![]()
![]()
![]()
Fish populations in reservoirs usually increase rapidly in numbers and mass after filling; thereafter fluctuations occur seasonally or from year to year. The latter fluctuations are influenced by the biology of the species and the vagaries of the environment. Therefore, knowledge of these changes is necessary in order to improve long-term forecasts, to plan catches, and to develop rational ways of fishing and to conserving the stocks.
The formation of fish populations in reservoirs is basically from the parental river and lake populations of the inundated river and watershed areas. The formation of the fish stocks is usually characterized by strengthening of the lacustrine species and the suppression or complete disappearance of the riverine species. A decline of riverine species is a result of changes in environmental conditions that do not favour them. For instance, the presence of the dam blocks their way downstream for feeding and spawning, and sedimentation reduces food supply and spawning grounds for benthophagous fish species. On the other hand, newly changed conditions are favourable to limnophilic species by providing good growth rates, successful reproduction and high survival rates. Thus the fish increase both in numbers of individuals and total biomass (Tomnatik, 1957; Kimsey, 1958; Zhadin and Gerd, 1961; Rzóska, 1966; Denyoh, 1969; Fitz, 1968; Beckman and Elrod, 1971; Evans and Vanderpuye, 1973; Balon and Coche, 1974; Petr, 1975a; Bhukaswan and Pholprasith, 1977, etc.).
The development of fish populations in temperate zone reservoirs has been divided into three stages (Sharonov, 1966) as follows:
A rapid rise in biomass due to an increase in growth rate and improved nourishment;
A sharp drop in growth of rheophilic species due to unfavourable environmental conditions including shortage in the food supply for benthophagous fishes;
An alteration of weak and strong year-classes, the stabilization of growth, and an increase in biomass of low-value and undesired fishes.
Sharonov described the first stage as being connected with the filling period. The sharp rise in biomass and growth rate was a consequence of better reproduction and feeding as well as high survival rate of young fish. The second stage resulted from a decline of benthos biomass and disappearance of the spawning grounds, which in subsequent years exerted an adverse effect on the abundance. The third stage presumed that feeding conditions had become relatively stable.
The establishment of fish population in reservoirs may take several years, depending primarily on how long it takes for individual species to sexually mature. Poddubnyi (1963) suggested that in the Soviet reservoirs the new fish stock will become established when the breeding stock consists of fish originating in the reservoir itself rather than from the original river. He cited examples of fish fauna development in the Ivankovo reservoir which stabilized in the ninth year after the filling; that of the Uglich reservoir between the eighth and ninth year; and that of the Rybinsk reservoir after 11–12 years. Patriarche and Campbell (1958) found that all species of the Clearwater reservoir in Missouri (USA) succeeded in spawning in the fifth year of the existence of the reservoir. In the tropics where the life-spans of many fish are short, the fish population will establish itself within a few years after the impoundment is formed (Balon and Coche, 1974; Petr, 1975a).
The year-class strength of fishes in the early stage of impoundment is not only influenced by the fertility of water and inundated soils, food availability, and water-level fluctuation; it is also dependent upon the period of filling. Poddubnyi (1963) stated that, if the temperate region reservoirs are flooded to the designated mark in one season, there would appear only one strong year-class of fish to live under initially favourable conditions. If the period of filling was extended over a few years and each year some parts of land were flooded which hitherto had not been inundated, then favourable conditions for a number of fish species would be maintained. During these years, the year-class of the young are abundant. Therefore, during the first few years, the fish stocks will develop in this type of reservoir better than in reservoirs that are filled in a single year. For example, the Ivan'kovo reservoir was filled in one spring and there was only one very abundant year-class of bream. The Gorki, Kuibyshev and Uglich reservoirs reached the full level after two delayed spring floods, and the bream year-class of the first two years was abundant. Il'ina and Gordeyev (1970) reported that the filling period of the Rybinsk reservoir took seven years, and during these years the newly flooded areas were increased every year, which supplied additional nutrients to impounded water, encouraging growth and reproduction of the fishes. Therefore, the generations of these years were abundant and ensured maximum catches over a number of years. Similar phenomena were found in Lake Oahe in the United States. Beckman and Elrod (1971) reported that the largest year-class of northern pike, carp, small-mouth buffalo, and big-mouth buffalo was associated with the rise and maintaining of high water level in the spring flooding of terrestrial vegetation (primarily grasses). Small year-classes occurred in years when no new terrestrial vegetation was flooded.
In the tropics, strong year-classes of all species also coincide closely with the filling period, which usually takes one to several years. The gradual flooding of new land over a number of years has the same impact on fish biological cycle and production as the annual formation of floodplains (Petr, 1975a).
A change in species composition of fish in a reservoir is the result of species reacting differently to the alteration of living conditions after impoundment. Some species whose life cycles have been particularly disrupted may be unable to adapt to the new conditions and may disappear from the population. But many species can adapt to those changes in varying degrees and may continue to exist at changed abundance. The fluctuations may range from a capability of some species to maintain their populations at about the same level when compared with the pre-impoundment period, to an increase in their numbers as a result of favourable environmental changes. There are several reasons for fewer fish species occurring in a reservoir than in the former riverine conditions. They include: (1) reservoirs have fewer habitat types than rivers with respect to current; (2) a reduction of turbidity in reservoirs due to sedimentation probably allows predators to reduce the population of small fish species more effectively than in the turbid rivers, and (3) an intensive fishery may remove rapidly growing valuable species, which will then become replaced by slow-growing undesirable fishes (Wajdowicz, 1964; Benson, 1968).
In tropical Africa the formation of a reservoir may lead to an explosive colonization of the new water body by cichlids, especially Tilapia and Sarotherodon, as noted for example in the Kariba (Balon and Coche, 1974), Volta (Petr, 1967), and Nyumba Ya Mungu (Petr, 1975) man-made lakes. In African lakes with a filling period extending over several years, tilapias come to dominate the commercial stock, while in lakes with a rapid filling (and usually a considerable draw-down) other fish will dominate. This seems to be related to unfavourable breeding conditions for tilapia in the latter type of lake (Petr, 1975a).
