![]()
![]()
![]()
Como ya se ha indicado, no es raro que los investigadores establezcan programas específicos sólo para observar la migración de los peces. La información disponible está diseminada en diversos textos y gran parte de ella es de segunda mano, notificando el escritor las observaciones transmitidas por los pescadores.
Rubião (1912) describió la “piracema” de diversas especies ícticas en el río Sapucaí en el estado de Minas Gerais, sin haberla visto. Aguirre (1936) describió la “piracema” en los rápidos de Pirapora (pira = pez; ora = abundante) en el río Sao Francisco en el estado de Minas Gerais, de octubre a diciembre, durante la estación de aguas altas. En este momento del año los peces migran aguas arriba para desovar. De nuevo, Aguirre (1938) describió peces marinos a considerable distancia aguas arriba del río Doce, en el estado de Espírito Santo.
Azevedo (1938) describió la migración de peces en el nordeste. Se refiere a la “curimata”, Prochilodus argentus, que espera para desovar que las aguas alcancen su más alto nivel, para pasar de los ríos a los embalses y viceversa; este pez puede subir por los rápidos y por las escaleras para peces. Azevedo y Vieira (1938) vinculan el éxito reproductivo de los peces a la cantidad de lluvia en el nordeste del Brasil.
Aguirre (1945) describió la caza y pesca en los pantanos del estado de Mato Grosso. Vinculó las migraciones ícticas reproductivas aguas arriba, de septiembre a febrero, con las precipitaciones. Localmente, la palabra “piracema” se convierte en “lufada”. Distintas especies ícticas migran en un orden establecido cuando desovan en febrero y marzo.
Kleerekoper (1949) describió la migración del “peixe-rei”, Odonthestes bonarienses, del mar al “Lagoa dos Quadros” en el estado de Río Grande do Sul, a partir de la segunda mitad de abril hasta el verano (diciembre). Estimó que esta migración es independiente de las condiciones meteorológicas, que, en su opinión, sólo afectan a las especies de río. Kleerekoper (1949) también manifestó que los vientos fuertes son especialmente favorables para la penetración del “peixe-rei” en los ríos.
Machado y Abreu (1952) manifiestan que la intensidad de pesca en el río Paraiba, estado de Sao Paulo, está inversamente relacionada con el régimen de precipitaciones porque el río transcurre con más lentitud en la estación seca. Los peces se capturan con más facilidad en las lagunas marginales durante este período.
Verissimo (1895), en su libro sobre pesca en el Amazonas, la describe como controlada por el régimen de los ríos, como consecuencia de que éstos afectan a los movimientos de los peces de los cuales las pesquerías dependen.
Geisler, Knöppel y Sioli, (1973) se ocuparon de la migración íctica en la cuenca del Amazonas, particularmente de Characidae. Saint Paul y Bayley (1979) se refieren a los movimientos ícticos laterales y longitudinales, que son importantes para las pesquerías en el Amazonas.
Goulding (1979) describió el complicado sistema de migración íctica en el río Madeira (estado de Rondonia), que también puede aplicarse a otros ríos de la cuenca del Amazonas con quizás algunas modificaciones locales.
En general, Goulding (1979) divide la migración íctica en dos tipos:
(i) Migraciones de desove mediante las cuales los peces van a los afluentes del río Madeira para desovar al principio de la temporada en la que las aguas alcanzan altos niveles. Aparentemente el desove tiene lugar cerca de las confluencias del río Madeira con sus afluentes y no hay pruebas de que los peces se trasladen en esta ocasión por el cauce principal. Después del desove, los peces vuelven aguas arriba en los afluentes. Goulding (1979) indicó que los huevos fertilizados y/o alevines se desplazan río abajo por la fuerte corriente del río Madeira. Estos peces se introducen en las llanuras anegadas de los tramos más bajos en donde crecen y por último migran hacia la parte superior del río compensando la mortalidad de los peces adultos en los afluentes de aguas arriba.
