![]()
![]()
![]()
Les chapitres précédents avaient pour objet de passer en revue l'information nécessaire à l'évaluation des ressources et à la définition des conditions optimales d'exploitation (données biologiques, statistiques de pêche, paramètres dynamiques, etc.). Ces données, dont dépend largement la qualité finale des évaluations et des diagnostics sont utilisées dans des modèles qui ne sont pas propres aux penaeïdes, mais au contraire, généralement utilisés pour les poissons, tropicaux ou non.
On distinguera les modèles globaux (ou modèles de production), les modèles analytiques (rendement par recrue, technique de simulation) et les méthodes approchées (à partir des biomasses, par comparaison, etc.). Plutôt que de s'attarder sur la formulation mathématique et les justifications biologiques des modèles, amplement traitées dans les ouvrages de base, on examinera les hypothèses implicites de ces modèles et la justification de leur utilisation dans le cas particulier des penaeïdes. Une discussion intéressante dans le cas plus général des invertébrés sera trouvée dans Hancock, 1979.
Le modèle de Schaefer (1954) et ses dérivés (Fox, 1970; Pela et Tomlinson, 1969) sont comparables à ceux utilisés pour l'étude des relations prédateur-proie. Ils considèrent le stock comme une entité dont l'abondance dépend de l'action de prédation exercée par l'Homme. Si la biomasse est exprimée par la prise par unité d'effort et la prédation par l'Homme par l'effort de pêche, on observe en général une relation négative, linéaire ou non entre ces deux grandeurs. La relation entre la capture totale (égale au surplus de production) et l'effort est une courbe parabolique ou non qui passe par un maximum pour un niveau d'abondance intermédiaire.
L'analyse de ces relations permet de calculer la prise maximale équilibrée (MSY des anglophones) et le niveau d'effort (FMSY) qui lui correspond. Ces modèles sont également la base d'une gestion des pêcheries par quotas de capture ou limitation de l'effort. Leur utilisation courante implique quelques hypothèses toujours acceptées bien que l'on sache parfois qu'elle ne sont pas réalistes. Il est intéressant de les reprendre ici pour voir dans quelle mesure elles sont respectées dans le cas des penaeïdes.
11.1.1 Les hypothèses propres aux modèles
On se référera ici aux travaux de Pella et Tomlinson (1969) et Fox (1974).
(a) Déterminisme des modèles: Les mécanismes de production sont en général considérés comme des processus déterministes. Toutes les variations de production annuelle autres que celles qui sont dues aux variations de l'effort de pêche sont négligées1. Dans le cas des penaeïdes, on a noté des variations de production d'ordre vraisemblablement climatiques. Cependant, la variabilité interannuelle semble avoir une amplitude très variable suivant les stocks considérés. Pour ceux où cette variation est telle qu'elle masque complètement la relation entre cpue et effort, le modèle n'est à l'évidence pas approprié car il est trop imprécis (Geibel et Heimann, 1976). Pour d'autres, les variations interannuelles paraissent faibles mais il existe des tendances à long terme de nature périodique, et liées aux oscillations climatiques. Dans ce cas, le danger réside surtout dans l'utilisation d'une série de données trop courte, conduisant à des conclusions erronées en matière d'évaluation. En ce qui concerne l'utilisation eventuelle du modèle pour une politique de gestion basée sur des quotas de capture, le modèle est inutilisable dans les deux cas car la performance du stock une année ne permet pas de prévoir celle de l'année suivante en l'absence de données supplémentaires.
(b) Conditions d'équilibre: On admet qu'à chaque niveau d'effort correspond un état stable de la structure démographique de la population. Ceci suppose que le principal facteur contrôlant la taille du stock à chaque instant est l'effort de pêche. Notons que lorsque de nombreux petits stocks voisins existent et que les flottilles peuvent se déplacer d'un stock à l'autre, il arrive parfois que ce soit l'abondance du stock qui règle le niveau de l'effort et non l'inverse. La relation cpue/effort ne donne pas dans ce cas l'image recherchée de l'évolution du stock sous l'action de la mortalité par pêche. Un exemple de ce type de phénomène peut être trouvé dans le golfe d'Exmouth en Australie (FAO/CPOI, 1973).
(c) Population unique: On suppose souvent qu'il n'existe qu'un seul stock, ou bien que l'ensemble des stocks étudiés régit comme un tout; on ignore, donc volontairement les intéractions entre stocks. Fox (1974) a montré l'importance du taux de melange entre stocks sur la validité des évaluations effectuées globalement pour plusieurs unités. Le problème peut être particulièrement ardu lorsque la ressource est constituée d'un grand nombres, de petits stocks aux interrelations peu ou pas du tout connues comme à Madagascar (voir Marcille, 1978) et pour les stocks où les données ne permettent pas de distinguer entre des espèces voisines (voir Fig. 78).
(d) Caractéristiques stables du stock: On suppose que la production dépend de la biomasse du stock considéré comme un tout, ou encore que la production et les prises sont indépendantes de la structure démographique. Dans la réalité, il est évident que la mise en exploitation d'un stock en provoque le rajeunissement et en modifie les caractéristiques de production. Le phénomène peut conduire à englober dans le même modèle des données qui, en fait, suivent des modèles différents (Daget et Le Guen, 1974; Fox, 1974).
D'autre part, il faut noter que dans le cas des penaeïdes, le “stock” exploité, sa “biomasse” théorique, sa structure d'âge, telle qu'elle est reflétée dans les captures annuelles par exemple, n'existent jamais en tant que tels. Au n'existent niveau annuel, on dispose tout au plus d'une image virtuelle de la population, car un stock de penaeïdes avec une seule classe d'âge est quelque chose d'extrêmement dynamique dont la biomasse, l'âge moyen, la fécondité, etc., varient considérablement au cours de l'année, ce qui nous éloigne beaucoup des conditions retenues par Schaefer pour son modèle.
