![]()
![]()
![]()
Pour des animaux à vie courte comme les pénaeïdes, où la capture annuelle dépend presque entièrement du recrutement de l'année, l'analyse des variations interannuelles d'abondance et l'établissement de modèles prévisionnels sont essentiels pour permettre un aménagement rationnel de l'exploitation de la ressource. En effet, les conséquences de ces variations sur l'efficacité des différentes mesures d'aménagement proposables sont importantes et seront discutées dans les chapitres suivants (11 et 12).
Dans un premier temps, nous rappellerons brièvement les diverses causes possibles des variations et ensuite nous passerons en revue les diverses corrélations établies à des fins de prédiction dans le cas de pénaeïdes et les mécanismes sous-jacents. Dans ce type de population, les variations les plus importantes sont vraisemblablement liées à des variations du recrutement annuel qui, à cause du nombre très faible de générations supportant l'exploitation, seront directement traduites par des variations pratiquement proportionnelles de la prise totale, si les systèmes d'exploitation restent stables. Les variations de la capturabilité existent et sont certainement en partie responsables des variations saisonnières de capture mais jouent, a priori, un rôle négligeable dans les variations interannuelles.
Il existe également d'autres causes de variations, liées à la dynamique propre des populations et bien qu'elles aient été encore peu ou pas du tout invoquées dans la littérature, il nous est apparu nécessaire de les souligner pour que l'exposé soit plus complet.
9.1.1 Nature des variations
L'exploitation industrielle des crevettes pénaeïdes est un phénomène récent dans la plupart des régions du monde intertropical. La capture de ces 20 dernières années a donc énormément varié, en fonction de l'effort de pêche, des améliorations apportées à la puissance de pêche des flottilles, de l'évolution progressive de la stratégie d'exploitation, du type monospécifique (à la naissance de la pêcherie) au type plurispécifique (lorsqu'elle arrive à maturité), etc. Si l'on ajoute à ceci le fait que les bonnes séries de statistiques de pêche suffisamment complètes et détaillées pour permettre une analyse fine sont peu fréquentes, on comprendra que les données permettant d'étudier la variabilité à long terme des captures sont rares.
L'une des premières questions est donc: l'abondance des stocks de crevettes varie-t-elle ou non indépendamment de l'effort? La réponse est ambiguë. Selon certains auteurs il existe des variations importantes, selon d'autres l'abondance est stable. La stabilité de la production artisanale de certaines lagunes de Côte d'Ivoire pendant des années est un argument en faveur d'un recrutement stable. En revanche, les variations sont importantes dans la pêche artisanale du Sénégal (Le Reste, 1979). De même Boerema (1974) indique que les variations d'abondance en mer sont faibles à Panama, mais fortes au moyen orient.
La différence d'opinions pourrait être liée en partie à la longueur de la série chronologique disponible et il faut vraisemblablement distinguer les variations interannuelles à courte période (ou bruit de fond) des variations à longue période (signal interannuel). Lorsque l'on dispose d'une courte série de données, le signal, souvent masqué par le processus de développement d'une pêcherie mal stabilisée n'apparaît pas et peut d'ailleurs être confondu avec les variations liées à l'effort de pêche, faussant ainsi totalement la relation cpue/effort et l'évaluation qui en découle.
Lorsqu'en revanche, les données disponibles le permettent (Figs. 63–66), le signal interannuel est nettement perceptible. Il est tentant de rechercher dans les modifications climatiques à longue période la cause de ce phénomène et c'est ce que de nombreux auteurs ont fait, comme on le verra plus loin.
Ceci étant, les spectres de variabilité climatique ne sont pas identiques en zone équatoriale et en zone tropicale ou tempérée. Dans ces conditions, il ne serait pas étonnant que l'abondance des stocks de crevettes de ces régions varie différemment. Si l'on examine la figure 63, par exemple, on voit que le bruit de fond, de part et d'autre des signaux interannuels est beaucoup plus important au Texas (± 30 pour cent environ entre 1930 et 1945) qu'en Louisiane (± 15 pour cent environ pendant la même période).
L'exemple utilisé ici l'est seulement à titre indicatif, sans préjugé de notre part quant aux causes réelles de cette variabilité à long et court termes. Notre intention est seulement d'attirer l'attention sur les divers types de variations possibles et leur combinaison car les conséquences sur l'aménagement et la recherche d'une stratégie optimale d'exploitation sont différentes. Il serait utile de caractériser les différents stocks mondiaux par leur spectre de variabilité (amplitude et fréquence du signal interannuel, amplitude du bruit de fond) mais ceci dépasse le cadre de ce travail. Notons cependant également que toute étude de ce type devrait tenter d'établir dans quelle mesure la stabilité “apparente” des captures d'un complexe multispécifique ne reflète pas la capacité d'une flottille à ajuster sa prise totale en ajustant sa stratégie de pêche, plutôt qu'une stabilité réelle du peuplement sous-jacent.
