![]()
![]()
![]()
L'une des conditions essentielles de l'évaluation ou de l'aménagement des ressources est l'obtention d'une bonne estimation de l'effort de pêche. Pour ne pas être biaisé, il doit satisfaire à la relation classique F = qf. Il en découle que toute variation de q, qu'elle soit périodique ou à long terme, entraîne pour un effort nominal donné une variation de la mortalité par pêche correspondante. D'autre part, la prise par unité d'effort C/f est souvent utilisée comme indice d'abondance dans la mesure où l'on admet la relation également classique C/f = qD où D est la densité (abondance par unité de surface). Il en résulte que toute variation de q se traduira par une variation “apparente” de la densité.
Pour ces raisons, il a donc paru intéressant aux auteurs d'examiner les différents types connus de variations de q chez les crevettes et leurs conséquences, même au prix de quelques répétitions. Nous distinguerons, pour la facilité de l'exposé, les variations nycthémérales, tidales, lunaires, saisonnières, spatiales, liées à l'âge et interannuelles. Les distinctions sont parfois arbitraires et il est clair en particulier qu'il y a, chez les animaux dont la durée de vie dans la pêcherie est proche de un an, superposition flagrante entre les variations saisonnières, spatiales, ou avec l'âge, car elles sont indissolublement liées à l'évolution des cohortes dans l'espace et dans le temps.
Elles ont été abordées au chapitre 2 de façon sommaire. Ces variations ont été mises en évidence depuis longtemps et par de nombreux auteurs car leur manifestation évidente est l'existence de variations nycthémérales de rendement. Les crevettes sont, en général, des animaux à activité nocturne qui restent enfouis dans le sédiment la journée. Les rendements, reflétant le rythme d'activité, sont en général meilleurs de nuit, en mer, et également dans les estuaires et les lagunes. Ce n'est cependant pas toujours le cas en mer et, lorsque les rendements sont plus élevés le jour, on peut penser que pour des raisons liées à l'éclairement (et à la transparence des eaux) les crevettes sont posées sur le fond pendant le jour, au repos, et nagent en pleine eau la nuit, particulièrement hors de portée des chaluts.
Le rythme d'activité “apparent” est alors l'inverse du rythme réel. Ces variations temporelles à courte période sont “gommées” par l'utilisation de données moyennes de prise et d'effort permettant de calculer un indice d'abondance journalier.
Garcia, Petit et Fonteneau (1973) ont étudié ces variations très en détail dans le golfe de Guinée, sur divers fonds de pêche, allant du Nigéria au Sénégal, et à diverses saisons. Ils ont montré que, dans cette région caractérisée par des eaux à haute charge solide, les rendements étaient le plus souvent maximaux le jour mais que des inversions du rythme d'activité “apparent” se produisaient périodiquement, en liaison avec les conditions hydrologiques saisonnières (température à la limite nord de distribution de l'espèce et turbidité). Nous verrons plus loin que ces modifications saisonnières sont la cause de variations saisonnières de q.
Les variations tidales liées au rythme des marées sont surtout nettes pour la pêche artisanale des juvéniles au filet fixe où les captures sont effectuées pendant le reflux. La capturabilité est susceptible de varier avec la vitesse du courant puisque Yokel (1969) a montré que si cette vitesse devenait inférieure à 0,31 m/seconde, les crevettes pouvaient nager à contre-courant hors des filets. Les variations saisonnières des courants de marée et les crues des fleuves sont de nature à induire en plus des variations saisonnières de capturabilité pour ce type de pêche.
1 Le terme capturabilité est pris ici dans son sens le plus large
Pratiquement tous les pêcheurs parlent de ces variations liées aux phases lunaires mais elles sont difficiles à mettre en évidence de façon indiscutable en mer. Leur existence a cependant été clairement démontrée par certains auteurs (chapitre 2). Elles sont, en revanche, très évidentes en lagune ou en estuaire pour la pêche des juvéniles en migration ou des maximums principaux et secondaires ont été observés en nouvelle et pleine lune respectivement.
Elles sont connues de longue date pour pratiquement tous les stocks exploités. Elles sont pourtant le plus souvent négligées car, en dynamique classique, l'utilisation d'indices d'abondance moyens annuels, pour l'étude d'espèces à vie longue permet de contourner la difficulté.