Une difficulté supplémentaire dans le cas des penaeïdes est que la pêche en mer porte sur un stock dont la production annuelle dépend beaucoup du prélèvement effectué sur les juvéniles en estuaire par une exploitation plus ou moins artisanale. Une modification de ce taux d'exploitation faussera la relation cpue/effort en mer par une modification du recrutement indépendante du stock parental et de l'effort en mer.
11.1.2 Hypothèses concernant les données
(a) Capturabilité constante: C'est l'un des points les plus délicats de ce type de modèle. La capture est toujours à peu près bien définie (bien que la modification progressive des schémas de rejets puisse être une source de biais) et l'effort est supposé obéir à la relation F = qf tout au long de la période étudiée. Si l'on se reporte au paragraphe 4.1.1.2, on verra que les sources de biais identifiées chez les crevettes sont nombreuses (changement du nombre d'heures de pêche effectives par jour, de la répartition journalière ou saisonnière de l'effort de pêche, amélioration de l'efficacité des engins, etc.).
Une autre difficulté importante réside dans la définition de l'effort appliqué à chaque espèce dans une pêcherie multispécifique. Lorsque les données précises n'existent pas, l'effort global est pris en considération or sa répartition réelle par espèce est très susceptible de changements progressifs au cours du temps en liaison avec les variations relatives d'abondance des espèces considérées. Un exemple de tentative de solution de ce problème est donné par Hall et Penn (1979) dans le cas de la pêcherie crevettière de Shark Bay (Australie).
Hancock (1979) rappelle enfin que pour les crevettes à comportement grégaire très marqué comme Penaeus merguiensis la notion de temps de recherche doit être prise en considération (comme c'est le cas pour les espèces pélagiques) pour le calcul de l'effort de pêche, car la raréfaction du stock se traduit par un changement du rapport entre le temps de recherche et le temps de chalutage.
(b) Pas de temps de latence: On suppose que le stock réagit à toute variation de l'effort pour atteindre immédiatement son état d'équilibre. Ce phénomène est d'autant plus susceptible de créer des distorsions que les espèces ont une longévité importante, que l'âge à la première capture est élevé et que les variations de l'effort de pêche sont grandes.
Ce problème volontairement négligé dans l'approche de Schaefer (1954, 1957) et de Pela et Tomlinson (1969) est en partie résolu par les approches de Gulland (1961, 1969), Le Guen et Wise (1967), Fox (1974) et Walter (1975). Les conditions d'équilibre sont alors approchées en reliant la prise de l'année à l'effort de l'année en cours et des années précédentes, combinés de manière diverse suivant les auteurs.
Les stocks de penaeïdes étant en général composés d'une seule classe d'âge annuelle, les temps de latence peuvent être considérés comme nuls. Lorsque les variations du recrutement sont importantes, on peut raffiner les données en considérant non plus des années calendaires mais des années “biologiques” commençant avec le recrutement et correspondant ainsi pratiquement à la prise réalisée sur une génération.
11.1.3 Conclusions
Au cours de l'énoncé des diverses hypothèses implicites des modèles de production, des objections ont été soulevées tendant à limiter leur utilisation dans le cas des penaeïdes. Pourtant, à cause du manque fréquent de données détaillées plus appropriées, les modèles de production ont été très utilisés (Boerema, 1969; Joseph, 1971; Cadima et al., 1972; Marcille, 1972; Marcille et Stequert, 1974; Garcia et Lhomme, 1977, 1979; Marcille, 1978). L'utilisation de ce modèle par Griffin et al. (1973) pour les pêcheries du golfe du Mexique a été critiquée par Eldridge, 1974. Klima et Parrak (1978) l'ont cependant réutilisé pour la même région.
Un exemple d'utilisation de ce modèle pour les pêcheries du Salvador est donné sur la figure 78.
La qualité de l'évaluation du potentiel obtenue sera en grande partie liée à la qualité des données de prise annuelle et celle du niveau d'exploitation correspondant dépendra de la stabilité réelle de la relation entre F et f au cours du développement de la pêcherie.
L'évaluation obtenue par ce modèle sera d'autant plus valable que l'analyse portera sur une longue série de valeurs. La production crevettière étant susceptible de variations liées au climat, une évaluation basée sur une période courte risque d'entraîner une confusion entre les variations liées au climat et celles qui sont dues à l'effort, conduisant à une évaluation erronée (un exemple est donné par Hancock, 1975).
L'inconvénient de ce type de modèle est l'impossibilité d'intégrer les données provenant de la pêche en estuaire sur les juvéniles et celles de la pêche en mer sur les adultes. Le plus souvent, faute de données, seule la pêcherie maritime est prise en considération (admettant ainsi implicitement que le niveau de la pêche artisanale n'a pas changé pendant toute la période d'observation). Un essai d'intégration a été tenté en additionnant les deux captures annuelles artisanale et industrielle et calculant l'effort total correspondant en divisant la prise totale ainsi obtenue par la cpue des chalutiers en mer (FAO/CPOI, 1973). Cette méthode n'est en réalité utilisable que lorsque les structures démographiques exploitées sont identiques, ce qui n'est pas le cas ici. L'effort ainsi calculé sous-estime en réalité la mortalité par pêche qu'il est sensé représenter.
Le dernier défaut, et non le moindre, des modèles de production est qu'ils ne permettent pas d'envisager les conséquences des modifications du vecteur de mortalité par pêche en fonction de l'âge, par réglementation du maillage, fermetures saisonnières, etc. Leur utilisation comme outil d'aménagement est envisagée au chapitre suivant où leurs avantages et inconvénients seront discutés.
