![]()
![]()
![]()
QUELQUES ASPECTS BIOLOGIQUES DES STOCKS DE MERLUS DANS LA DIVISION STATISTIQUE DU SAHARA LITTORAL
(34.1.3)
en 1979–1982
par
J. Bourdine, AtlantNIRO, KALININGRAD, URSS
1. LA PECHE
Qua tre périodes peuvent être distinguées dans l'exploitation des merlus au large des côtes de l'Atlantique de Nord-Quest :
Avant 1965, les stocks ont été faiblement exploités par les flottes portugaises et marocaines dans la zone septentrionale. Le tonnage capturé variait alors entre 5.000 et 10.000 tonnes.
Pendant la période 1965–1971, les prises ont brusquement augmenté atteignent 30.000 t par an. Cette augmentation s'explique par la mise en exploitation de la région du Cap Blanc, où 10–15.000 tonnes furent enregistrées.
Une nouvelle augmentation des captures intervient à partir de l'année 1972. Un niveau maximum de 121.500 tonnes fut atteint, grâce aux fortes générations de 1969, 1970, et 1971 et à l'intensification de la pêche. Les prises soviétiques de merlus atteignent 80.000 tonnes.
A partir de l'année 1975, les prises de merlus de tous les pays diminuent à cause de la dispartition des fortes classes d'âge. Les captures soviétiques ne dépassent alors plus le niveau à 5.000 à 10.000 tonnes par an.
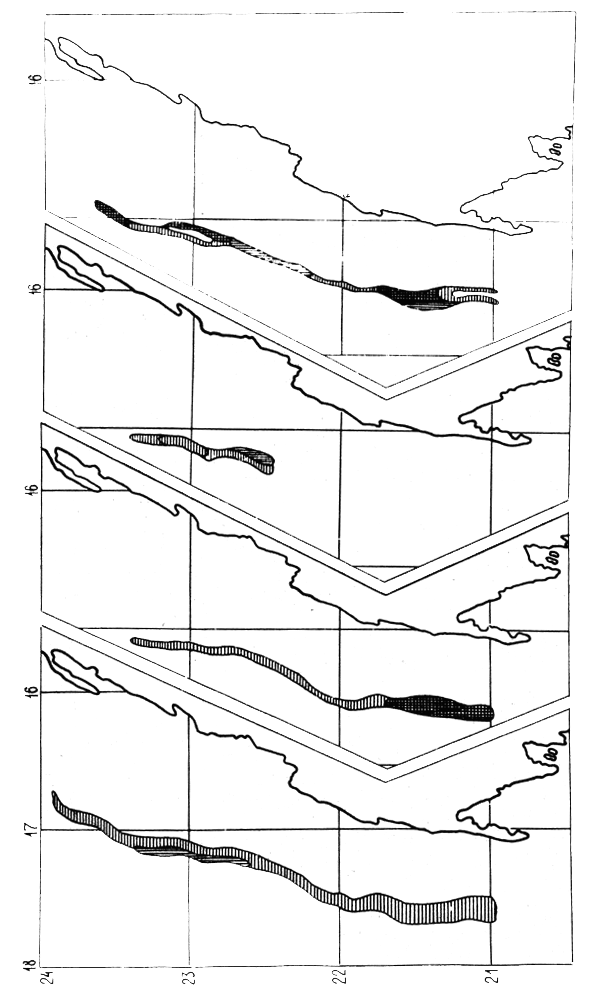
A l'heure actuelle, la pêche de l'URSS au chalut est pratiquée dans la zone de pêche libre, de la division statistique 34.1.3 ainsi que sous licence dans d'autres zones. Etant donné que la pêche est spécialisée sur les petites espèces pélagiques côtières, les merlus ne sont actuellement que des espèces secondaires, capturées accidentellement en petites quantités quand elles forment des concentrations trophiques ou de préponte. A la figure 1 sont illustrées les captures soviétiques de merlus dans la division statistique 34.1.3 constituant la zone principale de pêche.
2. COMPOSITION EN TAILLES ET AGES
Le tableau1 résume pour la période 1950–1982, fréquences de tailles des M. senegalensis dans les cptures des bateaux de pêche et les navires océanographiques dans la division 34.1.3 Au tableau 2 est présentée la fréquence de longueur d'un échantillon de 433 M. merluccius capturé lors d'une prospection par chalutage en mars 1982 dans cette même division. Ces deux tableaux montrent que de jeunes merlus ne sont présents qu'en petites quantités dans les prises des navires océanographiques qui emploient pour des buts de recherche des filets à mailles de 10 mm.
La composition en àge de M. senegalensis figure au tableau 3, elle est calculée à la base des clés taille-âge, dont les moyeenes sont présentées au tableau 4. Le tableau 5 illustre de la même façon la composition des âges de M. merluccius en mars 1982, basée sur les tailles moyennes du tableau 6.
3. MORTALITE NATURELLE
Le facteur le moins étudié jusqu'à présent, influant sur la dynamique des stocks halieutiques est la mortalité naturelle. Plusieurs méthodes existent pour en estimer l'order de grandeur. Rikhter en Efanov (1977) se basent sur l'âge de la maturation des poissons, tandis que d'autres (Pauly, 1980, (Lukachov, 1970) utilisent des paramètres de l'équation de von Bertalanffy. Le premier auteur utilise ensuite comme indice de l'anabolisme la température moyenne de l'eau dans laquelle habite le poisson, le second l'âge de la première maturation. Tiurin (1962, 1972) propose d'estimer le taux de mortalité natrelle pour chaque âge à l'aide d'un tableau exponentiel conçu par Baranov (1925) et d'un graphique logarithmique.
Pour l'estimation du taux instantané de mortalité M ches M. senegalensis nous avons utilisé les paramètres de l'équation de von Bertalanffy suivants W∞ = 1680, 3 g, L∞ = 61,4 K = 0,24 et to = - 0,26, ainsi que X (lâge de maturation de masse) =3,1 ans, t (âge de première maturation) = 2,9 ans et T (température moyenne de l'habitat du merlu) = 15°C, l'âge limite étant de ll ans. Cette espèce constituait moins de 2% de la biomasse relative calculés.
i. Selon Rikhter et Efanov 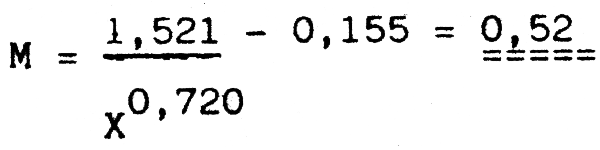
ii. Selon les méthodes de Pauly :
log M = - 0,2107 - 0,0824 log W∞ + 0,6757 log K + 0,4627 log T M = 0,45
log M = - 0,0066 - 0,278 log L∞ + 0,6543 log K + 0,4634 log T
iii. Selon Lukachov Qt (M) = 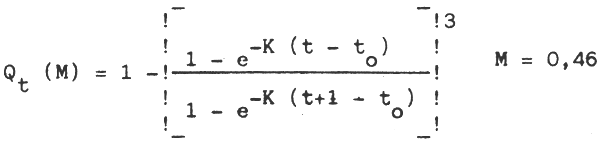
iv. l'application de la méthode proposée par Tiurin mène à un nombre de valeurs de M pour différents âges, dont la moyenne est de M = 0,410 (tableau 7).
4. AUTRES DONNEES BIOLOGIQUES
Trois espèces de merlus habitant la division stastique 34.1.3, mais leur importance relative est loin d'être égale. D'après les prospections par chalutage effectuées en 1979–1982, M. polli, et même compleètement absent sauf en mars 1982, quant M. merluccius constituait 39% dabs ke secteyr 22°35' – 23°40'. Le schéme de répartion des merlus est illustré à la firue 2. La figure 3 et le tableau 8 illustrent le sex ration chez M. senegalensis obtenu lors des observations en 1979–1982 en 34.1.3.
Les rapports taille-poids pour M. merluccius et M. senegalensis sont présentés en figure 4.
Huit propestions quantitatives par chalutage, effectuées en 1980– 1982 dans le secteur 21°23°50N de la division statistique 34.1.3 (en mars 1982, la prospection a été limité à la zone de 22°35'–23°40N), ont permis d'évaluer le “stock minimum” de M. merluccius et M. senegalensis. Les figures 5 et 6a, b illustrent la répartition et la densité des concentration des merlus et les tableaux 9 to 10 leur “stock minimum”. Des variations importantes apparente dans ces résultats s'expliquent par la répartition saisonnière des merlus et par la méthodologique de prospection. Les grands bateaux peuvent faire des chalutages à des profondeures d'au moins 600 mètres, les moyens jusqu'à 300 m.
Le volume maximal du stock coincide en effect avec la période d'apparition de concentrations commerciales.
5. REFERENCES BIBLIOGRAPHIQUES
Baranov, F. I., 1925 La pêche et l'âge lmite des poissons. Pytchevaya promytchlennost: 65–67. (en russe)
Lukachov, V. N., 1970 Méthode de calcul de la plus petite taille commercial du poisson. Tr. VNIRO 21 (2) : 281–293
Maurin, C., 1968 Ecologie ichthyologiues des fonds chalutables atlantiques (de la baie Ibero-marocaine à la Mauritanie) et de la Méditerranée Occidentale. Rev.Trav.Inst.Pêche Marit.Nantes., 32 (2) : 147p.
Pauly, D., 1980 On the interrelationship between natural mortality, growth parameters and mean environmental temperature in 175 fish stocks. J.Cons.CIEM 39 (2) : 175–192.
Rikhter, V.A. et V.N. 1977 Efanov, sur une des méthodes d'évaluation de la mortalité naturelle des populations des poissons. Tr. AtlantNIRO 23 : 77–85.
Tiurin, P.V., 1962 Le facteur de mortalité naturelle des poissons et son importance pour la régulation des pêches. Problème de l'Ichthologie 2, 3 (24) : 403–421.
Tiurin, P.V., 1972 Courbes “normales” de survivance et taux de mortalité naturelle des poissons comme base théorique de la régulation des pêches. Nouv.Cos.NIORKH, 71 : 71–128.
| IV–V 80x | VII-80xx | VI-81xx | VIII-81x | I-82xx | VI-VII 82xxx | VIII-82x | |
|---|---|---|---|---|---|---|---|
| 12 | |||||||
| 14 | |||||||
| 16 | 1 | ||||||
| 18 | 2 | 4 | |||||
| 20 | 2 | 6 | 12 | ||||
| 22 | 4 | 8 | 27 | ||||
| 24 | 3 | 1 | 33 | 36 | |||
| 26 | 9 | 2 | 3 | 152 | 40 | 29 | |
| 28 | 14 | 2 | 1 | 244 | 139 | 49 | |
| 30 | 42 | 15 | 4 | 254 | 240 | 59 | |
| 32 | 55 | 23 | 12 | 112 | 15 | 203 | 34 |
| 34 | 32 | 7 | 4 | 98 | 30 | 229 | 85 |
| 36 | 37 | 9 | 8 | 104 | 48 | 724 | 63 |
| 38 | 30 | 8 | 11 | 127 | 30 | 1462 | 79 |
| 40 | 50 | 9 | 31 | 181 | 19 | 1164 | 68 |
| 42 | 35 | 21 | 83 | 79 | 13 | 440 | 71 |
| 44 | 31 | 24 | 30 | 73 | 4 | 196 | 64 |
| 46 | 7 | 14 | 10 | 37 | - | 57 | 28 |
| 48 | 13 | 15 | 12 | 17 | 2 | 36 | 9 |
| 50 | 11 | 22 | 12 | 14 | 3 | 13 | 9 |
| 52 | 5 | 9 | 125 | 4 | 1 | 9 | 9 |
| 54 | 2 | 9 | 5 | - | 15 | 6 | |
| 56 | 2 | 4 | 2 | - | 9 | 4 | |
| 58 | 2 | 2 | 1 | 1 | 10 | 8 | |
| 60 | 2 | 1 | 1 | - | 4 | 4 | |
| 62 | - | 1 | - | - | 2 | 5 | |
| 64 | - | 2 | - | - | 1 | - | |
| 66 | - | 1 | 2 | - | - | 1 | |
| 68 | 1 | 1 | 1 | - | - | ||
| 70 | 1 | - | 1 | ||||
| 72 | - | ||||||
| 74 | 1 | ||||||
| Nmbre | |||||||
| 387 | 200 | 227 | 1558 | 167 | 4994 | 765 | |
| d'ex. |
x - des prises des navires scientifiques
xx - des prises des bateaux de pêche
xxx - Merluccius sp. des prises des bateaux de pêche
| III-1982X | |
|---|---|
| Longuear, cm | |
| 16 | 1 |
| 18 | - |
| 20 | - |
| 22 | 4 |
| 24 | 9 |
| 26 | 63 |
| 28 | 96 |
| 30 | 111 |
| 32 | 69 |
| 34 | 53 |
| 36 | 9 |
| 38 | 1 |
| 40 | 5 |
| 42 | 3 |
| 44 | 2 |
| 46 | 2 |
| 48 | - |
| 50 | - |
| 52 | 1 |
| 54 | 2 |
| 56 | - |
| 58 | - |
| 60 | - |
| 62 | 1 |
| 64 | - |
| 66 | 1 |
| 68 | |
| Nmbre d'ex. | 433 |
X - des prises des navires scientifiques.