The so-called stabilized new reservoir fish populations may undergo considerable changes even a number of years after the closure of the dam. In Zambia, the Upper Zambezi fish species, formerly considered geographically isolated by Victoria Falls from the Middle Zambezi, successfully invaded the man-made Lake Kariba, where some of them, such as Alestes lateralis, successfully competed and eventually displaced the Middle Zambezi Brachyalestes imberi, which initially successfully colonized the lake (Balon, 1978). In the long run, the number of fish species in Lake Kariba might become higher than the number of species recorded from the unregulated river. Even today, more than 20 years after damming the river, the fish population of the lake cannot be considered as stabilized.
Attention must be given to the effect of fisheries on possible changes of species composition in reservoirs after some years of existence. Rounsefell and Everhart (1953) stated that the intensive fishery exploitation of a water body effectively changed the relative numbers of different species. This occurs because practically all forms of fishing gear are more or less selective as to the species taken. Under certain circumstances a species that is especially vulnerable will decrease more rapidly than the others. Ostroumov (1957) reported that the fisheries had a remarkably negative effect on the formation of the fish population in the Uglich and Ivan'kovo reservoirs in the USSR. He found that intensive fishery exploitation in these reservoirs in the early years of their existence prevented an increase in number of bream (Abramis brama) and zander (Stizostedion lucioperca). As a result, he recommended a cautious approach to fishery exploitation of newly created reservoirs. Commercial fishing should be permitted to a limited extent only after the formation of stocks of the most valuable fishes is completed. Any premature intensive fishery may disrupt the process of formation of the ichthyofauna, and lead to overpopulation of the new reservoirs with low-value fishes. The formation of mature stocks in temperate latitude reservoirs takes a number of years, for bream usually 5 to 6 years. In the tropics, on the other hand, tilapias will mature in 6 to 12 months, and commercial fishing of the new stock in a new reservoir could proceed therefore much earlier than in the temperate zone reservoirs. A delay in fishing might result in a loss of this peak production, which is usually followed by depression. The change of species composition and abundance of each individual species derived from fishing may less endanger the fish as a whole in the tropics where reservoir fisheries exploit a multispecies stock than in temperate regions where the fisheries depend only on a few species of fish.
However, in Southeast Asia and the Indian sub-continent, fishing is among the important factors causing changes of fish species composition in reservoirs. Gillnet fishing and intensive capture of predatory fishes by using hooks and longline with live baits, in the early years of an impoundment, have been found to encourage the build-up of dense populations of small fishes.
A knowledge of fish distribution in a reservoir provides valuable information for devising more efficient methods of sampling populations and more efficient methods of harvesting the fish crops. Most of the studies on the distribution of fish have dealt with fish collected by several methods such as trawls, gillnets, electro-fishing, poisoning, or traced by echo-sounder.
Fish are not distributed evenly throughout reservoirs. Studies on the Tennessee Valley Authority (USA) reservoirs indicated that the greatest abundance of fish in the main stream reservoirs appeared midway between dam and headwaters, where there are big expanses of shallow water from 0.3 to 3 m in depth (Gabrielson, 1950). Circumstantial evidence implicates several factors governing the distribution of fish in reservoirs. They are: (1) species of fish; (2) stage in life history; (3) feeding habits; (4) physico-chemical conditions of water, and (5) seasonal changes.
Studies in India indicate that the overall index of fish concentration in Konar reservoir, as reflected by the index of catch per day, shows the average fish density to be greatest in the upper zone, followed by intermediate and near dam zones. The distribution of fish in this impoundment varies with time of year. The catch is superior in the intermediate zone in May, June, October, and November (i.e., prior and after the monsoon); the upper zone in June and August; and near dam zone in October and December.
The distribution of individual fish species within a reservoir varies. In Keystone reservoir (USA) Eley, Carter and Dorris (1967) found drum were abundant during summer in the upper reaches of the reservoir. White crappies were distributed generally throughout the reservoir from surface to 9 m deep. Black bullheads were abundant in the middle reaches in water warmer than 24°C. Carp were widely distributed. Gars were more numerous in the upper reaches near the surface. Channel catfish were present in the upper reaches of the reservoir at the shallower depths during the autumn and winter, but were found at all depths in the spring.
Variation in distribution of fishes in reservoirs is closely related to feeding habits and food availability. Summerfelt (1971) reported that the distribution of several species in Lake Carl Blackwell, Oklahoma, USA, was related to a habitat characterized by relatively shallow water with little organic matter. He found distribution of the flathead catfish, a highly piscivorous fish which feeds upon freshwater drum and gizzard shad, was directly correlated with the density of its forage. Distribution of white crappie was positively correlated to biomass of mayfly and gizzard shad. White bass were abundant in shallow littoral zones with abundance of mayflies. Freshwater drum was numerically correlated with abundance of oligochaetes and Chaoborus.
There are several physical and chemical factors influencing the distribution of fishes in reservoirs. Coke (1968) reported that in a bush-cleared area of Lake Kariba, Zimbabwe, the number of fish drastically decreased with increasing depth of water. A similar observation was reported by Balon (1972). With a large-scale echo-sounding survey, he found that fishes were limited to areas of shallow water close to shore and mainly to bays and coves. He stated that fishes seemed to concentrate in the evening along the shoreline and especially in the embayments, being distributed in remarkably high density from the surface to 15 m depth with maximum density at 6 to 7 m depth. Only at places shallower than 15 m were the fish distributed in the evenings throughout the whole water mass. At dawn fish had changed to a less dense concentration extending further offshore to a distance of about 2 km. It appeared that in the areas around islands with steep shores, fish were found only in coves or in strips a few metres wide close to shore.