9.1.2 Les variations du recrutement
Le cycle vital de la plupart des penaeïdes comporte une phase littorale importante dont le succès est lié à divers phénomènes.
9.1.2.1 Les variations de la reproduction
Il existe peu d'observations à long terme permettant d'étudier cette source de variations. Nous nous référerons ici essentiellement au travail de Lhomme (1979).
La figure 62 montre l'évolution du pourcentage de femelles mûres dans les captures, en fonction de la taille, de 1973 à 1977. Cette courbe est une information trop intégrée pour qu'un jugement trop précis soit porté mais la figure indique que le phénomène de reproduction varie et que, pour une taille donnée la proportion d'individus observés à l'état mûr dépend des années. S'il n'est pas possible de chiffrer la signification statistique des différences observées, il est clair que 1975 est une année anormale pour les deux stocks de St. Louis et Roxo-Bissagos et que 1977 est nettement anormale pour le dernier stock.
Nous ne savons pas dans quelle mesure ces anomalies dans la reproduction ont affecté le recrutement.
9.1.2.2 Survie et migration larvaire
Après la ponte, les larves doivent gagner les zones peu profondes où elles trouveront l'abri et la nourriture nécessaires à leur survie et à leur croissance. Il est évident que leurs chances de pénétrer dans les estuaires sont étroitement liées aux conditions hydrologiques (courants côtiers, courants de marée, bilans hydriques, etc.).
Les fortes pluies et les crues qu'elles provoquent apparaissent selon les auteurs comme un facteur favorable - car elles augmentent l'extension de la zone à fort gradient de dessalure dans laquelle les larves “pourraient” se diriger - ou défavorable, en empêchant tout simplement l'entrée des larves dans les estuaires.
9.1.2.3 Survie dans les estuaires
Il est clair que c'est au niveau larvaire en mer mais aussi très certainement en estuaire, que s'effectue l'essentiel de la régulation des nombres chez les pénaeïdes côtiers. De nombreux auteurs ont admis que des pluies et des crues importantes étaient de nature à favoriser la survie et la croissance, d'une part en augmentant la surface des nurseries disponibles (mais pour des espèces exigeantes en sels, ce peut être l'inverse) réduisant ainsi la compétition intraspécifique et d'autre part en augmentant la productivité du milieu par un lessivage intensif des alentours.
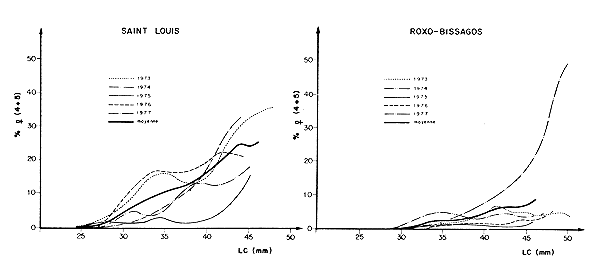
Fig. 62: Variations du % de femelles mûres avec la taille pour Penaeus notialis au Sénégal (d'après Lhomme, 1979b)
9.1.3 Les variations naturelles de la taille à la première capture
Ces variations sont essentiellement liées aux variations, d'une année à l'autre, de la taille moyenne à la migration des jeunes vers la mer.
La biomasse capturable juste avant ou pendant la migration dépend du nombre de postlarves recrutées, de leur survie jusqu'à la migration, de leur croissance et donc de la température rencontrée et de la durée du séjour en estuaire (Ford et St. Amant, 1971; Le Reste, 1979). Ce dernier auteur a montré que la taille moyenne à la migration variait beaucoup en fonction de la salinité ambiante avec, pour conséquence, des variations de biomasse capturable (Fig. 66). Il ne faut pas oublier que, dans ce cas précis, la pêche a lieu avec des filets fixes et que tout retard à la migration se traduit, en fait, par une augmentation naturelle de l'âge à la première capture en estuaire. Le fait que le résultat soit un accroissement de biomasse plaide d'ailleurs pour une mortalité naturelle moins élevée que celle que beaucoup d'auteurs ont calculée (cf. chapitre 7).
Nous insisterons plus loin (cf. paragraphe 10.1.2) sur le fait que le phénomène se répercute en mer. Les crevettes migrant plus tardivement, à une taille plus élevée, ont une courbe de recrutement décalée vers les grandes tailles ce qui se traduit là encore par des variations interannuelles de la taille à la première capture en mer. Bien que les données disponibles ne permettent pas de mesurer les effets de cette variation sur la production annuelle, il est vraisemblable qu'ils ne sont pas négligeables.