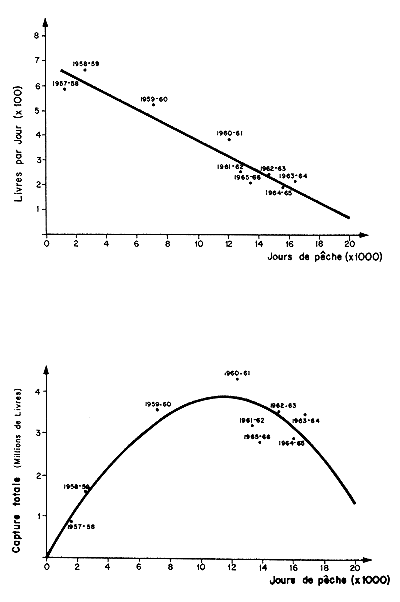
Fig. 78: Application du modèle de Schaeffer aux crevettes penaeides du Salvador (d'après Ellis, 1968). Il s'agit d'un mélange de P. occidentalis, P. vannamei et P. styliorostris.
Ainsi que cela a déjà été souligné de nombreuses fois les crevettes penaeïdes sont généralement exploitées à deux phases différentes de leur cycle vital (juvénile et adulte) par deux systèmes d'exploitation souvent très différents du point de vue socio-économique. La production de chacune des phases exploitées dépend, a priori, du niveau d'exploitation dans l'autre. Il est souhaitable de chercher à optimiser l'ensemble et les modèles analytiques permettant de résoudre le problème (notons que si l'exploitation des juvéniles a une action directe sur la production du stock d'adulte, la réciproque n'a encore jamais été démontrée).
11.2.1 Les modèles de rendement par recrue
Le cycle vital d'une cohorte est divisé en deux parties. La première, qui va de la naissance au recrutement, est en général mal connue et difficile à mettre en équation car l'estimation des paramètres est malaisée. La deuxième, qui va du recrutement à la fin de la vie dans la phase exploitable peut-être modelisée si l'on connaît les paramètres de croissance et de mortalité. Le nombre de recrues étant le plus souvent inconnu, les résultats seront exprimés en valeur relative, en rendement par recrue.
Les deux modèles plus utilisés sont ceux de Beverton et Holt (1957) et Ricker (1958). Le premier est mathématiquement plus élégant mais le deuxième, plus souple, permet de reproduire plus aisément la réalité, surtout depuis que l'utilisation des ordinateurs se vulgarise (Paulik et Bayliff, 1967). Le modèle de Thompson et Bell (1934 et in Ricker, 1975), moins utilisé et dont le principe est assez proche de celui de Ricker puisque basé sur une discrétisation de la vie en périodes courtes où les paramètres sont constants, a été utilisé par Berry pour Penaeus duorarum en Floride (in Lindner, 1966).
Les hypothèses de base de ces modèles sont les suivantes:
(a) Paramètres biologiques constants: Dans les versions les plus simples, les taux de croissance et de mortalité sont supposés constants d'une année à l'autre et indépendants du niveau d'exploitation et donc de la densité du stock (“density independent factors”). Le modèle de Beverton et Holt suppose en plus que M et F ne varient pas avec l'âge. En réalité, des variations peuvent être prises en considération en divisant également la vie d'une cohorte en segments dans lesquels les paramètres sont constants (cf. Beverton et Holt, 1966). Ces derniers peuvent eux-mêmes être des fonctions de la densité ou de l'environnement mais au prix d'une complexité extrême dans les calculs.
La notion de production par recrue permet d'éviter le problème de la relation stock-recrutement le plus souvent ignorée. Malheureusement, l'utilisation de cette notion conduit parfois à considérer que la prise totale évolue comme le rendement par recrue, ce qui n'est le cas que si le recrutement annuel est constant et indépendant de la taille du stock.
(b) Déterminisme du modèle: Bien que tous les paramètres des modèles puissent théoriquement être considérés comme des variables aléatoires, cette possibilité est rarement utilisée et les paramètres sont en général considérés comme fixes et identiques pour tous les individus d'une cohorte.
(c) Conditions d'équilibre: L'utilisation la plus courante des modèles de rendement par recrue suppose que le stock est en équilibre et que toutes les conditions sont stables. Dans ce cas, la production annuelle de toutes les classes d'âge présentes dans la pêcherie est égale à la production d'une seule classe d'âge pendant toute la phase exploitée. La variable temporelle utilisée dans les équations de capture est en général l'année, pour les poissons à vie longue. Dans le cas des penaeïdes à vie courte, cette variable doit être inférieure à l'année et le mois ou la semaine sont le plus souvent utilisées (pour les études de croissance, mortalité, recrutement, etc.) ce qui entraîne la nécessité de prendre en considération les variations saisonnières des paramètres de la population. Le recrutement est une fonction continue du temps mais on peut, en première approximation, admettre le recrutement de 12 cohortes mensuelles d'importance inégale. La production de chaque cohorte dépendra de son schéma saisonnier de croissance (lié à la date de naissance) et d'exploitation (lié aux variations saisonnières de q et de f). La production globale des 12 cohortes, ou production de la génération annuelle, dépendra donc du niveau global de recrutement et des variations saisonnières de l'effort de pêche, tous les autres facteurs et leurs variations saisonnières étant par ailleurs supposés fixés.
L'adoption de l'hypothèse d'équilibre pour les penaeïdes implique donc une stabilité de toutes les variations saisonnières, et en particulier celles de l'exploitation.
En conséquence, l'utilisation d'un modèle simple de rendement par recrue suppose que l'on peut correctement approcher la production complexe de 12 cohorte mensuelles pendant un an par la production d'une génération idéale théorique ayant des paramètres dynamiques “moyens”. La qualité des résultats dépendra beaucoup de la représentativité de ces derniers.