A reduction in reservoir volume during high withdrawal periods in summer may influence the distribution of fish as a result of increasing the relative thickness of the warm upper layer. Cady (1945) found the distribution of large-mouth bass, which is the most important species of the Norris reservoir game fish, was more abundant at 10 to 20 m depth than near the surface. Dendy (1945) marked temperature as the most significant factor in influencing fish distribution in Norris reservoir. He stated that most species of fish tended to move to deeper water as the summer progressed and as the isotherm move downward. Horak and Tanner (1964) found rainbow trout, kokanee salmon, and white sucker of the Horsetooth reservoir (765 ha), Colorado, USA, in deeper waters during summer. Kokanee salmon were found most abundant in water of 10.6°–12.8°C, white rainbow trout and white sucker were concentrated in 19°–21°C.
Changes in vertical distribution of fish between daytime and night-time are well-known. It usually appears that pelagic fishes are aggregated near the bottom in the daytime and dispersed over the whole volume at night.
Five species of small pelagic fishes in the new Volta Lake in Ghana showed a diel periodicity in catches, and were strongly attracted to light (Reynolds, 1971). The fish (clupeids Pelonulla afzeliusi and Cynothrissa mento, schilbeids Siluranodon auritus and Physailia pellucida, and cyprinid Barilius niloticus) spent the day either at the bottom or at the oxygen discontinuity which is usually situated at a depth of 20 to 25 m. At night the fish dispersed from their tight migrating schools into loose aggregations of individuals feeding at the surface in the same area. Fish in these associations are strongly affected by light; a bright moon causes total dispersal of the fish in the surface waters. It appears that a diel vertical movement occurs so that the fish can exploit a feeding ground at its richest moments, and at times when they are least conspicuous to predators.
Netsch and his colleagues (1971) reported that the night-time catch rates of metre nets and of midwater trawls on gizzard and threadfin shad in Beaver reservoir (USA) were 5–110 times greater than day-time catch rates. This may be a good example of how fish distribute in the reservoir between day and night.
The concentration of dissolved oxygen in water is considered as the most significant chemical factor influencing fish distribution in reservoirs. Dendy (1945) reported that, as oxygen depletion progressed beneath a density current in Norris reservoir, fishes tended to move through the current to the warm well oxygenated water immediately above it. However, he pointed out that the presence of dissolved oxygen of 3 ppm or more in reservoir water did not appear to influence warm-water fish distribution. A similar phenomenon has been found in Keystone reservoir (Eley, Carter and Dorris, 1967). They reported that fishes tended to move through the chemocline to upper aerated water as soon as the oxygen content of the hypolimnion reached 2 ppm or less. Results from year-long observations with echo-sounders and gillnets in Bull Shoals reservoir provided evidence that most warm-water fishes seldom occupy waters below the thermocline (Houser and Dunn, 1967). The night-time distribution of fish was found to be remarkably uniform over the entire lake. There was a definite order of decreasing density with depth and the vertical distribution was sharply limited between the surface and thermocline during summer.
In Thailand, fishes are usually found to be numerous in shallow areas of less than 3 m depth. Diurnal movements, both horizontal and vertical, are mostly for feeding. Pelagic species such as freshwater sardine (Corica goniognathus) remain in deeper zones during the day and move up to the water surface for feeding at night. Commercial fishing for this species is based on light attraction. Seasonal movement, mostly for breeding, occurs during the rainy season. Many species, such as striped silver catfish (Pangasius sutchi), striped carp (Probarbus jullieni), and golden breast carp (Puntius daruphani) migrate into rivers and streams and head to spawning grounds upstream. All phytophilous species will migrate to newly flooded areas with shrubs and bushes along the shore for spawning. Young fishes feed in shallow areas nearshore. When water level in reservoirs decreases in winter, fishes will migrate to deeper zones or to permanent pool areas and will remain there throughout the summer. The situation is probably similar to that existing in other countries in the region. More information is still needed for better understanding and planning for development of reservoir fisheries in Southeast Asia.
Robson and Regier (1968) stated that monitoring of the numerical changes which occur in a population through the course of time is essential to a basic understanding of population dynamics, production, yield and the rational management of a fishery. Several methods have been developed for estimating fish population, most of them attempting to estimate population size or relative abundance. Marking or tagging of fish and subsequent recapture is one of the field techniques employed in the estimation of freshwater fish populations. Another technique useful in measuring population density is the monitoring of the catch and fishing effort in an exploited population during a given period of time. A number of mathematical models, underlying the methods used in the estimation of size and other parameters of fish populations, are set up with certain assumptions concerning either the population, the fishery or both in an attempt to make reasonably close predictions of the changes in numbers, mass, and distribution of the populations. It should be borne in mind that many of the formulae and models constructed by workers in temperate regions have been established for monospecific commercial stocks, while in tropical water the fish stocks are mostly multispecific.
The simplest methods of estimating population size depend directly on enumeration of a whole or a selected portion of a population. Draining of a pond, for example, may result in complete recovery of all fish present, fish migration through a narrow channel or fish ladder may yield complete enumeration. These methods can be used only in small water bodies or with migratory species. Whenever fish population estimates are required in large water bodies, it is necessary to employ other means.
This method estimates total population by assuming that the population is non-migratory at least during the period of sampling, and that areas of sampling are similar habitats. The sampling is executed by completely blocking an area (at random) with nets and the total population in each section captured by poisoning (Henley, 1967; Hayne, Hall and Nichols, 1967; Balon, 1972), electric shocking (Klein, 1967; Vincent, 1971) or other means. The abundance of a population is estimated as the numbers or weights of fish per unit area of sampling and extrapolated to the whole body of water.
This method estimates by means of an echosounder or sector scan sonar (Cushing, 1963). It is a useful study of a whole water column. But there are two very serious problems in that identification cannot be made directly, and it is difficult to detect demersal fish in very deep water. However, the echosounder is common and very useful to commercial fisheries. It has been introduced to reservoir fishery investigation by several investigators, for example, Jenkins (1964), Houser and Dunn (1967), Balon (1972). Echosounder survey is a method of estimating the abundance of fish traces based on the presence or absence of fish within a given transmission. Abundance is estimated as the number of presences recorded per unit of distance traversed.
