![]()
![]()
![]()
La vie d'une espèce exploitée peut être schématiquement divisée en deux éléments principaux, une phase non disponible et une phase exploitable. On peut ranger, dans la première, la période qui va de la ponte à l'entrée sur le fond de pêche et, dans la seconde, celle qui va de cette entrée à la fin de l'existence. Pour des raisons pratiques, la modélisation de la pêche - dans le but d'évaluer les ressources ou de définir les conditions optimales d'exploitation - prend seulement en considération la phase exploitable, l'information provenant de la phase précédente étant réduite au nombre d'individus survivants à la fin de cette phase et qui constitue le recrutement. Lorsque l'information n'existe pas, on considère que le recrutement est constant; lorsqu'elles sont connues on prendra en considération les variations saisonnières et interannuelles de ce paramètre, dont l'importance pour la modélisation est considérable.
On appelle également “recrutement” le processus par lequel un groupe d'âge s'intègre pour la première fois dans le stock exploitable. C'est un phénomène plus ou moins progressif qui est fonction de l'âge (ou de la taille).
La présence d'un groupe d'âge dans une zone de pêche n'implique pas qu'il soit immédiatement capturable par les moyens d'exploitation mis en oeuvre, et on appelle sélection, le processus par lequel il entre dans la phase exploitée. Lorsque les deux processus de recrutement et de sélection se produisent dans des intervalles de taille qui se chevauchent, le processus d'entrée dans la phase exploitée sera finalement leur résultante.
Dans ce chapitre, on abordera d'abord la courbe de recrutement, puis celle de sélection pour discuter finalement de la relation entre le stock et le recrutement.
10.1.1 Recrutement et cycle vital
La première phase de l'existence des pénaeïdes côtiers se déroule en mer pendant trois semaines à un mois (à l'état larvaire) puis dans les zones littorales, les fonds de baie, les estuaires, sur des fonds de mangrove riches en nourriture ou dans des herbiers immergés (à l'état juvénile et subadulte). Au fur et à mesure que leur développement s'accomplit les crevettes migrent vers des profondeurs de plus en plus élevées. Lorsque les zones de répartition géographique des juvéniles et des adultes sont nettement séparées, il se produit une réelle migration vers la mer où se fera la reproduction (certains pénaeïdes se reproduisent cependant dans les estuaires (cf. chapitre 2)).
C'est par ce processus que s'effectue le recrutement, mécanisme par lequel les crevettes entrent dans les zones d'action des différentes pêcheries. Elles entreront ensuite dans la phase exploitée par un processus assez semblable à de nombreux égards: la sélection (cf. paragraphe 9.2).
Les modes d'exploitation (et les zones d'action des engins de pêche) étant très diversifiés, il existe en fait plusieurs phases successives de recrutement:
lorsque les crevettes quittent les bords des nurseries et deviennent accessibles à la pêche artisanale
lorsqu'elles gagnent les grandes baies où elles sont accessibles aux petits chalutiers
pendant la migration où elles sont capturées alors par les filets fixés
lorsqu'elles arrivent en mer, où elles sont enfin capturées par les chalutiers industriels.
Le processus d'entrée dans les différentes pêcheries est lié aux stades de développement des crevettes. Si on définit le recrutement comme la probabilité pour une crevette d'une taille donnée de se trouver dans le champ d'action d'un engin de pêche, on peut l'exprimer par le pourcentage de crevettes de cette taille dans l'ensemble de la population (en estuaire et en mer) present sur la zone de pêche. Si ces pourcentages sont portés en fonction de la taille, on obtient la courbe de recrutement.
(a) Recrutement dans la pêche artisanale
Bien que ce type de courbes n'ait jamais été calculé pour la phase estuarienne d'exploitation, on imagine aisément qu'elles sont unimodales et proches de celles que l'on observe pour les filets maillants (voir Gulland, 1969, par exemple). En effet, le pourcentage susceptible d'être capturé croit d'abord au fur et à mesure que la taille augmente et que les crevettes deviennent disponibles, passe par un maximum puis décroît avec la migration des crevettes les plus grosses vers le type d'exploitation suivant. L'information existante sur ce sujet est à notre connaissance pratiquement nulle malgré l'importance de ce phénomène pour la modélisation de l'exploitation de la phase estuarienne.
(b) Recrutement en mer
La phase de recrutement des estuaires vers la mer est mieux connue et on se reportera au paragraphe 2.3.4 pour un résumé des informations sur les variations saisonnières de ce phénomène. Elles sont importantes et représentent une donnée de base pour les modèles de simulation (chapitre 11). Dans le plupart des cas, le recrutement dure toute l'année mais présente des variations saisonnières plus ou moins marquées. On peut les matérialiser par les variations d'abondance des crevettes en migration (cf. paragraphes 2.3.4.2.3 et 2.3.4.3) ou par l'abondance de juvéniles dans les captures des chalutiers en mer (Fig. 6). Il est vraisemblable que ce recrutement se produit lorsqu'un stade donné de développement est atteint ou lorsque des conditions défavorables en estuaire l'imposent. Il se produit donc à un âge variable, dépendant des conditions de milieu. Pour les grands pénaeïdes cet âge est d'environ 2,5-4 mois (Kutkuhn, 1966 pour P. duorarum, Garcia, 1970, 1977 et Galois, 1975 pour P. notialis, Baxter, 1967, pour P. aztecus, Le Reste, 1971, 1973, pour Penaeus indicus, etc.).