(d) Conclusions: Si l'on dispose des données nécessaires (mortalité naturelle et par pêche, constantes ou fonctions de l'âge, croissance, taille au recrutement et sélectivité) on peut calculer l'évolution du rendement par recrue, de la fécondité théorique par recrue (Le Guen, 1971) en fonction de la mortalité par pêche et de l'âge à la première capture. On obtient ainsi les moyens de prévoir les conséquences d'un aménagement de la pêcherie par régulation de l'effort, réglementation des maillages, établissement de cantonnements, etc.
Dans le cas des pénaeïdes cependant, ces modèles ne permettent pas de prendre facilement en considération les variations saisonnières (de croissance, de recrutement, de capturabilité, etc.) et donc de prévoir les conséquences précises d'une modification de la répartition saisonnière de l'effort (par fermeture de la pêche par exemple) sur le vecteur “moyen” des mortalités par pêche en fonction de l'âge et sur la production globale. Les techniques de simulation que l'on abordera au paragraphe suivant permettent de résoudre ce problème.
Ce type de modèle a été utilisé dans le golfe du Mexique par Kutkuhn1(1966) et Berry (1967) pour les crevettes des Tortugas (Floride). Les résultats de ce dernier sont reproduits sur les figures 79 et 80. Une étude préliminaire avait été effectuée sur un modèle de Thompson et Bell (Lindner, 1966).
Les modèles de Ricker et de Thompson et Bell permettent de calculer également la fécondité théorique par recrue (notion introduite par Le Guen, 1971) et en particulier son évolution en fonction de F et de lc en intégrant dans le modèle des données sur la fécondité en fonction de l'âge (Fig. 81). L'utilité de cette donnée théorique a été discutée au paragraphe 10.1.3. Exprimée en valeur relative par rapport à la fécondité théorique du stock vierge, elle est un indice de l'état de dégradation du potentiel reproducteur sous l'action de la pêche.
Pour permettre un aménagement mieux adapté, les conséquences d'une mesure d'aménagement doivent pouvoir être exprimées non seulement en termes de production pondérale (absolue ou par recrue) mais également en termes économiques valeur des captures, coût des captures, produit marginal, etc. (cf. chapitre suivant).
Les modèles de rendement par recrue peuvent être utilisés dans ce sens (Fig. 79b). Kutkuhn (1966) par exemple, a remplacé dans le modèle de Beverton et Holt l'équation de la courbe de croissance en poids par celle de la croissance en valeur; cet artifice lui a permis d'exprimer ses résultats en valeur par recrue.
Les modèles plus souples de Thompson et Bell et de Ricker se prêtent particulièrement bien à cette utilisation puisqu'il n'est pas nécessaire d'adopter un modèle mathématique quelconque et en particulier celui de Von Bertalanffy pour représenter l'évolution de la valeur en fonction de l'âge, mais que l'on utilise simplement les valeurs observées (Fig. 81).
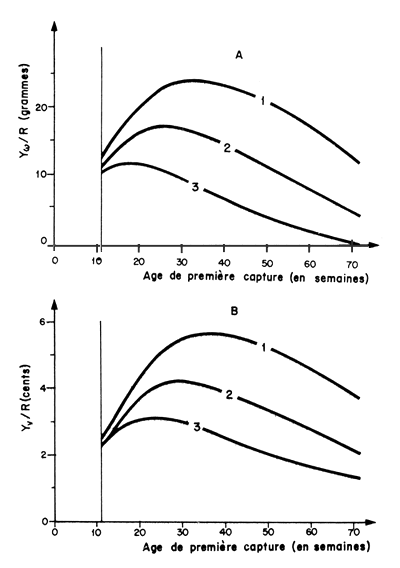
Fig. 79: Evolution du rendement par recrue en poids (A) et en valeur (B)
en fonction de l'âge de première capture, pour diverses combinaisons
observées, de F et de M (par semaine), chez Penaeus
duorarum en Floride (Berry, 1967)
1: F = 0,193 et M = 0,024
2: F = 0,227 et M = 0,039
3: F = 0,160 et M = 0,061
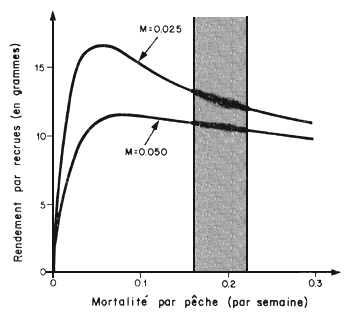
Fig. 80: Evolution du rendement par recrue pondéral en fonction de F pour 2 valeurs de M (par semaine) (d'après Berry, 1967)
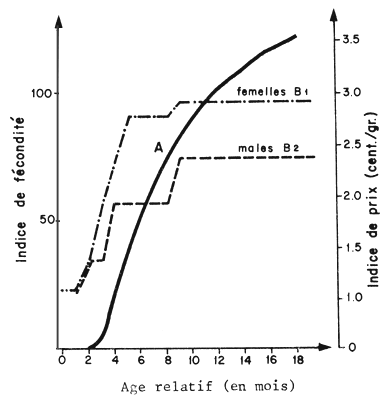
Fig. 81: Evolution de l'indice de fécondité (A) et de prix (B)en fonction de l'âge relatif Penaeus notialis (Garcia, 1977)
11.2.2 Les techniques de simulation
De plus en plus répandues depuis la vulgarisation des ordinateurs, elles permettent de s'afranchir de certaines contraintes imposées par les modèles précédents. Comme les modèles de rendement par recrue, les techniques de simulation utilisent le plus souvent une approche analytique.
Elles permettent, lorsque les données existent, de se rapprocher un peu plus de la réalité que ne le font les simples modèles de rendement par recrue. Une population de crevettes peut être reconstituée par exemple par 12 cohortes mensuelles d'importance inégale reproduisant ainsi le schéma saisonnier de migration et recrutement. Chaque cohorte peut suivre son propre schéma de croissance saisonnière et de mortalité en fonction de l'âge. La croissance des sexes étant différente (ce qui implique des âges à la première capture inégaux et une évolution différente de la biomasse), on les considérera séparément. Malgré la complication des calculs, les formules de base restent identiques et les résultats sont de même nature que précédemment. Si les variations saisonnières du recrutement sont connues seulement en valeur relative (en pourcentage du recrutement total par mois) on calculera la production pour un nombre donné de recrues (1 000 par exemple). Les résultats seront, en fait, de même nature qu'au paragraphe précédent et sont sujets aux mêmes restrictions liées à la forme réelle, et inconnue, de la relation stock-recrutement.