| Age | Année IV–V 80X | VII-80XX | VI-81XX | VIII-81X | 1–82XX | VIII-82XX |
|---|---|---|---|---|---|---|
| I | 1,3 | - | 0,4 | 3,2 | - | |
| II | 22,4 | 13,1 | 9,6 | 35,9 | - | 10,2 |
| III | 47,1 | 34,5 | 40,2 | 36,53 | 3,1 | 16,8 |
| IV | 26,0 | 33,0 | 34,5 | 18,8 | 41,1 | 20,7 |
| V | 5,7 | 11,0 | 11,4 | 4,0 | 41,2 | 29,2 |
| VI | 2,6 | 5,8 | 3,5 | 1,4 | 11,0 | 11,6 |
| VII | 0,3 | 2,6 | 0,4 | 0,1 | 1,8 | 5,7 |
| plus de VII 0,3 | plus de VII | plus de VII | ||||
| 1.8 | 5,8 |
X des prises des navires scientifiques
XX des prises des bateaux de pêche
| Année de | Âge, ans | ||||||
|---|---|---|---|---|---|---|---|
| l'élaboration | |||||||
| des clés | I | II | III | IV | V | VI | VII |
| 1980–1981 | 15,7 | 25,1 | 33,4 | 39,4 | 43,9 | 47,6 | - |
| 1982 | - | - | 31,0 | 37,8 | 39,3 | 43,1 | 46,2 |
| Age | III-82 |
|---|---|
| I | 0,2 |
| II | 14,3 |
| III | 38,1 |
| IV | 35,6 |
| V | 7,6 |
| V | 4,2 |
Clé “taille-âge” du merlu européen
| Année de | Âge, ans | |||
|---|---|---|---|---|
| l'élaboration | ||||
| de la clé | I | II | III | IV |
| 1982 | - | 25,1 | 29,7 | 31,5 |
| Age, ans | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI |
| 4,60 | 0,92 | 0,46 | 0,34 | 0,36 | 0,42 | 0,54 | 0,73 | 1,04 | 4,60 | |
| 1,63 | ||||||||||
| Proportion | Mois | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (‰) | I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII |
| 86,5 | 91,2 | 69,8 | 63,3 | 42,6 | - | - | 44,5 | - | 77,0 | 75,9 | - | |
| 13,5 | 8,8 | 30,0 | 36,7 | 57,4 | 55,6 | 23,0 | 24,1 | |||||
| Indices | Periode | |||||||
|---|---|---|---|---|---|---|---|---|
| III–IV 80 | IV–V 80X | V 80 | VIII 80 | X–XI 80X | VIII–IX 81X | III 82X | VIII 82X | |
| “Abondance | 0,4 | 1,1 | 0,3 | 1,0 | 2,4 | 16,0 | 0,4 | 17,4 |
| minimum”, | ||||||||
| millions d'ex. | ||||||||
| “Biomasse | ||||||||
| minimum”. | 0,08 | 0,5 | 0,07 | 0,5 | 1,2 | 4,8 | 0,2 | 5,8 |
| milliers de t | ||||||||
x Les prospections ont été accomplies par les bateaux à grand tonnage.
| Indices | Période | |
|---|---|---|
| X–XI 80 | III 82 | |
| “Abondance minimum”, milliers d'ex. | 33,0 | 508,0 |
| “Biomasse minimum”, t | 32,0 | 22,0 |
Figure 1 : Evolution annuelle des captures de merlus par la flotte soviétique dans la division statistique 34.1.3. En 1979 (*) la pêche n'a commencé qu'en Septembre.
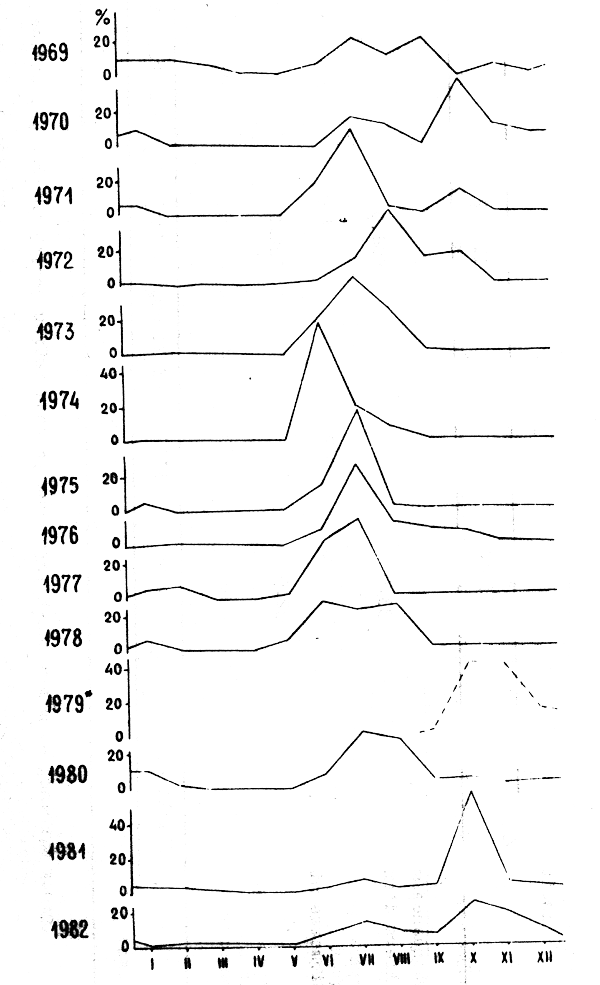
Figure 2 : Répartition géographique des trois espèces de merlus d'après les données soviétiques. 1. Merluccius merluccius, 2. Merluccius senegalensis, 3. Merluccius polli
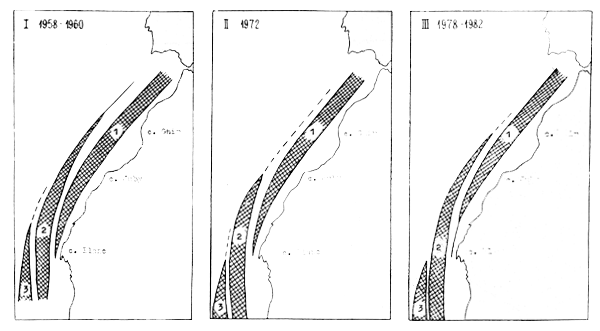
Figure 3 : Proportion taille-sexe chez M. senegalensis en 34.1.3
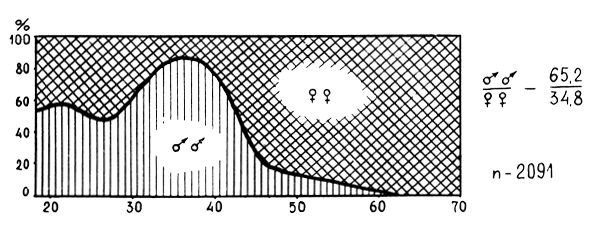
Figure 4 : Relation longueur-poids chez A) M. merluccius, B) M. senegalensis en division 34.1.3
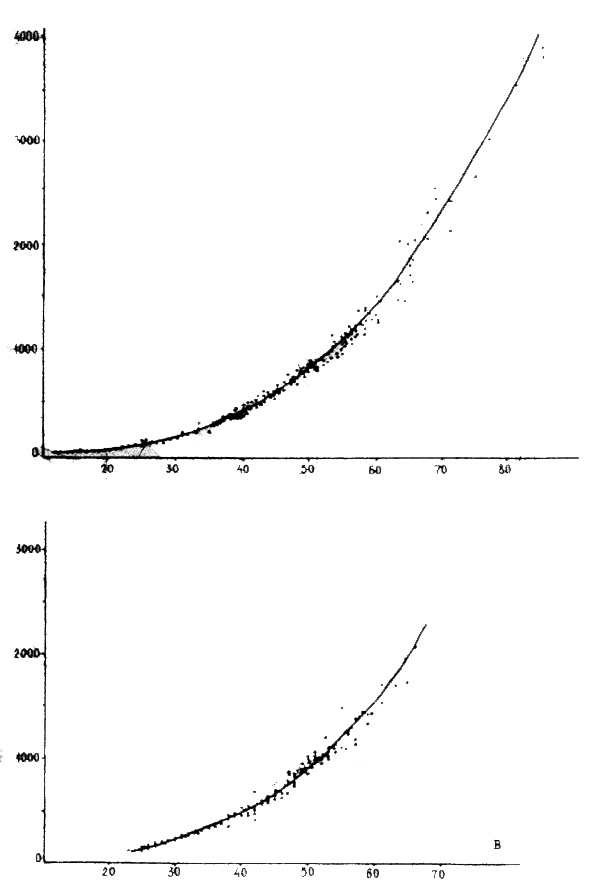
Figure 5 : Distribution de M. merluccius dans la division statistique 34.1.3 A : Octobre / Novembre 1980, B : Mars 1982
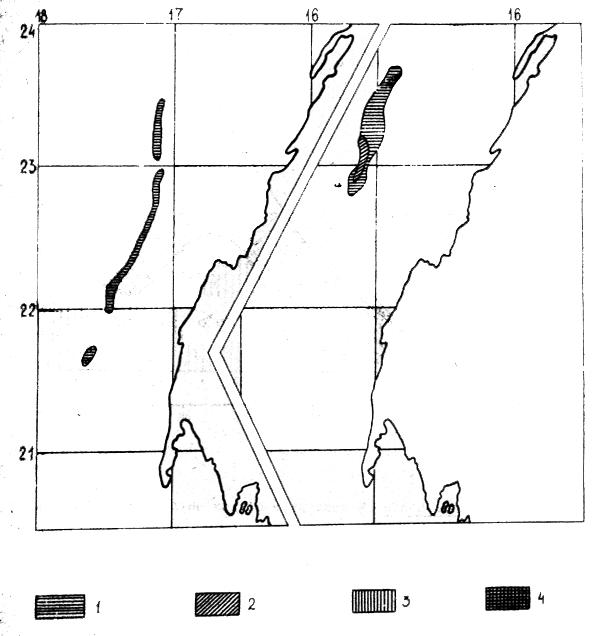
Figure 6a : M. senegalensis en division statistique 34.1.3
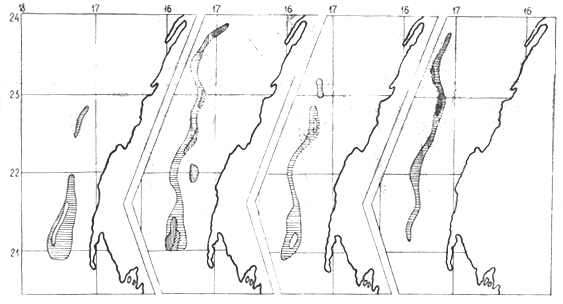
Figure 6b : M. senegalensis en division statistique 31.1.3
NOTE SUR LA TRANSFORMATION DES DISTRIBUTIONS DES
TAILLES EN DISTRIBUTIONS DE FREQUENCE DES AGES:
RAPPEL DES BIAIS ET DES SOLUTIONS
par
S. Garcia 1
1. INTRODUCTION
Compte tenu de la difficulté à lire l'âge des espèces tropicales et du nombre très important de ces dernières, il faut prévoir dans les années prochaines une intensification notable des travaux basés sur l'analyse des données de longueur. La méthodologie est en cours d'élaboration à l'heure actuelle (Jones 1985, pauly 1980). De nombreuses méthodes sont proposées mais les problèmes liés à leur utilisation commencent à être perçus et des solutions sont élaborées (voir Hoenig, Lawing et Hoenig 1983, Majkowski 1982, Bartoo et Parker 1983, Pereiro et Pallares 1984).
En particulier, l'utilisation des courbes de capture par classe de taille converties “length converted catch curves” (pauly 1983) ainsi que l'analyse des cohortes par classes de longueur se développera très certainement rapidement. Leur élaboration est relativement simple mais soulève quelques difficultés mineures que nous examinerons dans le paragraphe suivant.
2. ELABORATION DES COURBES DE CAPTURE PAR CLASSES DE TAILLE
Le principe a très souvent été utilisé dans le passé. Il consiste à utiliser une courbe de croissance (de von Bertalanffy par exemple) pour “découper” une distribution de fréquence des tailles en groupes d'âge dont les effectifs peuvent être alors approximativement calculés. La procédure la plus courante consiste à découper la distribution en tranches égales annuelles (pour les poissons à vie longue) ou trimestrielles ou même mensuelles (pour les animaux à vie courte). Il est alors possible d'établir une courbe de capture en portant le logarithme des effectifs calculés en fonction de l'âge et calculer ainsi la mortalité totale. Pauly (1983) propose d'utiliser les distributions de fréquence directement en calculant l'âge théorique correspondant au point médian de chaque classe de taille. Cette derniére procédure permet de conserver toute l'information et pourrait, en théorie, permettre d'apprécier des changements dans la mortalité totale à l'intérieur des grands groupes d'âge généralement adoptés par le passé. Cette méthode est celle qu'ont utilisé Turner et El Ouairi (1984) pour leur analyse des mortalités par pêches appliquées aux stocks de merlus du Maroc.
L'analyse des cohortes de Jones effectuée sur des données de longueur se rattache également à cette méthode dans la mesure où les classes de taille sont utilisées au lieu des groupes d'âge, et où on calcule le laps de temps Δt passé par les individus dans les classes de taille considérées, à l'aide des paramètres de croissance.