Marking experiments can be used for determining the size of fish populations in a variety of situations. This method has been used chiefly as a means for studying migrations and distribution, utilizing the return for growth studies, mortality and population size. The ability to account for the presence and recovery of marked fish has come to be recognized as a powerful tool for studying population dynamics. Types of tags presently in use which show promise in reservoirs are the double-barbed (spaghetti) dart, the vinyl tube spaghetti, the monel metal jaw, plastic streamer, and Swedish trailer (Jenkins, 1964). Experiments have been employed in many north American reservoirs such as, for example, in the TVA storage reservoirs (Manges, 1950).
Ricker (1958) recommended that in practice the methods of marking or tagging and recapture should be based on the following assumptions: (1) the marked fish suffer the same natural mortality as the unmarked; (2) the marked fish are as vulnerable to the fishing being carried on as are the unmarked ones; (3) the marked fish do not lose their marks; (4) the marked fish become randomly mixed with the unmarked, or the distribution of fishing effort (in subsequent sampling) is proportional to the number of fish present in different parts of the water body; (5) all marks are recognized and reported on recovery, and (6) there is only a negligible amount of recruitment to the catchable population during the time of recoveries.
Methods used in estimating fish populations by means of mark-recapture techniques include the Petersen method (Ricker, 1958) and the Schnabel method.
Determination from catch statistics (Nikolskii, 1969; Ricker, 1958) utilizes the primary data on catch and on the effort expended in obtaining various segments of the catch. The fluctuations in catches usually reflect fluctuations in population numbers when the fishing intensity exceeds a certain level, and may be used as a reasonably reliable index if taken at least from year to year (Nikolskii, 1969). He stated that variations in catch over shorter periods reflect short-term alterations in the annual cycle rather than fluctuations in numbers. The principle of this method is only applicable when a population is fished until enough fish were removed to reduce significantly the catch per unit effort. The accuracy of forecasting depends on the ability to estimate the slope of the regression of catch per unit effort. Whenever a very small proportion of the population is taken, the slope is either nil or slight. A change in slope makes a tremendous difference in the point at which the projection of the regression line cuts the X-axis. This type of estimation assumes that the population remains unchanged during the sampling period.
The Leslie and Davis method (1939) and the De Lury method (1947) have been given formulae by Ricker (1958) to calculate the initial population. Currently, Mahon (in press) tested in experimental fishing the accuracy of catch-effort methods for estimating fish density and biomass in streams. For De Lury's method ln or log catch per unit effort is plotted versus cumulative effort. If the proportion of fish taken by one unit of effort is greater than 0.02, De Lury's method is inappropriate and Ricker's (1975) modification should be used. The Leslie and Ricker estimates tended to underestimate. The results of Mahon's experiments and calculations suggest that the most appropriate way of decreasing error would be to increase the total effort and consequently the proportion of fish collected.
A knowledge of age and rate of growth of fish is extremely useful in fishery biology and fishery management. They are together the most important basic information for longevity, sexual maturity or spawning time, catchable size, environmental conditions of the water bodies, suitability for stocking and continuing studies in fishery production.
Carlander (1955) summarized the use of these data in fishery management: (1) to provide the general background needed for management decisions; (2) to aid in diagnosis of management needs such as the recognition of overcrowding and stunting; (3) to evaluate the effects of various environmental or hereditary factors by comparing growth rates and year-class abundance; (4) to evaluate the effects of abundance; (5) through the use of growth and mortality rates to estimate optimum yields and the possible effects of catch regulations; (6) to predict catch from year-class abundance; (7) with additional estimates of population numbers, to determine production rates; (8) to separate populations, although further growth and scale analysis are often needed, rather than just ageing.
Some problems associated with the ageing of fish were discussed at a special international symposium at Reading, UK (Bagenal, 1973).
Several methods have been used in age and growth studies of fish. Rounsefell and Everhart (1953) listed the methods of age determination of the fish as follows: (1) comparison of length frequency distribution; (2) marking or tagging and recapture; and (3) interpretation and counting of growth zones or annual marks which appear in the hard parts of fishes. Among these, the simplest method is the analysis of length frequency distributions. This method requires a unimodal size distribution of all fish of the same age (Tesch, 1968). It is easy to employ if there is no large overlap in the size of the individuals in adjacent age-groups, and is more generally useful in the younger age-groups of a population. However, it appears that the utility of this method declines with age for most teleost populations (Weatherly, 1972). Furthermore, it is relatively useless for those tropical fish which reproduce several times per year.
The marking or tagging method seems to be most satisfactory because it provides important data and explains several aspects of the mode of life of the fish, such as age, growth rate, migration, and distribution of fishes. Olsen (1954) stated that this method is almost the only approach to age determination in certain fish such as the elasmobranchs, in which hard parts are unavailable and length frequency distributions are suspect. However, some problems still occur, since most tagging methods are not applicable to small fish, or if they are, they may cause great mortality so that recaptures are few. Many of marked fish may lose their marks, and the marks often go unnoticed by the fishermen.
The most frequently used method of age determination of fish is the examination of annual marks on scales, otoliths, spine and finrays, opercular bones and vertebrae. The scale method is the most popular ageing technique of fish, but problems arise with many tropical fish (Fagade, 1974). In Lake Kariba in Africa, scales were used for age determination of Sargochromis codringtoni (Holcik, 1974), Tilapia rendalli (Bastl, 1974), Sarotherodon mossambicus mortimeri (Krupka, 1974) and some others.
The use of otoliths for age determination has been summarized by Williams and Bedford (1973). May (1967), Kim and Roberson (1968) determined age of cod and salmon from otoliths; Chitravadivelu (1974), Frank (1974), Pivnicka (1974) and Cerny (1974) studied age and rate of growth of a number of Lake Kariba fish from vertebrae; Bardach (1955) used opercular bone as a tool for age and growth studies of the yellow perch; Boyko (1950) estimated age of many fish species using finrays.