Si l'on dispose d'un moyen d'apprécier l'effectif absolu du recrutement annuel, la simulation peut alors être plus complète et les données calculées (prise, captures par unité d'effort, structures démographiques, variations saisonnières d'abondance, etc.) peuvent être comparées aux données observées dans le passé. Si la reproduction des événements passés est satisfaisante, on considère que le modèle peut être utilisé comme un outil de prévision pour évaluer les conséquences des divers schémas d'aménagement envisageables.
L'introduction dans la simulation d'une relation stock-recrutement permet de constituer un modèle autorégénérant. Dans le cas des crevettes penaeïdes cependant, cette relation n'est pas connue et son utilité est discutable (cf. paragraphe 10.1.3).
Les paramètres utilisés pour la simulation ne sont que des estimations; il est recommandé d'effectuer une étude de sensibilité de la simulation aux variations des divers paramètres. Cette procédure permet, de mettre en évidence les paramètres les plus “sensibles” conditionnant la qualité des résultats et sur lesquels l'effort de recherche ultérieur devra plus spécialement porter.
Rappelons que la simulation effectuée avec des paramètres obtenus par analyse des cohortes conduit forcément à une bonne superposition des résultats simulés et observés, mais que cette concordance est triviale et n'est pas une preuve de la bonne représentativité du modèle.
Les techniques de simulation ont été appliquées, par exemple, par Abramson et Tomlinson (1972) chez une crevette caridae (Pandalus sp.). Elles commencent à être utilisées chez les penaeïdes (Lluch, 1975; Garcia, 1977, 1978; Garcia et Ndiaye, 1977; Blomo et al., 1979; Grant et Griffin, 1979; Clark et Kirkwood, 1979).
Dans le cas des penaeïdes, elles présentent l'avantage de permettre l'intégration dans un seul modèle de l'exploitation des juvéniles et de celle des adultes pour évaluer les résultats globaux d'un schéma d'aménagement intégré (Fig. 82). Des exemples seront trouvés dans Garcia (1978), et dans Grant et Griffin (1979). L'organigramme du modèle bioéconomique mis au point par cet auteur est donné à titre d'exemple (Fig. 83).
Elles permettent également de modéliser et de chercher la stratégie optimale d'une exploitation simultanée de plusieurs stocks indépendants. C'est ainsi que Clark et Kirkwood (1979) en Australie, ont étudié par simulation le développement d'une pêcherie à entrée libre comprenant deux types de bateaux différents et exploitant deux stocks composés d'espèces d'intérêt économique inégal pour définir l'allocation spatiotemporelle optimale et la meilleure composition pour la flottille.
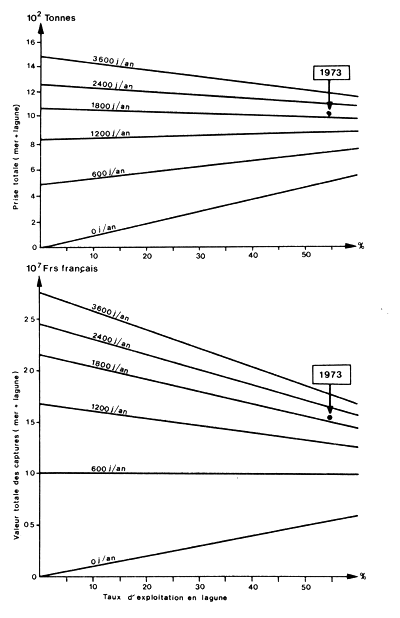
Fig. 82: Evolution de la prise totale (mer + lagune), en poids et en valeur, en fonction du taux d'exploitation en lagune et de l'effort annuel en mer (lignes obliques). La situation de la pêcherie en 1973 est indiquée (d'après Garcia, 1978)
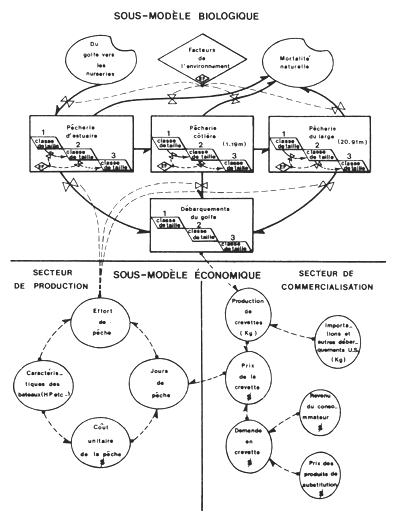
Fig. 83: Organigramme du modèle bioéconomique élaboré par Grant et Griffin (1979) pour la pêcherie crevettière américaine du golfe du Mexique (symboles d'après Forrester, 1961)
Dans le cas des animaux à vie courte, les variations saisonnières de la structure de la population, de sa biomasse, de ses effectifs, de son potentiel de reproduction, sont importantes. La simulation de ces variations est d'un intérêt certain pour la compréhension des mécanismes biologiques. Garcia (1977) a ainsi simulé l'évolution saisonnière théorique du nombre de femelles “adultes” (de taille supérieure à la taille de première maturité) dans la population et montré l'importance de ce phénomène pour l'interprétation des variations saisonnières de la reproduction.
En réalité, la seule limite à l'utilisation des modèles de simulation est la disponibilité et la qualité des données et l'on devra toujours veiller à ne pas trop s'éloigner de la réalité en demandant à un modèle plus que ce que les données sur lesquelles il repose ne l'autorisent à dire.