3. SOURCES DE BIAIS
L'utilisation des paramètres d'une courbe classique de von Bertalanffy pour le calcul des âges théoriques correspondant aux diverses classes d'âge pose quelques problèmes. Cette courbe est par définition une relation prédictive permettant de prédire la taille atteinte à un âge donné (âge = x → taille = Y).
Son utilisation pour une prédiction inverse (taille = x → âge = y) n'est pas exempte de biais et l'importance du biais est proportionnelle à l'erreur résiduelle, non expliquée par la relation (Bartoo et Parker 1983).
Jones (1985) souligne la difficulté créée par le fait qu'il est fréquent d'avoir un L∞ plus petit que les plus gros poissons capturés et mésurés. Dans ce cas, les classes de taille correspondantes doivent être rejetées, faute de pouvoir en calculer l'âge, ou regroupées artificiellement dans un même groupe d'âge (t+).
En réalité, le problème va bien au-delà car l'utilisation inverse
d'une équation de croissance classique introduit un biais dans l'estimation
des âges les plus grands, biais qui s'accroít très rapidement quand on
approche de L∞, taille au-delà de laquelle les calculs sont impossibles.
En effet, si des poissons d'âge t ont une taille moyenne, 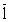 , les poissons
de taille l n'ont pas forcément un âge moyen t. Ceci a été souligné par
Bartoo et parker (1983). De leur côté,Majkowski et Hampton (1983) ont
démontré qu'en fait les erreurs dans la valeur calculée de l'âge s'accroissent
rapidement aux deux extrémités de la distribution de fréquence des tailles
convertie, c'est à dire que l'erreur commise s'accroît non seulement vers
les grandes tailles mais aussi vers les petites. Il est plus que vraisemblable
que ces erreurs de détermination conduisent également à des erreurs sur les
taux d'exploitation calculés par la méthode d'analyse des cohortes (en
longueur) de Jones.
, les poissons
de taille l n'ont pas forcément un âge moyen t. Ceci a été souligné par
Bartoo et parker (1983). De leur côté,Majkowski et Hampton (1983) ont
démontré qu'en fait les erreurs dans la valeur calculée de l'âge s'accroissent
rapidement aux deux extrémités de la distribution de fréquence des tailles
convertie, c'est à dire que l'erreur commise s'accroît non seulement vers
les grandes tailles mais aussi vers les petites. Il est plus que vraisemblable
que ces erreurs de détermination conduisent également à des erreurs sur les
taux d'exploitation calculés par la méthode d'analyse des cohortes (en
longueur) de Jones.
4. SOLUTIONS EXISTANTES
Il a été proposé, pour résoudre ces difficultés, des solutions plus ou moins simples, qu'il est bon de rappeler.
Lorsque les données de base sont disponibles, la solution la plus directe consiste à retourner aux données de base et à calculer la relation entre la taille (x) et l'âge moyen observé (y).
Lorsque les données de base ne sont pas disponibles, il existe plusiers possibilités. Jones (1985) propose de choisir un L∞ supérieur à la taille des poissons les plus gros et de calculer graphiquement la valeur de K compatible avec ce choix, c'est à dire reproduisant correctement la relation taille-âge observée pour le stock en question. La méthode consiste à calculer l'équation de la droite Ford-Walford forcée, passant par la valeur de L∞ ainsi choisie. La même procédure peut être appliquée si l'on utilise plutôt la relation entre les accroissements (lt+1 - lt) et la taille (lt).
Pereiro et Pallares (1984) montrent cependant que cette correction peut malgré tout conduire à des erreurs graves dans la détermination des mortalités si les valeurs “choisies” sont trop eloignées des valeurs réelles. Ils montrent également cependant que si l'analyse (régressive) des cohortes part d'un groupe de longueur (l+) dont la borne inférieure est située aussi loin que possible de la valeur de L∞ et aussi près que possible de l'intervalle réel de distribution des données observées, les vecteurs de F/âge calculés par la méthode de Jones sont alors relativement insensibles aux erreurs sur les paramètres de croissance.
Il semble donc que la solution simple et pratique au problème posé découlant des conclusions des auteurs cités précédemment soit:
de choisir un L∞ supérieur aux plus grandes tailles observées et de calculer une valeur compatible de K ou encore de calculer directement les paramètres de la relation taille-âge si les données sont disponibles;
de commencer l'analyse régressive à partir d'un groupe (l+) dont la borne inférieure soit située le plus loin possible du L∞ retenu;
de ne considérer comme strictement valides que les valeurs calculées (âge, durée du passage dans l'intervalle de taille, Δt, mortalité par pêche F. t, etc. ) situées dans l'intervalle des tailles observées pour le calcul des paramètres de croissance et reconnaître que les biais s'accroissent rapidement vers les plus grandes tailles et vers les plus petites.
Bartoo et Parker (1983), de leur côté, préconisent, pour résoudre le biais, une approche stochastique lorsque la distribution de fréquence des longueurs par classes d'âge est disponible.
5. REFERENCES
Bartoo, N.W. and K.P. Parker, 1983 Stochastic age frequency estimation using the von Bertalanffy equation. Fish.Bull., 81(1) :91–6
Hoenig, J.M., W.D. Lawing and N.A. Hoeing, 1983 Using mean age, mean length and medium length data to estimate the total mortality rate. ICES/CM 1983/D:23, 11 P. (mimeo)
Jones, R., 1985 Assessing the effects of changes in exploitation patterns using length composition data (with notes on VPA and cohort analysis). FAO Finsh.Tech.pap., 256 (in press)
Majkowski, J., 1982 Usefulness and applicability of sensitivity analysis in a multispecies approach to fisheries management. In Theory and management of tropical fisheries. Pauly, D. and G. Murphy (eds.). ICLARM/CSIRO:149-166
Majkowski, J. and Hampton, 1982 The effect of parameter uncertainties in an age-length relationship upon estimating the age composition of catches. Can.J.Finsh.Aquat.Sci., 40(3), 272–80
Pereiro, J.A. and P. Pallares, 1984 Consequences of introducing errors in the values of the input parameters in the “length distribution analysis” (R. Jones) as applied to the northern stock of hake. CIEM/CM:1984/G61, 7 p. (mimeo)
Pauly, D., 1980 A new methodology for rapidly acquiring basic information on tropical shrimp stocks: growth, mortality and stock recruitment relationships. In Stock assessment for tropical small-scale fisheries. Saila, S. and P. Roedel (eds.). Univ.Rhode Island Int.Center Mar.Res.Dev., 198 p.
Pauly, D., 1983 Length converted catch curves: a powerful tool for fisheries research in the tropics. Part I. Fishbyte, 1(2):9–13
Turner, J. and M. EL Ouairi, 1985 State of exploitation of Merluccius merluccius off the coast of norther Morocco. COPACE/PACE/85/33 (in press)
CONSIDERACIONES SOBRE LOS EFECTOS DE UN CAMBIO DE
MALLA EN LA PESQUERIA DE MERLUZA EUROPEA (Merluccius
merluccius) Y GAMBA BLANCA (Parapenaeus longirostris)
DEL ATLANTICO CENTRO-ORIENTAL. ASPECTOS ECONOMICOS.
por
R.GOÑI1
INTRODUCTION
La recomendación CECAF/6/1 sobre el tamaño de la malla a utilizar en la región del Comité, señala que este debe de ser de, al menos, 60 mm. Con el fin de estudiar la viabilidad de esta malla para la pesquería mixta merluza-gamba de caladeros situados al norte de Cabo Juby, se ha realizado una serie de estudios conjuntos Hispano-Marroquíes sobre la selectividad del arte de arrastre sobre estas especies. Los resultados de estas experiencias están parcialmente recopilados en los documentos de Belvèze, Bravo y Goñi y Delgado, 1981.
En la presente nota se hace un estudio de los efectos que un posible cambio de malla tendría sobre las capturas españolas de estas especies, a corto y largo plazo, así como una económica de los mismos.
Un estudio similar, referente unicamente a la gamba blanca, se presentó a la 6a Sesión del Grupo de Trabajo de Evaluación de Recursos de CECAF (CECAF, 1982). Este trabajo se consideró como preliminar ya que los datos de base correspondían al 4o trimestre de 1981; siendo este periodo el de iniciación del sistema de muestreo de las capturas españolas en caladeros marroquíes, la cobertura del mismo era todavía parcial.
MATERIAL Y METODOS
En este estudio se ha seguido el método propuesto por E.L. Cadima (1978) como modificación al método de Holt (1957).
Se ha tomado como malla actual la de 40 mm y se han analizado las consecuencias que tendría el cambio a las mallas de 50, 60 y 70 mm.
Los procentajes de retención utilizados en el caso de la merluza corresponden a curvas de selección paralelas a la obtenida para 40 mm durante la campaña ELID 8005 (Belvèze, Bravo y Goñi, 1980) (Tabla 1), considerando un valor del factor de selección de 3.6 (CECAF, 1978). Este valor del factor de selección se encuentra próximo a los calculados a partir de los datos extraidos de las experiencias de selectividad conjuntas mencionadas.
Para la gamba se han utilizado los porcentajes de retención de las curvas de selección teóricas calculadas a partir de los valores reales de las campañas ELID 8005, IBN SINA 8105 e IBN SINA 8112 (Tabla III).
1 Centro Costero de Canarias (IEO) Crta. San Andrés s/n Santa Cruz de Tenerife, España
La relación talla-peso aplicada en el caso de la gamba es:
W(gr)=0.00398 L3.08(cm)
Y en el de la merluza:
W(gr)= 0.013 L2.87(cm)
Las valoraciones de pérdidas inmediatas y posibles ganancias a la larga se han hecho sobre composiciones por tallas de la captura española actual de merluza y gamba de Marruecos, correspondiente al primer semestre de 1982, considerando que han sido realizadas con copos de malla de 40 mm.
Dada la falta de conocimiento de la tasa de explotación a que está sometido este stock se han probado tres valores de la misma: baja (E = 0.25), media (E = 0.50) y alta (E = 0.75).
La evaluación de las consecuencias económicas de los cambios de malla en la pesquería mixta se ha realizado considerando una capture conjunta de 100 kg, repartidos entre ambas especies según los porcentajes en que se presentaron en las capturas españolas del periódo 1981–1982. Los precios por kilogramo corresponden al valor medio del año 1982, teniendo en cuenta las fluctuaciones que sufre según la época del año y el puerto de descarga de quese trate.
RESULTADOS
En las Tablas I y III se incluyen las composiciones por tallas de la captura de merluza y gamba, el peso medio por ejemplar en el punto medio del intervalo de tallas y los porcentajes de retención estimados para las mallas estudiadas.
En la Tabla IV se presentan los resultados de la evaluación del cambio de la malla de 40 mm a las de 50, 60 y 70 mm correspondiente a la merluza europea. Como puede observarse en dicha Tabla, al aumentar la malla del copo las pérdidas iamediatas en peso van creciendo, desde el 14%, para la malla de 50 mm y una tasa de explotación baja, al 92% en el de 67.1 mm y tasa de explotación alta.
En la Tabla V se presenta la valoració n económica del cambio de malla contemplando en conjunto las dos especies. Se registran pérdidas inmediatas quevan desde el 25%, si se pasa a la malla de 50 mm al 61% si se pasa a la mayor.
DISCUSION
Del estudio de los efectos de un cambio de malla en la pesquería de merluza europea podría deducirse la rentabilidad, a largo plazo, de este tipo de medidas. No sucede lo mismo con la gamba blanca en que, siendo las pérdidas inmediatas muy superiores a las de la merluza, en ningún caso se registran en el futuro capturas superiores a las actuales.
Considerando la posibilidad de un cambio de la malla de 40 mm a la de 50 mm para una tasa de explotación media, en la merluza se perdería un 14%, ganándose después un 18%; sin embargo en la gamba, de inmediato, se perdería el 29% en peso, para después obtener capturas más bajas que la actual en un 12%.
En los otros posibles cambios a mallas más grandes las pérdidas en la gamba crecen mucho, tanto a corto como a largo plazo, no compensándose con las ganancias que se obtendrían en la captura de merluza al paso del tiempo.
La valoración económica del cambio de malla indica que la aplicación de esta medida conllevaría, en el mejor de los casos, unas pérdidas del 25% del valor de la captura de ambas especies en conjunto.