9.1.4 Conclusions
Les causes de variations interannuelles de production des stocks de pénaeïdes sont nombreuses. Les relations de cause à effet sont difficiles à démontrer même si elles relèvent parfois du simple bon sens. Il se peut que certains facteurs comme les crues par exemple aient des effets favorables ici, défavorables là, en fonction de leur intensité (phénomène de seuil) ou de la forme de l'estuaire (ria ou lagune, ouverture large ou passe étroite).
Ces variations ayant été décelées dans de nombreuses régions, on a tenté de les prédire à partir de modèles de régression simples considérant le stock marin ou d'estuaire comme une “boîte noire” réagissant à un facteur donné (pluie, crues, température annuelle, etc.) pour produire une biomasse et donc une capture annuelle.
Etablis par corrélation à partir des données historiques, ils ont été étudiés dans le but de parvenir à une prédiction des captures. Le principe est de mettre en évidence une relation quantitative suffisamment précise entre l'abondance et un paramètre biologique ou climatique pour prédire à l'avance les variations interannuelles de capture. Pour être utilisable, le modèle doit permettre une prédiction précise suffisamment tôt pour que l'information soit utile.
Dans le cas des crevettes, la prise annuelle dépend presque entièrement du recrutement de l'année et la prédiction ne peut être faite qu'à court terme. Seule la tendance générale pourra être prédite à long terme si on peut la rattacher à un paramètre climatique périodique (activité solaire par exemple).
On peut distinguer pour les crevettes deux types de modèles suivant que les relations sont établies respectivement avec un facteur climatique (pluie, crues, ensoleillement, activité solaire) ou biologique (en général, l'abondance d'un stade antérieur dans le cycle vital).
9.2.1 Prévisions à partir des données climatiques
9.2.1.1 Relations avec les crues et les pluies
Etant donné l'importance des zones côtières dans le cycle vital des crevettes et le rôle important que semblent jouer les gradients côte-large dans les processus de migration au cours du développement, l'influence des apports continentaux a été étudiée en premier lieu. Les résultats sont contradictoires car les relations mises en évidence sont soit positives soit négatives suivant les cas.
Aux U.S.A. dès 1953, Hildebrant et Gunter (1953), puis Gunter et Hildebrant (1954) montrent qu'il existe pour Penaeus setiferus, au Texas, une bonne relation entre les captures totales et les pluies des deux années précédentes (r = 0,66) ou les pluies de l'année en cours et des deux précédentes (r = 0,70). La relation est significative au niveau 1 pour cent et les prises importantes sont liées aux fortes pluies.
Les auteurs ont envisagé une relation de cause à effet par l'intermédiaire de la dessalure, le temps de latence observé pouvant correspondre au temps de latence entre les pluies et leur répercussion sur la dessalure puis sur la production.
Quelques années plus tard, Gunter et Edwards (1969) confirment ces résultats à l'aide de données plus nombreuses (1927–64) sur la même espèce et dans la même région. Analysant en revanche les données disponibles en Louisiane pour la même espèce, ils ne peuvent mettre en évidence de relation entre les captures annuelles et les déversements du Mississipi. Ils en concluent logiquement que la dessalure doit être, là, toujours supérieure au minimum nécessaire.
Le même type d'analyse appliqué par les auteurs à P. aztecus au Texas et en Louisiane ne donne pas de relation significative et suggère même qu'avec plus de données la relation serait négative, résultat estimé normal pour une espèce que l'on admet adaptée à une zone où les apports en eau douce sont faibles et la salinité élevée.
Ce problème a été de nouveau abordé plus récemment par Barret et Gillespie (1973, 1975) qui indiquent qu'il existe en fait une corrélation négative, en Louisiane, entre la décharge fluviale et les captures si l'on considère la décharge de printemps (pour P. aztecus) et cette d'été (pour P. setiferus)1.
Selon ces deux auteurs, des décharges excessives, dans cette région où elles sont déjà importantes limiteraient les nurseries disponibles, limitant ainsi la production. Ceci serait confirmé, d'après les auteurs, par le fait que la production de P. aztecus dans les estuaires de Louisiane est d'autant meilleure que la salinité est élevée au printemps.
Les apports d'eau douce apparaissent donc favorables à la production au Texas et défavorables en Louisiane et ce pour la même espèce, indiquant que la relation positive entre la dessalure et la production de P. setiferus ne serait pas une fonction continue mais qu'elle présente un seuil au-delà duquel elle s'inverse.