Le développement de l'exploitation d'une ressource est en général beaucoup plus rapide que celui du potentiel de recherche destiné à fournir aux administrateurs les éléments scientifiques d'une gestion raisonnée. Il existe donc toujours le risque qu'une information scientifique précise mais trop tardive, ne permette en définitive qu'un diagnostic “post mortem”.
Il sera d'autre part souligné, au chapitre suivant, qu'il est préférable d'aménager une ressource dès le début de son développement et sans attendre que l'exploitation anarchique conduise à un état de crise difficile à résoudre.
Comme l'indique Troadec (1977) “lorsqu'il s'agit de suivre l'évolution d'une pêcherie, exactitude, précision et opportunité sont trois qualités interdépendantes dont l'importance varie au cours du développement d'une pêcherie”. Dans la mesure où dans la recherche excessive d'une précision accrue, l'accroissement de l'information nécessaire “et donc son coût et ses délais d'acquisition croissent plus vite que la précision qu'on en tire, on comprend que la précision puisse être ennemie de l'exactitude et surtout de l'opportunité”.
Des méthodes approchées peuvent être utilisées pour l'évaluation des paramètres de base, et cet aspect a été examiné dans le chapitre 7, ou pour une première appréciation des potentiels et des niveaux d'exploitation. Un résumé des possibilités sera trouvé dans Saetersdal (1973) ou Troadec (1977) et nous ne reprendrons ici que les aspects applicables aux penaeïdes avec quelques chances de succès.
11.3.1 Relation entre la biomasse obtenue par prospection et le potentiel
Cette méthode implique la connaissance de la biomasse d'un stock que l'on peut obtenir à l'aide des prospections par chalutage.
11.3.1.1 Commentaires sur les prospections
Elles peuvent fournir de bonnes évaluations préliminaires. Elles permettent d'identifier les espèces intéressantes, leur répartition et leur abondance relative. Correctement planifiées, elles permettent une estimation de la biomasse du stock si l'efficacité réelle du chalut utilisé est connue, ce qui, reconnaissons-le, est assez rare. Si l'on doit supposer, faute de données précises, que l'efficacité est totale et que tout le poisson présent devant le chalut est capturé la biomasse calculée doit être considérée comme une estimation minimale car, ainsi que cela a été souligné au paragraphe 7.7.3 cette hypothèse n'est vraisemblablement pas réaliste et l'efficacité pourrait bien être plus près de 0,5 que de 1.
Les documents de base sur la théorie des prospections sont nombreux: Alverson et Pereyra (1969), Alverson (1971), Mackett (1975), Holden et Raitt (1974), Gulland (1975a) et Ulltang (1977a) etc., et nous n'y reviendrons pas. Rappelons seulement que lorsque les prospections sont régulièrement répétées avec des techniques standard pendant plusieurs années, elles fournissent un outil de contrôle de l'état des ressources. L'indice d'abondance obtenu par le bateau de recherche peut remplacer la capture par unité d'effort de la flottille quand les données de la pêche professionnelle n'ont pas les qualités requises, et quand la flottille et le bateau de recherche exploitent bien la même population.
Rappelons également que le rendement moyen obtenu par un bateau de recherche pendant une prospection planifiée, sur un stock vierge ne peut être comparé au rendement d'un bateau de pêche en opération commerciale car la distribution des efforts respectifs est vraisemblablement différente, la pêche professionnelle sélectionnant les strates les plus riches. Si la comparaison est nécessaire, une sélection des stations à meilleur rendement peut corriger l'erreur mais avec excès car le rendement ainsi calculé sera supérieur à celui qu'obtiendra en moyenne une flottille de crevettiers en compétition sur la même zone.
En fait, le rendement calculé par sélection des meilleures zones d'un stock vierge se rapproche de l'indice d'abondance théorique pour F = O (dans un modèle de production) et il faut noter que cette valeur peut être très différente (surtout pour des animaux à vie courte) de celle qu'obtiendra plus tard une pêcherie bien développée. Lindner (1971) rappelle que si les rendements obtenus sur les stocks vierges du golfe du Mexique atteignirent deux tonnes de queues par jour en 1936, ils n'étaient plus que de 450 kg/jour en 1946. Garcia (non publié) a également noté, lors du démarrage de la pêcherie de Côte d'Ivoire des rendements très élevés atteignant certains jours 700 kg/heure et une baisse très rapide dès les premiers mois de la mise en exploitation régulière. Les rendements actuels, à un niveau proche de l'optimum d'exploitation sont de 30 kg/heure.
11.3.1.2 Utilisation des résultats des prospections pour évaluer le potentiel ou le niveau d'exploitation
Lorsqu'on dispose d'une évaluation de la biomasse Bo d'un stock vierge, Gulland (1970a) indique que, si l'on admet que la ressource obéït au modèle de Schaefer et qu'au niveau du maximum de production les mortalités naturelle et par pêche sont voisines (M Fmax)1, la biomasse à ce niveau (Bmax) étant égale à Bo/2, l'expression classique Cmax = Fmax.Bmax devient
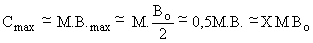
où M peut être obtenu suivant l'une des méthodes approchées indiquées au chapitre 7. Le coefficient X, ci-dessus fixé à 0,5 peut être affiné pour tenir compte des caractéristiques dynamiques des espèces considérées et en particulier de 1c/L∞ et M/K. On peut en effet écrire
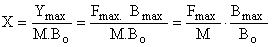
Si l'on se place dans le cas d'un modèle à recrutement constant, les tables IV de Beverton et Holt (1964) donnent un rapport (Y'/F)Emax/(Y'/F)o égal Bmax/Bo et la valeur (Fmax/M) correspondante, pour toute valeur de M/K et lc/1∞.
