![]()
![]()
![]()
by
MARIA SZUMIEC
Experimental Farm Ochaby
Laboratory of Water Biology
Polish Academy of Sciences
Kraków, Poland
Abstract
Observations on temperature variations were carried out in spawning, nursery and rearing ponds at the Solysz Experimental Farm of the Laboratory of Water Biology, Polish Academy of Sciences. The results indicate that in all ponds distinct differences exist in time-space temperature distribution. These differences depend on the size of the pond, depth being the decisive factor. Diurnal amplitude and the vertical gradient of water temperature change in inverse proportion to depth. Horizontal gradients are mostly caused by depth variations in different parts of ponds. The extent of pond surface influences thermodynamics only when wind velocity is high. Nursery ponds have more favourable thermal conditions than spawning ponds. Vertical thermal sections of all investigated ponds indicate that convections include the whole stratum of water. Increasing wind velocity leads to the formation of turbulent movements. Its influence increases with decrease in relative depth of ponds.
CARACTERISTIQUES THERMIQUES DES ETANGS DE CARPICULTURE
Résumé
Des observations sur les variations de température de l'eau ont été effectuées dans des étangs de reproduction, d'alevinage et de production de la ferme expérimentale Solysz dépendant du Laboratoire de biologie hydrologique de l'Académie des Sciences de Pologne. Les résultats indiquent que dans tous les étangs il existe des différences marquées de répartition des températures dans le temps ou dans l'espace. Ces écarts dépendent de la dimension de l'étang et notamment de sa profondeur. L'amplitude diurne et le gradient vertical de la température de l'eau varient en proportion inverse de la profondeur. Les gradients horizontaux sont généralement dus à des variations de profondeur dans les différentes parties de l'étang. La superficie de l'étang ne joue un rôle thermodynamique que lorsque la vitesse du vent est élevée. Dans les bassins d'alevinage les conditions thermiques sont plus favorables que dans les bassins de reproduction. La section thermique verticale de tous les étangs étudiés indique que des phénomènes de convection intéressent l'ensemble du plan d'eau. Lorsque la vitesse du vent s'accroît, il entraîne la formation de turbulences. Son influence est d'autant plus forte que le bassin est moins profond.
CARACTERISTICAS TERMICAS DE LAS CARPAS DE ESTANQUE
Extracto
Han sido llevadas a cabo observaciones sobre las variaciones de la temperatura en estanques de desove, de cría y de desarrollo en la Granja Experimental de Solysz del Laboratorio de Biología Acuática de la Academia Polaca de Ciencias. Los resultados indican que en todos los estanques existen señaladas diferencias en la distribución de la temperatura en el tiempo y el espacio. Estas diferencias dependen del tamaño del estanque, siendo la profundidad el factor decisivo. La amplitud diurna y el gradiente vertical de la temperatura del agua varían en proporción inversa a la profundidad. Las gradientes horizontales son determinados en su mayor parte por las variaciones de profundidad en las distintas partes de los estanques. La extensión de la superficie del estanque influye en la termodinámica solamente cuando la velocidad del viento es elevada. Los estanques de cría tienen condiciones termales más favorables que los de fecundación. Las secciones verticales termales de todos los estanques investigados indican que las convecciones abarcan todos los estractos del agua. El aumento de la velocidad del viento determina la formación de movimientos turbulentos. Su influencia aumenta al disminuir la profundidad relativa de los estanques.
In carp rearing, ponds of different sizes and depths are used according to the changing environmental requirements of fish at different stages of their life. The thermo-dynamics of these ponds show significant differences, which result from differences in the heat capacity of individual ponds related to the volume of water. Other factors such as the relation of depth to area, development of the bank line, and relation of the height of dyke to the surface of the water are also of importance (Geiger, 1942; Tuszko, 1952; Starmach, 1956). Certain differences in the thermal balance of different types of pond result from differences in the transfer of heat turbulence between air and water. This transfer is directly proportional to the vertical gradient of air temperature over the water, which is also a function of the size of the pond (Szumiec, 1962).
This paper describes the thermo-dynamics of different ponds used for the rearing of carp and discusses the thermal differences and the relationships between time-space alterations of temperature and the size of the pond.
