grass carp (Ctenopharyngodon idella), Pelecus cultratus, the twaite shad (Alosa alosa), whitefish (Coregonidae), the goldeye (Hiodon alosoides). The eggs of the European smelt are adhesive when spawned, but they may become free floating later (Lillelund, 1961).
A survey of the semi-pelagic eggs of the twaite shad in the river Elbe has been described (Hass, 1968). A plankton net was set horizontally from an anchored boat heading against the direction of the current (Fig. 6). The net could be set at different depths and the volume of water filtered was measured with a current meter and water turbulence. Eggs were caught up to 9.5 m above the bottom depending on the current. More than 97 percent of the total, however, occurred in water layers between 0 and 2.5 m above the bottom. However, the number of eggs that lie directly on the bottom and therefore not available to this catching method is unknown and such a complicated depth distribution of eggs make it diff cult to get representative samples from which the total number of eggs spawned can be calculated.
Adhesive Eggs:
Adhesive eggs can be sampled by gathering the spawning substrate from weed beds or shallows. Such a technique, of course, can by no means be called quantitative. To overcome this disadvantage sampling units placed throughout the spawning area may provide representative samples of eggs.
Enumeration of eggs on such units give an estimate of egg production and egg survival parameters. For smelt eggs sampling units of glazed black ceramic tile, 11.5 cm square, with two egg-depositional surfaces of heavy black canvas strips, 5 cm wide and about 12.5 cm long were used (Rothschild, 1961). The narrow ends of each strip were joined by a rubber band stapled to each end, which formed a collar-like arrangement that permitted each strip to be slipped easily on and off the tile. When using these sampling units two assumptions have to be made. The first is that the attachment of eggs to the depositional surfaces is equivalent to their attachment to the natural substrate. Secondly, it has to be assumed that both surfaces on each unit are equally available for egg deposition. The validity of these assumptions has been proved. Suspected large variability in egg densities suggested that a maximum number of sampling units be used. The eggs on one surface of a unit were enumerated each morning, this surface was then replaced by a new one. The second surface remained, so that the eggs accumulated. The temporary surface gave data on egg production and the permanent one was used to determine egg survival. The estimate of egg production followed methods of representative but disproportional sampling (Schumacher and Chapman, 1954).
In many species females often distribute eggs over a considerable area and the spawning period is usually protracted with extrusions of small numbers of eggs each time mating occurs (Bagend and Braun, 1971). So it is difficult to get representative samples by exposing sampling units, unless such units are distributed very accurately in the lake or river. Sampling of horring eggs is similar to the sampling of adhesive eggs of fresh-water fish. They are also non-buoyant, and quantitative sampling is almost impossible. When water is sufficiently clear and spawning beds are not too deep, free diving techniques and direct observations had helped to overcome the problem (Tibbo, Scarratt and McMullon, 1963).
Buried Eggs:
In salmonids, which bury their eggs in the gravel of river beds, egg numbers may be estimated by counting the redds soon after spawning took place within a certain section of the river. Average numbers of eggs in a redd must be gained by digging out whole redds. The eggs which are relatively light are carried downstream over a certain distance and can be sampled by nets properly arranged in the current (Briggs, 1953). Redds may be marked by stakes.
Some aspects of the biology of fresh water fish larvae:
Once the eggs have hatched most species of fresh water fish start their life as larvae. That means, from the morphological point of view they are plankton-like animals with restricted swimming ability (Schaperclaus, 1940; Elss, 1967). But the biological situation in fresh-water habitats is so different from the marine environment that fresh-water fishlarvae must not be expected to appear necessarily in the pelagic zone, where planktonic animals normally live. Compared to the sea the pelagic space is much smaller in inland waters, i.e., the proportion of extension of shore line or bottom to water mass is much larger. This might be one reason why almost no planktonic fish eggs exist. Furthermore the many larvae of several fresh-water fish species are not planktonic larval pike and many larval cyprinids, for instance, have adhesive organs on the head (Elss, 1967) and while the yolk-sac is being absorbed they attach themselves to plants. Thereafter they are probably already able to maintain their position in water and probably never occur in high numbers in the open. It is obvious, therefore, that marine sampling methods can not also be expected to be feasible in rivers and lakes. Factors depending on the behaviour of the species certainly play a great part in the distribution of fresh-water fish larvae, and this, certainly, will influence their catchability.
