![]()
![]()
![]()
J. J. HAVEL
J. J. HAVEL es Director de la Escuela de Montes de Bulolo (Nueva Guinea). W. Finlayson prestó su ayuda leyendo y corrigiendo el manuscrito.
SE DICE, con cierta verosimilitud, que el monte higrofítico tropical es el sueño del botánico, pero la pesadilla del forestal. La excesiva riqueza de especies obstaculiza la evaluación de los recursos, el aprovechamiento inicial del monte y su consiguiente regeneración y tratamiento. La simple evaluación de un sitio de prueba se convierte en una compleja empresa taxonómica, pero la evaluación volumétrica sin la identificación de especies no basta para indicar la probable cosecha. La corta y elaboración de un producto tan variado, y especialmente su comercialización, presentan dificultades casi insuperables, con el resultado de que generalmente sólo se explota una pequeña proporción del bosque. La regeneración de las masas cortadas suele ser a veces un enigma, en apariencia sin solución, que frecuentemente induce a los forestales a las cortas rasas y a la replantación con especies exóticas.
La situación dasonómica, a menudo insatisfactoria en los países tropicales, es, al menos en parte, atribuible a la incapacidad de los forestales de identificar de manera fidedigna incluso una proporción ínfima de todas las especies. La mayoría de los forestales profesionales se han formado en países de la zona templada y antes de graduarse no han tenido ocasión de enfrentarse con el monte higrofítico tropical. Para agravar más esta situación, en los países donde generalmente trabajan no existen publicadas floras completas. El empleo de nombres vernáculos y la utilización de los lugareños como señaladores de árboles no dejan de presentar dificultades. En muchas zonas, particularmente en Nueva Guinea, la fragmentación lingüística es tal que el ajuste de los nombres tribales con los científicos es tan difícil como la propia identificación sobre el terreno. Además, los conceptos científicos y tribales no coinciden siempre. En el caso de árboles de importancia económica para los indígenas, existen nombres distintos para variedades casi indiferenciables, mientras que especies e incluso géneros sin valor económico están englobados bajo un nombre común, como por ejemplo el término malayo medang. Sin embargo, la causa principal de las dificultades estriba en la magnitud misma del problema.
En contraste con los montes de las zonas templadas o subtropicales, el bosque higrofítico tropical es sumamente rico en especies y familias. Excepto en condiciones especiales de clima y suelo, no hay especie de importancia abrumadora, ya sea ecológica o económicamente. El número de especies arbóreas de Malaya ha sido estimado por Menon (1961) en 2.000 a 2.500; las estimaciones referentes a Nueva Guinea son incluso mayores. Pertenecen a una grandísima diversidad de familias, entre ellas muchas que en los países templados están formadas principalmente de especies herbáceas, como por ejemplo, las datiscáceas.
Como consecuencia, salvo la estructura de la flor, no hay caracteres suficientemente variados para servir de base en la identificación. Menon ha estudiado la cuestión en Malaya y llegó a la conclusión de que los tejidos leñosos son demasiado simples para que la identificación pueda abarcar hasta la especie, y en muchos casos es incluso imposible separar géneros próximos. Endert (1956) ha creado una clave aplicable a los géneros de árboles de Indonesia, que se basa sólo en los órganos vegetativos, pero exige un examen morfológico y anatómico de los ramos tan detallado que su empleo queda restringido a los botánicos profesionales.
Si bien últimamente varios botánicos (Wood, 1952; Wyatt-Smith, 1954; Whitmore, 1963) han pensado utilizar la corteza para identificar árboles, este método en sí no permite una identificación exacta y fidedigna de todas las especies.
Es evidente que un forestal, para quien la botánica taxonómica no es sino una de las innumerables esferas de su trabajo, no puede pretender conocer cabalmente todas las especies del bosque higrofítico con las que ha de enfrentarse. No obstante, paradójicamente, los forestales sienten preferencia por las especies individuales. Existen muchas razones para ello. Muchas características que utiliza el forestal para la identificación, como la corteza externa o la forma de un árbol, suelen ser propias de especies individuales. Es a menudo más fácil identificar una especie determinada que captar el concepto de todo un género. Dos especies pertenecientes al mismo género difieren con frecuencia notablemente en cuanto a su valor económico.
En cambio, los botánicos se inclinan más al estudio de las familias por razones igualmente válidas. En muchas de las familias comunes, como las lauráceas y las meliáceas, la clasificación y nomenclatura están actual mente muy lejos de ser completas y la identificación válida de las especies resulta difícil, si no imposible.