10.1.2 La courbe de recrutement en mer
La donnée généralement recherchée est la taille (et l'âge) au recrutement (lr, tr). On l'obtient en traçant la courbe de recrutement indiquant pour chaque taille le pourcentage de l'ensemble de la population qui est présent sur le fond de pêche (Fig. 72). La courbe peut théoriquement revêtir des formes variées mais elle est le plus souvent sigmoïde. La taille moyenne au recrutement (lr)est celle pour laquelle le pourcentage total des individus de taille inférieure à lr déjà recrutés est égal au pourcentage des individus de taille supérieure à lr non encore recrutés. Lorsque la courbe est symétrique, on admet en général lr = lr50, la taille pour laquelle 50 pour cent des individus sont recrutés.
Par nature, la courbe est difficile à définir car les informations quantitatives nécessaires sont très difficiles à obtenir et c'est la raison pour laquelle, à notre connaissance, il n'en existe aucune dans la littérature concernant les penaeïdes. On peut cependant l'approcher: si l'on sait que l'on ne trouve presque jamais en mer de crevettes de taille inférieure à, disons, 15 mm (LC) et qu'on ne trouve jamais en estuaire de crevettes de taille supérieure à 26 mm (LC), on a donc les deux bornes 0 et 100 pour cent de la courbe de recrutement. Entre ces deux points, la courbe de recrutement peut être approchée par une sigmoïde ou une droite. Cette dernière solution a été choisie par exemple par Lhomme (1978) pour Penaeus notialis au Sénégal. La courbe sigmoïde pourrait être approchée par la courbe des fréquences cumulées de la distribution des tailles des crevettes en migration lorsque celle-ci est connue.
Il est bon de rappeler que la taille à la migration et donc au recrutement varie saisonnièrement et d'année en année en fonction des conditions climatiques (cf 2.3.4.2.4) et que ce phénomène est responsable des variations naturelles de la taille à la première capture dans certaines pêcheries artisanales. Le cas des crevettes P. notialis de Casamace (Sénégal) est particulièrement net. Celle-ci varie de 24,3 mm (LC) en période de pluviométrie normale (Lhomme, 1978) à 30 mm (LC) en période sèche. Cette variation équivaut à une modification du vecteur de mortalité par pêche en fonction de l'âge dont les conséquences sur la production annuelle ne sont pas négligeables (cf. paragraphe 9.2.1.1).
10.1.3 La relation stock-recrutement
Cette relation, qui doit être normalement établie entre le nombre de géniteurs dans une population et le nombre de géniteurs qui leur ont donné naissance, est d'une importance capitale pour la modélisation des stocks exploités. Elle peut, selon les espèces, être asymptotique (c'est le type décrit par Beverton et Holt pour les poissons plats de la Mer du nord), en dôme (c'est la courbe classique de Ricker, que l'on rencontre chez les saumons) ou pratiquement-linéaire (comme l'indique Cushing, 1971 et 1973, pour les clupeïdes). La forme de cette courbe conditionne la résistance d'un stock à l'exploitation puisque le pouvoir de reproduction1 sera plus ou moins affecté ou même amélioré (dans le cas de la courbe en dôme) par l'exploitation.
Dans le cas des crevettes penaeïdes, on n'a jamais observé de stock dont le recrutement ait été, avec certitude, affecté par la pêche sur les adultes même sur des stocks très exploités sauf peut-être récemment dans le golfe entre l'Iran et la peninsule Arabique (Van Zalinge, communication personnelle). Ceci laisserait supposer que la courbe est de type Beverton et Holt, pour laquelle le pouvoir reproducteur reste constant même pour des réductions importantes de la population mère. C'est en tous cas ce que suggèrent Neal (1975) et Le Reste et Marcille (1973) respectivement aux U.S.A. et à Madagascar.
On tend plus généralement à admettre que le recrutement n'est pas lié à l'abondance des reproducteurs dans les gammes normales de niveaux d'exploitation et qu'il paraît plus affecté par les conditions du milieu (voir chapitre 9) et la dégradation des estuaires (voir paragraphe 12.4.1.3).
On a vu, au chapitre précédent, que le recrutement des crevettes dépendait vraisemblablement pour une bonne part, des conditions hydrologiques et de leurs répercussions sur la reproduction, la survie des larves pendant leur trajet vers les estuaires ainsi que la croissance, la compétition et la durée du séjour en lagune. On verra d'autre part plus loin (12.4.1.3.3) que la production annuelle de certains stocks paraît nettement affectée par la destruction des biotopes d'estuaires par l'activité humaine (Doï et al., 1973). Tous ces éléments laissent entrevoir une action de régulation puissante au niveau de la phase estuarienne et pourraient expliquer que l'on n'observe pas de relation entre l'abondance du stock et le recrutement. Gunter (1966) supposait déjà que l'exploitation ne pourrait affecter le renouvellement du stock et rappelait qu'après la destruction presque totale des ressources crevettières de la côte Atlantique des Etats-Unis par la vague de froid de l'hiver 1940, les captures de l'automne 1941 furent normales, indiquant par là que les quelques survivants suffirent à repeupler le stock2.
Bakun et Parrish (1981) indiquent d'ailleurs que si les nurseries ont une capacité biologique limitée et relativement constante la relation de type Beverton et Holt est la plus appropriée. Cette remarque nous paraît pouvoir parfaitement s'appliquer aux crevettes penaeïdes côtiers étant toutefois bien entendu que la capacité biologique des estuaires n'est que “relativement” constante et varie en fonction des conditions climatiques. Ces variations étant importantes, il faudrait dans le cas où les crues sont par exemple le facteur décisif, définir non pas une relation stock-recrutement mais une famille de courbes correspondant aux diverses valeurs possibles des crues.
Sur un plan pratique, cependant, s'il n'existe pas de relation entre le stock et le recrutement pour des niveaux raisonnables d'exploitation, cette relation peut être totalement négligée et le recrutement peut alors être considéré comme dépendant seulement de l'environnement. Cette conclusion, même si elle n'est jamais clairement exprimée, sous tend en fait toutes les utilisations de modèles prévisionnels basées sur les relations entre la production et les conditions climatiques (voir chapitre précédent).