Plus récemment, Lhomme et Garcia (1981) ont montré, au Sénégal, que les anomalies de pluviomètrie et d'abondance en mer étaient positivement correlées pour le stock de St. Louis, où les apports d'eau douce sont faibles) et négativement correlées pour le stock de Roxo-Bissagos où les apports sont plus importants.
Y = 3365 D-0,5064 (1,0 - 0,994746E)
où E est l'effort pêche et D la décharge annuelle. Dans ce modèle, la possibilité suggérée plus haut que la fonction reliant production et décharge annuelle ne soit pas monotone n'a pas été prise en compte.
Le travail de Gunter et Edwards (1969) appelle un autre commentaire en ce qui concerne P. setiferus. En effet, la comparaison des évolutions annuelles des prises en Louisiane et au Texas indique une bonne concordance des variations à longue période (Fig. 63). En conséquence, si les données semblent indiquer une réaction des stocks aux apports fluviaux, différente suivant les régions, il n'en reste pas moins qu'il existe apparemment des oscillations pratiquement synchrones qui laisseraient supposer l'action d'un facteur commun, vraisemblablement d'ordre climatique.
Des variations synchrones sont également montrées par Berry et Baxter (1969) pour les captures annuelles de P. aztecus dans la partie nord-ouest du golfe du Mexique (Fig. 64) mais ce synchronisme pourrait provenir de l'existence de migrations importantes dans tout ce secteur (Barret et Gillespie, 1975).
Il aurait été intéressant de savoir si il y avait aussi synchronisme des variations entre deux espèces différentes mais l'observation positive à ce sujet de Barret et Gillespie (1973) est contredite par les mêmes auteurs en 1975.
En Australie, Thompson (1956) avait, depuis longtemps, mis en évidence une liaison positive entre les variations des pluies et celles des prises annuelles de juvéniles (de 1924 à 1941) sans faire d'hypothèses sur le mécanisme sous-jacent. Il a observé, en particulier, une bonne relation entre la prise et les pluies des deux années précédentes (r = 0,76), ou les pluies tombées 6 à 12 mois avant le début de la saison de pêche. Il a également montré, sur six ans seulement (1948–53), les variations similaires des pluies et des prises d'adultes en mer l'année suivante, dans la même région.
Cette observation préliminaire est confirmée par Ruello (1973) avec une série de données plus longue (Fig. 65). Il a observé les meilleures corrélations avec les pluies de l'année précédente (r = 0,64) ainsi que celles de la même année plus celles de l'année précédente (r = 0,69). Les débits des rivières montrent également une bonne relation positive.
L'observation par Thompson (1956) d'une rélation positive entre les pluies et la production de juvéniles dans les estuaires est en contradiction avec les observations de Castello et Olinto Möller (1978) au Méxique, Le Reste (1979) au Sénégal (Fig. 65), et Barret et Gillespie (1975) en Louisiane qui montrent une relation négative entre la pluviométrie et la production en estuaire respectivement au Mexique et au Sénégal (Fig. 65).
Là encore, les observations disponibles à travers la littérature indiquent une influence favorable ou défavorable suivant les régions ou les espèces.
Plusieurs hypothèses ont été proposées pour expliquer les relations observées: action directe sur la fécondité, sur la migration larvaire facilitée par une augmentation des gradients de dessalure ou bloquée par les crues excessives, sur l'habitat disponible en estuaire et donc sur la compétition, la croissance, la survie, etc., toutes ces actions pouvant d'ailleurs être concomitantes.
L'apparente généralisation de variations à longue période à une grande région du golfe du Mexique laisse supposer l'action de facteurs climatiques agissant à grande échelle et dont les modes d'actions seront très difficiles à déterminer en raison de la complexité du cycle et des écosystèmes dans lesquels elle se déroule.
L'ensemble des hypothèses proposées suppose que l'action climatique a lieu au niveau du recrutement dont elle commande les effectifs. Certains facteurs, comme les crues, apparaissant favorables ou défavorables suivant les travaux, on peut penser que la relation n'est peut-être pas linéaire et que si la production est accrue dans une certaine gamme, elle est freinée au-delà d'un certain seuil qui dépend des conditions locales.
Un mécanisme sensiblement, différent, faisant intervenir des variations naturelles de l'âge à la première capture liées à la salinité; peut être proposé au vu des résultats de Le Reste (1979). Cet auteur a étudié les variations de production d'une importante pêcherie artisanale en Casamance (Sénégal), estuaire sursalé une partie de l'année. Il a montré que l'évolution de la taille moyenne annuelle des crevettes capturées, observée depuis le début de la sécheresse sahélienne, était liée à celle de la salinité de l'estuaire. Il a également montré que les années sèches, où la taille à la migration (et donc à la première capture) est élevée, la production est forte et vice versa. La salinité de la Casamance, alimentée surtout par la nappe phréatique, dépend des pluies des années précédentes et Le Reste a montré une bonne corrélation négative entre les prises d'une année et les pluies des années précédentes. Ceci est interprété comme une action de la salinité sur la durée du séjour en estuaire, hypothèse avancée et étayée par Garcia (1977) pour la Côte d'Ivoire et le Nigeria. Le séjour plus long permettrait une meilleure production, par le jeu de la croissance et de la mortalité. La relation utilisée comme modèle a permis d'obtenir une bonne concordance entre les prises observées et prédites (Fig. 66). La qualité et la fiabilité réelles de la prédiction devront être testées dans les années futures.