Los forestales y los botánicos también parecen estar divididos en cuanto a los medios de identificación. Los forestales generalmente evitan el empleo de material fértil, argumentando que las flores y los frutos de los árboles forestales grandes son virtualmente inaccesibles y, en muchos casos, disponibles sólo estacionalmente. Prefieren basarse en el aspecto general de los árboles y en la estructura de su corteza y leño, a los que puede accederse fácilmente desde el suelo, suplementando el examen con características de las hojas caídas. En cambio, los botánicos tienden a basarse en las características perceptibles en los ejemplares fértiles de un herbario, es decir, hojas, flores y frutos. Con frecuencia no reconocen en condiciones de campo una especie que han manipulado repetidamente en el herbario. No se puede decidir cuál de los dos está equivocado - lo están ambos. Los frutos y flores caídos son características más útiles, frecuentes y fidedignas de lo que los forestales están dispuestos a admitir. En cambio, las objeciones de los botánicos sobre el empleo de las características vegetativas pierden su peso después de haberse establecido una correlación fidedigna entre el material fértil y la corteza y leño.
Sin embargo, queda el hecho de que la clasificación y la nomenclatura científicas se basan en ejemplares fértiles, y de que una identificación positiva inicial sólo es posible sobre esa base. En palabras sencillas, los forestales bailan al son de la gaita de los botánicos. Después de la identificación inicial positiva, no sólo son posibles las identificaciones subsiguientes basadas en una correlación entre las características reproductoras y las vegetativas, sino que en la práctica ordinaria resultan mucho más eficaces.
Uno de los motivos de que no se haga un mayor empleo de tales correlaciones es que no siempre son sencillas. Es posible que géneros estrechamente relacionados en cuanto a la estructura floral difieran mucho en la leñosa o, al contrario, que exista una similitud anatómica entre géneros que, por otra parte, no guardan la menor relación entre sí (Menon, 1961). Esto, al fin y al cabo, no es una novedad para los taxonomistas: la identificación y la clasificación no se basan, y en realidad no pueden basarse, en un solo carácter, sino más bien en una combinación de caracteres más o menos compleja.
Cuanto menos comunes y más llamativos sean los caracteres se necesitará un número menor de ellos. A medida que se pasa de la especie al género y a la familia, la combinación de caracteres va volviéndose cada vez más importante, aunque también más difícil de captar, y ésta es la principal razón por la cual se recurre a conceptos más amplios, más generales y por tanto definidos menos claramente.
Aquí tocamos el punto crucial del problema: individualmente las especies comprendidas en los montes de los países tropicales son demasiado numerosas para ser tratadas como entidades distintas y no relacionadas. Existe, por tanto, la necesidad de una generalización utilizando los conceptos de género y familia.
Dicha generalización facilita la retención del conocimiento adquirido, proporcionando un marco en que los datos pueden integrarse significativamente. También reduce el número de hechos que hay que aprender de memoria, puesto que muchos de los caracteres son comunes a todos los miembros de la familia. Por último, permite ampliar posteriormente nuestros conocimientos, ya que todo árbol que se encuentra por vez primera puede ser asignado a una familia y seguidamente identificado mediante una clave para determinar los géneros y especies de la familia. Sin embargo, antes de poder hacer esto hay que establecer el concepto difícil y a veces evasivo de una familia.
Pretender exclusivamente alcanzar esta fase avanzada, puede ser demasiado arriesgado. Sin embargo, cuando se utilizan especies importantes desde el punto de vista ecológico o económico para establecer el concepto, se adquiere pericia en la identificación, aun cuando la fase más avanzada de la formación del concepto pueda resultar demasiado difícil.
La formación de los conceptos de género y familia es una tarea ardua, que exige un empeño a fondo. Se sugieren dos principios. Primeramente, los árboles deben tratarse como un todo, y no simplemente como piezas de madera cubiertas por corteza, o ramos pegados en una hoja de un herbario. Wyatt Smith (1954) nos da un ejemplo excelente del empleo de una amplia gama de caracteres en la identificación de árboles. En segundo lugar, la enseñanza o el aprendizaje de los conceptos debe ajustarse a los descubrimientos de la psicología pedagógica en este campo.