Notons que Abramson et Tomlinson (1972) ainsi que Geibel et Heiman (1976) concluent également à l'absence de relation stock-recrutement utilisable pour les crevettes pandalides de Californie, bien que la biologie des espèces concernées et, en particulier, les contraintes liées au recrutement soient totalement différentes.
Cette hypothèse d'un recrutement relativement indépendant du stock serait contredite par les observations de Cushing (1973) qui relie la forme de la relation stock-recrutement à la fécondité individuelle moyenne d'un stock. Chez les grands penaeïdes la fécondité individuelle partielle varie de 100 000 à 500 000 oeufs environ selon l'âge. La fécondité partielle moyenne serait de l'ordre de 200 000 oeufs. On ne connaît cependant pas le nombre d'oeufs réellement pondus (bien que selon Penn, 1980, il y ait peu d'ovules atrésiques en conditions naturelles) parce qu'une crevette pond plusieurs fois dans l'année et que sa fécondité totale individuelle n'est pas connue. Elle est en tout cas certainement très supérieure à 200 000. Il faudrait donc admettre, sur ces bases, que la relation stock-recrutement chez les penaeïdes pourrait être du type en dôme de Ricker, caractéristique des animaux à grande fécondité. Beverton et Holt (1957) ont d'ailleurs attribué cette forme en dôme au cannibalisme, comportement très caractéristique des pénaeïdes en élevage au moment des mues.
Ces remarques toutes spéculatives ne peuvent cacher le fait que l'on ignore totalement la forme de l'éventuelle relation.
Il n'en reste pas moins que les penaeïdes sont le plus souvent soumis à une exploitation intensive qui commence avant, et le plus souvent bien avant, la taille de première maturité sexuelle en particulier lorsque l'exploitation artisanale ou semi-industrielle des juvéniles est autorisée. Dans ces conditions, le potentiel de reproduction est fortement réduit et même si la relation stock-recrutement n'est pas connue, il peut tomber à des niveaux où une grande prudence est de rigueur.
La notion de fécondité théorique par recrue (Le Guen, 1971) présente ici un intérêt dans la mesure où, exprimée en valeur relative par rapport au stock vierge, elle permet de se rendre compte de la réduction du potentiel sous l'action de l'exploitation, pour divers niveaux de la mortalité par pêche.
La figure 71 donne un exemple concernant le stock de P. notialis de Côte d'Ivoire, où la fécondité théorique (en pourcentage de celle du stock vierge) est evaluée en fonction du taux d'exploitation en lagune (en abscisse) et de l'éffort de pêche simultané en mer (de 0 à 3 600 jours de pêche d'un crevettier de 280 ch par an. Le point corréspondant à la situation de 1973 est indiqué.
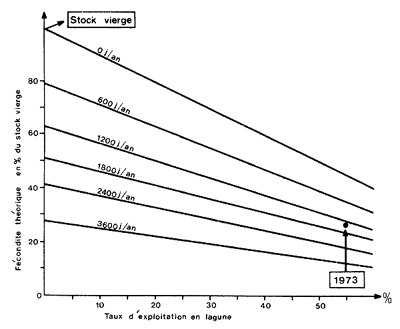
Fig. 71 Evolution de la fécondité théorique en fonction du taux d'exploitation en lagune (en %) et de l'effort en mer pour Penaeus notialis en Côte d'ivoire (d'après Garcia, 1978). Le point indique la situation en 1973.
La sélection est le processus par lequel les crevettes recrutées entrent progressivement dans la phase exploitée. Pour différencier ce processus de celui qui consiste à sélectionner une espèce cible ou un groupe d'âge cible par application préférentielle de l'effort de pêche, on peut préférer le terme de rétention. Le taux de rétention d'un poisson de taille donnée est la probabilité qu'il soit retenu par le chalut (une fois qu'il l'a rencontré et qu'il y est entré). On l'exprime par le pourcentage de poissons retenus dans le cul de chalut pour chaque taille. Le phénomène est, en fait, de même nature que le recrutement et se traduit par une courbe sigmoïde plus ou moins symétrique. Cette courbe est obtenue, en général, en comparant la distribution de fréquence de taille des crevettes retenues dans le chalut à la distribution de l'ensemble de la population ayant pénétré le chalut. On utilise le plus souvent la méthode de la double poche ou bien le chalutage avec deux engins, traînés simultanément ou alternativement, dont l'un est sélectif et l'autre est censé retenir tous les animaux. La grandeur généralement recherchée est l'âge moyen de sélection = ts qui est aussi souvent l'âge moyen d'entrée dans la prise (ou l'âge de première capture, tc) lorsqu'il n'y a pas d'interférence avec le phénomène de recrutement (cf. paragraphe 10.3). Dans la pratique, on utilise une courbe de rétention en fonction de la taille et la taille ls est définie comme le point où le nombre de poissons retenus par le chalut, de taille inférieure à ls, est égal au nombre de poissons relâchés de taille supérieure à cette taille. Si on définit par Yi les pourcentages de rétention observés pour chaque classe de taille, par ln+1 la borne supérieure du premier intervalle de taille où la valeur de 100 pour cent est obtenue et si l'intervalle de classe est égal à l'unité, on a:
ls = ln+1 - ∑Yi
Dans le cas où la courbe est symétrique Ls est égal à Ls50, la taille pour laquelle 50 pour cent des individus sont retenus.