BIBLIOGRAFIA
Belvèze, H., J. Bravo 1980 de Laguna y R. Goñi, Selectividad del arte de arrastre sobre merluza europea (Merluccius merluccius) y gamba blanca (Parapenaeus longirostris) en la zona norte de CECAF. Comunicación presentada en la 2a Reunión del Grupo de Trabajo Especial de Merluza de CECAF. Dakar. Septiembre 1980
Cadima, E.L., 1978 The effect on yield of a change in the age at first capture. Manual for fish stock assessment. FAO, Fisheries. Circular No. 701
Goñi, R. Y A. Delgado, 1981 Informe de los trabajos realizados en la campaña IBN SINA 8105. Mimeo
Goñi, R., 1982 Consideraciones sobre los efectos de un cambio de malla en la pesquería de gamba blanca (Parapenaeus longirostris) del Atlántico Centro-Oriental. Comunicación presentada a la 6a Sesión del Grupo de Trabajo Especial de Evaluación de Recursos. Dakar. Febrero 1982
Merluccius merluccius
| Talla | C1 | P1 | P2 | P3 | P4 | |
|---|---|---|---|---|---|---|
| LT.cd. | № ej/100 |  gr gr | 89,2 mm | 50 mm | 59,7 mm | 67,1 mm |
| 7 | 41 | 4,22 | 0,2 | |||
| 8 | 393 | 6,04 | 0,4 | |||
| 9 | 2474 | 8,32 | 0,07 | |||
| 10 | 6264 | 11,09 | 0,11 | 0,1 | ||
| 11 | 12394 | 14,39 | 0,18 | 0,02 | ||
| 12 | 18875 | 18,28 | 0,27 | 0,04 | ||
| 13 | 23512 | 22,80 | 0,40 | 0,07 | ||
| 14 | 21386 | 27,90 | 0,53 | 0,11 | 0,01 | |
| 15 | 17343 | 33,90 | 0,66 | 0,18 | 0,02 | |
| 16 | 18107 | 40,56 | 0,77 | 0,27 | 0,04 | 0,1 |
| 17 | 15704 | 48,42 | 0,86 | 0,40 | 0,07 | 0,02 |
| 18 | 14095 | 56,33 | 0,91 | 0,53 | 0,11 | 0,04 |
| 19 | 12698 | 65,52 | 0,95 | 0,66 | 0,18 | 0,07 |
| 20 | 11837 | 75,63 | 0,97 | 0,77 | 0,27 | 0,11 |
| 21 | 10710 | 86,70 | 0,98 | 0,86 | 0,40 | 0,18 |
| 22 | 9788 | 98,79 | 0,99 | 0,91 | 0,53 | 0,27 |
| 23 | 10014 | 111,92 | 0,99 | 0,95 | 0,66 | 0,40 |
| 24 | 7523 | 126,14 | 1,00 | 0,97 | 0,77 | 0,53 |
| 25 | 5624 | 141,49 | 0,98 | 0,86 | 0,66 | |
| 26 | 4393 | 158,00 | 0,99 | 0,91 | 0,77 | |
| 27 | 3689,175,72 | 175,72 | 0,99 | 0,95 | 0,86 | |
| 28 | 4017 | 194,69 | 1,00 | 0,97 | 0,91 | |
| 29 | 2719 | 214,95 | 0,98 | 0,95 | ||
| 30 | 2090 | 236,53 | 0,99 | 0,97 | ||
| 31 | 1997 | 259,47 | 0,99 | 0,98 | ||
| 32 | 1872 | 283,82 | 1,00 | 0,99 | ||
| 33 | 1938 | 309,61 | 0,99 | |||
| 34 | 1752 | 336,89 | 1,00 | |||
| 35 | 1263 | 365,68 | ||||
| 36 | 1289 | 396,03 | ||||
| 37 | 1225 | 427,97 | ||||
| 38 | 940 | 641,55 | ||||
| 39 | 823 | 496,79 | ||||
| 40 | 702 | 533,75 | ||||
| 41 | 620 | 572,46 | ||||
| 42 | 521 | 612,94 | ||||
| 43 | 545 | 655,25 | ||||
| 44 | 401 | 699,42 | ||||
| 45 | 349 | 745,48 | ||||
| 46 | 342 | 793,48 | ||||
| 47 | 304 | 843,44 | ||||
| 48 | 253 | 895,41 | ||||
| 49 | 174 | 949,42 | ||||
| 50 | 195 | 1005,52 | ||||
| 51 | 124 | 1063,73 | ||||
| 52 | 105 | 1124,09 | ||||
| 53 | 93 | 1186,64 | ||||
| 54 | 76 | 1251,41 | ||||
| 55 | 55 | 1318,45 | ||||
| 56 | 53 | 1387,79 | ||||
| 57 | 48 | 1459,45 | ||||
| 58 | 36 | 1533,49 | ||||
| 59 | 21 | 1609,93 | ||||
| 60 | 17 | 1688,81 | ||||
| 61 | 16 | 1770,17 | ||||
| 62 | 12 | 1854,04 | ||||
| 63 | 6 | 1940,46 | ||||
| 64 | 11 | 2029,46 | ||||
| 65 | 2 | 2121,07 | ||||
| 66 | 2 | 2315,37 | ||||
| 67 | 7 | 2312,31 | ||||
| 68 | 9 | 2411,99 | ||||
| 69 | 2 | 2514,43 | ||||
| 70 | 1 | 2619,67 |
Tabla I (Cont.). - Datos de base para la evaluación de un cambio de malla. Merluccius merluccius. C1 = Captura actual (malla de 40 mm.) que corresponde al período Enero-Julio 1982. P1, P2, P3, P4 = Tantos por uno de retención para las mallas de 39,2, 50, 59,7, 67,1 mm. respectivamente. Estas retenciones has sido calculadas a partir de los co rrespondientes para 40 mm. suponiendo un factor de selección de 3,6. Relación talla-peso:
W(gr) = 0,013 L (cm)2.87
Merluccius merluccius.
1. Cambio de malla 1 (39,2 mm) a malla 2 (50 mm).
C1 = 25389600 C2 = 13403500 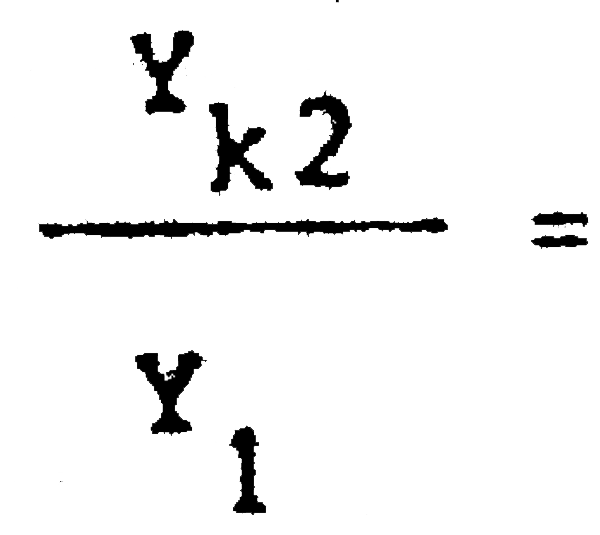 = 0,86 Pérdida inmediata del 14% de la captura actua
= 0,86 Pérdida inmediata del 14% de la captura actua
Y1 = 2288703 Yk2 = 1977484
(E = 0,25) = 1,01 Captura a la larga = 101% de la de equilibrio con 40 mm
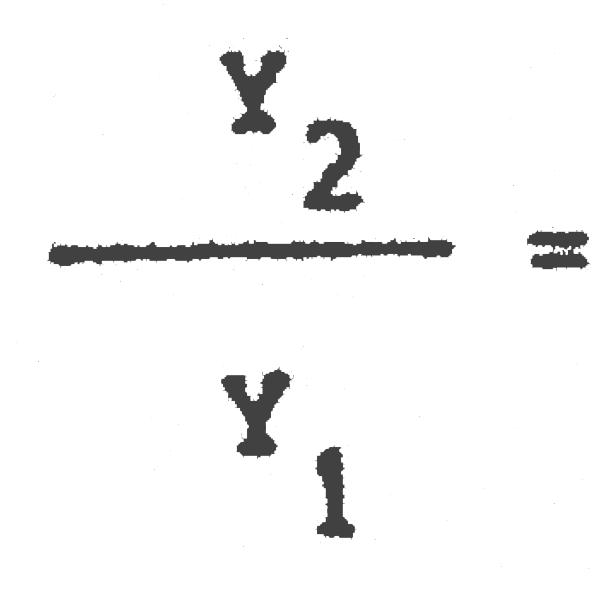 = (E = 0,50) = 1,18 Captura a la larga = 118% de la de equilibrio con 40 mm
= (E = 0,50) = 1,18 Captura a la larga = 118% de la de equilibrio con 40 mm
(E = 0,75) = 1,38 Captura a la larga = 138% de la de equilibrio con 40 mm
2. Cambio de malla 1 (39,2 mm) a malla 3 (59,7 mm).
C1 = 25389600 C3= 7186500 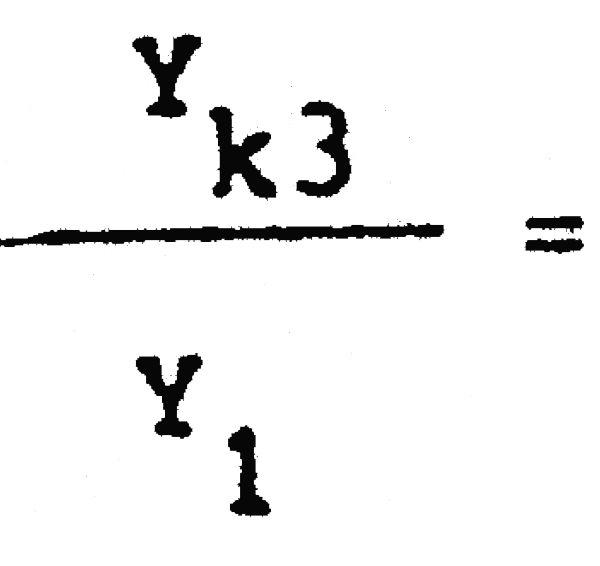 = 0,67 Pérdida inmediata del 33% de la captura actual
= 0,67 Pérdida inmediata del 33% de la captura actual
Y1 = 2288703 Yk3 = 1533363
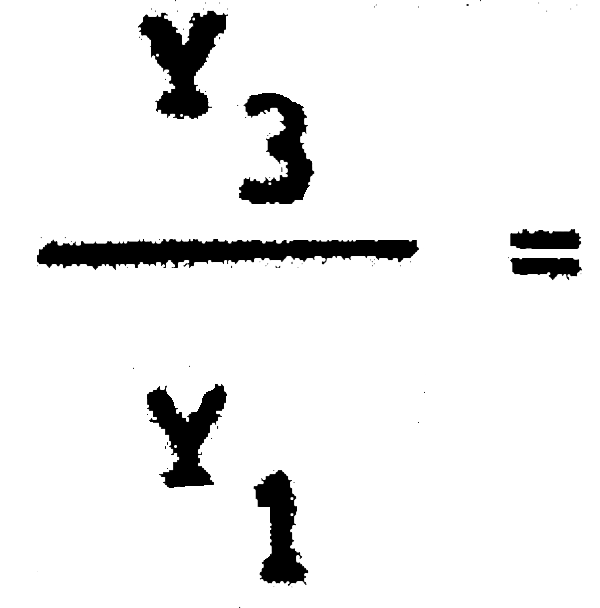
(E = 0,25) = 0,92 Captura a la larga = 92% de la de equilibrio con 40 mm.
(E = 0,50) = 1,26 Captura a la larga = 126% de la de equilibrio con 40 mm.
(E = 0,75) = 1,73 Captura a la larga = 173% de la de equilibrio con 40 mm.
3. Cambio de malla 1 (39,2 mm) a malla 4 (67,1 mm).
C1= 25389600 C4= 5247500 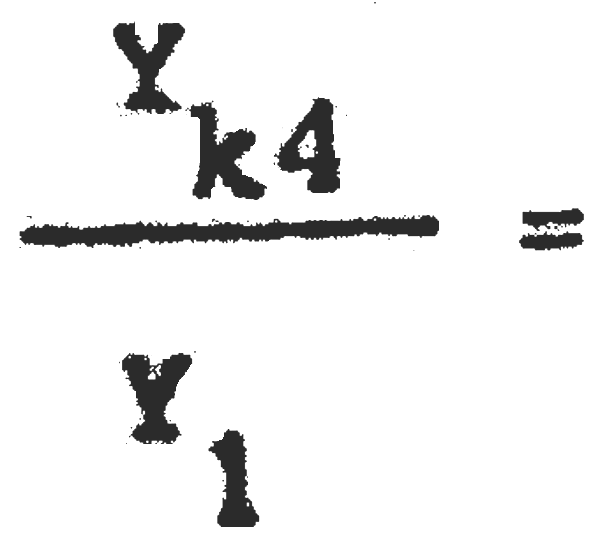 = 0,59 Pérdida inmediata del 41% de la captura actual.
= 0,59 Pérdida inmediata del 41% de la captura actual.
Y1= 2288703 Yk4 = 1350268
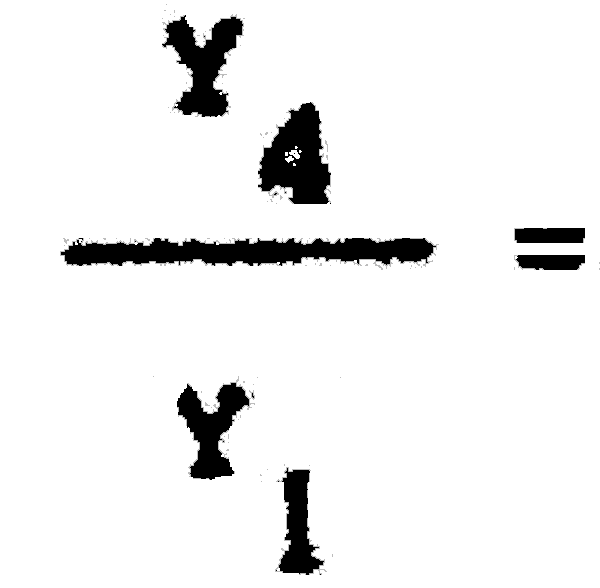
(E = 0,25) = 0,87 Captura a la larga = 87% de la de equilibrio con 40 mm.