En revanche, l'étude d'espèces à vie courte comme les crevettes requiert l'utilisation d'un intervalle de temps inférieur à la période d'oscillations des phénomènes biologiques (l'année), ce qui met en relief le rythme saisonnier qu'il n'est plus possible de masquer par un effet de moyenne au sein d'un intervalle annuel.
Les causes des variations saisonnières de q sont nombreuses. Un premier inventaire a été tenté par Garcia (1977b) que nous reprenons ici en le complétant. Les différentes sources de variations saisonnières sont:
les conditions hydrologiques et le comportement
le recrutement et la sélectivité
les variations d'abondance numérique
les variations de concentration de la pêche.
8.4.1 Les conditions hydrologiques et le comportement
8.4.1.1 La turbidité
On a vu au paragraphe 1 de ce chapitre que les variations nycthémérales de rendement pouvaient donner une image correcte ou inversée du rythme d'activité des crevettes. Garcia et al. (1973) ont montré l'existence de variations saisonnières nettes du rythme d'activité “apparent” dans divers fonds de pêche du golfe de Guinée. Suivant que la turbidité est élevée ou faible ce rythme présente un maximum de jour ou de nuit. Ces auteurs ont montré que le passage d'un comportement “apparent” diurne (rendements plus élevés le jour) à un comportement apparent nocturne (rendements plus élevés la nuit) se traduisait par une baisse de la densité apparente liée à une baisse de la capturabilité. Ils ont également indiqué qu'il existait des saisons à rythme stable et des saisons à rythme instable. Il est vraisemblable que cette instabilité soit également de nature à influer sur l'efficacité de l'effort.
Dans un travail plus récent sur la même région, Garcia (1978) a montré par l'analyse des cohortes l'existence de variations saisonnières de capturabilité synchrones des variations de turbidité (Fig. 59).
Marcille (1978), à Madagascar, a également montré qu'en saison sèche la puissance de pêche relative de certains bateaux est augmentée par les averses qui agissent sur la turbidité des eaux côtières. Ces changements, liés au comportement des crevettes et en particulier aux changements dans l'extension verticale des bancs, peuvent affecter différemment des chaluts dont l'ouverture verticale est différente, perturbant les relations de standardisation (voir paragraphe 4.1.1.2).
8.4.1.2 La température
Son action sur la capturabilité a été envisagée de manière contradictoire. Garcia et al. (1973) ont suggéré qu'au Sénégal, les basses températures hivernales (14–15°C environ) amélioraient la capturabilité globale journalière en supprimant les rythmes d'activité. Les crevettes engourdies seraient alors également capturables de jour comme de nuit.
En revanche, Penn (1976) en Australie a attribué l'augmentation de capturabilité observée à une augmentation de la température.
8.4.1.3 Les courants
On peut rappeler ici, pour mémoire, le cas particulier des crevettes en migration. Lorsque les juvéniles quittent les lagunes ou estuaires, et que la capture s'effectue à l'aide de filets passifs, la capture obtenue par heure de pêche, ou par jour, dépendra d'une part de l'abondance réelle des crevettes dans l'estuaire (grandeur liée au cycle saisonnier de reproduction) mais également du volume d'eau filtrée par unité de temps de pose. Elle dépendra donc de la vitesse du courant et la capturabilité sera plus élevée en période de vives eaux qu'en mortes eaux. Elle sera également plus élevée en période de crue qu'en période d'étiage.
Pour une analyse plus détaillée des problèmes d'évaluation de l'effort de pêche dans ce type de pêcherie on se référera au chapitre 4.1.2.2.
8.4.2 Les variations liées au recrutement et à la sélectivité
Les variations de q d'une cohorte avec l'âge en fonction de la sélectivité sont connues. Les courbes d'évolution sont en général de type sigmoïde, symétriques ou non. Dans le cas d'animaux à vie longue où la classe en cours de recrutement ne représente qu'une faible partie du stock exploité, ce phénomène n'a pas d'incidence saisonnière notable sur la capturabilité globale de la population. Dans le cas d'animaux à vie courte comme les crevettes, où la durée de vie dans la pêcherie est souvent inférieure à un an, la structure d'âge du stock est sujette à des variations saisonnières importantes liées au mécanisme du recrutement (Garcia, 1977). Une illustration de ce phénomène et de son amplitude est donnée par la figure 77. Sur de tels stocks, la combinaison des phénomènes de sélectivité et de recrutement peut entraîner des variations saisonnières de la capturabilité globale de la population et en particulier, une augmentation de q avec l'âge moyen de la population jusqu'à ce que l'essentiel du recrutement soit terminé. Etant donné que la participation d'une cohorte dans la pêcherie est peu supérieure à un an, cette évolution de q avec l'âge liée à la sélectivité se traduit en fait par une variation saisonnière.