On admet en général que la relation entre ls, la longueur moyenne de sélection ou de rétention et la taille de la maille (m) est linéaire et de la forme
ls = b.m où b est le coefficient de sélection
Quelques informations disponibles sur ce sujet ont été portées sur la figure 74, où les longueurs des crevettes sont exprimées en longueur totale ou cephalothoracique suivant les travaux considérés. Une certaine variabilité apparaît dans la relation, due vraisemblablement aux différences de morphologie des espèces considérées, aux conditions dans lesquelles les expériences ont été réalisées (type de chalut et nature des fils utilisés) et à l'utilisation de ls ou l50 suivant les cas. Rappelons que la probabilité de capture pour une maille donnée et en particulier la valeur de ls dépend de la durée du chalutage (Berry et Hervey, 1965) à cause du colmatage progressif. Ceci implique que les courbes soit calculées avec une durée de trait égale à celle que pratiquent en moyenne les professionnels.
Des renseignements complémentaires sur les études de sélectivité et en particulier les aspects pratiques seront trouvés dans les manuels de Beverton et Holt, Ricker, Gulland ainsi que dans ICNAF (1963) et Pope (1975).
Selon Gulland (1972) les courbes de sélectivité de crevettes ne sont pas assez bien définies pour que la réglementation des maillages se justifie - Mistakidis (1958) indique également que la sélectivité n'est pas “parfaite” pour les caridés et que les chaluts retiennent toujours quelle que soit la maille, une partie des juvéniles. Cependant leur pourcentage dans les captures diminue quand la taille des mailles utilisées augmente. Kurk et al. (1965) confirment l'existence de courbes de sélectivité bien définies pour les caridés. Chez les penaeïdes, la réglementation des maillages semble être une méthode utilisable pour l'aménagement si l'on en croit les travaux de Roelofs (1950) et Berry et Hervey (1965) sur Penaeus aztecus, Regan et al. (1957) sur P. duorarum, Simpson et Perez (1975) et Lhomme (1978) sur Penaeus notialis Gorman (in Hynd, 1975) espèce non précisée, Aoyama (1973), sur P. orientalis et Lluch (1975) sur Penaeus vannamei et P. californiensis. Tous ces travaux ont porté sur des espèces de grande taille mais George et al. (1974) et Garcia et Lhomme (1977a) ont étudié la sélectivité des filets fixes sur pieux en estuaires vis-à-vis des juvéniles (Fig. 75). Les résultats indiquent que, même dans le cas de juvéniles les maillages sont susceptibles d'effectuer une sélection des tailles capturées. Notons que les filets fixes travaillent en pleine eau, avec peu de colmatage (sauf en saison des pluies où les débris végétaux sont nombreux). Il est vraisemblable qu'en revanche la sélectivité des chaluts utilisés dans les petites baies, près des nurseries, doit être très perturbée à cause de la forte charge du fond en détritus végétaux, coquilles vides, etc. D'autre part, Simpson et Perez (1975) ont montré que l'échappement à travers le corps du chalut pouvait être important.
Il est intéressant de comparer les coefficients de sélection obtenus par les différents auteurs.
| Auteur | Espèces | Coefficient de sélection | |
| Longueur éphalothoracique | Longueur totale | ||
| Aoyama (1973) | P. orientalis | 0,40 | |
| Lhomme (1979) | P. notialis | 0,37 – 0,52 | 2,0 à 2,61 |
| Regan et al. (1957) | P. duorarum | 0,35 – 0,37 | |
| Lluch (1975) | Penaeus spp. | 2 à 2,42 | |
| Kurk et al. (1965) | Crangon crangon | 2,15 | |
1 Transformation effectuée par les auteurs en utilisant la relation LC/LT de Garcia,1970
2 Calculs effectués par les auteurs à partir des figures de la publication originale
Ces valeurs correspondent toutes à des grands pénaeïdes du genre Penaeus et il n'est pas certain qu'elles s'appliquent aux autres espèces. Notons cependant que la valeur obtenue pour le petit caridé C. crangon est dans la gamme observée pour les pénaeïdes. Ceci peut laisser espérer que les valeurs ci-dessus soient généralisables à toutes les crevettes de morphologie classique mais des expérimentations supplémentaires seraient nécessaires pour pouvoir l'affirmer.
Si l'on considère les droites qui limitent les nuages de points sur la figure 74 et les droites moyennes (tracées à main levée) on obtient les valeurs du coefficient de sélection suivants:
1,90 ± 0,40 si les mesures sont en longueur totale
0,40 ≡ 0,10 si elles sont en longueur céphalothoracique.
L'intérêt d'une telle généralisation serait de permettre l'obtention rapide d'une valeur approximative de la taille moyenne de sélectivité pour des espèces pour lesquelles l'information de base manque.
La donnée traditionnellement importante pour les modèles d'évaluation est tc l'âge à la première capture (correspondant à une taille lc). Lorsque le recrutement et la sélection sont suffisamment distincts et que le recrutement est terminé lorsque la sélection commence, le premier phénomène peut être négligé pour la gestion du stock marin. Lorsque ce n'est pas le cas, il faut calculer la courbe résultante obtenue en multipliant pour chaque taille individuelle le pourcentage de recrutement par le pourcentage de rétention (Fig. 72). Le même procédé qui a permis de calculer ls, permettra de calculer lc (puis tc). Si la courbe résultante peut être considérée comme symétrique lc50 sera également une bonne approximation de lc.
L'utilisation du concept de taille (ou âge) à la première capture implique que le phénomène de recrutement (sélection) est assimilable à un phénomène en “lame de couteau”, c'est-à-dire que le pourcentage recruté est nul avant cette taille et égal à 100 au delà. Cette approximation permet de simplifier les calculs et d'utiliser les tables de rendement par recrue de Beverton et Holt (1964).
Dans le cas des penaeïdes, cette approche peut cependant ne pas être tout à faire satisfaisante si l'intervalle de taille (et d'âge) dans lequel se produit la sélection couvre une part trop importante de la phase exploitée.