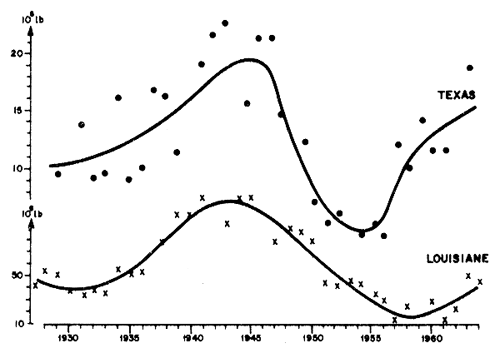
Fig. 63: Variations à long terme des débarquements de crevette Penaeus setiferus au Texas (points) et en Louisiane (croix) (d'après des données de Gunter et Edwards, 1969)
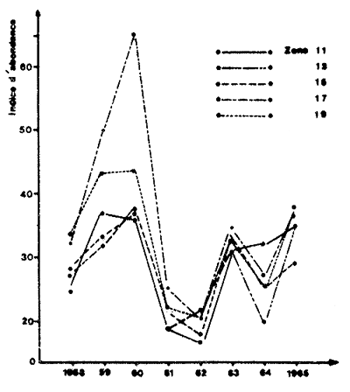
Fig. 64: Variations interannuelles de l'abondance de Penaeus aztecus dans différentes zones statistiques du nord et nord-ouest du golfe du Mexique, de l'Alabama au Mexique (d'après Berry et Baxter, 1969)
Il faut noter enfin que certains auteurs ont cherché à relier les captures et les pluies, mois par mois, avec un certain décalage. La corrélation est en général excellente car on relie entre eux deux phénomènes saisonniers, mais elle ne peut être utilisée pour prédire les captures totales annuelles.
9.2.1.2 Relation avec la température
Williams (1969) a montré, aux Etats-Unis, une relation entre le “contenu thermique” d'une année et l'abondance exprimée par la prise par unité d'effort. L'introduction d'un décalage dans le temps n'améliore pas sensiblement la relation. Lorsqu'il regroupe les données de la Georgie, des Carolines et du Texas, les années chaudes correspondent nettement à des captures élevées. Cet auteur note cependant que les relations, bien qu'apparentes, sont moins bonnes si on considère les régions séparément.
Cette relation semble, en fait, peu utilisable pour prédire les variations interannuelles dans chaque région prise séparement, avec une bonne précision, et n'être significative qu'à l'échelle de toute la face sud-est des Etats-Unis car c'est alors seulement que la gamme des bilans thermiques annuels est suffisamment étendue pour que se dégage une relation de type morpho-édaphique qui indique, en fait, que la richesse globale en crevettes d'une région est proportionnelle à son bilan thermique annuel (Fig. 67). Ceci se comprend aisément quand on sait que les mécanismes essentiels de la croissance et de la reproduction de ces animaux tropicaux sont liés à la température. L'auteur a d'ailleurs signalé que l'échelonnement des données le long de la droite d'ajustement correspond à un étagement latitudinal. Ces observations sont à rapprocher de celles de Turner (1977) reprises au paragraphe suivant.
Une relation de type différent a été établie au Texas par Baxter (1969) qui montre la liaison existant entre la date du pic de migration annuelle des juvéniles vers la mer et la température de l'air, en avril de la même année (Fig. 68). Cette prédiction peut être utile pour déterminer avec précision la période pendant laquelle devrait s'effectuer l'échantillonnage visant à définir un indice d'abondance des juvéniles en migration. On peut citer, dans le même ordre d'idées, le travail effectué en Louisiane (Barret et Gillespie, 1973, 1975) qui indique que la capture en estuaire en mai dépend de la température en avril pour P. aztecus.
9.2.1.3 Relation avec la latitude
Turner (1977) a montré l'existence, au niveau mondial, d'une relation entre la production (Y) et la latitude (x, en degrés), de la forme
Y = 159e-0.070(x).