Existen dos medios de establecer el concepto de una familia; de éstos, el enfoque formal, deductivo es el utilizado más comúnmente, especialmente en los libros de texto. Consiste en la enumeración de los caracteres de la familia, desglosándolos mediante una clave, y describiendo los géneros y especies que la componen. Al parecer, esto es sencillo y metódico. Su principal inconveniente es que pocas personas pueden hacerse una imagen clara de la familia partiendo de la definición, ya que los caracteres definidores rara vez son tajantes. No es raro que dentro de un género de una familia se encuentren hojas alternas, opuestas y verticiladas, o cuatro, cinco o seis pétalos. Se trata pues de un proceso de razonamiento incompleto, adecuado quizás para el especialista, pero inapropiado a un nivel inferior.
El otro método aplicable es el inductivo. Consiste en estudiar un grupo de árboles que se sabe pertenecen a una determinada familia, y llegar, basándose en la observación y el razonamiento, al concepto de la familia. Este es un enfoque natural, posible incluso cuando es difícil una descripción exacta de los caracteres. Pueden utilizarlo, y así lo hacen, personas sin una educación formal; por ejemplo, son pocos los estudiantes de Nueva Guinea que no aciertan a colocar Viticipremnanovae pomeraniae después de Vitex cofassus, a pesar de que estas dos especies difieren en que tienen hojas palmaticompuestas y sencillas respectivamente. Muchos reconocen al género Tectona introducido como «hermano» del género indígena Gmelina; algunos llegan a incluir los cuatro géneros citados en un «clan», aunque jamás hayan oído hablar de la familia de las verbenáceas.
El desarrollo inductivo de un concepto puede desglosarse en cinco fases:
a) observación de los diversos objetos,
b) formación de la hipótesis acerca de sus elementos comunes,
c) prueba de la hipótesis en casos específicos,
d) selección de una categoría general que conviene a todos los objetos,
e) dar un nombre a la categoría.
En el estudio de la taxonomía hay que seguir esta misma secuencia. Se llegará al concepto de la familia más rápidamente cuando un profesor pueda ayudar al alumno, pero con una capacitación básica suficiente y un libro de consulta adecuado, éste puede emprender sólo la tarea. Seguidamente se hacen algunos comentarios sobre la aplicación del razonamiento anterior.
OBSERVACIÓN DE ÁRBOLES QUE SE SABE PERTENECEN A UNA FAMILIA
Las instrucciones acerca de cómo hay que observar los árboles pueden parecer superfluas, pero en realidad no lo son. Hay que enseñar al principiante a reconocer los caracteres importantes; por ejemplo, muchas leguminosas tienen flores espectaculares, cuyo color atrae inmediatamente la mirada y, sin embargo, los colores tienen poca importancia. Lo que importa es la disposición de las partes florales, el tipo de fruto y la naturaleza del leño. Los innumerables caracteres observables en todo árbol pueden dividirse en tres categorías principales:
a) caracteres principales útiles para la identificación del árbol como miembro de una familia o género particulares;b) caracteres notables útiles para la identificación de la especie de que se trate;
c) caracteres comunes útiles sólo para la confirmación de la identificación preliminar.
Dicha observación fragmentaria debe, sin embargo, compensarse por la percepción de todo el árbol como entidad, ya que el conjunto es más de una simple suma de sus partes componentes. Es posible identificar a simple vista los miembros de las familias más claramente definidas, como las miristicáceas, sin necesidad de comprobar mentalmente los caracteres de la familia. El carácter en que se basa dicha identificación - la inclinación de las ramas, la disposición de las hojas, la naturaleza de la corteza - son difíciles de describir. Por esta razón, el reconocimiento a simple vista sólo se logra gracias a la experiencia, reduciendo gradualmente el número de indicios necesarios para la identificación positiva y haciendo más hincapié en un carácter detallado y definido que en una impresión general. Para dicho estudio, naturalmente, son indispensables un buen herbario y una colección de muestras de madera, así como el fácil acceso al bosque, que resulta menos difícil cuando se trata de instituciones de capacitación a nivel inferior que en el caso de universidades.
El material del herbario, las muestras de madera y de árboles deberían observarse en una estrecha proximidad espacial y temporal. Si se ha reservado una tarde para el estudio de una determinada familia, hay que prescindir de los miembros de otras familias, aunque se encuentren a proximidad muy conveniente. La claridad del concepto adquirido de esta manera justifica sobradamente el esfuerzo suplementario. Debido a que la identificación rápida sólo se consigue mediante observaciones repetidas, vale asimismo la pena crear un paseo botánico, con cada árbol identificado con una etiqueta en la que conste su nombre.