8.4.3 Les variations d'abondance numérique
Pour les mêmes raisons que précédemment, l'abondance numérique du stock disponible est sujette à des variations saisonnières d'autant plus marquées que le recrutement est saisonnier. Si l'on considère que tous les recrutements élémentaires dans une année constituent une génération théorique, il est clair que la proportion de cette génération disponible pour la pêche varie au cours de l'année. Ceci implique que la proportion de la génération annuelle totale capturée par une unité d'effort variera avec le temps, passant par un maximum juste après le recrutement puis décroissant jusqu'au recrutement suivant. Ces variations sont assimilables à des variations de q et un effort donné appliquera une mortalité par pêche globale différente, selon la période de l'année considérée.
8.4.4 Les changements d'indices de concentration
Lorsque la ressource est constituée de plusieurs espèces différentes et que l'effort n'est connu que globalement, la variation saisonnière de la distribution relative de l'effort total sur les différentes espèces entraîne des variations saisonnières cachées de q. On verra aux paragraphes 8.5 et 8.6 que les changements de concentration de l'effort sur les différentes cohortes ou groupes d'âge d'une espèce donnée provoquent des variations de q dans l'éspace et avec l'âge.
8.4.5 Conclusions
Les sources de variations saisonnières de la capturabilité, prise ici dans son sens le plus large, sont nombreuses. Bien que la plupart d'entre elles n'aient pas été quantifiées, leurs sens de variation peuvent être qualitativement appréciés. Les variations liées à la dynamique du stock (abondance numérique et âge moyen) ont tendance à s'opposer. Celles qui sont liées aux conditions hydrologiques peuvent se combiner ou s'opposer entre elles suivant les cas et la résultante saisonnière de toutes ces variations est a priori difficile à définir analytiquement. Elle peut cependant être calculée par des marquages successifs ou par analyse des cohortes (voir chapitre 6).
Les conséquences de ce type de variations sont les suivantes:
les variations de F avec l'âge seront différentes pour chaque cohorte élémentaire composant une génération;
la mortalité totale appliquée au stock par un effort annuel donné dépendra de la distribution saisonnière de ce dernier, de même que la prise totale et le rendement par recrue;
le résultat d'une mesure d'aménagement telle qu'une fermeture saisonnière dépendra essentiellement de sa position dans l'année.
Ces variations saisonnières doivent donc être prises en compte autant que possible lorsque les techniques de simulation sont utilisées pour déterminer l'effet probable de variatins dans les strategies de pêche et des mesures d'amènagement.
Dans le cas de la pêche au chalut, elles peuvent être liées aux difficultés du chalutage et dépendent de la configuration du fond, de sa nature, de la présence de végétaux, de courants violents, etc. C'est un problème généralement peu abordé parce que facile à contourner par effet de moyenne, si le schéma de distribution relative de l'effort sur l'espèce considérée ne change pas dans le temps (indices de concentration constants).
Cependant, lorsque la distribution est hétérogène et que la pêche est parfaitement dirigée, comme c'est le cas pour les crevettes grâce au chalut d'essai, l'effort est en général concentré sur les biomasses les plus fortes (Fig. 60). La conséquence est que si en chaque point la capturabilité est la même (car la proportion de l'effectif présent en ce point, capturée par unité d'effort est la même) globalement elle est supérieure dans les zones à forte abondance (car la proportion de l'ensemble du stock qui y est capturée par unité d'effort y est plus élevée). Il en résulte que tout changement dans la distribution de l'effort sur celle de la biomasse entraîne des variations dans la rélation entre F et f. C'est pour résoudre ce problème que Beverton et Holt (1957, page 148) ont crée la notion d'intensité de pêche globale effective (overall effective fishing intensity).
La variabilité spatiale de q est encore plus nette dans les pêcheries artisanales, d'une part à cause de la morphologie des estuaires et d'autre part à cause de la compétition entre engins de pêche. En effet, la quantité capturée par les filets fixes couramment utilisés en Inde et en Afrique, dépend du temps de pose mais aussi et surtout du volume filtré. C'est-à-dire, du flux d'eau au point de pêche. Ce flux étant plus élevé dans les zones étroites, on observe fréquemment une hausse des rendements au niveau des passages étroits (goulets, passes, ponts, etc.). Cela complique d'ailleurs singulièrement la règlementation éventuelle de la pêche car la conséquence d'une réduction du nombre des filets variera suivant la zone considérée et la mortalité totale dépendra de la localisation des engins retirés et de ceux qui restent.