(ii) Las migraciones de “arribacão”, en las que participan los peces que proceden de los afluentes que desembocan en el Madeira, se dirigen hacia la parte superior del río para introducirse en otro afluente. Los peces, en esta ocasión, se conocen como “peces gordos” porque han pasado varios meses alimentándose. Esta clase de migración se considera como un complejo ecológico. El nivel y transparencia del agua pueden tener cierta influencia en el proceso.
Goulding (1979) también describió las características de la migración íctica de algunas especies comerciales importantes. La migración del “jatuarana”, Brycon spp., parece ser muy compleja, con siete fases distintas de migración en distintas épocas del año. En general, la migración del “curimatá”, Prochilodus nigricans, sigue la misma pauta anteriormente descrita y es una de las primeras especies que abandonan los bosques sin inundar cuando el nivel del agua comienza a descender (esto se produce dos o tres meses antes que cualquier otra especie íctica). También es distinta de muchas otras especies ícticas, porque puede superar los rápidos del río Madeira.
Goulding (1981) de acuerdo con Bonetto, Verón y Roldán (1981), describe las migraciones de bagre que se traslada cerca del fondo, haciendo mucho más difícil la detección de sus movimientos estacionales. Sólo dos familias de bagres, Pimelodidae y Doradidae, efectúan migraciones aguas arriba en los rápidos del río Madeira; las migraciones más largas son durante el período de bajo nivel del agua entre julio y noviembre. El “babao”, Goslinia platynema, es el único bagre conocido que migra durante el período en que el nivel de las aguas es alto. Goulding (1981) duda de que el desove sea la razón de estas migraciones aguas arriba.
Goulding y Carvalho (1982) describieron la migración del “tambaqui”, Colossoma macropomum, que es la especie íctica más importante descargada en el mercado de Manaus.
La secuencia de la migración es como sigue:
desove a principios del período de aguas altas;
después del desove se introducen en los “igapós” (bosques inundados) de todo tipo de aguas, negras o blancas;
después de las inundaciones migran a los canales del río con aguas blancas en donde buscan refugio en las trozas situadas en el cauce;
durante toda la estación seca permanecen en las trozas del cauce;
cuando el agua comienza a subir de nuevo se dirigen aguas arriba hasta el desove.
En la Figura 3 se describe este modelo de migración.
Ribeiro (1983) describió tres tipos de migración del “jaraqui”, Semaprochilodus spp., en el río Negro de Manaus: (i) reproductiva; (ii) trópica y (iii) de dispersión.
Cuando el nivel del agua comienza a subir, el “jaraqui” se desplaza corriente abajo en los afluentes del Solimoes-Amazonas para desovar en el río principal. Después del desove, las larvas se transportan hacia el río principal entrando en los lagos de las llanuras anegadas en donde crecen. Mientras tanto, el pez adulto migra aguas arriba en el mismo afluente y se dispersa a través de los bosques recientemente inundados en donde están casi tres meses alimentándose. Después de este período, estos “jeraquis” se dirigen hacia los afluentes de nuevo, y luego a los ríos principales en donde se desplazan aguas arriba hasta llegar y ascender a otro afluente para dispersarse por todos los hábitat de predesove, como son las orillas, lagos, pequeños ríos, etc. El ciclo se repite en la próxima inundación. Los peces jóvenes también dejan al final los viveros de los lagos de las llanuras anegadas para comenzar su desplazamiento aguas arriba. Este proceso tiende a producir un aumento del tamaño medio del pez aguas arriba.
Ribeiro (1983) indica que la migración de adultos y jóvenes en la parte superior de los ríos principales y el movimiento de las larvas hacia la parte inferior de ellos, contribuye a contrarrestar las pérdidas causadas por mortalidad natural (y pesca) de “jaraquis” en el ecosistema. Con ello también se refuerza la variabilidad genética manteniéndose el equilibrio energético del ecosistema escaso de nutrientes en donde vive esta especie.