Dans l'exemple de la Fig. 72 on voit que l'intervalle s'étend de 15 mm (LC) à 45 mm (LC) environ. Dans la mesure où peu de femelles, et pratiquement aucun mâle, n'atteignent cette taille, cela revient à admettre que l'intervalle de sélection couvre pratiquement toute la vie exploitable de l'animal.
Dans ces conditions, la simplification de la courbe de recrutement à une courbe en “lame de couteau” et l'hypothèse que 100 pour cent des crevettes sont recrutées à une taille lc risquent d'être la source d'un biais qui n'est peut-être pas négligeable.
Il appartiendra donc au biologiste de juger dans quelle mesure la courbe de recrutement peut être assimilée à une “lame de couteau”. Lorsque cette approximation ne paraît pas admissible, on peut lui préferer une approximation linéaire (Beverton et Holt, 1957) assimilant la partie ascendante de la courbe de sélection une droite. Une approximation “en escalier” peut également être utilisée par laquelle la probabilité de capture (exprimée en pourcentage) est supposée constante pour chaque âge (Fig. 76). Cette approche permet de donner une meilleure image du phénomène considéré mais interdit pratiquement l'usage des tables de rendement et rend nécessaire l'utilisation de méthodes de calcul plus complexes (modèle de Ricker, par exemple).
Les méthodes permettant d'effectuer les calculs sont nombreuses et le choix dépend des données disponibles. Les résultats les plus directement intéressants pour les pêcheurs sont, d'une part, les effets immédiats de la réglementation et, d'autre part, les effets prévisibles à long terme, sous le nouveau régime d'exploitation. Beverton et Holt (1957) ont montré avec élégance l'évolution de la population dans le temps après l'introduction d'une changement de maillage. Allen (1953), Holt (1958), Gulland (1961, 1969), Jones (1974) et Cadima (1977a) ont proposé des méthodes simples de calcul exigeant un minimum de données.
10.4.1 Calcul des effets immédiats
Le principe général en est le suivant. Connaissant la distribution annuelle de fréquence des tailles dans les captures réalisées avec le maillage traditionnel, sa courbe de sélection ainsi que celle du nouveau maillage que l'on se propose d'imposer, on peut calculer les conséquences immédiates du changement de maillage. En particulier si, pour une taille donnée, les pourcentages retenus par l'ancienet le nouveau maillage sont respectivement rl et rk et si C1 est la capture en nombre réalisée avec l'ancien maillage, on peut calculer la capture en nombre Ck avec le nouveau maillage, immédiatement après le changement, lorsque la structure de la population est encore celle qui prévalait sous l'ancien régime d'exploitation:
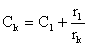
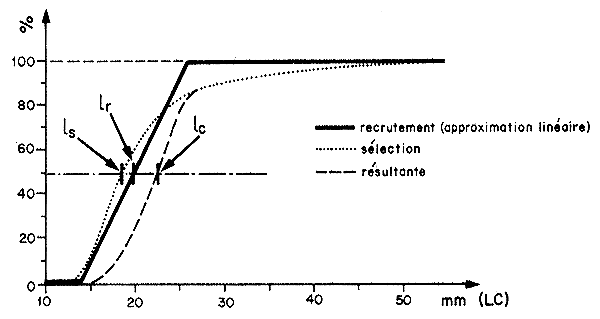
Fig. 72: Courbe de recrutement (approximative) et de sélectivité pour la maille de 25 mm de côté et courbe résultante pour Penaeus notialis au Sénégal, d'après Lhomme (1978)
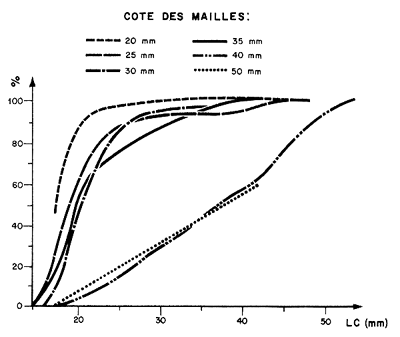
Fig. 73: Courbes de sélectivité observées pour des mailles de 25 à 50 mm de côté chez Penaeus notialis (d'après Lhomme, 1978)
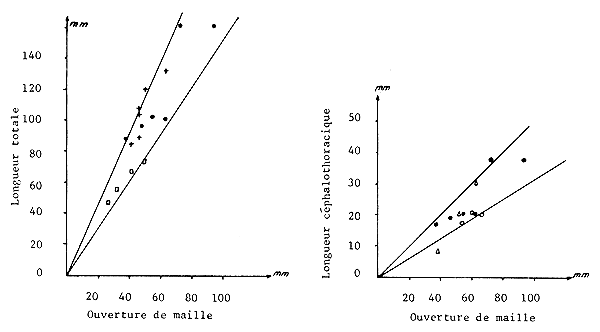
Fig. 74: Relation entre la taille moyenne de sélectivité et l'ouverture de maille
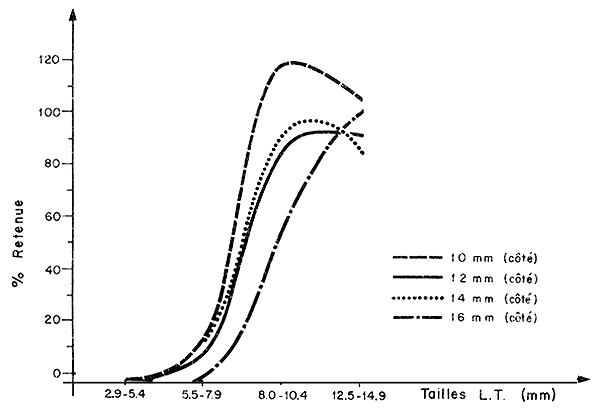
Fig. 75: Sélectivité des filets fixes utilisés en lagune Ebrié (Garcia et Lhomme, 1977)
Connaissant le poids moyen individuel correspondant à chaque taille on peut transformer ces captures en nombre en captures en poids et les captures totales réalisées sous l'ancien régime d'exploitation et juste après le changement de maillage seront
| Y1 = ∑ C1ω | et | Yk = Σ Ckω |
Le résultat immédiat en poids de la nouvelle réglementation en valeur relative par rapport à la capture dans la situation première, sera donc exprimé par
L = (Y1 - Yk)/Y1
Dans le cas des pénaeïdes, il est évident que l'utilisation d'une distribution annuelle des tailles pour calculer un effet “immédiat” n'a pas de sens. Chez les animaux à vie courte et à croissance rapide, la population constituée pratiquement d'une seule classe d'âge n'est jamais en équilibre et sa structure d'âge et de taille varie saisonnièrement. La Fig. 77 donne, à titre d'exemple, les variations de taille moyenne sur le plateau continental d'Arabie.