Elles sont d'origines diverses et les variations relatives au phénomène de sélectivité et de recrutement ainsi que les variations liées à l'abondance numérique de la population et à ses variations au cours de l'année ont déjà été signalées aux paragraphes 8.4.2 et 8.4.3. En outre, il est nécessaire de souligner que la conséquence d'une concentration de l'effort sur les abondances les plus élévées est l'existence de variations de q liées à l'âge. On a vu, en effet, au paragraphe précédent que la pêche est concentrée sur les strates bathymétriques et géographiques où la densité est la plus élevée (Fig. 60). Comme il existe une stratification parallèle des tailles et des âges, la conséquence est une concentration de l'effort sur les principales cohortes, pendant la période où leur biomasse est la plus importante (Fig. 61).
La perception de ces variations est importante dans l'optique de l'utilisation des données d'effort annuel dans un modèle global d'évaluation des ressources (type Schaefer ou Fox par exemple).
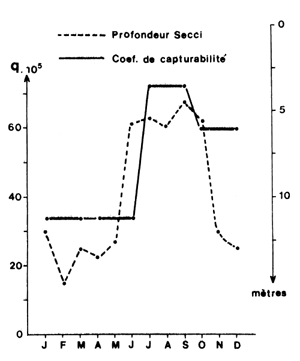
Fig. 59: Variations saisonnières du coefficient de capturabilité et de la turbidité chez Penaeus notialis (d'après Garcia, 1979)
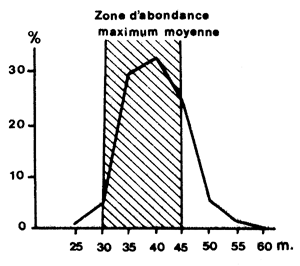 | 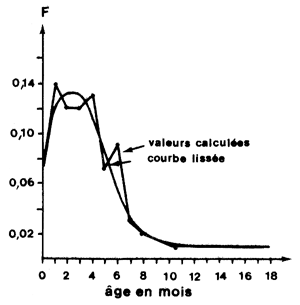 |
Fig. 60: Répartition bathymétrique de l'effort annuel et de l'abondance en Côte d'Ivoire (d'après Garcia, 1977) | Fig. 61: Variations de F avec l'âge chez Penaeus notialis (d'après Garcia, 1977) |
8.7.1 Changement de variations saisonnières de l'effort de pêche
A cause de l'existence de variations saisonnières de q, tout changement dans la distribution relative de l'effort à l'intérieur de l'année aura pour conséquence une modification de la relation en l'effort nominal et la mortalité qui en découle.
8.7.2 Changements d'espèce cible
Lorsqu'il existe plusieurs espèces de penaeïdes mélangées, l'expérience montre que bien qu'elle les exploite plus ou moins simultanément, la pêche est en général plus particulièrement dirigée vers les lieux où les espèces sont les plus rentables et dérivera ensuite vers les espèces secondaires au fur et à mesure que l'abondance des premières diminuera. Un phénomène identique a été décelé au Sénégal où une seule espèce est exploitée mais où le transfert s'effectue de la crevette vers les espèces de poissons accompagnatrices ou existant au voisinage des fonds à crevette (Lhomme, communication personnelle).
Sans une bonne mesure de la répartition réelle de l'effort par espèce l'évolution de ce dernier pour une espèce donnée sera en général mal appréciée et la baisse des rendements des cibles principales sera exagérée. Un exemple de tentative de solution de ce problème en Australie est donné par Hall et Penn (1979).
8.7.3 Modification du rapport entre le temps de recherche et le temps de pêche
Hancock (1979) indique que ce concept classiquement retenu pour les pélagiques est valable pour les crevettes à comportement grégaire très marqué comme Penaeus merguiensis, car la diminution du stock avec les années s'accompagne d'une raréfaction des bancs et donc d'une augmentation du temps de recherche dont l'évaluation devient primordiale au détriment du temps de pêche lui-même qui ne suffit plus à indiquer correctement l'évolution de la mortalité par pêche.
![]()
![]()
![]()