Observations were carried out in carp ponds at the Golysz Experimental Farm of the Laboratory of Water Biology of the Polish Academy of Sciences (49° 51'North, 18° 46'West).
Measurements were made in spawning ponds with a surface area of 0.023 ha and a mean depth of 30 cm, there types of nursery ponds (nursery ponds I with a surface area of 0.47 ha and a mean depth of 40–50 cm, nursery ponds II with a surface area of 1.2 ha and a mean depth of 60 cm, nursery ponds III with a surface area of 1.7 ha and a mean depth of 80 cm), and ponds stocked with one- and two-year old carp (commercial fish) with a surface area of three to four ha and a mean depth of 100 cm.
The spawning ponds were used for 10 to 14 days, usually in the second half of May or first half of June, the dates depending on the meteorological conditions. Nursery ponds I are used in June or sometimes in the early part of July, nursery ponds II from the beginning or the second part of July to 15 August, nursery ponds III until the spring of the next year. Rearing ponds are used during the whole growing season, from the middle of April to the middle or the end of October. The present studies covered the period of flooding of spawning ponds, from the middle of May till the end of the growing season at the end of October.
Spawning and nursery ponds observed in this study belonged to a complex specially organised for research and production. The rearing ponds were on a neighbouring farm at a distance of 500 m from the first one. At both places the surroundings were similar and the ponds were not overgrown nor shaded by trees or buildings.
In conditions of sunny and windless weather all ponds stratified during the day, but with variations in the vertical temperature gradients (Fig. 1). At noon the mean difference between surface and bottom temperatures in shallow spawning ponds amounted to 2.9°C, and in rearing ponds to 1.5°C (Table I).
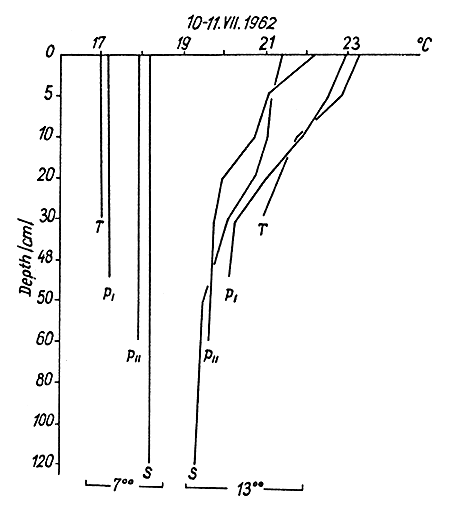
Fig.1 Vertical thermal stratification of water in conditions of sunny weather in a spawning pond (T), in nursery pond I (PI), in nursery pond II (PII), and in a rearing pond (S), (simultaneous measurements)
Table I
Differences of temperature (°C) between surface and bottom strata, sunny weather, in various types of ponds (mean noon values for a number of years)
| Spawning | Nursery I | Nursery II | Nursery III | Rearing |
| 2.9 | 2.3 | 1.6 | 1.3 | 1.5 |
Assuming that one m is the unit of depth, it was calculated that the vertical temperature gradient was seven times as great in the spawning as in the rearing ponds (Table II), and that the vertical gradient diminishes in proportion to the distance from the pond surface (i.e. the temperature gradient decreases with increasing depth, being greatest near the surface of the pond).
Table II
Vertical gradients of water temperature (°C/1m), sunny weather, in various types of ponds (mean noon values for a number of years)
| Spawning | Nursery I | Nursery II | Nursery III | Rearing |
| 9.7 | 5.8 | 2.7 | 1.6 | 1.3 |
The gradient calculated per meter depth and based on the entire water column is greatest in spawning ponds, reaching more than 12°C/m. (It is the equivalent of more than 30°C/m in the bottom strata where the fall in gradient is steepest in these ponds). The gradient is less steep in nursery ponds and ponds with flowing water than in spawning ponds (Fig. 1 and Table II), but in ponds with no current it also reaches the equivalent of 30°C/m in certain strata. Spawning ponds and larger ponds also differ in that there is no thermocline in spawning ponds. This begins to appear in nursery pond I; it occurs in these ponds at a depth of 5–10 cm, in nursery ponds II and III at a depth of 5–20 cm, and in rearing ponds at a depth of 20–40 cm. Isothermy, or possibly poikilothermy with a minimum gradient, is generally observed in all ponds, the latter being mostly observed in deeper ones.