(E = 0,50) = 1,30 Captura a la larga = 130% de la de equilibrio con 40 mm.
(E = 0,75) = 1,92 Captura a la larga = 192% de la de equilibrio con 40 mm.
Tabla II.-Resultados de la evaluación de un cambio de malla de 40 mm a las de 50, 59,7 y 67,1 mm. respectivamente. C1 = Captura actual en no de ejemplares (40 mm.). C2, C3 Y C4 = Capturas en no de ejemplares con las mallas de 50, 59,7 y 67,1 mm. respectivamente. Y1 = Peso de la captura actual en Kgr. Yk2, Yk3, Yk4= Pesos de las capturas inmediatamente después del cambio a las distintas mallas. Y2, Y3, Y4 = Pesos correspondientes a largo plazo.
| Talla | C1 | P1 | P2 | P3 | P4 | |
|---|---|---|---|---|---|---|
| LT.cm. | № ej/1000 | gr | 39,2 mm | 50 mm | 59,7 mm | 67,1 mm |
| 5 | 28 | 0,66 | 0,02 | 0,01 | 0,02 | |
| 5.5 | 67 | 0,87 | 0,02 | (0,01) | 0,01 | 0,03 |
| 6 | 80 | 1,13 | 0,04 | (0,01) | 0,02 | 0,04 |
| 6.5 | 42 | 1,43 | 0,06 | 0,02 | 0,02 | 0,05 |
| 7 | 354 | 1,78 | 0,09 | 0,02 | 0,03 | 0,05 |
| 7.5 | 1133 | 2,18 | 0,13 | 0,04 | 0,04 | 0,06 |
| 8 | 3656 | 2,65 | 0,19 | 0,06 | 0,05 | 0,08 |
| 8.5 | 9262 | 3,17 | 0,26 | 0,09 | 0,07 | 0,19 |
| 9 | 20070 | 3,76 | 0,35 | 0,13 | 0,09 | 0,11 |
| 9.5 | 30171 | 4,43 | 0,45 | 0,19 | 0,13 | 0,13 |
| 10 | 44704 | 5,16 | 0,56 | 0,26 | 0,16 | 0,15 |
| 10.5 | 63740 | 5.98 | 0,66 | 0,35 | 0,21 | 0,17 |
| 11 | 83682 | 6,88 | 0,74 | 0,45 | 0,27 | 0,20 |
| 11.5 | 75157 | 7,86 | 0,82 | 0,56 | 0,34 | 0,23 |
| 12 | 68168 | 8,94 | 0,87 | 0,66 | 0,41 | 0,27 |
| 12.5 | 51898 | 10,11 | 0,91 | 0,74 | 0,49 | 0,31 |
| 13 | 39505 | 11,38 | 0,94 | 0,82 | 0,57 | 0,35 |
| 13.5 | 21435 | 12,76 | 0,96 | 0,87 | 0,64 | 0,39 |
| 14 | 10240 | 14,24 | 0,97 | 0,91 | 0,71 | 0,43 |
| 14.5 | 4317 | 15,84 | 0,98 | 0,94 | 0,77 | 0,48 |
| 15 | 2465 | 17,55 | 0,99 | 0,96 | 0,82 | 0,53 |
| 15.5 | 1021 | 19,39 | 0,99 | 0,97 | 0,86 | 0,57 |
| 16 | 524 | 21,35 | 0,99 | 0,98 | 0,90 | 0,62 |
| 16.5 | 280 | 23,43 | 1,00 | 0,99 | 0,92 | 0,66 |
| 17 | 98 | 25,66 | 0,99 | 0,94 | 0,70 | |
| 17.5 | 22 | 28,02 | 0,99 | 0,96 | 0,74 |
Tabla III.- Datos de base para la evaluación de los efectos de un cambio de
malla. C1 = captura actual (mall de 40 mm.) correspondiente al
período Enero-Julio 1982. = peso por ejemplar en el punto me
dio del intervalo de tallas. P1, P2, P3, P4 = Tantos por uno de
retención para las mallas de 39,2, 50, 59,7 y 67,1 mm. respecti
vamente. Estas proporciones han sido obtenidas a partir de datos
reales de campaña de selectividad (ELID 8005, IBN SINA 8105 y
IBN SINA 8102). = peso medio por ejemplar. Relación talla-peso
utilizada W(gr)= 0.00398 L3.08 (cm)
1. Cambio de malla 1 (39,2 mm) a malla 2 (50 mm).
C1 = 531114000 Ck2 = 347306000 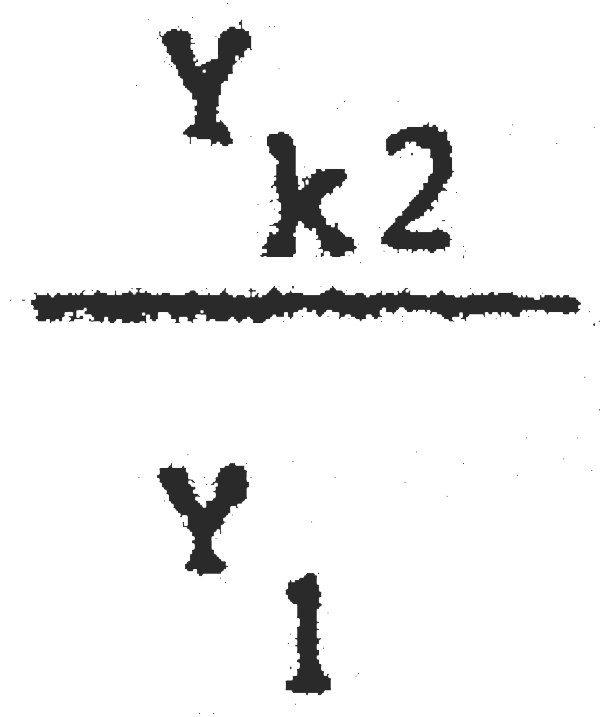 = 0,71. Pérdida inmediata del 29% en peso.
= 0,71. Pérdida inmediata del 29% en peso.
Y1 = 4185159 Yk2= 2972394
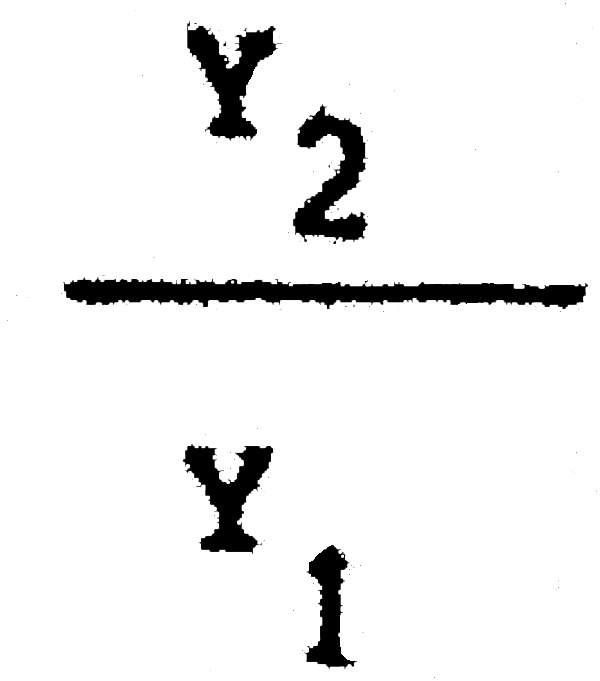
(E = 0,25) = 0,79. A la larga la captura será el 79% de la actual.
(E = 0,50) = 0,88. A la larga la captura será el 88% de la actual.
(E = 0,75) = 0,98. A la larga la captura será el 79% de la actual.
2. Cambio de malla 1 (39,2 mm) a malla 3 (59,7 mm).
C1 = 531114000 Ck3 = 225482000
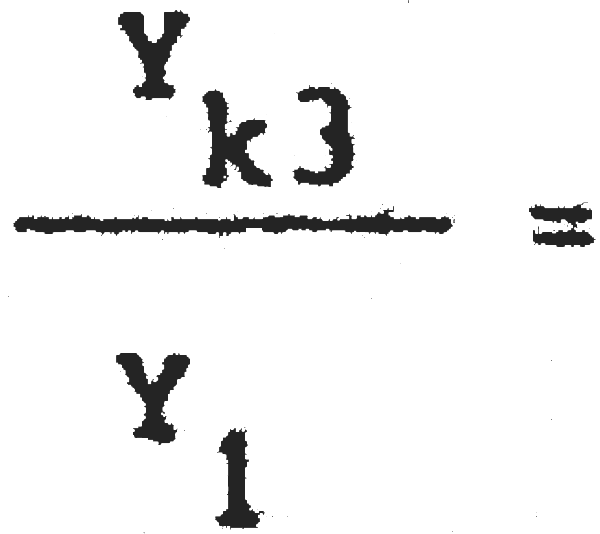 = 0,47. Pérdida inmediata del 53% en peso.
= 0,47. Pérdida inmediata del 53% en peso.
Y1 = 4185159 Yk3 = 1968254
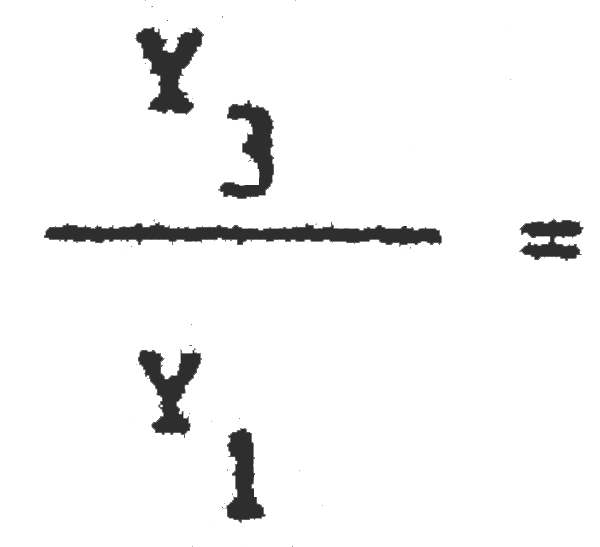
(E = 0,25) = 0,58. A la larga la captura será el 58% de la actual.
(E = 0,50) = 0,72. A la larga la captura será el 72% de la actual.
(E = 0,75) = 0,89. A la larga la captura será el 89% de la actual.
3. Cambio de malla 1(39,2 mm) a malla 4 (67,1 mm).
C1 = 531114000 Ck4 = 167805000
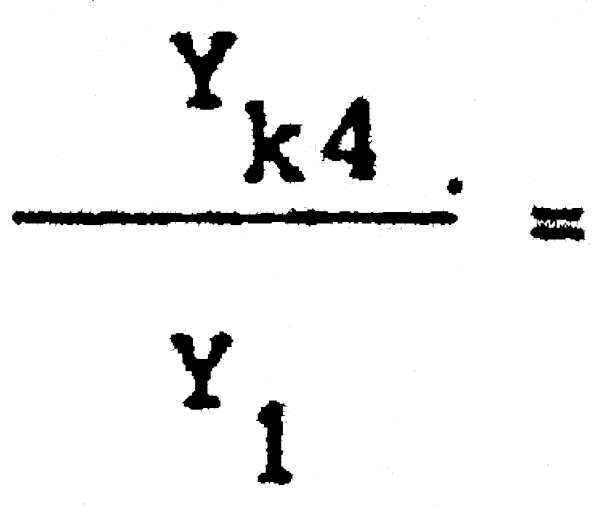 = 0,33. Pérdida inmediata del 67% en peso.
= 0,33. Pérdida inmediata del 67% en peso.
Y1 = 4185159 Yk4 = 1363498
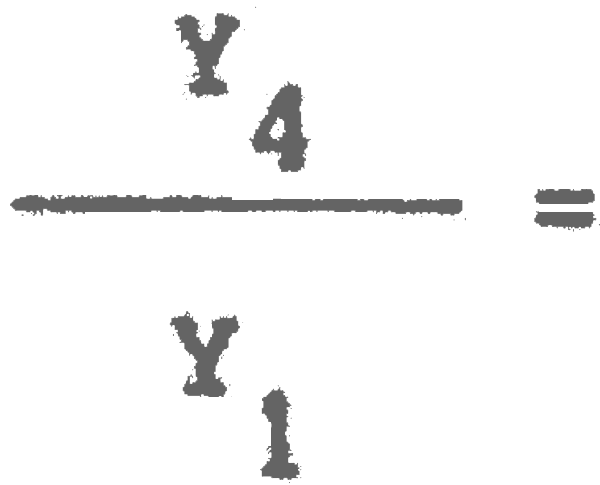
(E = 0,25) = 0,44. A la larga la captura será el 44% de la actual.
=(E = 0,50) = 0,59. A la larga la captura será el 59% de la actual.
(E = 0,75) = 0,79. A la larga la captura será el 79% de la actual.