9.2.1.4 Relation avec l'activité solaire
Bien que ces études n'aient pas été réalisées sur les crevettes penaeïdes, il est intéressant de noter que Driver (1979) a montré la bonne corrélation existant entre la capture de Crangon crangon (crevette caridae) et le rythme d'activité solaire. Une étude antérieure avait montré qu'une prédiction pouvait être faite un ou deux ans à l'avance en se basant sur la pluviométrie et la température de l'air. Les variations interannuelles du climat étant bien liées à l'activité solaire, l'auteur a donc cherché et a obtenu une bonne corrélation à ce niveau (r = 0,86) avec une période de 11 ans.
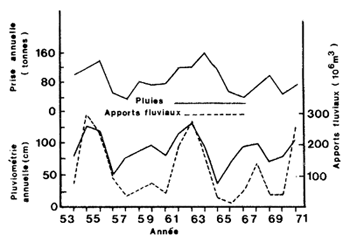
Fig. 65: Variations interannuelles des pluies, des apports fluviaux et de la capture annuelle chez Metapenaeus mackleyi (d'après Ruello, 1973)
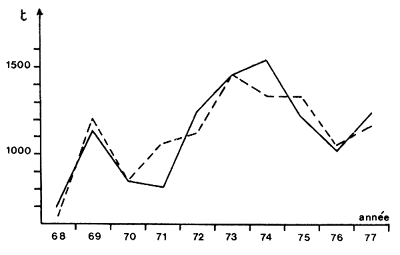
Fig. 66: Evolution des prises réelles (---) et simulées (—) dans la pêcherie de juvéniles de Casamance (Sénégal) (d'après Le Reste, 1979)
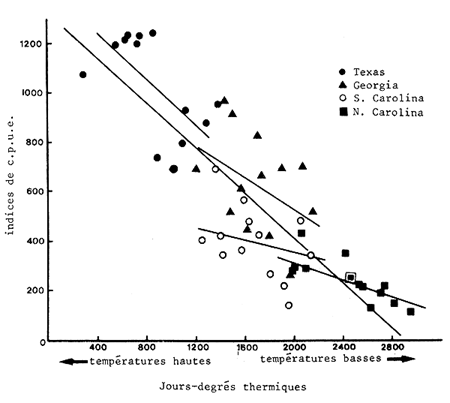
Fig. 67: Relation entre le bilan thermique annuel et les indices d'abondance (d'après Williams, 1969)
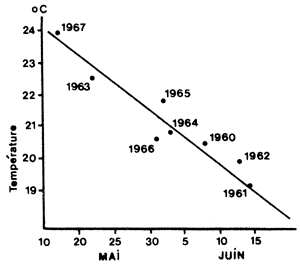
Fig. 68: Relation entre la température en avril et la date du maximum d'abondance des juvéniles dans la baie de Galveston (d'après Baxter, 1969)
9.2.1.5 Relation avec la surface des zones intertidales
Turner (1977) a montré également que la capture en lagune au niveau régional était liée à la superficie des zones intertidales. Il admet d'ailleurs également que cette relation reflète peut-être simplement la relation entre la production de l'estuaire et l'importance de “l'interface” mer-estuaire. De même Barret et Gillespie (1973, 1975) ont montré une relation entre les variations interannuelles de superficie d'estuaire propices à la vie des crevettes (salinité supérieure à 10 ppm) et celles de la capture.
9.2.1.6 Conclusions
Il existe suffisamment d'indices prouvant que la production annuelle des crevettes varie d'une année à l'autre en relation avec des facteurs climatiques à grande échelle. On peut supposer qu'il y a là une relation sous-jacente de cause à effet qui sera toujours très difficile à prouver car les mécanismes sont certainement complexes et concernent les modifications par les conditions climatiques de nombreux paramètres (fécondité, recrutement post-larvaire, chronologie des migrations, croissance et survie, capturabilité).
Les paramètres climatiques mesurés, qu'il s'agisse des pluies, des crues, du bilan thermique annuel ou de l'activité solaire, peuvent n'être finalement que des indicateurs globaux de variations climatiques affectant bien d'autres paramètres encore (ensoleillement, donc production primaire et photopériode, salinité, chronologie ou longueur relative des saisons, déplacement de fronts, position ou force des courants et sous-courants) tout aussi susceptibles d'influencer la production biologique des stocks.
Mais il n'est finalement pas nécessaire de comprendre les mécanismes pour utiliser une relation empirique à titre prédictif si la précision du diagnostic est jugée suffisante. C'est vraisemblablement en ce sens que les relations précédentes ont été établies. Il faut cependant souligner que selon Marr (1972) ce type de relation n'est souvent précis que dans les extrêmes et que, dans les conditions moyennes, la variance la plus forte est le plus souvent liée à des facteurs biotiques (compétition, etc.). Il en découle qu'un paramètre donné peut n'être suffisamment explicatif que dans les zones où il est un facteur limitant et qu'ainsi seules les années anormales seront prédites avec une précision suffisante.