No es extraño que se hayan visto tiburones en el Amazonas. Myers (1952) identificó un macho adulto de Carcarhinus (probablemente C. leucas) mediante una fotografía tomada de un ejemplar en Iquitos (4 300 km del mar) en 1943, y Thorson (1972) presenta pruebas verbales, informes de periódicos locales, descripciones fotográficas y fórmulas dentales de mandíbulas de tiburón recuperadas que confirman la presencia de C. leucas.

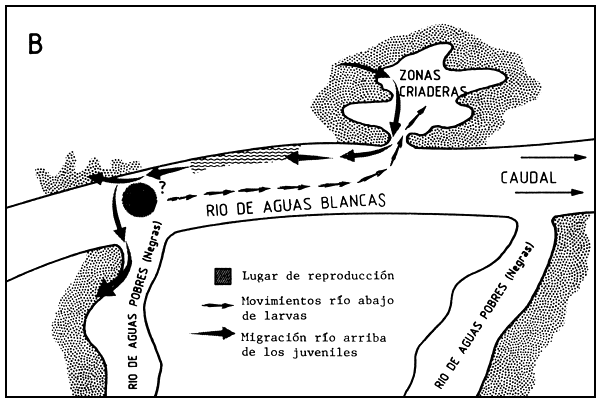
Figura 3 Modelo cartográfico de las migraciones de los adultos, juveniles y larvas de Colossoma macropomum en la cuenca amazonica centro-occidental. A. Se indica la migración de los adultos que dejan los planos inundados de la foresta migrando río arriba en un río de aguas claras hasta refugiarse en los cauces donde permanecen durante el período de agua baja. Cuando el agua comienza a subir de nuevo, se alejan de los cauces y migran río arriba hasta el desove; después de desovar entran en los planos inundados de la foresta donde se alimentan. B. Se indican los movimientos corriente abajo de las larvas inmediatamente después de eclosionar. Las larvas encuentran su camino en un área inundada de un río de aguas claras, donde permanecen por 5 ó 6 años (según hipótesis) antes de migrar en el río principal. No está claro si los juveniles de C. macropomum desovan durante el primer año de su migración o solo después (desde Goulding and Carvalho, 1982)
Thorson (1974) también se refiere a varios informes que indican la presencia del pez sierra Pristis perotetti a l 340 km aguas arriba de la desembocadura del Amazonas. También piensa que el P. pectinatus se introduce en el río, pero se translada a distancias mucho más cortas, y también menciona que en el Museo de Zoología de la Universidad de Sao Paulo hay un ejemplar de 116,5 cm de P. perotetti capturado en Oriximina en el río Trombetas.
Colombia: Miles (1947) describió dos “subiendas” (migraciones aguas arriba) y dos “bajadas” (migraciones aguas abajo) en el río Magdalena, y cada una tuvo lugar aproximadamente en una de las cuatro estaciones hidrológicas. La principal subida se produce entre febrero y marzo y a ella sigue la “bajada” de abril a junio. La segunda “subienda” es menos intensa que la primera y se conoce como “mitaca”1; se produce en julio–septiembre y la “bajada” final tiene lugar en octubre–diciembre. Miles(1947) también se refiere a invasiones de tiburones en el río Magdalena.
Dahl (1963) describió la migración del “bocachico”, Prochilodus reticulatus magdalenae, en los ríos San Jorge y Uré. Cuando el agua sube en el verano, a finales de enero, el “bocachico” se introduce en las ciénagas que son sus criaderos naturales. Después, abandonan estas lagunas y van arriba del río durante un breve período. Cuando vuelven, tiene lugar la reproducción después de la “candelada”. La migración aguas arriba es seguida por una serie de depredadores, y es de tipo irregular, porque los peces se detienen para comer.
Parece ser que los ejemplares más pequeños son los primeros que comienzan a migrar aguas arriba. Aparentemente no todos los peces van en este sentido porque se encuentran siempre algunos en las ciénagas situadas en la parte inferior de los ríos. Los ejemplares más viejos tienen un comportamiento de retorno al lugar de procedencia. La gran mayoría de las hembras descienden por el río durante la “bajada”, pero algunas permanecen en la parte superior. En mayo, tiene lugar el desove de “bajança”. Dahl (1963) indica que el desove tiene lugar en el medio ambiente más tranquilo de las ciénagas y afluentes, como consecuencia de la protección ofrecida por la vegetación acuática y también porque las larvas se alimentan del zooplancton abundante en este medio ambiente.