Distribution of fish larvae in inland waters:
For our own fish larval investigations in the Schlei, a brackish water fjord near Schleswig (40 km in length) we used a 1-m mouth diameter plankton net which was towed at a speed of two knots just below the surface from a small research vessel. Good catches of smelt larvae, 18 to 42 mm long, and perch larvae, 8 to 11 mm long were obtained in May and June. No larvae of cyprinids were caught by this method, however, though several species of this family inhabit the inner parts of the fjord. The salinity at which smelt and perch larvae were found was 7 to 8 percent, which concentration might have been too high for the young stages of cyprinids. Some cyprinid larvae, however, were gathered with a push net in the littoral zone among the reeds, and although high numbers of smelt and perch larvae were also caught it is not possible to say whether they were more abundant in the open water than in the littoral.
Similar observations were made in the Kiel canal which is also inhabited by several kinds of fresh-water fish species where the salinity is no longer an important ecological factor. We caught high numbers of smelt larvae and young sticklebacks in July, but only one cyprinid 50 mm long.
Another inland water in Northern Germany where a fish larvae survey was made is the Eider (Langford and Jermoloyer, 1966). This is a river which is now separated from the North Sea and opened only periodically. Limnologically it is a very long lake, some 80 km in length and up to 250 m wide, rather than a river.
In May, July, September, and October 1969 plankton samples were taken monthly using the same method as that described for the Schlei-fjord. Fish larvae of the following fresh water species were caught: Osmerus eperlanus, (several thousands), Acerina cernua (“few”), Lucioperca lucioperca (four), Gasterosteus aculeatus (“many”), Abramis brama (one), and Gobio gobio (one). These investigations took place in the lower part of the river which is still influenced slightly by the sea, but because salinity is below 2 percent degrees in most parts of the area, reproducing stocks of fresh-water fish certainly exist (Nellen, 1965). These catches were under-representative of all species except the smelt and the stickleback.
During the second half of May and in June 1969 a neuston net survey was performed in the upper part of the Eider, which is not influenced by the sea. Three hauls with a double neuston net were made at any of five different stations, two parallel and as close as possible to both of the shores and one in the main channel.
The catches were to investigate the influence of the wind on fish larval distribution. Even holoplanktonic organisms - (D'Ancona, 1955) - may not be scattered evenly throughout the upper layer of a lake. The wind regime contributes much to the distribution of these organisms; concentrations of zooplankton, for instance, having been observed at the wind exposed side of lakes (Langford and Jermolejet, 1966; Colebrook, 1960).
Considerable numbers of larvae of Osmerus eperlanus (6–11 mm), Acerina cernua (5–7 mm), and Cyprinids (Abramis brama and Rutilus rutilus, 7–11 mm long) were caught. Additionally some larvae of Lucioperca lucioperca and Perca fluviatilis (18–23 mm) and one pike larva (17 mm) were found, as well as adults of Alburnus alburnus which were in spawning condition.
Distribution of fish larvae was as follows: Percids were most abundant at the weather shore. The ratio of numbers of larvae caught at this side of the river to numbers caught at the lee-side were 12 to 0 in the upper net and 118 to 16 in the lower net. In the main channel a total of 27 period larvae were caught, but only by the lower net.
Smelt larvae were found almost entirely in the lower net. No influence of the wind had been observed, numbers sometimes were high at the lee-side as well as at the weather side as in the water way.
Larvae of cyprinids, interestingly enough, were missing completely in mid-channel. Numbers of specimens caught at the weather-side to specimens caught at the lee-side were 34 to 52 in the upper net and 25 to 17 in the lower net.
Larvae of the smelt and of percids were less abundant in the uppermost surface layer than larvae of cyprinid.