La distribution des tailles dans les captures totales annuelles est donc toute théorique et donne une image virtuelle d'une population “annuelle” qui n'existe, en fait, jamais. D'autre part, les effets définitifs (à “long terme”) de la réglementation seront obtenus dans l'année même de mise en application de la nouvelle maille et la notion suivant laquelle la population “immédiatement” après le changement de maillage reste identique à elle-même ne peut être admise que pour une période de un ou deux mois au maximum et impliquerait que les calculs soient effectués, au mieux, avec la distribution de fréquence des tailles dans les captures réalisées, par le passé, dans le ou les deux premiers mois qui suivent le changement de maillage, encore que la portée pratique de ce calcul soit limitée.
Le calcul des résultats réels, transitoires, du changement de maillage à partir de son application et jusqu'à la nouvelle situation d'équilibre peuvent parfaitement être effectués de façon plus précise en utilisant, par exemple, les techniques de simulation sur un modèle de Ricker (voir chapitre 11) à condition de disposer des données concernant la croissance, la mortalité naturelle et par pêche, le cycle annuel de recrutement avec leurs variations saisonnières éventuelles. On peut cependant se demander si ces résultats immédiats et transitoires ont une utilité pratique en matière de prédiction dans la mesure où les stocks de pénaeïdes sont hautement dynamiques et présentent parfois une grande variabilité interannuelle du recrutement (voir paragraphe 9.1.2).
Il est important, en revanche, de connaître le résultat définitif dans le nouveau régime d'exploitation, résultat qui sera de toute façon acquis très rapidement (environ six mois après le changement de maillage si celui-ci intervient juste avant le principal recrutement).
10.4.2 Calcul des effets à long terme
Le bénéfice à long terme de ce changement de maillage dépend du nombre d'individus “relâchés” par la nouvelle maille Ck (et qui étaient auparavant capturés par l'ancienne), de leur croissance, de leur mortalité naturelle et du taux d'exploitation. On peut calculer (Cadima, 1977) la capture en poids sous le nouveau régime d'exploitation (Y2) par rapport à la capture théorique juste après le changement de maillage (Yk) par1
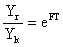 | d'après Holt (1958) | avec T = (tc2 - tc1), ou encore |
| d'après Gulland (1961), | 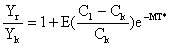 | |
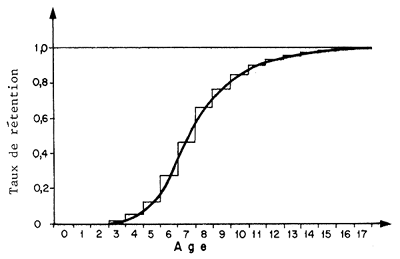
Fig. 76: Approximation de la courbe de sélection par une fonction en escalier
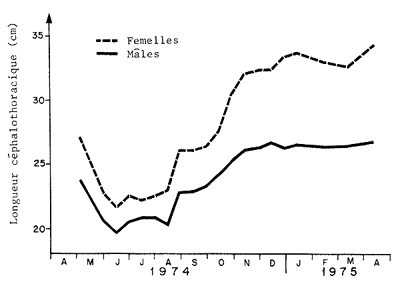
Fig. 77: Variations saisonnières de la taille moyenne chez Penaeus semisulcatus (mâles et femelles) en Arabie Saoudite, d'après Price & Jones (1975)
ou T* est égal à (tc2 - tr), tr étant l'âge moyen des poissons “relâchés” par la nouvelle maille, ou encore par
Yr/Yk = (C1/Ck)E = eE.loge(C1/Ck) Cadima (1977)
Etant donné que Y2/Y1 = (Y2/Yk) (Yk/Y1) et que Y1 et Yk sont connus (cf 10.4.1) le rapport de Y2 et Y1 permet d'apprécier l'effet à long terme découlant du changement de maillage.
L'application de ces méthodes suppose que le recrutement soit en lame de couteau, que les structures de taille des captures avant le changement de maillage soient connues, de même que M, E et T (méthode de Gulland), F et T (méthode de Holt) et E (méthode de Cadima). Cette dernière est donc la moins exigeante en données de base.
Si l'hypothèse d'une mortalité constante avec l'âge au delà de tc paraît peu plausible, en particulier si la courbe de sélection couvre une partie trop importante de la vie de l'animal pour admettre un recrutement en “lame de couteau” ou, en d'autres termes, si une partie importante des captures est effectuée dans l'intervalle de sélection, la formule de Holt surestimera le changement réel alors que les deux autres sous-estiment les résultats en poids mais pas en nombres (Gulland, 1961).