Venezuela: Mago Leccia (1970) describió migración íctica en los “llanos” de Venezuela. El comienzo de la estación lluviosa (mayo–junio) desencadena la migración de varias especies ícticas, particularmente las masivas de Prochilodus spp., aguas arriba a fines reproductores. Mago Leccia (1970) también parece aceptar una hipótesis fisiológica similar, como la que sugiere Godoy (1967), para explicar la migración íctica aguas arriba. Mago Leccia (1970) también menciona la mortalidad íctica catastrófica de ejemplares atrapados en las lagunas y canales de ríos en la estación seca, que es un fenómeno común en los trópicos.
Novoa y Ramos (1978) se refieren a las grandes migraciones de “zapoara”, Semaprochilodus laticeps, durante julio–agosto, que permiten la captura del pescado en grandes cantidades en el río Orinoco (Figura 4).
Novoa y Ramos (1982a) describen la migración íctica a fines reproductivos para una serie de especies del río Orinoco. No observaron la migración íctica aguas abajo, aunque algunos pescadores dicen que se produce.
El río Orinoco presenta su más bajo nivel en febrero–marzo, cuando muchas especies ícticas van aguas arriba, notablemente el “morocoto”, Colossoma brachypomum, cuyo movimiento migratorio es discontinuo, ya que los peces se detienen para alimentarse y descansar. Novoa y Ramos (1982) también piensan que los niveles de luz pueden estar implicados en este proceso migratorio.
En julio–septiembre, cuando el nivel de las aguas es más alto, se observan numerosos juveniles de varias especies en el canal del río, desde el cual migran a los lagos de las llanuras anegadas en donde permanecen durante el próximo verano. También son igualmente importantes lo movimientos laterales y longitudinales en el ciclo vital del pez. En Novoa y Ramos (1982) se pueden encontrar descripciones más interesantes mereciendo los cuadros 1–4 especial atención:
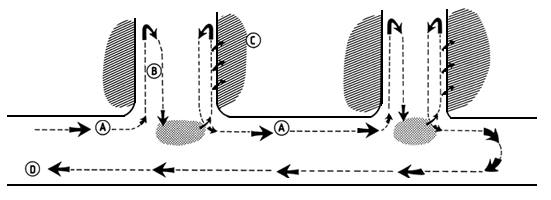
Figura 4 Secuencia de migración de “Piracema” en el río Madeira y sus tributarios (desde Golding, 1980) y en el río Negro y
tributarios (Ribeiro, 1983)
A. Migración río arriba de stocks de peces adultos cuando el agua es baja (Piracema)
B. Migración de desove río abajo y cuando el agua es alta
 Lugares hipotéticos de desove
Lugares hipotéticos de desove
C. Migración alimentaria río arriba con dispersión en las zonas inundadas de la foresta durante la inundación
D. Movimientos hipotéticos río abajo y movimientos de los huevos, larvas y juveniles

Figura 5 Esquema ilustrativo del movimiento migratorio de la Zapoara y su relación con la distribución de tallas y proceso de desove a lo largo del río Orinoco deducido de programas de muestreo biológico y de marcaje
Guatemala: Thorson, Watson y Cowan (1966a) informan de invasiones marinas de peces espada, Pristis perotetti, y tiburones, Carcarhinus leucas en el sistema del lago Izabal-río Dulce, en Guatemala.
Guyana: Lowe-McConnell (1964) presentó una descripción extensiva de la fauna íctica de la sabana Rupununi y también describió los movimientos ícticos durante la crecida y bajada del agua. La migración comienza en un orden establecido durante las inundaciones. Cuando el agua vuelve a su estado primitivo aparecen distintas especies que se mueven aguas abajo en momentos determinados del día.
![]()
![]()
![]()