The age composition of an exploited fish population is normally determined by analysing age composition of the catches taken at stated intervals over the season or year. Several methods have been recommended for determining the age composition of a commercial stock (Gulin, 1968). They are:
Age composition of the stock may be determined by converting age composition of the catches over a season or year;
Age composition may be obtained by methods entailing calculation of the absolute numbers of the population from catches in a unit effort, and size and age of fish in the catch. Age composition of the commercial stocks is then estimated by extrapolating the samples of age analyses to total stocks;
Age composition of the stock may be calculated from the mean of samples taken to determine the age composition of catches with standard fishing gear. Trawls are best suited, but there is a limit on their use in freshwater reservoirs with drowned trees.
Growth is an important factor influencing the time of sexual maturation, reproduction, production and fishing regulations. The growth process is specific for each species of fish. In natural waters, the growth of a fish population is dependent upon the abundance of the stock, food supply, space, temperature, growing season, and other environmental factors. The methods used in growth studies in freshwater investigations fall into three main groups (Tesch, 1968):
Direct observation - Growth may be studied experimentally by tracing the seasonal and annual increase of fish of known size and age in tanks, ponds, etc.
Tagging and marking - These methods require certain techniques which cause little or no retardation of growth such as the internal tags, colouring with dyes, and fin clipping. Study is then made by measuring the increment in length or weight of marked fish between liberation and recapture. This method is costly to apply on a large scale. Other constraints are that the growth data are usually obtained over relatively short intervals, and growth of tagged fish may not be representative of the untagged population (Parrish, 1958).
Back-calculation of lengths or weights - This method is done by taking length measurements on skeleton structures and then calculating the total length of fish at the end of successive years of life. Thus, its validity depends on the relationship between the growth in length or weight of fish and the growth of the skeleton structures over a certain period of time.
The skeleton structure most used in this method is the scale, but otoliths and opercular bones also have been successfully used (Parrish, 1958). Much attention has been given to the nature of scale and body relationship. Tesch (1968) found when fish length was plotted on the Y-axis against scale radius on the X-axis, the relationship obtained may be either: (a) linear and passing through the origin (of the form: L = bS), or linear but not passing through the origin (of the form: L = a + bS); (b) curved with increasing slope; or (c) S-shaped with slope at first increasing, later decreasing. The back-calculation of growth history can be done by a number of methods (for details see Ricker, 1958, 1968).
Chugunova (1963) stated that the average number of individuals (expressed in percentages) spawning for the first time in any fish population is directly proportional to the growth during their feeding period before reaching maturity. This statement appears to be true for the majority of fish, but not for all and not in all cases. Sometimes, on the contrary, maturity occurs early with poor growth, for example, in Crucian carp (in basins containing little food), and dwarf males of salmon. It is usually remarked that the age of maturing or rate of sexual maturity of a particular fish species depends on some external factors, the most important of which are: food, temperature, photoperiod and water currents. A greater number of experiments have been carried out at the Kalarne Fishery Research Station in Sweden to study the connexion between first sexual maturity, size (growth rate), and age in fish (Alm, 1959). The results are summarized as follows:
Fish of a good growth rate reach maturity at a lower age and a smaller average size than fish of medium growth rate which reach maturity at a higher age. Again, fish with poor growth become mature at a higher age and their average size is smaller than that of fish matured at medium growth rate;
The age of maturity is usually higher in large-sized and often fast-growing fishes than in small-sized and slow-growing species;
The age of maturity in different populations of the same species depends mainly on size and thus indirectly on growth rate. The maturity of fish with an initially good growth rate is reached at an earlier age than of fish with a beginning poor growth rate;
The average size of the individuals in the year-class or population with good growth rate is greater than in cases with poor growth rate;
Fish, on account of better growth rate, reached maturity at a lower age and spawned a greater number of times than fish of a poorer growth rate and with later maturity;
The earlier or later age of maturity has not had any perceptible influence on mortality and longevity, and in spite of the fact that the males practically always reach maturity one or two years before the females, mortality has not been perceptibly higher in the former than in the latter.
Two methods are known (Chugunova, 1963): (a) determination on the basis of spawning checks; and (b) an overall determination of the maturity of the sexual products by dissecting the fish. The determination on the basis of spawning checks is sometimes difficult because spawning checks are not seen well on all fish, and are often absent in some individuals of a given population or in some tropical fish.
The fecundity of fish is defined as the number of ripening eggs found in the female just prior to spawning (Bagenal, 1967). Several parameters have been used in fecundity evaluation for fish, including individual fecundity, relative fecundity, and specific fecundity (Nikolskii, 1969). Individual fecundity refers to the number of eggs for the generation of that year present in the ovaries, or that should be laid that year. The relative fecundity is the number of eggs per unit body weight of fish. The specific fecundity is interpreted as the following relationship:
SF = (1 + r)1/pjs
Where, r is the individual fecundity, p is the period between laying times, j is the age of onset of sexual maturity, and s is the ratio of the sexes.
The fecundity of individual female fish shows significant variations depending upon genetics, length, weight, age of fish, season and food supply in the previous season (Le Cren, 1965).
The number of eggs gradually increases with age in the majority of fishes, but as the individual approaches senility it usually begins to decrease (Nikolsky, 1963). Typical results of fecundity investigations have shown that the number of eggs are approximately proportional to the cube of the length, or linearly proportional to weight or to age of the fish (Bagenal, 1968). Bagenal (1967) plotted fish length against fecundity, and found their relation: F = aLb. His further analysis using logarithmic transformation, fits a straight line by the method of least square, and fecundity can be estimated from the equation:
log F = log a + b log L
Nikolskii (1969) reported that the length of the spawning period and the mode of deposition of the eggs are very important features in population studies. Prolonged or repeated spawning is an adaptation to unstable spawning conditions for a survival of the eggs. In many fishes the survival rate of the larvae is dependent on the time during the spawning interval when the eggs are laid. Le Cren (1965) stated that the number of eggs laid is influenced by several factors, the more important of which are: the number of spawning females, their age and size, the food supply in the period before spawning, access to spawning grounds, the area of suitable grounds available, and competition for space on the spawning grounds.