It is obvious from these investigations, that fish larvae are not affected much by the wind but may be found in areas which seem to be suitable for their ecological demands. On the other hand the invertebrate plankton in the surface layer was found to be concentrated always close to the lee-shore. Adults of Alburnus alburnus and young eels were also caught here and it is likely that this was because of high aggregation of prey animals. A similar observation was made for smelt larvae caught in the Kiel Canal and in the lower part of the Eider where high numbers were correlated with concentrations of mysids. From several other investigations it is known that the distribution of larval fish in inland waters may be biased by patterns of behaviour: In the multiple impounded River Main high concentrations of cyprinid and other fish larvae seemed to be more common for groynes than for open-shore line sections (Hofbauer, 1965). Larvae of the common Perch (Perca fluviatilis) have been described as active, and intensive swimming organisms (Schindler, 1934). Soon after hatching they move from the shores into the open water. This may happen partly passively with transport by surface currents because perch fry are said to be free floating due to the large oil droplet in their yolksac (Schneider, 1922). So far the larvae of this species may disperse like plankton occuring for a while more or less solitarily rather than in schools, but they soon move back to the shore zone when 8 to 20 mm long. This probably is related to high temperatures on the shallows (Schneider, 1922). The observation that 8 mm long perch larvae are already able to migrate corresponds with sampling results according to which young yellow perch begin to avoid plankton nets at this size (Noble, 1970). The distribution of walleye fry is probably biased even more than that of the fry of the former species and in Oneida Lake, New York, they mainly occupy protected bays. It is said of its European relative, Lucioperca lucioperca, that the larvae stay close to the sandy spawning grounds at a depth of two to four metres during the summer. In autumn they move into the reed-zone, preying on O-group cyprinids (Schneider, 1922).
Observations analogous to those made on the distribution of walleye larvae were made on young sockeye salmon in the Babine lake system, British Columbia (Johnston, 1956). Their distribution may in general be dependent on the distribution of the spawning parent population, the morphometry of the lake, and a limited dispersal of young sockeyes from their points of entrance into the lake as fry. This results, by the way, in inefficient utilization of nursery facilities in the lake.
Spawning of pike (Esox lucius) takes place in shallow and small backwaters or on seasonally submerged grasslands. After the larvae have moved down or become washed into the adult habitat, a lake or a river, they start a solitary life in wait for prey at locations near shore (Schindler, 1934). Pike may, therefore, not be expected to be pelagic at any stage of their life history.
Techniques suitable for fish larval surveys in inland waters:
The above examples of larval distribution show that sampling techniques have to be different in inland waters from those commonly used in the marine environment. Nevertheless, well proved standard methods known from marine fish larval investigations may be recommended also for inland waters whenever they appear to be meaningful. This probably would be the case in a coregonid larval survey, for instance where the fry of white fish can be expected to inhabit the epipelagic zone as do the adults.
In addition to the biological obstacles there are also some of a technical nature. Powerful research boats are needed, for example, to handle a Gulf III-sampler or a Bongo-net. Such boats may not always be available in fresh water research areas.
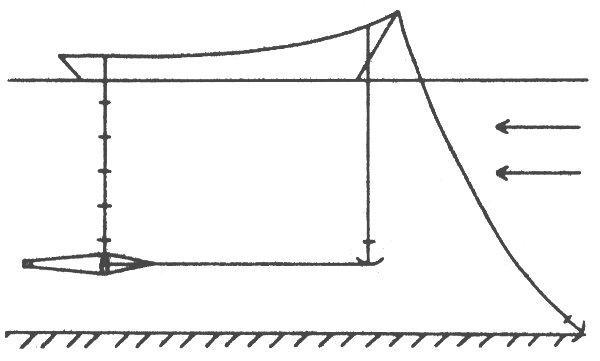
A method for using a plankton sampling gear similar to the CalCoFi net in lakes is described for a sockeye survey (Johnson, 1956). The 1–m mouth diameter net (Fig. 7) was towed by means of two boats powered with outboard motors (15– to 25–horsepower), the boats running parallel about 30 m apart. Nothing preceded the mouth of the net while fishing. For surface tows each boat was equipped with a 30–m tow line of 6 mm nylon rope. In order to fish at greater depths, it is necessary to attach a depressor to the tow lines, which requires greater towing power, longer tow lines, and winches for hauling the net. Experience has shown, however, greater fishing success with surface tows. At dusk, fishing success for young sockeye increased quickly and reached a maximum during the first moments of darkness then decreased and at night catches were rare. No corresponding period of high catchability had been observed for the morning. The same daily pattern of catchability had been observed in several sockeye lakes.