Lorsque les données sont disponibles (L∞, lc1, lc2, K, M, F) on peut aussi, bien entendu, utiliser les tables de Beverton et Holt (1964) pour calculer les indices de rendements par recrue correspondant aux deux régimes d'exploitation considérés et effectuer la comparaison. Si la courbe de sélection est approchée par une fonction en escalier, on pourra alors diviser la vie de l'animal en quelques périodes à l'intérieur desquelles Z est constant. Les calculs peuvent ensuite être effectués à l'aide des tables de rendement, le rendement par recrue total étant égal à la somme des rendements partiels. Le processus de calcul est indiqué dans les tables (Beverton et Holt, 1964, p. 7). Les calculs pourront également être simplement effectués avec les mêmes données en utilisant les formulations de Ricker (1958, 1975) pour le calcul du rendement par recrue ou encore plus simplement en adoptant la méthode approximative de Thompson et Bell (in Ricker, 1975). Ces deux dernières méthodes ont l'avantage de ne pas exiger la représentation de la croissance par le modèle de Von Bertalanffy et surtout de permettre l'expression des résultats du changement de maillage en termes monétaires en remplaçant simplement dans les calculs la clé âge-valeur. De la même manière, les variations de la fécondité théorique par recrue liées au changement de maillage peuvent être estimées.
Un exemple de calcul par la méthode de Ricker sera trouvé dans Ricker (1975, p. 245). Un exemple d'utilisation de la méthode de Thompson et Bell est donné en annexe I, à la fin du chapitre, avec un rappel des principales équations utilisées.
En réalité, les exemples de données sur les variations de mortalité par pêche avec l'âge sont très rares pour les pénaeïdes et nous avons vu au chapitre 7 qu'elles sont difficile à séparer des variations saisonnières.
L'analyse des cohortes effectuée sur le vecteur des captures par classes de tailles (méthode de Jones, 1974) qui est rappelée au paragraphe 7.6.2, permet d'obtenir un vecteur des mortalités par pêche en fonction de la taille. Dans ce travail, l'auteur indique que l'on peut calculer ensuite les effets d'une modification de l'effort et/ou du maillage. Jones (1979) a appliqué cette méthode à la langoustine (Nephrops sp.) et sa démarche est donnée à titre d'exemple en annexe II à la fin du chapitre, où les principales équations utilisées sont reproduites.
Notons, enfin, que les calculs peuvent être effectués de manière plus précise à l'aide des techniques de simulation. Ils sont basés sur les mêmes principes mais ils ne peuvent être facilement réalisés, que sur ordinateur. Ils permettent d'utiliser, en plus des données sur la croissance et les mortalités, des informations sur les niveaux de recrutement et les variations saisonnières des paramètres de base (capturabilité, effort, recrutement).
L'étude de la sélectivité des engins de pêche et la détermination des courbes de sélection spécifiques fournissent des résultats directement utilisables pour l'aménagement des pêcheries. Ce sont des résultats relativement faciles à obtenir, résultant de techniques dont la mise en oeuvre est simple. Ces données sont pourtant très rares en ce qui concerne les pénaeïdes et il serait nécessaire d'intensifier les études de sélectivité, en particulier pour les stocks de crevettes multispécifiques constitués d'espèces de taille moyenne très différentes.
Si l'effet à long terme d'un changement de maillage peut être facilement calculé de manière approximative, une analyse fine des événements pendant la période de transition nécessite le recours à des méthodes plus élaborées, dont le principe et simple, mais qui exigent beaucoup plus de données.
L'utilité d'une réglementation des maillages comme outil d'aménagement sera abordée en détail au chapitre 12.
Annexe I
Calcul de l'effet d'un changement de maillage en utilisant la méthode de Thompson et Bell (1934)
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 |
| Age | S1 | Ft1 | Zt1 | Popul. en nb Nt1 | Nb morts Dt1 | Capture (Nb) Ct1 | Poids individuel Wt1 | Capture en poids Yt2 | S2 | Ft2 | Zt2 | Nt2 | Dt2 | Ct2 | Yt2 |
| 1000 | 1000 | ||||||||||||||
| 2,5 | 0 | 0 | 0,3 | 259 | 0 | 5 | 0 | 0 | 0 | 0,3 | 259 | 0 | 0 | ||