La relation n'étant pas forcément linéaire, il convient de ne pas effectuer de prévision en déhors de l'intervalle des données déjà observées.
Le cycle vital des crevettes passe par plusieurs phases, larvaire, postlarvaire, juvénile, adulte, dont deux, correspondant aux migrations, sont des phases de concentration, facilement échantillonnées. Depuis les années 60, de nombreux travaux ont tenté de mettre en évidence des relations entre l'abondance d'une phase et celle d'une phase ultérieure exploitée, de façon à disposer d'un indice de pré-recrutement et d'un modèle prévisionnel simple.
9.3.1 Relation entre l'abondance des adultes et celle de leur progéniture larvaire et postlarvaire
D'après Neal (1975) aucune relation n'a pu être mise en évidence. Selon lui, cela indique que la variabilité à ce stade est essentiellement liée à l'environnement. Aucune observation à ce sujet n'est disponible, à la connaissance des auteurs. Il est cependant exact que le phénomène de reproduction est susceptible de fortes variations d'une année à l'autre (paragraphe 9.1.2.1).
9.3.2 Relation entre l'abondance des larves en mer et celle des adultes de la même génération
Toujours selon Neal (1975) aucune relation n'a été observée. Il faut noter que le principal handicap réside dans l'obtention d'un indice suffisamment précis d'abondance larvaire en mer, à un coût raisonnable. Les problèmes rencontrés découlent d'une part du type de distribution du plancton (cf. chapitre 2) et d'autre part du développement rapide (accompagné d'une mortalité larvaire élévée) à travers les nombreux stades larvaires successifs qui caractérisent le développement des penaeïdes. En effet, à un instant précis, l'abondance d'un stade donné (protozoé par exemple) dépend de l'intensité de la ponte qui lui a donné naissance mais aussi du temps écoulé entre la ponte et l'échantillonnage, en raison de la migration permanente dans et hors du stade considéré et de la mortalité larvaire élevée.
9.3.3 Relation entre l'abondance des postlarves et celle des juvéniles ou des adultes
Lorsque l'on dispose de suffisamment de données concernant les variations d'abondance de deux stades successifs au cours de l'année, il est possible de relier entre eux les maximums d'abondance (Fig. 69).
Lorsque l'on dispose ensuite de longues séries d'abondance de divers stades du développement sur plusieurs années, on peut calculer des indices d'abondance réspectifs dont la corrélation, si elle est précise, peut servir à des fins de prédiction (Fig. 70).
Baxter (1963), au terme d'une étude de trois ans sur P. aztecus au Texas, montre une bonne corrélation des variations saisonnières d'abondance des postlarves, des juvéniles en lagune et des adultes. Il suggère qu'une prédiction semble possible mais que seule une étude approfondie peut permettre de l'affirmer (Fig. 69). Plus tard, Berry et Baxter (1969), après sept années d'études dans la même région, notent l'absence de relation utilisable entre l'abondance des postlarves et celle des captures. Ils suggèrent que l'échantillonnage au filet Renfro (cf. 2.3.2.4) est à revoir. Ces résultats sont corroborés par ceux de St. Amant et al. (1965) qui montrent qu'en Louisiane la relation entre l'abondance des postlarves et celle des juvéniles n'est pas excellente, ce qui indiquerait que la production dépend beaucoup de la survie des larves en estuaire et qu'une importante régulation des nombres se produirait à ce stade là. Subrahmaniam (1966) et Subrahmaniam et Rao (1970) montrent que pour deux années successives l'abondance des postlarves et celle des captures varient dans le même sens. Ces travaux portent cependant sur des périodes trop courtes pour juger de la valeur quantitative de la relation. Il en est de même pour le travail de Christmas et al. (1966) qui porte sur deux ans, dont les résultats sont cependant confirmés par Christmas et al. (1976) qui obtiennent, sur trois ans, une prédiction valable à 10 pour cent près.
Plus récemment encore, après une étude sur deux ans, Roessler et Rehrer (1975) ont montré une bonne relation entre les variations saisonnières d'abondance des postlarves dans les Everglades et celle du recrutement des subadultes en mer. La corrélation élevée qu'ils ont observée (r = 0,78) est normale car elle est établie entre deux phénomènes saisonniers.
En définitive, en dehors du travail de Berry et Baxter il n'existe pas, à notre connaissance, d'autres exemples probants portant sur une période suffisamment longue, et le problème de l'utilité pratique réelle des indices d'abondance postlarvaires, qui peut d'ailleurs varier suivant le type d'écosystème considéré reste donc posé.