FORMACIÓN DE LA HIPÓTESIS
En esta fase, más aún que en la primera, el profesor puede ser de gran ayuda señalando la atención sobre los caracteres comunes de la familia. Antes de poder hacerlo, los estudiantes necesitarán conocer la terminología taxonómica fundamental. Los términos empleados deben abarcar el árbol entero y no limitarse a los caracteres reproductivos, pero no merece la pena una precisión y detalle excesivos. Es mejor descartar los caracteres que no se perciban fácilmente a simple vista o con lupas de 10 aumentos, ya que son poco útiles para la identificación sobre el terreno. Merece la pena observar que muchos de los términos utilizados en taxonomía y anatomía de la madera son ya de por sí conceptos bastante complejos que describen la relación mutua entre las partes componentes, como epiginia en las flores y parénquima confluente en la madera. Como tales son pasos útiles en el desarrollo del concepto de familia.
Cuando se utilizan libros de texto, el paralelismo de las descripciones e ilustraciones ayuda a poner de relieve las analogías. Como la formación del concepto implica la combinación de muchas experiencias concretas en una abstracción generalizada, el empleo de ayudas tales como fórmulas o diagramas floreales aceleran el proceso.
Durante el estudio de la familia y en los sucesivos encuentros con ella, conviene proceder a una revisión continua del concepto, que permita la fusión de las percepciones anteriores con las nuevas observaciones. El nuevo concepto resultante será más amplio o más exacto que el anterior, y dependerá de la selección de los ejemplos anteriores. El profesor podrá justificadamente, dada su mayor experiencia, evitar ejemplos que conduzcan a un concepto innecesariamente amplio y general. En la mayoría de las familias, el estudio de los árboles madereros de un país determinado llevará a un concepto más estrecho y exacto que el de las especies herbáceas y arbóreas del mundo entero.
El concepto de familia es más fácil de desarrollar en algunas familias que en otras. Las familias pequeñas y compactas, como las miristicáceas, son mucho más fáciles de agrupar que las más amplias y variables, como las euforbiáceas.
PRUEBA DE LAS HIPÓTESIS
Esta es una fase importante, ya que no sólo permite una comprobación de la eficacia del estudio anterior, sino que estimula la participación activa de los alumnos. Surgen espontáneamente dos posibles enfoques: la omisión de una especie importante desde la formación inicial del concepto, o bien el empleo de una especie secundaria relativamente poco importante. En ambos casos, la especie debe introducirse casualmente de forma que los alumnos puedan tratar de situarla sin ayuda alguna. El descartar los caracteres que sólo tengan un valor específico ayuda a aclarar el concepto de familia.
DEFINICIÓN Y NOMBRE DE LA FAMILIA
Los estudiantes pueden intentar definir la familia, pero en general necesitarán la ayuda del profesor. En esta etapa, el nombre de la familia puede añadirse al concepto. Esto puede parecer un enfoque de atrás a adelante, pero sin desarrollar previamente el concepto, un nombre no es sino una cosa hueca y vacía.
Anteriormente se ha hecho mención del empleo del concepto de familia como medio mnemotécnico, ya que los hechos resultan con ello más significativos. Merece la pena considerar si el mismo principio se aplica a unidades de clasificación más grandes, como los órdenes. En casi todos los países, los herbarios utilizan el sistema Engler y Prantl, útil porque agrupa a las familias de acuerdo con sus similaridades. Por ejemplo, en la clase de las dicotiledóneas, las familias progresan desde las carentes de perianto, pasando por las de perianto de una serie, las de perianto de dos series helicoidales hasta las de disposición cíclica en dos series, primero las que tienen los segmentos de la serie interna distintos y por último las que los tienen fundidos. Tal disposición proporciona un nuevo marco más amplio para la debida ordenación de los datos. Existen también series paralelas, pero menos constantes, desde la apocárpica a la sincárpica y de la hipógina pasando por la perígina a la epígina. Dado el gran volumen de datos que se necesita recordar, merece la pena ensayar todo enfoque que sea mnemotécnico.
FASES FINALES
Durante el establecimiento del concepto familia, es conveniente facilitar el paso a la segunda etapa de enseñanza en que el estudiante ha de trabajar solo. Por lo tanto, la terminología utilizada inicialmente debe seguir lo más posible las obras de consulta de las que el estudiante dependerá en definitiva. En el caso de la región de Malasia, éstas son sin lugar a dudas la Flora Malesiana y la carta clave de la Commonwealth Scientific and Industrial Research Organization (CSIRO) para identificar las maderas de la región del Pacífico suroccidental. El principal problema, en este aspecto, es la gran variedad de términos técnicos utilizados por los botánicos profesionales.