Tabla IV.- Resultados de la evaluacion de los efectos del cambio de la malla de 40 mm. a las de 50, 59,7 y 67,1 mm. C1 = captura actual (malla de 40 mm.) en no ej. × 10-3; C2, C3 y C4 = captura inmediata con las mallas de 50, 59,7 y 67,1 mm respectivamente; Y1 = captura actual (malla de 40 mm.) en Kgs.; Yk2.; Yk3 e Yk4= peso de la captura inmediatamente después del cambio a las mallas de 50, 59,7 y 67,1 mm. respectivamente ; Y2, Y3 e Y4= peso de las capturas esperadas a largo plazo con las mallas de 50, 59,7 y 67,1 mm. respectivamente.
EVALUACION DEL EFECTO ECONOMICO DE UN CAMBIO DE MALLA EN LA PESQUERIA MERLUZA GAMBA CONSIDERANDO UNA CAPTURA DE 100 Kgr. DE AMBAS ESPECIES.
| Precios (1982) | Porcentajes relativos de las dos especies (1981–1982) | |||
|---|---|---|---|---|
| Merluza = 418 P./Kgr. | Merluza = 35,6% en peso. | 35,6 Kgr merluza - 14880,8 P, | ||
| 100 Kgr | 6210,4 P, | |||
| Gamba = 734 P,/Kgr. | Gamba = 64,4% en peso. | 64,4 Kgr gamba - 47269,6 P, | ||
| MALLAS | 50 mm. | 59,7 mm. | 67.1 mm. | |||
|---|---|---|---|---|---|---|
| Especies | Merluza | Gamba | Merluza | Gamba | Merluza | Gamba |
| Pérdidas | ||||||
| Pérdida por especie | 14% | 29% | 33% | 53% | 41% | 67% |
| (‰) | ||||||
| Pérdida por especie | ||||||
| (P.) | 2083,3 | 13708,2 | 4910,7 | 25052,9 | 6101,1 | 31670,6 |
| Pérdida total | ||||||
| P. | 15791,5 | 29963,6 | 37771,7 | |||
| Pérdida total | ||||||
| ‰ | 25,4% | 48,2% | 60,8% | |||
6. BIBLIOGRAFIA
Alverson, O.L., and Carney, M.J., 1975 A graphic review of the grow and decay of population cohorts. J.Cons.int.Explor.Mer. 36(2) : 133–143
Andreu, B., 1955 The sexuality of sardines, Proc.Tech.Pap.gen.Fish.Coun.Mediterr. 3 : 45–60
Belvèze, M., 1972 Le facteur de condition de la sardine et son évolution à Safi et Agadir. Bull.Inst.Pêches.Marit. du Maroc, No. 20
Belvèze, M., 1974 L'état des stocks de la sardine marocaine à la fin de l'année Trav.Doc.Rev.Pêches.Maroc, 6 : 22
Belvèze, M., et Rami, M., 1978 Détermination de l'âge de la sardine marocaine (Sardina pilchardus Walb.). Eléments de croissance comparés. Bull.Inst. Pêches Marit.Maroc. 23 : 57–81
Bravo de Laguna, J., Fernandez, M.A.R. and Santana, J.C., 1976 The Spanish Fishery on Sardine (Sardina pilchardus Walb.) off West Africa. ICES, C.M. 1976/J : 13
Bravo de Laguna, J., Fernandes, M.A.R., y Delgado, A., 1977 Problematica de las investigaciones sobre la sardina (Sardina pilchardus Walb.) del Banco Pesquero Sahariano. Com. I mesa redonda sobre Dinamica de Poblaciones Barcelona
Delgado, A., Fernandez, M.A.R., y Goni, R., 1980 Contribucion al estudio de la sardina (Sardina pilchardus Walb.) en aguas de Africa Occidental. III Estudio del crecimiento por lectura de otolitos y por retrocalculo mediante escalimetria de escamas. Bol.Inst.Espa.Oceano.T.IV.
Domanevski, L.N., et Barkova, N.A., 1981 Etat du stock de sardine Sardina pilchardus Walb. dans la région de l'Afrique du Nord-Ouest. COPACE/TECH/81/131 : 19–30
Fernandez, M.A.R., Delgado, A., y Bravo de Laguna, J., 1979 Contribucion al estudio de la Sardina (Sardina pilchardus Walb.) en aguas de Africa Occidental. II Estimacion de la abundancia de la poblacion explotada por la flota espanola. Bol.Inst.Espa. Oceano. IV, 260 : 45–61
Furnestin, J., 1950 Premières observations sur la biologie de la sardine marocaine. Rapp. p.V.Rem.CIEM, 126 : 57–61.
Furnestin, J. 1959 et Furnestin, M.L., La reproduction de la Sardine et de l'Anchois des côtes atlantiques du Maroc. Rev.Trav.Inst. Pêches Marit. 23–1
Furnestin, J., et Furnestin, M.L. 1967 La Sardine marocaine et sa Pêche. Migrations trophiques et génétiques en relation avec l'hydrologie et le placton Rapp.Proc.Verb.CIEM. 159
Gomez Larrañeta, M., 1977 Natural mortality in a Mediterranean sardine population ICES, C.M., 1977/J : 8
Holden, M.J., et Raitt, D.F.S., 1974 Manuel de science halieutique. Doc.Tech.FAO. Pêches (115). Rev. 1 : 223
Holzlöhner, S., 1975 On the recent stock development of Sardina pilchardus Walb. off Spanish Sahara. ICES, C.M./J : 13
Howard, G. y Landa, A., Estudo de la edad, el crecimiento, la madurez sexual y el desove de la anchoveta (Cetengraulis mysticetus) en el golfo de Panama.
Krzeptowski, M., 1975 Sardine in Polish catches in the Spanish Sahara Region during the years 1971–1974. ICES.C.M./1975/J : 18
Macer, C.T. 1974 Some observations on the fecundity of the pilchard (Sardina pilchardus Walb) off the South-west coast of England. ICES, C.M. 1974/J : 9
Rikhter, C., and Efanov, V., 1976 On one of the approaches to estimation of natural mortality of fish populations. ICNAF, Res.Doc. 76/8
Saccardo, A.A., (comp), 1983 Biologia y disponibilidad de Sardina (Sardinella brasiliensis, Steindachner, 1897) en la costa Sudeste del Brasil Comunicacion Consulta recursos de peces neriticos. Costa Rica, 1983
Taylor, C.C., 1959 Temperature and Growth. The Pacific Razor Clam. Jour.Cons. 25 (1).
BIBLIOGRAPHIE CHOISIE CONCERNANT LES MERLUS ET LES CREVETTES PROFONDES
DANS LA REGION DU COPACE
AtlantNIRO, 1978 Les merlus (Merluccius spp.) de la côte Nord-Ouest de l'Afrique Rome, FAO, CECAF/ECAF Series 78/9 : 43–63
Belvèze, J. et J. Bravo de Laguna, 1980 Les ressources halieutiques de l'Atlantique contre—Est. Deuxième partie : les ressources de la côte ouest afriscaine entre 24°N et le Détrioit de Gibralar. FAO.Doc.Tech.Pêches, (186.2) : 64p.
Belvèze, H. et J. Bravo de Laguna y R. Goni, 1981 Selectividad de arte de arrastre sobre Merluza (Merluccius merluccius) y Gamba blanca (Parapenaeus longirostris) en la zona norte de CECAF. FAO.Fish.Rep., (244) : 110–127.
Bravo de Laguna, J., 1981 Informe de los trabajos realizados en la campaña IBN SINA 8112. Doc.Int.C.C. Canarias IEO
Bravo de Laguna, J., 1982 Distribution and abundance of demersal resources of the CINECA region. Rapp.P.-v. Réun.Cons.int.Explor.Mer., 180 : 432–446
Brethes, J.C., 1978 Estimation de la biomasse de M. merluccius entre le Cap Spartel (36°N) et le Cap Juby (28°N), d'après les campagnes du chalutier scientifique “El Idrissi”. Rome, FAO, CECAF/ECAF Series 78/9 : 35–42
Cervantes, A., y R. Goni, 1983 Descripcion de las pesquerias espanolas de merluza y crustaceos de Africa Occidental al norte de Cabo Blanco. Comunicacion presentada al Simposio international sobre las areas de afloramiento mas importantes del oeste Africano, Barcelona, Noviembre, 1983 : 24p.
Chaali, A., 1982 Approche analytique de l'état d'exploitation du stock de merlu (Merluccius merluccius) atlantique du Maroc. Manuscrit, ISPM, Casablanca
Christy, F.T., 1979 Economic benefits and arrangements with foreign fishing countries in the northern sub-region of CECAF : a preliminary assessment. Rome, FAO, CECAF/ECAF Series 79/19 : 39p.
Collingnon, J., 1969 Première note sur le peuplement en poissons benthiques du plateau continental atlantique marocain. Bull.Inst.Pêch.Marit.Maroc, 17 : 11–44
Collingnon, J., 1971 Les poissons benthiques du plateau continental marocain, 2ème note : le peuplement des sables côtiers. Bull.Inst.Pêche.Marit.Maroc 18 : 11–35.
Domanevsky, L.N., 1970 Biology and distribution of the main commercial fishes and peculiarities of their fishing by trawl on the shelf from Cape Spartel to Cape Verde. Rapp.P.-v. Réun.CIEM, 159 : 223–6
FAO, 1973 Fishery Committee for the Eastern Central Atlantic (CECAF). Working Party on Resource Evaluation. Report of the first session of the Working Party on Resource Evaluation. Rome, 14–20 April 1972. FAO.Fish.Rep., (136) : 69p.
FAO 1975 Fishery Committee for the Eastern Central Atlantic (CECAF). Working Party on Resource Evaluation. Report of the Second Session of the Working Party on Resource Evaluation. Rome, 3–6 December 1973. FAO.Fish.Rep., (158) : 91p.
FAO, 1976 Fishery Committee for the Eastern Central Atlantic (CECAF). Report of Third session of the Wroking Party on Resource Evaluation. Rome, 9–13 February 1976. FAO.Fish.Rep., (183) : 51p.
FAO 1978b Report of the ad hoc Working group on hakes (Merluccius merluccius Linn aeus) Merluccius senegalensis (Cadenat) and Merluccius cadenati (Doutre) in the northern zone of CECAF. Rome, FAO, CECAF/ECAF Series 78/9 : 93 p.
FAO 1979a Fishery Committee for the Eastern Central Atlantic (CECAF). Report of the Fourth Session of the Working Party on Resource Evaluation. Dakar, 23–27 April 1979. FAO.Fish.Rep. (220) : 200 p.
FAO 1981b Fishery Committee for the Eastern Central Atlantic (CECAF). Report of the Fifth Session of the Working Party on Resource Evaluation. Dakar, 15–17 October 1980. FAO.Fish.Rep., (244) : 127p.
FAO 1983 Fishery Committee for the Eastern Central Atlantic (CECAF). Report of the Sixth Session of the Working Party on Resource Evaluation. Dakar, 2–6 February 1982. FAO.Fish.Rep, (264) : 147 p.
Goñi, R., 1983a Growth studies on European hake (Merluccius merluccius L.) from the Northwest African shelf. ICES CM 1983/G : 10 : p. 16 (mimeo)
Goñi, R., 1983b Selectividad de la merluza europea (Merluccius merluccius Linnaeus, 1758), con artes de arrastre de poliamida en las pesqueria de Africa Occidental, al norte de Cabo Blanco.
Goñi, R., 1983c Consideraciones sobre los efectos de un cambio de malla en la pesquerias de gamba blanca (Parapenaeus Pongirostris) del Atlantico Centro-Oriental. FAO.Fish.Rep., (264) : 58–62
Greboval, D., 1982 Analyse bioéconomique des principales pêcheries dans la région Nord du COPACE. CECAF/TECH/82/45: 118p
Griffin, W.L., J.P. Warren and W.E. Grant, 1979 A bioeconomic model for fish stock management : the cephalopod fishery of Northwest Africa. Dakar, CECAF Project, CECAF/TECH/79/16 : 42p.
Gulland, J.A., 1979a Problems of managing the CECAF hake and seabream stocks. CECAF/ECAF Series 79/13, Rome, FAO, 7p.
Maurin, C., 1954 Les merlus du Maroc et leur pêche. Bull.Inst.Pêche.Marit.Maroc 2 : 38 p.
Maurin, C. 1968 Ecologie ichthyologique des fonds chalutables atlantiques (de la baie Ibero-marocaine à la Mauritanie) et de la Méditerranée occidentale Rev.Trav.Inst.Pêch.Marit., 31 (1) : 47p.