The survival of the young is governed by the number and quality of the eggs and sperm, as well as the biotic and abiotic conditions. Nikolskii (1969) stated that variations are prominent in species with a very labile food supply, the mechanisms being substantially related to the supply of internal food received from the egg. Large fluctuations occur for species whose egg-fat content is low and whose fry cannot live long without an external food supply. It is evident that the amount and quality of the yolk are related to the feeding conditions of the parent population. Food supply depends on the concentration of the food, variation in feeding season, feeding area, and density of population. Deterioration in food supply after the larval stage usually reduces activity and growth together with increased losses due to predators, diseases and parasites.
The variation in recruitment depends on the number of parental stock, the number of eggs they lay, and the subsequent survival of the young stages up to the recruit stage (Le Cren, 1958). In most fish populations, particularly those species that lay a large number of eggs, survival in larval and young stages appears to decide the strength of the brood at the age of recruitment. The survival of a brood and its strength usually depends on changes of climatic and hydrological conditions.
Timeliness is often more important than precision in making yield estimates for reservoirs with extant fisheries (Henderson et al., 1973). Consequently, a holistic approach estimating first approximation yield will be more useful to the fisheries manager than some of the traditional demographic, and more time-consuming methods. Some of these holistic methods are detailed in Regier, Cordone, and Ryder (1971), Henderson, Ryder, and Kudhongania (1973), Ryder et al. (1974), Ryder and Henderson (1975), and Ryder (1978). While initially precision may be lost in the application of these methods, estimates may be continually refined from the feedback information from the fishery itself, and management policy deriving from such estimates will be more realistic and applicable (R.A. Ryder, pers. comm.).
The production of fish in reservoirs is affected by morphological, physical, chemical and biological factors. Rawson (1952) found a highly significant inverse relationship between fish production and mean depth. Carlander (1955) found a significant increase in standing crop per acre with increase in carbonate content of the water in mid-western reservoirs of the United States. Cooper (1966) pointed out that high production of fish in newly created reservoirs was a result of water enriched with nutrients from newly flooded substrata which favour production of plankton. This condition consequently results in good survival of fish fry and young, and good growth rate of fish. Jenkins (1967) studied the influence of some environmental factors on standing crop and harvest of fish in 210 US reservoirs by using regression analysis. He found dissolved solids, area and shore development were positively correlated with total standing crop. Sport harvest was negatively correlated to age of reservoir and area. Commercial harvest was positively correlated to age, but negatively to mean depth and storage ratio.
The effects of engineering design and operation, together with selected reservoir environmental variables, on fish standing crop have been investigated in 140 large reservoirs (Jenkins, 1970). Data were interpreted by logarithmic partial correlation and multiple regression analyses. He found logarithmic partial correlation revealed highly significant (0.01 confidence level) positive effects of outlet depth, shoreline development and dissolved solids on total standing crop in all samples. At 0.20 confidence level, the crop of all sport fishes was positively influenced by outlet depth, storage ratio and shoreline development, but negatively by mean depth. In 54 hydropower reservoirs with a stable thermocline, positive effects of increased storage ratio and dissolved solids on total crop were evident at the 0.05 confidence level. Increase in thermocline depth had a negative effect.
Brylinsky and Mann (1973), in their interpretation of data collected from 43 lakes and 13 reservoirs distributed from the tropics to the Arctic, showed variable solar energy input was a greater influence on production than variables related to nutrient concentration in the water. Similar results were also demonstrated by Henderson, Ryder and Kudhongania (1973). They showed the predicted potential yields, for six sets of natural lakes representing six of the world's climate zones at any constant MEI, are highest in the tropics and then decrease continuously from the tropics toward the polars (Figure 4).
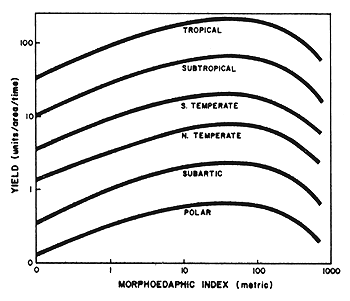 | Figure 4. Predicted yield curves for sets of natural lakes representing six of the world's climatic zones (from Henderson, Ryder and Kudhongania, 1973) |
Jackson (1966) observed greater sport harvest in tropical reservoirs than in reservoirs of temperate regions. He compared average annual sport harvest of 40.4 kg/ha in TVA reservoirs in the United States to 44.9–56.1 kg/ha in Lake Kariba, Africa.
In Latin America, Paiva (1976) has identified several factors, both biotic and abiotic, influencing fish yield in Brazilian reservoirs. He listed 10 positive and five negative factors affecting fish yield. They are:
| Augmentative Factors | Reductive Factors |
|
|
He has given equal weight to these factors for the evaluation of fishery potential, and each factor has been arbitrarily assigned a value of 10 kg/ha in terms of its positive or negative influence.
The major factor influencing fish production in reservoirs of Southeast Asia may be water-level fluctuation. An annual flooding of dry marginal land in reservoirs during the rainy and flood seasons will have a similar effect on fish stock in the reservoir as on fish stock of rivers overflowing onto adjacent floodplains. The importance of floodplains for especially tropical fisheries has been described in detail by Welcomme (1979). It is apparent that high fish yield usually occurs following the high water-level year. This can explain that most freshwater fishes in Southeast Asia spawn during the rainy season and they are mainly phytophilous species. In high water-level year, there are more spawning grounds as well as nursing and feeding areas available along the shore and connected flooding areas nearby. Inundation of large areas of dry lands results in a release of more nutrients into reservoirs, resulting in a high production of phytoplankton, followed by zooplankton and benthos, which are the major sources of food for fish. There will be a high survival rate of fry and fingerlings and an increase in growth rate. Fish production can be increased by additional measures such as stocking with indigenous species (Indian major carps in India, carps and catfish in Thailand, etc.), and exotic species (Tilapia in Sri Lanka, Indonesia, India, Chinese carps in Singapore, etc.). In some countries where reservoir fishery resources are underutilized, promotion of fishing activities is needed. This may be done by introducing fishing techniques and fishing gears to local people and teaching them how to fish as well as how to utilize the catches.