9.3.4 Relation entre l'abondance des juvéniles et celle du stock adulte en mer
Yokel et al. (1969) et Yokel (1969) sur P. duorarum en Floride montrent que si l'on tient compte des variations dans les temps de latence, découlant des variations saisonnières de la taille à la migration, on obtient une bonne corrélation entre l'indice numérique d'abondance des juvéniles en migration et celui des jeunes recrutés sur le fond de pêche (Fig. 70). Les rendements de la pêche sur les juvéniles peuvent donc servir d'indices de pré-recrutement. Berry et Baxter (1969) sur P. aztecus au Texas, ont montré qu'il y avait une bonne rélation entre l'abondance des juvéniles dans la pêche en estuaire (du 25/4 au 31/7) et l'abondance des adultes en mer (de juillet à septembre). Caillouet et Baxter (1973) indiquent les équations des relations obtenues pour le Texas et la Louisiane. Au Texas, la relation est excellente (r = 0,85) pour P. aztecus si l'on relie la prise annuelle en mer à l'indice d'abondance des juvéniles en baie de Galveston1, et médiocre (r = 0,28) si on relie cette prise à la prise annuelle en estuaire2. La relation est moyenne (r = 0,58) pour P. setiferus avec ce même type de données. En Louisiane, la relation entre la prise annuelle en estuaire et la prise annuelle en mer est excellente pour P. aztecus (r = 0,73) et P. setiferus (r = 0,86), résultat confirmé par Barret et Gillespie (1975).
1 Cet indice est calculé entre le 25 avril et le 12 juin de chaque année, pendant la période de
migration le plus importante
2 Selon les auteurs ceci serait dû à l'effet de la réglementation sur les captures imposée dans les
estuaires du Texas
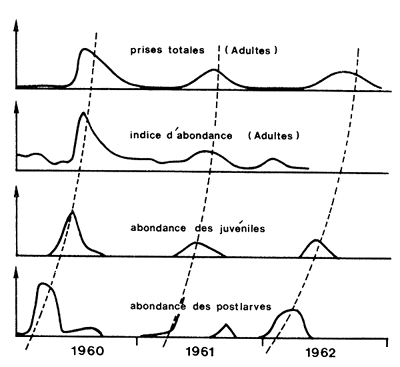
Fig. 69: Relations entre les variations saisonnière des indices d'abondance des postlarves, juvéniles et adultes (simplifié de Baxter, 1963)
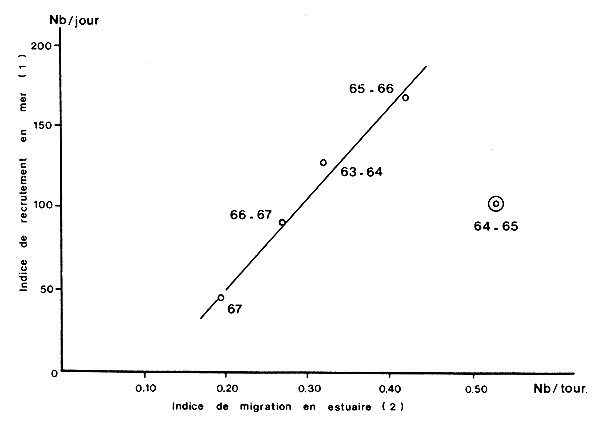
Fig. 70: Relation entre la migration et le recrutement (modifié de Yokel, 1969)
1. Prise annuelle moyenne/jour des subadultes (68/livre et au-delà)
2. Indice d'abondance annuel des subadultes en migration (Nb/tour de
fluxmètre)
Les corrélations entre prises annuelles en estuaire et en mer indiquent que les bonnes années en estuaire sont aussi de bonnes années en mer (et vice versa) mais ne peuvent pas servir pour des prédictions car les deux variables portent sur la même période de temps.
9.3.5 Conclusions
Lorsque l'on cherche à relier la production d'une année à l'abondance d'une des phases antérieures du cycle vital, il semble que les subadultes en migration donnent un indice utilisable pour des prédictions à court terme (environ 3 mois à l'avance). L'utilité des indices d'abondance postlarvaire reste encore à confirmer avec une longue série de données.
En effet, les conclusions optimistes de Berry et Baxter (1969, p. 797) quant à l'utilisation possible de l'abondance postlarvaire pour des prédictions n'ont été ni réellement confirmées ni infirmées car aucun autre travail de cette envergure n'a été entrepris. Compte tenu du coût relativement élevé d'obtention de ces données par rapport à celles de l'exploitation des juvéniles, il n'est pas certain qu'un tel programme soit remis sur pied un jour et puisse être poursuivi en routine.
![]()
![]()
![]()