If fish fry are to be expected at the surface the David neuston net (Fig. 5) can also be successfully used. Another surface plankton sampler has been described recently (Miller, 1973). This gear is especially suitable for inland and inshore waters. It lacks the disadvantages of a preceding bridle and wire and operates outside the boat's wake. It can be used for relatively fine-scale pattern, it is self-cleaning, and can be operated by two investigators from a relatively small, shallow-draft catamaran or other twinhulled boat. The major components of the sampling system are two standard Bongo-nets, a frame of galvanised pipe, and a catamaran (5.3 m) powered by two 40 hp motors (Fig. 8). The mouth area of each side of the net frame is 0.36 m2. The net frame is suspended from the main frame by cables, which allows for vertical oscillation producing only gentle undulations of the nets. This also contributes to lack of clogging and to good condition of the fish larvae. While sampling one investigator steers the boat, the other one removes the samples from the nets through the well in the catamaran floor. Net mouths remain continuously in the water, permitting sampling of contiguous quadrants. At an operating speed of 3 mph 505 μ Nitex nets yielded fish larvae 2 to 25 mm long and frequently alive after 3 min towing. Net bags were found to be most suitable when made out of 303 μ Nitex for in larger mesh bags fish larvae were damaged to a greater extent. The principle advantage of the sampling technique is its utility in detecting small-scale variations in abundance of surface zooplankton.
Another gear which has been used successfully in inland waters is the Miller high-speed sampler (Miller, 1961). Because of its small size (10-cm mouth aperture) it can be operated from a small boat powered by outboard motor and also allows sampling of deeper water layers in a larval survey of yellow perch and walleye in Oneida Lake, New York. The Miller sampler has been tested for efficiency (Noble, 1970). Equipment has been designed for this purpose which allows high sampling speed and maintains sampling depths more precisely than is possible when towing on cables. Two aluminium pipes were suspended vertically from a hinged clamp on each side of the stern. The poles reached to a depth of 3 m. A sampler was attached 1 m from the bottom of each pole (Fig. 9). The pole apparatus was used for several tests:
For comparison of several towing speeds.
For comparison of the standard Miller sampler with a Miller type sampler having a sampler body and aperture area 1.5 times the standard size.
For testing an electric shocker which produced an electric field in front of the sampler and which was expected to positively influence the efficiency of the gear. The shocker consisted of two 61 cm diameter grids held 23 cm apart by polystyrene rods, which also insulated the shocker from the aluminium pole (fig. 10). An AC generator in the boat supplied a current of 220 Volts.
For evaluating the effect of conspicuousness of the sampler by utilizing translucent and clear samplers instead of the conventional opaque, dark-green fibreglass samplers.
with the following results:
A standard Miller sampler towed at 8 mph during daylight hours could be avoided by yellow perch and walleye fry. Avoidance apparently begins when fry are less than 8 mm long.
Higher speeds were shown to be effective in increasing catches per volume of water filtered.
Both night/day comparisons and comparisons of translucent or clear samplers with dark samplers indicated that avoidance was a response to a visual stimulus.
Avoidance could be decreased by immobilizing and disorienting the fry through the electric shocker.
The bigger sampler did not appear to be more efficient.
Speeds up to 11 mph apparently did not eliminate avoidance by yellow perch fry. Relative efficiencies at 11 mph were lower than those in shocker evaluation and night sampling.
Differences in avoidance with depth were illustrated by the clear sampler evaluations, where avoidance was consistently higher at 0.5 m than at 1.5 m. Light intensity differed by 25 percent between the two depths.
In biological oceanography the abundance of fish eggs and larvae is usually referred to as specimens per m2 of sea surface. Whenever larvae of fresh water fish species are irregularly dispersed within the water column of a lake the best measure for abundance would also be number per m2 of water surface. This permits vertical or oblique tows of the sampling gear through the whole water column or down to a depth at which larvae still occur. But powerful winches are necessary for plankton hauls of this kind, which therefore are difficult to realize in most limnological research programmes. In this respect a “simple”vertical haul with a Heligoland larvae net, for instance, may be even more difficult to perform, because a which has to be used not only for hauling the net but also to haul at a speed of at least 1 m/sec-1 to hold avoidance factors to a minimum. In oblique tows, the speed of the net can be regulated more easily by the motor of the boat.