The ability to estimate fish production is very important in permitting management personnel to make a more accurate appraisal of the expected harvest from reservoirs. It should be clarified here that when discussed in terms of fish production it means at the community level rather than at the traditional species or population level as was clearly stated by Ryder (1978). Several methods for estimating fish production in lakes and reservoirs have been recommended (Ryder, 1965; Jenkins, 1967; Gulland, 1971; Sheldon, Prakash and Sutcliffe, 1972; Melack, 1976; Oglesby, 1977), with specific assumptions and variables. All estimates interrelate the physical factors to chemical properties and biological conditions of the impoundment and then relate them to standing crop and/or to the catch. The results of estimations show various degrees of significance, which mainly depends on quality of morphoedaphic conditions of impounded water and accuracy of records of the catches.
Ryder (1965) has established the Morphoedaphic Index (MEI) and demonstrated it to be an effective method for yield prediction in north temperate lakes. The index comprises two basic and easily measured parameters of a lake system, namely, total dissolved solids (TDS), a correlate of nutrient levels (edaphic factor), and mean depth (z), a morphometric factor. He related fish production (in pounds per acre per year) to total dissolved solids (in ppm) and mean depth (in feet). The relationship is expressed as follows:
MEI = TDS/z
and the fish yield can be calculated from the following equation:
Y = kxa
where y = fish yield; x = MEI; k = constant that represents an appropriate coefficient for climatic effects, and a an exponent approximating 0.5 or the square root.
He demonstrated that the yield production for north temperate lakes could be calculated from the equation:
y = 2.094 x0.4460
Thereafter, this equation has been transformed to metric units (Ryder et al., 1974) as follows: MEI (metric) = 3.281 MEI (English); the metric expression is therefore y ~ 1.24 x0.446; where yield is expressed as kg/ha/year, z as metres, and TDS as mg/1. When plotted, the relationship of the logarithms of fish production to the logarithms of the morphoedaphic index approximates a straight line (Figure 5).
| Figure 5. Yield and/or biomass curves for four regional lake groups (from Ryder et al., 1974) | 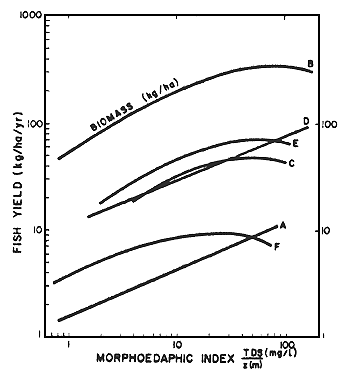 |
The morphoedaphic index has provided the simplest and most general approach to the problem of initial estimates of potential yield in lakes and reservoirs (Henderson, Ryder and Kudhongania, 1973). A modification of the morphoedaphic index equation has been used in prediction of potential fish yield of 31 African lakes because of the greater availability of data on the conductivity of African waters compared with total dissolved solids (Henderson and Welcomme, 1974). Ryder's original equation was modified as follows:

For these lakes the total catch in kg/ha against MEI resulted in the equation:
y = 8.7489 x0.3813
with a correlation coefficient = 0.5073 which is significant at the 0.01 level.
The total catches fluctuated with the density of fishermen operating on the lakes (intensity of exploitation), and when the recorded catches were examined, it was found that at densities less than one fisherman/km2 the catch per unit area was less than maximum. Above this level the catches are optimal until the density of about three fishermen per km2 is reached when they fall abruptly. To compensate for the variation introduced by diversity in intensity of exploitation, catch against MEI was replotted by excluding the 14 lakes with less than one fisherman/km2 to obtain the equation:
y = 14.3736 x0.4681
with an improved correlation coefficient = 0.6864, and is also significant at the 0.01 level.
Further, they made a comparison between the predicted yield of tropical African lakes and of north-temperate lakes (Ryder, 1965). They found a meaningful relationships between them which indicated that the yield from tropical African waters is approximately ten-fold higher than that of north temperate waters. This is largely explicable by differences in seasonality and mean temperature (about 25°C for the tropics and 10°C for north-temperate zone) between the two regions.
The morphoedaphic index technique has become popular and gained recognition as a tool for predicting fish yield in lakes and reservoirs in all regions. The greatest use for this method is a rapid assessment of potential yield where detailed data on the fish stocks are lacking, as computation of the index requires knowledge only of the mean depth and a mean value of total dissolved solids or any of its correlates such as conductivity or total alkalinity (Henderson and Welcomme, 1974; Ryder and Henderson, 1975).
The application of MEI as a tool for predicting fish yield requires the establishment of a family or set of lakes and reservoirs suitable for regression treatment. Consequently, it will be necessary to ascertain if specific criteria are satisfied, i.e., the lake or reservoir is subject to a relatively homogeneous set of environmental conditions (Ryder, 1978). These conditions might include a characteristic and reasonably constant proportionality of ions, a flushing regime roughly proportional to the lake volume, inorganic turbidity levels on about the same order of magnitude for all lakes in the set, and moderately intensive to intensive fishing effort on most abundant species over a period of years. Furthermore, he recommended that in cases where the regression for a family of lakes at any given latitude has not been previously described, it may be useful to do so. A series of ten or more lakes are preferably selected that appear to be homogeneous with respect to most external abiotic factors, especially climate, and a regression is calculated between fish yield and the MEI.
Jenkins (1967) had developed models describing the influence of some important environmental factors on reservoir fish standing crop and harvest. He found the relationship between standing crop and morphoedaphic index to be the most meaningful regression of predictive utility. He developed an equation for calculating yield as follows:
y = 2.07 + 0.164 x
where y equals log standing crop, and x equals log morphoedaphic index. The coefficient of correlation (r) is 0.325, significant at the 0.01 level.
The regression of sport harvest in 121 reservoirs on morphoedaphic index was:
y = 0.88 + 0.183 x
where r = 0.23, significant at 0.02 level.