Poinsard, F., et L. Villegas, 1975 Analyse de la pêche côtière au chalut dans l'Atlantique marocain. Trav.Doc.Dév.Pêch.Marit., Casablanca (12) : 37p
RESUMEN DE LOS DATOS DE BASE Y PARAMETROS BIOLOGICOS DE LA PESQUERIA DE GAMBA
EN LA DIVISION 34.1.1. DE CECAF.
par
A. Cervantes et R. Goni
| PUERTO | № MEDIO | № MEDIO | CARACTERISTICAS MEDIA | CALADEROS | PROFUNDIDAD | DP/MAREA | ‰CAPTURA | ESPECIES OBJECTIVOS | ||
|---|---|---|---|---|---|---|---|---|---|---|
| DESCARGA | BARCOS | MAREAS/MES | TRB | HP | ESL.(m) | PREFERIDOS | PESCA(m) | TOTAL ANUAL | (‰RELATIVO) | |
| HUELVA | 106 | 123 | 97 | 318 | 23 | 1oCasablanca | 100–600 | 11 | 35 | Gamba blanca (68%) |
| 2o Tafelneh-C.Sim | Otros crust.*(18%) | |||||||||
| 3o Cabo Ghir | Merluza Europ.(14%) | |||||||||
| 4o Kenitra | ||||||||||
| PTO.STA.MARIA | 57 | 97 | 70 | 213 | 18 | 1o Kenitra | 100–250 | 7 | 20 | Gamba blanca (59%) |
| 2o Casablanca | Merluza europ.(37%) | |||||||||
| 3oArcila-Larache | Otros crust. * (4%) | |||||||||
| ALGECIRAS | 65 | 106 | 109 | 413 | 21 | 1o Arcila-Larache | 100–500 | 9 | 30 | Gamba blanca (63%) |
| 2o Kenitra | Merluza Europ.(33%) | |||||||||
| 3o Casablanca | Otros crust.*(4%) | |||||||||
| 4o Cabo Ghir | ||||||||||
| MALAGA | 52 | 41 | 95 | 373 | 20 | 1o Arcila-Larache | 100–450 | 8 | 11 | Gamba blanca (53%) |
| 2o Kenitra | Merluza europ.(46%) | |||||||||
| 3o Casablanca | Otros crust.*(4%) | |||||||||
| CADIZ | 6 | 7 | 188 | 543 | 27 | De Cabo Cantin | 300–700 | 16 | 4 | Merluza europ.(61%) |
| a Cabo Noun | Otros crust. | |||||||||
* Carabinera alistado, moruno y cigala)
Tabla I - Datos sobre la actividad de la flota española de arrastre al fresco que explota la pesqueria de gamba, con malla 40 mm., en Africa Occidental alnorte de Cabo Blanco. Año 1982.
| PUERTOS | HUELVA | ALGECIRAS | PTO.STA.MARIA | MALAGA | CADIZ | ALMERIA | TOTAL | |
|---|---|---|---|---|---|---|---|---|
| C (t) | 2 721 | 1 472 | 813 | 613 | 13 | 12 | 5 644 | |
| 981 | f (DP) | 14 047 | 11 415 | 8 666 | 4 382 | 1 831 | 181 | 40 522 |
| CPUE | .19 | .13 | .09 | .14 | .01 | .07 | .14 | |
| C (t) | 2 889 | 1 773 | 1 171 | 584 | 16 | 21 | 6 453 | |
| 1982 | f (DP) | 16 293 | 11 616 | 8 574 | 3 941 | 1 404 | 447 | 42 275 |
| CPUE | .18 | .15 | .14 | .15 | .01 | .05 | .15 | |
| ENERO | FEBRERO | MARZO | ABRIL | MAYO | JUNIO | JULIO | AGOSTO | SEPBRE. | OCTUBRE | NOVBRE. | DICBRE. | 1982 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CAPTURA | |||||||||||||
| (KG) | 502 | 513 | 657 | 562 | 636 | 586 | 503 | 530 | 523 | 460 | 396 | 588 | 6455 |
| (DP) | 405 | 351 | 453 | 313 | 365 | 382 | 339 | 357 | 368 | 400 | 360 | 518 | 4611 |
| ESFUERZO (№M) | 3 620 | 3 605 | 4 275 | 2 724 | 3 433 | 3 487 | 3 320 | 3 144 | 3 479 | 3 723 | 3 132 | 4 315 | 42 257 |
| CPUE (C/DP) | 0.139 | 0.142 | 0.154 | 0.206 | 0.185 | 0.168 | 0.151 | 0.169 | 0.150 | 0.123 | 0.126 | 0.136 | 0.153 |
Tabla III :- Captura (t); Esfuerzo (NM = № de mareas; DP = dias pesca) y captura por unidad de esfuerzo (t/DP) de la flota española de arrastre que pesca gamba blanca en la división 34.1.1. de CECAF. Año 1982.
| Lt (cm) | № ej. |
|---|---|
| 3.0 | 0 |
| 3.5 | 0 |
| 4.0 | 0 |
| 4.5 | 0 |
| 5.0 | 27927 |
| 5.5 | 67593 |
| 6.0 | 82656 |
| 6.5 | 42885 |
| 7.0 | 433289 |
| 7.5 | 2118300 |
| 8.0 | 6253032 |
| 8.5 | 19049802 |
| 9.0 | 39196596 |
| 9.5 | 64818598 |
| 10.0 | 102714424 |
| 10.5 | 123469104 |
| 11.0 | 139863175 |
| 11.5 | 121730347 |
| 12.0 | 110349689 |
| 12.5 | 87623861 |
| 13.0 | 63859555 |
| 13.5 | 31205611 |
| 14.0 | 14857158 |
| 14.5 | 6519247 |
| 15.1 | 3651128 |
| 15.5 | 1556405 |
| 16.0 | 755851 |
| 16.5 | 400612 |
| 17.1 | 174141 |
| 17.5 | 45891 |
| 18.0 | 14694 |
| 18.5 | 0 |
| 19.0 | 2235 |
| 19.5 | 0 |
| 20.0 | 0 |
Tabla IV .- Composicion por tallas de la captura de gamba blanca. Division 34.1.1 de CECAF, Ano 1982.
| Lt (cm | %♂ | %♀ | № EJEMPLARES |
|---|---|---|---|
| 7 | - | - | 2 |
| 7.5 | 62.5 | 37.5 | 8 |
| 8 | 60.0 | 40.0 | 25 |
| 8.5 | 65.7 | 34.3 | 67 |
| 9 | 68.9 | 31.1 | 193 |
| 9.5 | 71.9 | 28.1 | 185 |
| 10 | 78.3 | 21.7 | 378 |
| 10.5 | 82.9 | 17.1 | 293 |
| 11 | 69.3 | 30.7 | 433 |
| 11.5 | 47.8 | 52.2 | 251 |
| 12 | 22.5 | 77.5 | 440 |
| 12.5 | 5.2 | 94.8 | 310 |
| 13 | 7.8 | 92.2 | 461 |
| 13.5 | 3.8 | 96.2 | 236 |
| 14 | 4.3 | 95.7 | 351 |
| 14.5 | 1.8 | 98.2 | 163 |
| 15 | 1.1 | 98.9 | 179 |
| 15.5 | - | 100.0 | 76 |
| 16 | 5.1 | 94.9 | 79 |
| 16.5 | - | 100.0 | 32 |
| 17 | - | 100.0 | 20 |
| 17.5 | - | 100.0 | 12 |
| 18 | - | 100.0 | 7 |
| 18.5 | - | 100.0 | 2 |
| 19 | - | 100.0 | 2 |
| TOTAL No | 1 152 | 3 053 | 4 205 |
| % | 27% | 73% |
Tabla V :-Relación de sexos por intervalo de talla de gamba blanca en la división 34.1.1. de CECAF. Año 1982.
| Relación talla total-peso | № ej. | r | rango (cm) | |
|---|---|---|---|---|
| ♂ | W(g) = 0.00942 . LT (cm)2.65 565 | 561 | 0.85 | 8.5 – 14.0 |
| ♀ | W(g) = 0.00467 . LT (cm)2.9893 | 1 119 | 0.97 | 8.5 – 19.0 |
Tabla VI .- Relación longitud total-peso de gamba blanca en la división 34.1.1. de COPACE. Año 1982.
| Relación talla caparazón-peso | № ej. | r | rango (cm) | |
|---|---|---|---|---|
| ♂ | W(g) = 0.00561 . Lc (mm)2.20453 | 599 | 0.74 | 16 – 45 |
| ♀ | W(g) = 0.00344 . Lc (mm)2.41686 | 1 129 | 0.90 | 15 – 30 |
Tabla VII .- Relación longitud del caparazón-peso de gamba blanca en la división 34.1.1. de COPACE. Año 1982.
| Relación talla total-talla | № ej. | r | rango LT (mm) | |
|---|---|---|---|---|
| caparazón | ||||
| ♂ | Lo (mm) = 0.27823 . LT (mm)0.93047 | 1 475 | 0.79 | 70 – 165 |
| ♀ | Lo (mm) = 0.13953 . LT (mm)1.08325 | 2 370 | 0.90 | 76 – 194 |
Tabla VIII .- Relación talla total-talla del caparazón de gamba blanca de la división 34.1.1. de COPACE. Año 1982.
| Talla | C1 | P1 | P2 | P3 | P4 | |
|---|---|---|---|---|---|---|
| LT.cm. | № ej/1000 | gr | 39,2 mm | 50 mm | 59,7 mm | 67,1 mm |
| 5 | 28 | 0,66 | 0,02 | 0,01 | 0,02 | |
| 5.5 | 67 | 0,87 | 0,02 | (0,01) | 0,01 | 0,03 |
| 6 | 80 | 1,13 | 0,04 | (0,01) | 0,02 | 0,04 |
| 6.5 | 42 | 1,43 | 0,06 | 0,02 | 0,02 | 0,05 |
| 7 | 354 | 1,78 | 0,09 | 0,02 | 0,03 | 0,05 |
| 7.5 | 1133 | 2,18 | 0,13 | 0,04 | 0,04 | 0,06 |
| 8 | 3656 | 2,65 | 0,19 | 0,00 | 0,05 | 0,08 |
| 8.5 | 9262 | 3,17 | 0,26 | 0,09 | 0,07 | 0,19 |
| 9 | 20070 | 3,76 | 0,35 | 0,13 | 0,09 | 0,11 |
| 9.5 | 30171 | 4,43 | 0,45 | 0,19 | 0,13 | 0,13 |
| 10 | 44704 | 5,16 | 0,56 | 0,26 | 0,16 | 0,15 |
| 10.5 | 63740 | 5.98 | 0,66 | 0,35 | 0,21 | 0,17 |
| 11 | 83682 | 6,88 | 0,74 | 0,45 | 0,27 | 0,20 |
| 11.5 | 75157 | 7,86 | 0,82 | 0,56 | 0,34 | 0,23 |
| 12 | 68168 | 8,94 | 0,87 | 0,66 | 0,41 | 0,27 |
| 12.5 | 51898 | 10,11 | 0,91 | 0,74 | 0,49 | 0,31 |
| 13 | 39505 | 11,38 | 0,94 | 0,82 | 0,57 | 0,35 |
| 13.5 | 21435 | 12,76 | 0,96 | 0,87 | 0,64 | 0,39 |
| 14 | 10240 | 14,24 | 0,97 | 0,91 | 0,71 | 0,43 |
| 14.5 | 4317 | 15,84 | 0,98 | 0,94 | 0,77 | 0,48 |
| 15 | 2465 | 17,55 | 0,99 | 0,96 | 0,82 | 0,53 |
| 15.5 | 1021 | 19,39 | 0,99 | 0,97 | 0,86 | 0,57 |
| 16 | 524 | 21,35 | 0,99 | 0,98 | 0,90 | 0,62 |
| 16.5 | 280 | 23,43 | 1,00 | 0,99 | 0,92 | 0,66 |
| 17 | 98 | 25,66 | 0,99 | 0,94 | 0,70 | |
| 17.5 | 22 | 28,02 | 0,99 | 0,96 | 0,74 |
Tabla IX: Datos de base para la evaluación de los efectos de un cambio
de malla. C1 = captura actual (malla de 40 mm) correspondiente
al periodo Enero-Julio 1982. = peso por ejemplar en el punto
medio del intervalo de tallas. P1, P2, P3, P4 = retención para
las mallas de 39.2, 50, 59.7 y 67,1 respectivamente. Estas
proporciones han sido obtenidas a partir de datos de la campañ a
de selectividad (ELID 8005, IBN SINA 8105 y IBN SINA 8102).
Relación talla-peso utilisada W(gr) = 0.0398 L 3.08 (cm)
RESUMEN DE LOS DATOS DE BASE Y PARAMETROS BIOLOGICOS DE LA
PESQUERIA DE MERLUZA EUROPEA EN LA DIVISION 34.1.1 DE CECAF
par
A. Cervantes et R. Goni
| ESPECIES | MODALITES | |||||
|---|---|---|---|---|---|---|
| MODALITIES | TRAWL 40 m/m | TRIO 60 m/m | GILLNET | LONG-LINE | T O T A L | |
| MODALIDADES | ARRASTRE 40 m/m | TRIO 60 m/m | VOLANTA | PALANGRE | TOTAL | |
| Merluccius merluccius | 2 972 | 1 118 | 1 093 | 95 | 5 278 | |
| P. longirostris | 6 455 | - | - | - | 6 455 | |
| M. senegalensis | - | 63 | 373 | - | 436 | |
| Crustaceos varios (1) | 1 150 | - | - | - | 1 150 | |
| Cefalópodos varios (2) | 813 | - | - | - | 813 | |
| Trachurus trachurus | 691 | - | - | - | 691 | |
| Lophius piscatorius | 557 | - | - | - | 557 | |
| Micromesistius poutassou | 540 | - | - | - | 540 | |
| Dentex macrophtalumus | - | 659 | 75 | - | 734 | |
| Argyrosomus regius | - | 229 | - | - | 229 | |
| Zeus faber | 60 | 201 | - | - | 261 | |
| Pagellus spp. | 356 | 172 | 120 | - | 684 | |
| Mullus barbatus | 50 | 115 | - | - | 165 | |
| Varios | 2 652 | 319 | 220 | 41 | 3 232 | |
| TOTAL | 16 296 | 2 876 | 1 881 | 136 | 21 189 | |
(1) Nephrops norvegicus + Plesiopenaeus edwardsianus + Aristeus antennatus + Aristeomorphafoliacea.