An increase in wind velocity above a certain minimum value equalises the temperature differences between surface and bottom even during favourable insolation. The wind velocity causing isothermy varies for different types of pond, and depends on the temperature difference between surface and bottom waters (the calculated “relative depths”, after Starmach, 1958, given in Table III). In spawning ponds where this is greatest, a wind velocity greater than 6m/sec still does not cause isothermy, and the steepest gradient continues to be observed near the bottom (Fig. 2). In rearing ponds a wind velocity of about 3m/sec causes isothermy. Sellmayer (1934) mentions a similar influence of wind in a large pond.
Table III
Wind velocities causing isothermy in ponds of different relative depths
| Spawning | Nursery I | Nursery II | Nursery III | Rearing | |
| Relative depth 1 × 105/m | 130 | 11 | 6 | 5 | 5 |
| Wind velocity m/sec | > 6 | 5 | 4 | 4 | 3 |
Temperature profiles of these ponds with wind velocities of 4–6 m/sec are shown in Fig. 2, based on simultaneous measurements made during the noon hours. Only the rearing ponds were isothermal the whole time. Besides the differences in relative depth between the types of pond, the resistance caused by the water at high temperature (Kühlman, 1961) in the spawning ponds during sunny weather also counteracts mixing.
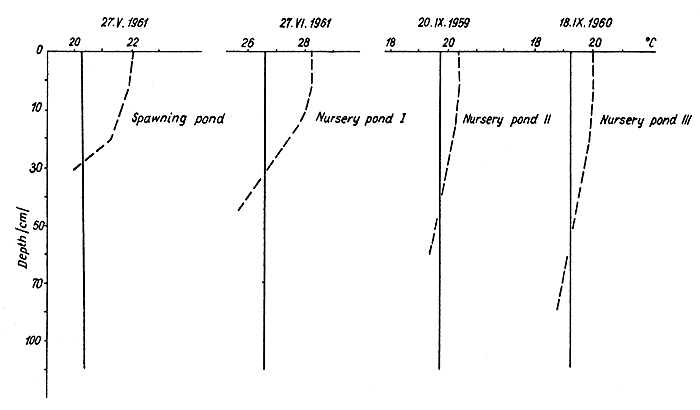
Fig.2 Vertical thermal stratification in spawning and nursery ponds ( - - ) and rearing ponds ( —— ), in sunny and windy weather, (simultaneous measurements)
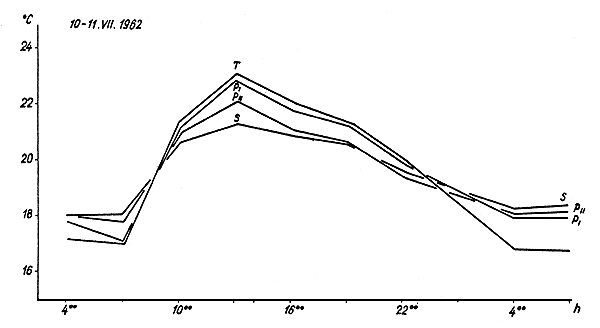
Fig.3 Diurnal changes in the temperature of the surface stratum of water in a spawning pond (T), nursery pond I (PI), nursery pond II (PII) and in a rearing pond (S), in conditions of sunny weather (simultaneous measurements)
Temperature changes per unit time in sunny weather are greatest in spawning ponds, least in rearing ponds (Table IV); this is mostly due to differences in depth between the ponds.
Table IV
Maximum differences of temperature (°C) in the surface layers observed in shallow and deep ponds, sunny weather, (mean noon figures for a number of years)
| Spawning | Nursery I | Nursery II | Nursery III | Rearing |
| 1.2 | 2.2 | 2.8 | 4.0 | 5.0 |
The diurnal temperature variations in all the ponds investigated follow the same sinusoidal curve (Fig. 3), differing only in amplitude. This is greatest in spawning ponds and decreases with depth of pond (Table V).