Commercial harvest versus the index of 46 US reservoirs was expressed as:
y = 0.52 + 0.210 x
where r = 0.21, not significant at 0.05 level.
However, his attempt to relate commercial catches and sport catches separately to morphoedaphic index did not give convincing results. With some modification by using total annual catches (average annual commercial plus sport catches) from his data plotted against the morphoedaphic index, Regier, Cordone and Ryder (1971), found a significant relationship between the total catches and the index.
In 1971 Jenkins and Morais reported their studies on the influence of environmental variables in relation to reservoir sport harvest. They derived an equation:
y = 0.2775 - 0.2401x1 + 1.0201x2 - 0.2756x3
where y is log total harvest in kilogrammes per hectare, x1 is log area, x2 is log growing season, and x3 is log age of reservoir. By using this equation they predicted the harvest for Beaver reservoir, Arkansas, through the first six years of impoundment as 208 kg/ha compared to field estimated total yield for that period of 176 kg/ha. For Bull Shoals reservoir, the predicted harvest from the second through sixth year was 136 kg/ha compared to field estimated total yield of 145 kg/ha.
A simple model of the relation of potential yield to known biomass of a virgin fish stock has been developed by Gulland (1971). He gave an equation:
y = kMB
where y is estimated total fish yield on a long-term basis; k is a constant which probably lies between 0.3 and 0.5 (or equals 0.4 typically); M is the natural mortality coefficient; and B is the fish biomass prior to fishing.
Regier, Cordone and Ryder (1971), in applying the model to the unexploited multispecies standing stocks of Lake Victoria, used a community mortality coefficient based on individual species mortality coefficients weighted on the basis of their proportionality within the total experimental catch. Ryder and Henderson (1975) used this method in obtaining one of their estimates of the potential yield of Lake Nasser (now called the “High Dam Lake”) (Table 1). Lacking direct data on mortality, mortality rates were approximated for the major species or species groups entering the fishery on the basis of their average known life-span following the attainment of exploitable size. Further, these authors, lacking data on the standing stock of fish, obtained a first approximation by equating fish standing stocks to the estimate of zooplankton standing stock obtained by Entz et al. (1971). This argument was based on the hypothesis of Sheldon, Prakash, and Sutcliffe (1972) that in the aquatic environment roughly equal concentrations of material occur at all particle sizes within the ranges from 1 μ to about 106 μ, i.e., from bacteria to whales. The resulting estimate of a potential fish yield of 11 000 t, or about 26 kg/ha, seemed reasonable. Current catches in Lake Nasser are about 20 000 t. Clearly such approaches can only be expected to give “order-of-magnitude” indications.
Table 1
Data used in estimating long-term potential yield for Lake Nasser,
Egypt, based on the Gulland (1971) model
Mortality coefficients were approximated as in Regier et al. (1971) After Ryder and Henderson (1975)
| Species | Approximate mortality coefficienta | Relative biomassb | M × B |
| Tilapia sp. | 0.25 | 0.56 | 0.1400 |
| Alestes sp. | 0.40 | 0.18 | 0.0720 |
| Labeo sp. | 0.25 | 0.09 | 0.0225 |
| Lates niloticus | 0.10 | 0.07 | 0.0070 |
| Bagrus bayad | 0.15 | 0.04 | 0.0060 |
| Clariidae | 0.30 | 0.01 | 0.0030 |
| Schilbeidae | 0.60 | 0.01 | 0.0060 |
| Others | 0.50 | 0.04 | 0.0200 |
∑MB = 0.2765 |
a Based on known longevity of fish following recruitment to exploitable size
b Based on 1972 percentage composition in the commercial fishery
The relation of fish yield to primary production has been investigated. Henderson, Ryder and Kudhongania (1973) found that most trophic transfer efficiencies may tend to be low in waters of low primary productivity and high in waters of high primary productivity. Melack (1976) showed a regression equation describing the relation between fish yield (FY) and gross photosynthesis (PG). He found the relation for eight African lakes as follows:
log FY = 0.113PG + 0.91, with
a coefficient of determination of 0.57. A very similar relation was found for fifteen tropical Indian lakes:
log FY = 0.122PG + 0.95, with
a coefficient of determination of 0.82. He pointed out that, in using these equations for predicting fish yield, caution should be exercised as:
If estimations of primary productivity have been made at a few stations repeatedly for several times in one year, and then converted to a lakewide average annual production, errors are exaggerated;
At low fishing effort, commercial catch records may only indicate fishing effort, not potential yield, and may apply to only one region of the lake.
Oglesby (1977) has studied the relationships of fish yield to summer phytoplankton standing crop in temperate lakes ranging from oligotrophic to highly eutrophic, small to very large in area, and very shallow to very deep. He found high correlation of fish yield with summer phytoplankton standing crop as expressed in the following equation:
log = Y1 = - 1.98 + 1.17 log CHLs
where Y1 is annual yield of fish expressed as dry weight per square metre of lake surface, CHLs is summer phytoplankton standing crop; the correlation coefficient (r) = 0.91, significant at 0.05 level.
The analyses of the relations between fish yields and primary productivity for temperate and tropical lakes show promise as a method for assessing a fishery's potential. Oglesby (1977) recommended that this technique needs more research and development before it can be used as an effective predictive tool. Points that need to be clarified include better definite boundary conditions and the probability that this function is curvilinear (with important implications for fishery management).
These measurement techniques for prediction of fish potential yield in lakes and reservoirs might be very important to the development and management of freshwater fisheries in Southeast Asia. In this region, the studies of fish production in the past were mainly based on the measurement of fish standing crop by the area density method and catch landing statistics. Some authors estimated fish production by other methods such as the frame survey, and catch per unit of effort (Dubey and Chatterjee, 1977). There is a number of studies of changes of physico-chemical and biological conditions in lakes and reservoirs in this region. Therefore, it might be possible, at least in some cases, to estimate fish potential yield in lakes and reservoirs.
![]()
![]()
![]()