Como paso final habría que enseñar a los estudiantes un método adecuado para la identificación de especies que encuentren por primera vez. Uno de estos métodos, que el autor utiliza, es el siguiente: Si se puede incluir la especie desconocida, basándose en una intuición inicial, en un género o familia determinados, esta «intuición» debe inmediatamente ser comprobada comparando los caracteres de la especie desconocida con los de las especies conocidas del género o familia. Si no existe inmediata «intuición», se anotan los caracteres de la especie desconocida y se comparan con los conocidos de las diversas familias. A menudo la simple enumeración conduce a ver más claro el problema. Si entonces puede hacerse una identificación aproximada, debe hacerse otra vez una comprobación a fondo. Cuando se dispone de un herbario, no hay mejor comprobación que la directa con los ejemplares reales; en los demás casos, deben bastar los libros de consulta. Una parte integral de la segunda fase es el empleo de claves taxonómicas. Como éste es un tema que tratan detalladamente la mayoría de los libros de texto, no insistiremos sobre él.
En conclusión, cabe dudar de la factibilidad del procedimiento sugerido. El autor comprobó que algunos cursillistas que demostraban interés y eran inteligentes, con una educación de escuela secundaria elemental, después de estudiar durante un año 150 de las especies más importantes de Nueva Guinea, podían incluir correctamente muchas especies vistas por vez primera en una familia o género determinados. Otros eran capaces de reconocer las especies que se les había enseñado realmente y no podían captar el concepto de familia. Incluso cuando se proporciona la identificación, no siempre ésta es correcta y se aconseja el envío de un ejemplar de un herbario para confirmación. Lo principal es que se ha sentado una base y se ha establecido una estructura, lo que permite nuevos progresos. Cuando se trata de estudiantes de educación más avanzada, es lógico prever resultados mejores.
ENDERT, F.H. 1956. Key to the tree genera of Indonesia. Traducción al inglés por R.D. Hoogland, CSIRO, Canberra.
KRECH, D. y CRUTCHFIELD, R. S. 1959. Elements of psychology. Knopf, New York
LAWRENCE, G.H.M. 1951. Taxonomy of vascular plants. Macmillan, New York..
MENON, P.K.B. 1961. Taxonomic value of wood anatomy seen through Malayan woods. Malayan Forester, 24: 290-301.
MERRILL, E.D. 1954. Plant life of the Pacific. New York, Macmillan.
WHITMORE, T.C. 1963. Bart, morphology as an aid to forest recognition and taxonomy of Dipterocarpaceae. Flora Malesiana Bulletin, 18: 1017-1019.
WOOD, G.H.S. 1952. Bark as means of tree identification. Journal of the Oxford University Forest Society. Ser. 3, 6: 15-27.
WYATT SMITH, J. 1954. Suggested definitions of field characteristics. Malayan Forester, 17: 170-183.
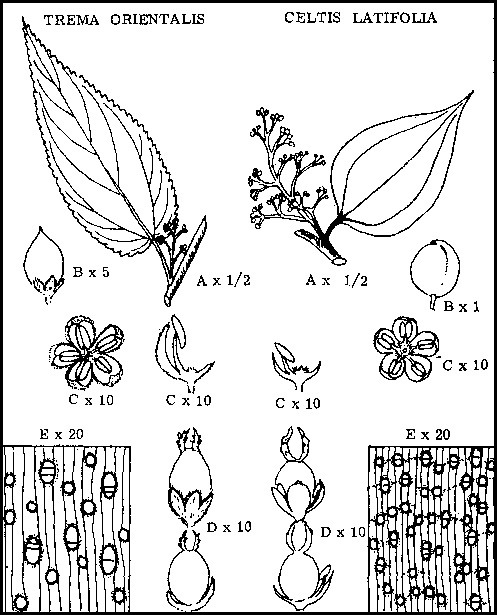
Existen otros dos géneros poco en Nueva Guinea que se parecen a Trema, que juntamente con Celtis pertenecen a la familia de las ulmáceas. Los caracteres de la familia, observados sobre Celtis y Trema son:
Hojas simples, alternadas, con margen entero o serrado.
Arboles monoicos, flores, en su mayoría unisexuales, consistentes en periantos de 4 a 6 lóbulos con igual número de estambres en las flores masculinas, y con un simple ovario do una célula con dos estilos en las flores femeninas.
El fruto consiste casi siempre en drupas pequeñas, de ligera capa carnosa externa y cáscara interna dura.