If vertical tows are performable we have to keep in mind that these also show increasing efficiency with darkness. In a marine fish larval survey where a Heligoland larvae net had been used average numbers of species in a certain area were found to be 2.9 times higher in night hauls than in day hauls, and numerically the night hauls were 5.3 times as rich as the day hauls (Murphy and Clutter, 1972).
The general lack of mechanical energy on fresh-water research boats and the scattered distribution of larvae in a multiple habitat environment make it difficult to sample larvae by the classical approach, that is the towed net. A gear which would overcome many of the disadvantages of tow nets in inland waters, is a purse seine. A seine is an extremely effective large volume grab sampler which, if not too large, may be operated without the help of any machine and by which further the whole assemblage of organisms occuring in a small section of an environment can be gathered. Avoidance of organisms to such a gear can be expected to be low, so that samples are less biased than tow net samples are. A plankton purse seine for catching anchovy larvae has been described (Murphy and Clutter, 1972). The body of the net was constructed of 333 μ Nitex. The length of the Nitex section was 30 m and the depth 7 m. A small cod end was placed near the end of the net. The net was set in the form of an incomplete circle. Closure of the circle was effected during the first stages of hauling. The operation from starting the set to pursing generally took less than 5 min. Hauling in the net required 15 to 20 min with two men at work. No plankton was caught on the net as it was being hauled up, apparently because the plankton avoided the mesh. During the final stage it was necessary to carefully wash the catch toward and into the cod end.
The seine was not only easy to handle, but comparison of day purse seine and tow net (1-m mouth aperture) data indicated remarkable undersampling especially of the larger larvae by the tow net. At night the purse seine was also superior. This kind of gear seems to be almost ideal for fresh-water fish larval surveys, though, to our knowledge, it has never been used yet in corresponding programmes.
But even a plankton purse seine can not be an allround fish larval sampling gear for inland waters. Several species - especially cyprinids - are to be expected first of all in the shore zone among floating and emergent vegetation. Here no type of moving gear can be operated, and larval fish may be caught only by traps. A very effective one has been constructed from pieces of clear plastic (Bagenal and Braum, 1971; Breder, 1960), as shown in fig.11. The wings are removable, being attached to the box by rubber bands that encircle them and hook on to lugs cemented to the outside of the trap. The trap can be set on the bottom or suspended at any depth from a float. It is, of course, difficult to quantify such samples, although relative abundance of larvae of certain areas of a water may be outlined by this method.
For the very special case of sampling emergent fry of salmon or trust another trap is known from the literature (Heard, 1964) a modified version of which is given in fig. 12. The trap is stuck into the bottom of the river bed at locations where disturbed gravel indicate salmon redds. The lower part of the trap is open at the bottom. Because salmon may deposit their eggs in pockets, which results in a patchy distribution throughout a redd, the trap has to be tested at several points within a redd if it does not cover the redd completely.
A trap for the capture of fish and other organisms moving downstream has been constructed in Sweden (Wolf, 1951). This is a very heavy device, suitable not only for sampling fish larva but also bigger specimen.
Methods for sampling fish larvae in inland waters have to be manifold. This is because in fresh water the fry of different species stay at different habitats or niches in their environment.
The difficulty of catching fresh-water fish larvae proportionally to abundance is reflected by many publications dealing with all kinds of different sampling devices. This and the fact that heavy gears such as tow nets are usually not easily handled from the boats available in fresh water research may have contributed to the reluctance with which fish larval studies have been carried out in inland waters. But gear such as a plankton purse seine might become a fairly useful standard sampling device for fish larval surveys in relatively quiet-watered lakes and rivers.
For marine fishery biologists it is quite fascinating to see how diverse a distribution fish larvae may show in a lake or a river. Though regarded as plankton-like creatures without effective swimming ability larvae of at least some fresh-water fish species show that they obviously have the capability to move into certain areas of the environment and (or) stay there regardless of wind-induced turbulence or currents. This raises the question to what extent a fish larvae assembly in the sea might be structured too. It could be possible for instance that fish larvae are able to stay close to or within patches of planktonic organisms whenever these are suitable for them to prey on. It also may be that larvae do search for such patches and that they find them more or less by chance.