Dans le cas des penaeïdes, les valeurs les plus vraisemblables de M (voir chapitre 7) et de K conduisent à un rapport M/K compris entre 1 et 2 alors que lc/L∞ est proche de 0,4–0,5 pour la pêche en mer. Si l'on se reporte à la table 1 de Gulland (1970a) cela conduit à des valeurs de X comprises entre 0,32 et 0,44. Notons qu'en première approximation l'erreur faite en utilisant X = 0,5 est certainement inférieure aux erreurs sur la valeur de la biomasse elle-même. Notons également que le calcul conduit à des productions maximales qui sont de l'ordre de grandeur de la biomasse vierge dès que M est supérieur à 3,0 sur une base annuelle (ou 0,25/mois). Il est intéressant de noter, en conséquence, que pour des taux d'exploitation inconnus mais compris entre 0 et Emax' l'éventail des valeurs possibles du potentiel se situe approximativement entre B(pour E = O) et MB (pour E = Emax).
Une difficulté importante est rencontrée lorsque l'estimation de biomasse porte sur un stock déjà exploité. Les équations simples précédentes ne peuvent s'appliquer. Si la capture annuelle réalisée est connue, on peut se rappeler que Ci = Fi.Bi et donc que Ci/Bi = Fi et comparer cette valeur de Fi à une estimation de M pour l'espèce considérée. Si Fi est proche de M, on peut penser que la capture totale ne pourra guère être augmentée. En revanche, si Fi est nettement inférieur à M l'effort exercé doit pouvoir être multiplié par M/Fi pour approcher le niveau de production maximale équilibrée. Celle-ci ne peut être estimée ainsi mais on sait, par la courbe du modèle de production, que l'augmentation de capture sera inférieure à l'augmentation de l'effort, donc Cmax sera inférieur à Ci.M/Fi.
On sait d'autre part que Cmax sera compris entre B et MB selon le taux d'exploitation exercé au moment de la prospection (si M est égal ou supérieur à 0,25/mois).
11.3.1.3 Remarques concernant la méthode
Il convient de terminer ce paragraphe par quelques remarques importantes. La première concerne l'évaluation de la biomasse. Il s'agit ici d'une valeur moyenne annuelle. On sait que pour des animaux à vie courte comme les penaeïdes elle varie beaucoup dans l'année et la biomasse moyenne ainsi que son intervalle de confiance devront donc être estimés par des observations répétées au cours d'un cycle annuel complet. D'autre part, la capture réalisable sur un stock donné dépendra des variations saisonnières de l'effort. Ces commentaires, ajoutés aux remarques précédentes concernant les difficultés rencontrées à évaluer l'efficacité réelle du chalut comme estimation de la densité, indiquent que les valeurs obtenues, aussi bien pour la biomasse que pour le potentiel de capture, doivent être considérées comme de toutes premières approximations à utiliser avec précaution et en prenant des marges de sécurité suffisantes.
La deuxième remarque a trait au fondement même de la méthode qui admet qu'au niveau du maximum de production M est proche de F. Ce théorème est discuté par Francis (1974) qui démontre qu'il n'est acceptable que dans le cas d'un recrutement indépendant de la densité du stock (relation “stock-recrutement” de type Beverton et Holt). Il a été indiqué au chapitre 9 que la forme de la relation stock-recrutement des penaeïdes n'était pas connue mais qu'on tendait à considérer le recrutement comme indépendant du stock parental. Si ceci est vérifié, l'utilisation de la formule approchée Cmax = X.M.Bo serait donc pleinement justifiée pour les penaeïdes.
11.3.2 Evaluation à partir des indices de productivité
Il est toujours tentant d'essayer de réduire le temps et l'information nécessaires à l'évaluation des potentiels de capture d'une région en la comparant à une autre bien connue, de productivité et de caractéristiques écologiques analogues.
11.3.2.1 Les indices morphoédaphiques
Utilisés dans les systèmes lacustres et les rivières ils permettent, à partir de quelques caractéristiques morphoédaphiques simples (teneur en solides dissous, profondeur moyenne d'un lac, longueur du cours d'eau, surface du bassin versant) de calculer la production (en kg/ha/an par exemple) de systèmes situés dans des contextes climatiques analogues et exposés à des taux d'exploitation identiques. Un exposé détaillé des hypothèses sur lesquelles repose l'utilisation de ces indices sera trouvé dans Ryder et al. (1974) et un exemple concernant les rivières africaines dans Welcomme (1976).
La diversification des milieux marins et la complexité de leur étude n'a pas encore permis la généralisation de tels indices. On peut trouver dans le travail de Cushing (1969) un exemple des difficultés que présente le calcul des potentiels halieutiques des zones d'upwelling à partir des données de productivité planctonique.
En ce qui concerne les crevettes penaeïdes, leur liaison évidente avec les mangroves a été examinée par MacNae (1974) dans l'océan Indien. La production capturée varie de 1,3 à 10,6 tonnes/km2 de mangrove suivant les régions. Cette dispersion des valeurs n'est pas surprenante quand on sait qu'une même espèce apparaît étroitement inféodée aux mangroves dans telle région et abonde aussi ailleurs, où les mangroves n'existent pas. Notons par exemple que les penaeïdes juvéniles sont rencontrés dans les plages de sable pur (au Japon), les phanérogames marines (dans le golfe du Mexique, au Koweït) ou les palétuviers (en Malaisie, Inde, Afrique de l'Ouest). En outre, dans une même région des genres voisins (Penaeus et Parapenaeopsis par exemple) ont des exigences différentes.
La seule liaison universelle est celle que l'on observe partout entre les pénaeïdes juvéniles et l'interface mer-continent. C'est ainsi que Turner (1977) a montré une liaison entre la production lagunaire de crevettes et la surface des zones intertidales au niveau régional.