Las maderas tienen radios estrechos, poros de tamaño pequeño a mediano en pequeños múltiplos y parénquima rodeando los poros.
Obsérvese que en las siguientes descripciones más detalladas de Celtis latifolia y Trema orientalis el subrayado continuo se refiere a los caracteres específicos importantes, mientras que el espaciado indica los caracteres de la familia.
CELTIS LATIFOLIA PLANCH.
|
Porte |
Arbol grande hasta 140 pies (42 m) de altura y 3 pies (1 m) de diámetro. Tronco con costillas basales, largo, esbelto, ahusado, copa de anchura mediana. |
|
Corteza |
½ pulgada (1,27 cm) de espesor. Tez color gris, lisa salvo pústulas y algunas acanaladuras salientes horizontales. Corteza intermedia color verde, yema entre crema y pajizo, con un preponderante aspecto pardo-rojo «sal y pimienta», dura y no fibrosa. Cortada, oscurece rápidamente. |
|
Madera |
Color entre aroma y paja, moderadamente dura y posada, radios medulares estrechos, poros de tamaño mediano, relativamente numerosos dispuestos en radios cortos múltiples, parénquima rodeando los poros, en forma de ala y confluentes. |
|
Hojas |
Simples, alternadas, ovadas, a menudo asimétricas, de 1 ½ x 3 pulgadas (3,8 x 7,62 cm), ápice acuminado, base redonda, margen entero, multinerviales, ternadas, de espesor mediano, correosas, haz satinado verde oscuro, envés glabro verde claro. |
|
Flores |
En panículas axilares o terminales, pequeñas, verde amarillo, perfume tenue. Las flores unisexuales y monoicas o aparentemente bisexuales. Las flores masculinas consisten en cinco segmentos periánticos, con un estambre opuesto a cada segmento Las flores femeninas también con cinco segmentos periánticos, con un ovario grande ovoide con dos estilos y estambres poco desarrollados. |
|
Fruto |
En drupa globosa con ápice puntiagudo y poca carne, negro-azul brillante cuando está maduro. |
|
Distribución |
Montes higrofíticos de las tierras bajas por todo el territorio, asociada con las diversas especies frondosas. |
|
Otras especies |
Existen seis especies de Celtis en el territorio, que difieren principalmente en cuanto al tamaño y forma de las hojas. En dos de ellas, la nervadura no es marcadamente tornada. La identificación en el arbusto se basa en el aspecto «sal y pimienta» de la corteza, en la estructura de la madera y, en muchos casos, en la nervadura tornada. |
TREMA ORIENTALIS (Lino) Bl.
|
Porte |
Arbol de tamaño pequeño a medio, hasta 70 pies (21 m) de altura y 15 pulgadas (38 cm) de diámetro. Fuste espolonada y recto; copa entre mediana y amplia, follaje ralo. |
|
Corteza |
De ¼ a 1/3 de pulgada (0,63 a 0,84 cm) de espesor; tez suave de color gris; yema color crema verdoso, muy fibrosa que se utiliza como atadura. |
|
Madera |
Blanda y ligera, color paja, fibra basta, poros de tamaño medio dispuestos en cadena aorta, radios medulares estrechos, ondulantes, parénquima rodeando los poros. |
|
Hojas |
Simples, alternas, dísticas, ovadas, de ápice acuminado, base cordada y margen serrado, uninervias pero con dos venas inferiores fuertemente desarrolladas, de espesor mediano, verdes, el haz ligeramente velloso, el envés plateado, densamente velloso. |
|
Flores |
Dispuestas en panículas axilares, flores diminutas color verde amarillento, unixesuales, pies monoicos. Flores masculinas consistentes en un perianto pentalobulado con un estambre opuesto a cada lóbulo. Flores femeninas consistentes en un perianto pentalobulado con un ovario ancho con dos estilos. |
|
Fruto |
Pequeño, 1/8 de pulgada (0,32 cm) de diámetro, ovoide, drupa negra con una ligera capa carnosa exterior y cáscara interior dura, con lóbulos periánticos persistentes. |
|
Otras |
especies Existen otras dos especies de Trema en el territorio, que difieren principalmente en cuanto a vellosidad de las hojas. |
|
Distribución |
Las tres especies de Trema se dan en toda Nueva Guinea, en las tierras bajas y en las montañas, hasta una altitud de 5.000 pies (1.500 m) aproximadamente, son muy comunes en los antiguos terrenos de huerta. |
Trema orientalis - Celtis latifolia
![]()
![]()
![]()