Towed nets, generally in use for marine surveys, integrate the organisms, living along a transect of considerable length, making it difficult to consider the catch from a tow as representing an assemblage of co-existing organisms (Murphy and Clutter, 1972). It is the fresh-water environment which creates the suspicion that special assemblages can be typical even for very small organisms. Results of fish larval surveys from there may contribute to a general knowledge of larval ecology and may stimulate biological oceanographers dealing with fish larvae to look at new aspects of their subject.
Fig. 1: Heligoland Larvae Net
 | 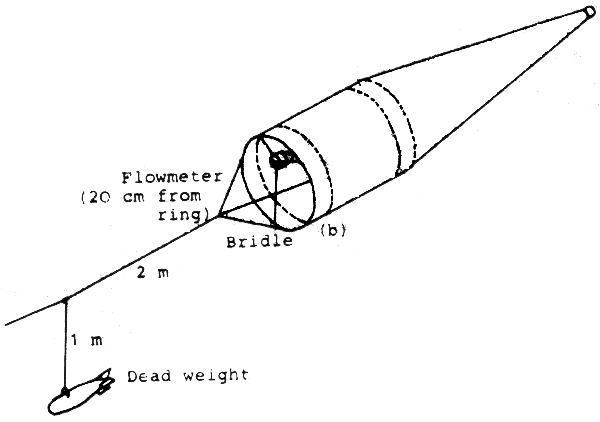 Fig. 2: CalCoFi Net |
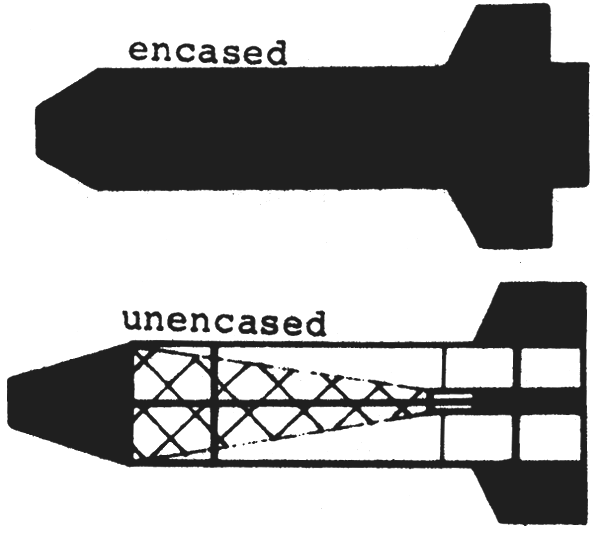 | 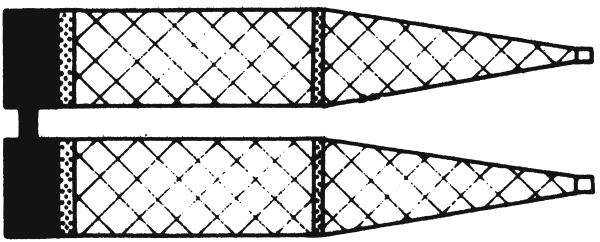 |