D'autre part, il est évident que la production des crevettes dépend de nombreux facteurs imbriqués tels que temperature, pluies et apports fluviatiles, reproduction, croissance, taille des estuaires et de la zone de répartition des adultes, etc. Il est clair qu'il existe un gradient latitudinal de ces paramètres et que les zones chaudes et humides sont aussi celles où les apports fluviatiles sont importants et où les lagunes, deltas et fonds sablonneux abondent, où la croissance est rapide et la reproduction continue. C'est ainsi que Turner (1977) a montré, au niveau mondial, une bonne corrélation entre la production crevettière (Y) d'une région (en kg/ha/an) et la latitude en degrés (X), de la forme
Y = 159 e-0,070(X).
Cette relation explique à elle seule 55 pour cent de la variance observée, ce qui est important. Elle est intéressante d'un point de vue théorique mais peu utilisable d'un point de vue pratique car, bien que la variance autour de la courbe n'ait pas été donnée, les valeurs de Y observées pour une latitude donnée varient de 1 à 10, “précision” très largement insuffisante pour une évaluation même grossière du potentiel d'une région.
Notons enfin que de telles études devraient utiliser pour la production biologique des indices standards (potentiels et prises maximales équilibrées par exemple), et non les captures annuelles, ce qui introduit une variabilité parasite.
11.3.2.2 Les comparaisons directes des productions
On peut également examiner la relation qui existe, dans une région donnée, entre la superficie des fonds de pêche connus et leur production maximale équilibrée pour s'en servir comme outil de prédiction du potentiel de ressources moins bien connues. Garcia et Lhomme (1979) et Lhomme (1980) notent que pour plusieurs fonds à crevettes roses (P. notialis) de la côte ouest africaine, la productivité potentielle maximale moyenne (MSY/mille carré/an) varie seulement de 1,6 (à l'extrême limite septentrionale de la distribution de l'espèce) à 4,5 t/mille carré/an. Il est intéressant de comparer ces valeurs à celles obtenues dans l'océan Indien (FAO/CPOI, 1973). Si l'on considère les données disponibles du Mozambique à la Thailande et y compris l'Australie les captures “optimales” varient de 1,7 à 3,8 t/mille carré/an, la valeur moyenne étant de 2,4 t/mille carré/an. Les valeurs extrêmes diffèrent seulement par un facteur de 2 ou 3.
L'utilisation de tels indices simples devrait permettre d'évaluer l'ordre de grandeur du potentiel d'un stock connaissant l'aire de distribution de la ressource. Cette approche a été utilisée par exemple par Garcia et Lhomme (1979) sur la côte ouest-africaine.
On ne peut cependant nier les difficultés liées à l'usage de ce type d'approche et qui sont les suivantes:
Si les superficies des stocks exploités ne sont pas toujours connues et sont souvent imprécises, celles des ressources non exploitées risquent d'être encore plus délicates à définir. On peut se baser sur des cartes sédimentologiques si la bionomie de la région est connue mais la relative adaptabilité apparente des crevettes pour divers types de sédiments meubles rend la délimitation des fonds sur cette seule donnée trop hasardeuse.
La production maximale moyenne réelle du stock dépendra finalement beaucoup de la répartition de l'effort de pêche annuel dans l'espace et sur les différentes espèces, et du niveau de la pêche artisanale et des rejets, lesquels varient avec les espèces, les régions, les types d'exploitation et les marchés. En réalité, dans quelques cas où elle a été calculée, la prise totale (artisanale sur les juvéniles et industrielle sur les adultes) est peu sensible à des variations raisonnables du taux d'exploitation sur les juvéniles (cf, chapitre 12.5).
Malgré toutes ces causes d'erreurs, sur une quinzaine de stocks observés, les résultats concernant la productivité sont suffisamment semblables pour qu'on accorde quelque crédit à la méthode étant bien entendu que le résultat doit être considéré comme provisoire et doit être vérifié, d'une part par des prospections, avant mise en exploitation (cf. paragraphe précédent) ou par les techniques classiques d'évaluation dès son démarrage.
L'approche proposée ici pour évaluer très grossièrement les potentiels a été également suggérée par Boerema (1974) pour évaluer le niveau d'effort à ne pas dépasser. Il indique qu'on peut supposer que la mortalité par unité d'intensité de pêche appliquée par un chalutier standard est la même pour toutes les espèces à comportement équivalent (et on pourrait ajouter, à des latitudes similaires). Si ces espèces ont, en plus, des propriétés biologiques voisines (K, L∞, M, fécondité, etc.) le niveau de production maximale devrait être obtenu pour des intensités de pêche équivalentes.
Cette méthode évidemment grossière permettrait de fixer, dès le début d'une exploitation l'ordre de grandeur de l'effort à ne pas dépasser et de limiter ainsi le nombre de licences à un niveau raisonnable. Une telle démarche appliquée en 1968 en Côte d'Ivoire avait permis, en comparant les fonds de pêche de ce pays à ceux du Sénégal et de Floride, de limiter à 10 le nombre de bateaux autorisés alors que le nombre de demandes atteignait 32 (Garcia, non publié). Beaucoup plus tard, les résultats d'une étude analytique par simulation basée sur des années de recherche confirmaient les prévisions (Garcia, 1977).
11.3.3 Conclusions
La nécessité de fournir, dans bien des cas, un premier diagnostic rapide pour éviter un regrettable surinvestissement plaide pour l'utilisation (prudente) de méthodes simples, rapides et peu coûteuses en information. Les résultats obtenus doivent cependant être considérés comme peu précis et très provisoires, donnant seulement des ordres de grandeur et destinés à éviter de grossières erreurs d'aménagement. Leur utilisation doit seulement être la toute première phase d'un programme de recherches visant à mieux identifier la ressource et ses réactions à l'exploitation de manière à affiner le diagnostic et à apporter les éléments d'un aménagement plus sophistiqué si nécessaire.
![]()
![]()
![]()