| 741 | 741 | ||||||||||||||
| 3,5 | 0,5 | 0,15 | 0,45 | 269 | 90 | 10 | 900 | 0 | 0 | 0,3 | 192 | 0 | 0 | ||
| 472 | 549 | ||||||||||||||
| 4,5 | 1,0 | 0,3 | 0,60 | 213 | 106 | 20 | 2120 | 0 | 0 | 0,3 | 142 | 0 | 0 | ||
| 259 | 407 | ||||||||||||||
| 5,5 | 1,0 | 0,3 | 0,60 | 117 | 58 | 28 | 1624 | 0,5 | 0,15 | 0,45 | 148 | 49 | 1372 | ||
| 142 | 259 | ||||||||||||||
| 6,5 | 1,0 | 0,3 | 0,60 | 64 | 32 | 35 | 1120 | 1,0 | 0,3 | 0,60 | 117 | 58 | 2030 | ||
| 78 | 142 | ||||||||||||||
| 7,5 | 1,0 | 0,3 | 0,60 | 35 | 17 | 40 | 680 | 1,0 | 0,3 | 0,60 | 64 | 32 | 1280 | ||
| 43 | 78 | ||||||||||||||
| 8,5 | 1,0 | 0,3 | 0,60 | 20 | 10 | 45 | 450 | 1,0 | 0,3 | 0,60 | 35 | 17 | 765 | ||
| 23 | 43 | ||||||||||||||
| 9,5 | 1,0 | 0,3 | 0,60 | 10 | 5 | 48 | 240 | 1,0 | 0,3 | 0,60 | 20 | 10 | 480 | ||
| 13 | 23 | ||||||||||||||
| 10,5 | 1,0 | 0,3 | 0,60 | 6 | 3 | 50 | 150 | 1,0 | 0,3 | 0,60 | 10 | 5 | 250 | ||
| 7 | 13 | ||||||||||||||
| 11,5 | 1,0 | 0,3 | 0,60 | 3 | 1,5 | 52 | 78 | 1,0 | 0,3 | 0,60 | 6 | 3 | 156 | ||
| 4 | 7 | ||||||||||||||
| 12,5 | 1,0 | 0,3 | 0,60 | 2 | 1 | 53 | 53 | 1,0 | 0,3 | 0,60 | 3 | 2 | 106 | ||
| Total | 7415 | 6439 | |||||||||||||
Colonnes 2 et 10 = Courbes de sélectivité pour l'ancien maillage (S1) et le nouveau (S2)
| 3 et 11: Ft = F base × S1 ou F base × S2, F base = 0,30/mois | 6 et 14: | Dt = Nt+1 - Nt |
| 4 et 12: Zt = Ft + Mt, = Cte = 0,3/mois | 7 et 15: | Ct · Ft/Zt |
| 5 et 13: Nt = Nt-1e-Zt, 1000 recrues à 2 ans | 9 et 16: | Yt = Ct · Wt |
| Résultat du changement de maillage = (6439 – 7415)/7415 = - 13,9% |
Annexe II
Calcul de l'effet d'une augmentation de l'effort (de 40 pour cent) et du maillage (de 20 mm)
sur le rendement par recrue de la langoustine (d'après Jones, 1979)
| 1 Length (mm) | 2 N × W (kg) | 3 F1DT | 4 S1 | 5 S2 | 6 F2DT | 7 F1DT-F2DT | 8 ∑(7) | 9 exp (8) | 10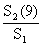 | 11 (2)×(10) |
| 10–15 | 1 | 0 | 0.0019 | 0 | 0 | 0 | 0 | 1.0000 | 0 | 0 |
| 15–20 | 480 | 0.0085 | 0.1170 | 0.0039 | 0 | 0.0085 | 0.0041 | 1.0041 | 0.0035 | 16 |
| 20–25 | 8 714 | 0.0792 | 0.6950 | 0.1038 | 0.0166 | 0.0626 | 0.0395 | 1.0403 | 0.1554 | 1 354 |
| 25–30 | 47 126 | 0.2943 | 0.9864 | 0.0557 | 0.2321 | 0.0622 | 0.1019 | 1.1073 | 0.6238 | 29 400 |
| 30–35 | 89 290 | 0.5323 | 1.0 | 0.9382 | 0.6992 | - 0.1669 | 0.0495 | 1.0508 | 0.9858 | 88.022 |
| 35–40 | 88 170 | 0.6520 | 1.0 | 0.9984 | 0.9113 | - 0.2593 | - 0.1636 | 0.8491 | 0.8477 | 74 745 |
| 40–45 | 69 660 | 0.7647 | 1.0 | 1.0 | 1.0706 | - 0.3059 | - 0.4462 | 0.6401 | 0.6401 | 44 589 |
| 45–50 | 48 750 | 0.9828 | 1.0 | 1.0 | 1.3759 | - 0.3931 | - 0.7957 | 0.4513 | 0.4513 | 22 011 |
| 50–55 | 24 830 | 1.2130 | 1.0 | 1.0 | 1.6982 | - 0.4852 | - 1.235 | 0.2908 | 0.2908 | 7 221 |
| 55–60 | 9 829 | 1.7580 | 1.0 | 1.0 | 2.4612 | - 0.7032 | - 1.829 | 0.1606 | 0.1606 | 1 579 |
| 60–65 | 1 343 | 1.0780 | 1.0 | 1.0 | 1.5092 | - 0.4312 | - 2.396 | 0.0911 | 0.0911 | 122 |
| 65–70 | 508 | 1.0780 | 1.0 | 1.0 | 1.5092 | - 0.4312 | - 2.828 | 0.0591 | 0.0591 | 30 |
| Total | 388 701 | 269 109 |
1. Données de base
| Colonnes 2: | N = Nombres capturés, par classes de taille |
| W = poids moyen individuel | |
| Capture en poids en régime d'exploitation 1 = 388 701 | |
| 3: | Vecteur de F par intervalle de taille DT en régime d'exploitation 1 |
| (mailles de 70 mm) obtenu par analyse des cohortes selon la méthode | |
| de Jones (1974) | |
| 4 et 5: | proportions retenues pour chaque taille par les maillages de 70 mm |
| (S1) et 90 mm (S2). |
2. Calculs
| 6. | Vecteur de F en régime d'exploitation 2 (maille de 90 mm et effort × 1,4) |
F2DT = (1 + x/100)S2 (F1DT)/S1 où × est l'augmentation d'effort prévue, en %. Par exemple pour la classe de taille 25–30 mm | |
F2DT = 1,4 . 0,5557 . 0,2943/0,9854 = 0,2321 | |
| 7. | Différences entre F1DT et F2DT |
| 8. | Cumul des différences = ∑ (F1DT - F2DT) jusqu'au point médian de l'intervalle considéré = soit 0,1019 = 0,85 + 0,0626 + (0,0622/2) |
| 9. | e∑(F1DT - F2DT) |
| 10. | 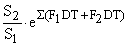 |
| 11. | 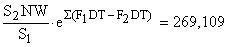 |
3. Résultat final
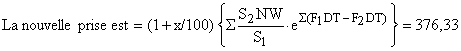
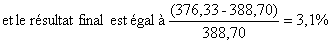
![]()
![]()
![]()