| Fig. 3: Gulf III-Sampler | Fig. 4: Bongo-Net |
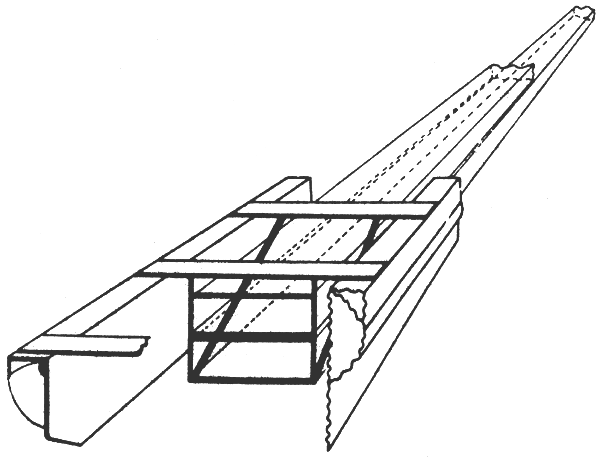 |  |
| Fig. 5: Neuston catamaran with two nets. Diagram (H8) el, 1972) | Fig. 6: Arrangement for sampling semibuoyant eggs in a stream (Hass, 1968) |
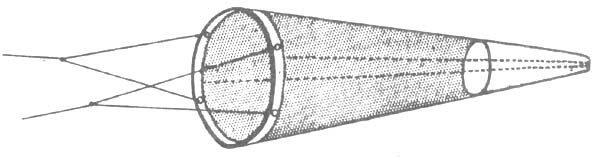
Fig. 7: Tow-net used for catching young sockeyes (Johnson, 1956)
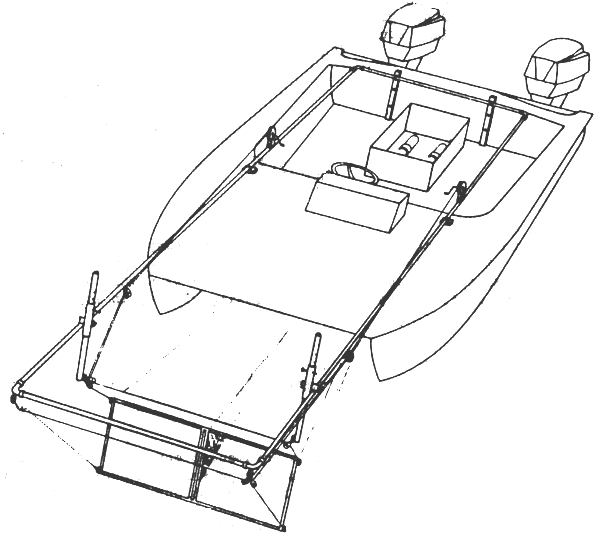
Fig. 8: Surface plankton sampling device (Miller, 1973)
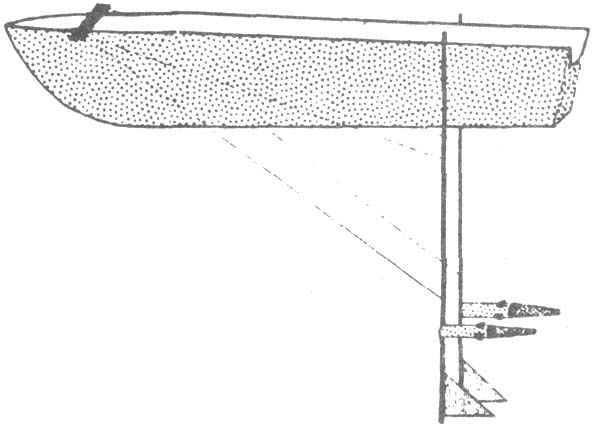 | 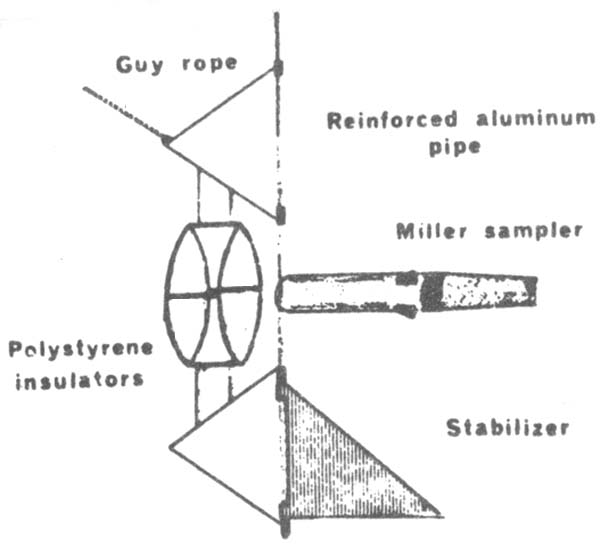 |
| Fig. 9: Pole apparatus developed for evaluation of Miller high-speed sampler (Miller, 1970) | Fig. 10: Electric fry shocker mounted in front of a fish larvae sampling gear (Miller, 1970) |
![]()
![]()
![]()