Table V
Diurnal amplitude of temperature (°C) in the surface layers of various types of ponds (mean figures for a number of years)
| Spawning | Nursery I | Nursery II | Nursery III | Rearing | |
| Sunny days | 6.8 | 5.9 | 3.6 | 2.9 | 2.8 |
| All days | 4.6 | 3.9 | 2.1 | 1.9 | 1.9 |
In the investigated ponds the amplitude of diurnal temperature variation (A) × depth of reservoir (Z) is constant and the constant in the formula (AZ = constant) amounts to about 200.
Besides variations in diurnal amplitude there are also differences in the speed of temperature changes in individual ponds (Table VI), changes occurring most rapidly in spawning ponds (up to 30°/h) and least rapidly in rearing ponds (not exceeding 1°C/h). Temperature extremes appear somewhat later in larger ponds than spawning ponds, but not exceeding three hours later.
Table VI
Changes of temperature per time unit (°C/1h) in morning hours, sunny weather, in various types of ponds(mean figures for a number of years)
| Spawning | Nursery I | Nursery II | Nursery III | Rearing |
| 0.8 | 0.6 | 0.4 | 0.3 | 0.3 |
The influence of meteorological factors (such as overcast skies) is exerted most rapidly and distinctly on the thermo-dynamics of the spawning ponds. As the size of the pond and therefore its thermal capacity increases, this influence is proportionately greater and is observed only after a longer lapse of time.
These observations show that in the ponds used for carp rearing the time-space temperature distributions vary according to the size of pond, depth being the decisive factor. In the ponds investigated diurnal amplitude and vertical temperature gradient both change in inverse proportion to their depth. Depth differences also cause stratification. The surface area of the pond only influences the thermo-dynamics when the wind velocity is high, isothermy being observed in large ponds at velocities which are not sufficient for equalising temperatures in nursery or spawning ponds.
The temperature ranges indicate that nursery ponds have more favourable thermal conditions than the spawning ponds. In nursery ponds the temperature does not fall appreciably at dawn, as in spawning ponds, and the water warms up more gradually during the morning.
The thermal characteristics described here permit conclusions on the dynamics of carp ponds. On a windless sunny day, at about 0600 or 0700 hours, the surface layers of water begin to warm up and their density diminishes until the hour of maximum temperature. The water strata near the bottom are then cooler and heavier and so a stable equilibrium is established; convection currents are not observed. After exceeding the maximum temperature (after 1400 or 1500 hours) the direction of the warm current alters, the surface strata cool down and, being heavier, move to the bottom. This state of unstable equilibrium and the convection movements connected with it last throughout the night, up to the moment when the water temperature exceeds its minimum value. Vertical thermal sections of all investigated ponds indicate that convection movements affect the whole water column, as in all ponds isothermy is usually observed during the night.
The influence of wind being greater in them, larger movements are observed in rearing ponds. Turbulent mixing is also facilitated by smaller differences of density between the surface and bottom layers of the water in the ponds. In spawning ponds, as indicated by temperature measurements, mixing does not take place even with a considerable wind velocity. A similar influence of wind on the thermodynamics of shallow ponds has already been observed by Gieysztor (1934).
Geiger, R., 1942 Das Klima der bodennahen Luftschicht. Braunschweig.
Gieysztor, M., 1934 Limnologische Untersuchungen an einigen Kleingewässern. Arch.Hydrobiol.Ryb., 8:75–148
Kühlmann, D.H.H., 1961 Thermodynamische Untersuchungen in Kleingewässern verschiedener landschaftlicher Exposition. Verh.Int.Ver.Limnol., 15:87–94
Sedlmayer, K.A., 1934 Ein Beitrag zur Klimatologie des Teiches. Z.Fisch., 29:805–15
Starmach, K., 1956 Staw jako środowisko hodowlane. Hodowla ryb stawowych. Warszawa, PWRiL
Starmach, K., 1958 Hydrobiologiczno podstawy uzytkowania przez wodociagi wód plytkich zbiorników rzecznych. Polsk.Arch.Hydrobiol., 4(17):9–66
Szumiec, M., 1962 Czasowo-przestrzenny rozklad temperatury nad woda i gleba w najnizszych warstwach powietrza. Przegl.geofiz., 7(15):3, 157–68
Tuszko, A., 1952 Budowa stawów rybnych. Warszawa, PWRiL
![]()
![]()
![]()