(2) Pulpo + choco + calamar + volador + pota.
Tabla I .- Captura española (en miles de t por especie y tipo de flota, en 1982, en la división 34.1.1. de CECAF.
| MOD. | TOTAL | ||||
|---|---|---|---|---|---|
| ANNEE YEAR AÑO | GILLNET | LONG-LINE | ARR. 40 | (TRIO + BOU) ARR.60 | TOTAL |
| VOLANTA | PALANGRE | ARR.40 (1) | (TRIO + BOU) ARR.60 | TOTAL | |
| 1970 | n.d. | n.d. | 1 714 | 4 916 | n.d. |
| 1971 | " | " | 1 455 | 4 616 | " |
| 1972 | " | " | 1 356 | 4 515 | " |
| 1973 | " | " | 1 426 | 3 576 | " |
| 1974 | " | " | 2 421 | 3 657 | " |
| 1975 | 1 294 | " | 1 796 | 4 620 | " |
| 1976 | 1 082 | " | 1 142 | 3 840 | " |
| 1977 | 764 | " | 670 | 3 641 | " |
| 1978 | 680 | " | 1 155 | 1 732 | " |
| 1979 | 364 | " | 1 035 | 1 552 | " |
| 1980 | 742 | " | 957 | 1 435 | " |
| 1981 | 832 | " | 3 947 | 735 | 5 514* |
| 1982 | 1 093 | 95 | 2 972 | 1 173 | 5 333 |
Tabla II .- Capturas españolas de merluza europea en la división 34.1.1. de COPACE.
| ESFUERZO (DP) | CPUE (t/DP) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| MOD. | |||||||||
| ANNEE YEAR AÑ | GILLNET | LONG-LINE | ARR.40 (1) | ARR.60 | TOTAL | GILLNET | LONG-LINE | ARR. 40 (2) | ARR. 60 |
| (TRIO + BOU) | (TRIO + BOU) | ||||||||
| VOLANTA | PALANGRE | ARR. 40 (1) | ARR.60 | TOTAL | VOLANTA | PALANGRE | ARR. 40 (2) | ARR. 60 | |
| (TRIO + BOU) | (TRIO + BOU) | ||||||||
| 1970 | n.d. | n.d. | 10 416 | n.d. | n.d. | n.d. | n.d. | 0.16 | n.d. |
| 1971 | " | " | 10 024 | " | " | " | " | 0.15 | " |
| 1972 | " | " | 7 540 | " | " | " | " | 0.18 | " |
| 1973 | " | " | 7 042 | " | " | " | " | 0.20 | " |
| 1974 | " | " | 5 040 | " | " | " | " | 0.31 | " |
| 1975 | " | " | 5 670 | " | " | " | " | 0.25 | " |
| 1976 | " | " | 6 412 | " | " | " | " | 0.18 | " |
| 1977 | " | " | 6 860 | 4 905 | " | " | " | 0.10 | 0.74 |
| 1978 | " | " | 4 438 | 2 925 | " | " | " | 0.26 | 0.59 |
| 1979 | 5 729 | " | 4 046 | 3 465 | " | 0.15 | " | 0.26 | 0.45 |
| 1980 | 3 780 | " | 3 710 | 2 205 | " | 0.20 | " | 0.26 | 0.65 |
| 1981 | 2 824 | " | 40 522 | 1 680 | 45 026 | 0.29 | " | 0.97 | 0.44 |
| 1982 | 3 015 | 298 | 47 531 | 2 120 | 52 584 | 0.36 | 0.32 | 0.63 | 0.55 |
(1) Hasta 1981, el esfuerzo corresponde solo a las “bacas” de Cádiz.
(2) Hasta 1981, la CPUE corresponde solo a las “bacas” de Cádiz.
Tabla III.- Esfuerzo y rendimiento español de merluza europea en la división 34.1.1 de COPACE.
| ENERO | FEBRERO | MARZO | ABRIL | MAYO | JUNIO | JULIO | AGOSTO | SEPBRE | OCTUBRE | NOVBRE | DICBRE | 1982 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CPUE (t/DP) ARR.60 | 0.52 | 0.59 | 0.51 | 0.60 | 0.84 | 0.80 | 0.51 | 0.71 | 0.45 | 0.35 | 0.33 | 0.31 | 0.55 |
| CPUE (t/DP) ARR.40 | 0.07 | 0.09 | 0.08 | 0.07 | 0.07 | 0.07 | 0.07 | 0.09 | 0.08 | 0.06 | 0.06 | 0.05 | 0.07 |
| CPUE (t/DP) VOLANTA | - | 0.36 | 0.37 | 0.25 | 0.46 | 0.39 | 0.36 | 0.44 | 0.32 | 0.42 | 0.33 | 0.18 | 0.36 |
| CUPE (t/DP) PALANGRE | - | - | - | - | - | - | - | - | - | 0.27 | 0.32 | 0.37 | 0.32 |
Tabla IV - Evolución mensual de la captura por unidad de esfuerzo (t/DP) de merluza europea capturada por los distintos tipos de flota española en 1982.
a) Arrastre con malla de 60 m m; b) Arrastre con malla de 40 m m (pesqueria de gambas); c) Volanta; d) Palangre
| TALLA | ARR60 | ARR40 | VOLANTA | PALANGRE | TOTAL | TALLA | ARR60 | ARR40 | VOLANTA | PALANGRE | TOTAL |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 5 | 0 | 0 | 0 | 0 | 0 | 54 | 13949 | 12531 | 49185 | 4066 | 79731 |
| 6 | 0 | 0 | 0 | 0 | 0 | 55 | 8013 | 11418 | 46453 | 2785 | 68669 |
| 7 | 0 | 27830 | 0 | 0 | 27830 | 56 | 5002 | 11214 | 48889 | 3381 | 68486 |
| 8 | 0 | 177904 | 0 | 0 | 177904 | 57 | 5990 | 9232 | 38239 | 2575 | 56036 |
| 9 | 0 | 538238 | 0 | 0 | 538238 | 58 | 5865 | 5334 | 36222 | 2848 | 50269 |
| 10 | 0 | 1602180 | 0 | 0 | 1602180 | 59 | 3669 | 3617 | 27789 | 2900 | 37975 |
| 11 | 1682 | 2539760 | 0 | 0 | 2541442 | 60 | 3669 | 3579 | 22971 | 1544 | 31763 |
| 12 | 5045 | 3153560 | 0 | 0 | 3158605 | 61 | 1914 | 2893 | 19053 | 2368 | 26228 |
| 13 | 7572 | 3596000 | 0 | 0 | 3603572 | 62 | 1643 | 3005 | 17251 | 995 | 22894 |
| 14 | 13453 | 3308130 | 0 | 0 | 3321583 | 63 | 1075 | 1182 | 15873 | 472 | 18602 |
| 15 | 21025 | 2548330 | 0 | 0 | 2569355 | 64 | 1800 | 1441 | 14886 | 703 | 18830 |
| 16 | 10935 | 2362988 | 0 | 0 | 2373923 | 65 | 1345 | 584 | 10373 | 1079 | 13381 |
| 17 | 6726 | 2026430 | 0 | 0 | 2033156 | 66 | 406 | 605 | 7281 | 199 | 8491 |
| 18 | 5045 | 1889780 | 0 | 0 | 189825 | 67 | 511 | 611 | 7742 | 242 | 9106 |
| 19 | 5405 | 1767490 | 0 | 0 | 1772895 | 68 | 824 | 882 | 6341 | 256 | 8303 |
| 20 | 1494 | 1718960 | 0 | 0 | 1720454 | 69 | 307 | 583 | 5389 | 155 | 6434 |
| 21 | 3072 | 1563240 | 0 | 0 | 1566312 | 70 | 323 | 106 | 3752 | 54 | 4235 |
| 22 | 5864 | 1350510 | 0 | 0 | 1356374 | 71 | 268 | 360 | 2985 | 3667 | |
| 23 | 11376 | 1379330 | 117 | 0 | 1390823 | 72 | 0 | 108 | 3556 | 33 | 3697 |
| 24 | 28050 | 1158350 | 72 | 0 | 1186472 | 73 | 625 | 212 | 876 | 69 | 1782 |
| 25 | 27683 | 959510 | 94 | 0 | 987287 | 74 | 680 | 0 | 848 | 27 | 1555 |
| 26 | 50886 | 795157 | 229 | 0 | 846272 | 75 | 234 | 101 | 665 | 0 | 1000 |
| 27 | 49874 | 646526 | 221 | 0 | 696621 | 76 | 466 | 13 | 811 | 6 | 1296 |
| 28 | 77101 | 622034 | 264 | 0 | 699399 | 77 | 0 | 0 | 756 | 0 | 756 |
| 29 | 83715 | 487281 | 364 | 0 | 571360 | 78 | 275 | 0 | 857 | 16 | 1148 |
| 30 | 107779 | 386532 | 701 | 0 | 495012 | 79 | 0 | 0 | 467 | 6 | 473 |
| 31 | 112966 | 337122 | 1165 | 0 | 451253 | 80 | 0 | 0 | 280 | 6 | 286 |
| 32 | 140152 | 282680 | 1372 | 267 | 424471 | 81 | 178 | 0 | 240 | 0 | 418 |
| 33 | 140859 | 240459 | 2205 | 223 | 383746 | 82 | 169 | 0 | 93 | 6 | 268 |
| 34 | 151708 | 23637 | 2012 | 202 | 387559 | 83 | 0 | 0 | 191 | 0 | 191 |
| 35 | 134453 | 185720 | 3118 | 1481 | 324772 | 84 | 0 | 215 | 13 | 0 | 228 |
| 36 | 135334 | 172682 | 4736 | 3095 | 3145847 | 85 | 88 | 0 | 55 | 0 | 143 |
| 37 | 147227 | 146797 | 4315 | 2083 | 300422 | 86 | 217 | 0 | 25 | 0 | 242 |
| 38 | 141289 | 144870 | 4319 | 3840 | 294318 | 87 | 178 | 0 | 7 | 0 | 185 |
| 39 | 107952 | 126803 | 7353 | 4137 | 246245 | 88 | 274 | 0 | 0 | 0 | 274 |
| 40 | 125600 | 114616 | 8988 | 4505 | 253709 | 89 | 183 | 0 | 13 | 0 | 196 |
| 41 | 100201 | 102906 | 12036 | 4540 | 219683 | 90 | 403 | 0 | 0 | 0 | 403 |
| 42 | 86733 | 89913 | 20494 | 5564 | 202704 | 91 | 0 | 137 | 0 | 0 | 137 |
| 43 | 87608 | 93845 | 24662 | 3580 | 209695 | 92 | 0 | 0 | 7 | 0 | 7 |
| 44 | 83604 | 66682 | 29248 | 5049 | 184583 | 93 | 0 | 0 | 0 | 0 | 0 |
| 45 | 63139 | 59448 | 39430 | 4688 | 166705 | 94 | 0 | 0 | 0 | 0 | 0 |
| 46 | 63863 | 620216 | 35319 | 4446 | 163844 | 95 | 0 | 0 | 7 | 0 | 7 |
| 47 | 52538 | 56443 | 32657 | 4630 | 146268 | 96 | 0 | 0 | 0 | 0 | 0 |
| 48 | 45169 | 49273 | 48614 | 4952 | 148008 | 97 | 0 | 0 | 0 | 0 | 0 |
| 49 | 33710 | 34092 | 52142 | 2618 | 122562 | 98 | 129 | 0 | 0 | 0 | 129 |
| 50 | 28795 | 34617 | 58459 | 3911 | 125782 | ||||||
| 51 | 24683 | 24578 | 53614 | 4368 | 107243 | ||||||
| 52 | 24816 | 22719 | 55693 | 3916 | 107143 | ||||||
| 53 | 16507 | 16336 | 54376 | 3658 | 90877 |
Tabla V. - Composicion por tallas de la captura de merluza europa realizada por la flota espanola en la division 34.1.1 de CECAF. Ano 1982. ARR 60 = arrastreros con copos de 60 mm ; ARR. 40 arrastreros con copos de 40 mm.
| SIZE | TOTAL CAPTURE |
|---|---|
| LT (cm) | (№) |
| TALLA | CAPTURA TOTAL |
| LT (cm) | (№) |
| 5–9 | 743 972 |
| 10–14 | 14 227 400 |
| 15–19 | 10 644 200 |
| 20–24 | 7 220 430 |
| 25–29 | 3 800 950 |
| 30–34 | 2 142 050 |
| 35–39 | 1 481 610 |
| 40–44 | 1 070 380 |
| 45–49 | 748 889 |
| 50–54 | 510 776 |
| 55–59 | 281 435 |
| 60–64 | 118 316 |
| 65–69 | 45 715 |
| 70–74 | 14 936 |
| 75–79 | 4 673 |
| 80–84 | 1 391 |
| 85–89 | 1 040 |
| 90–94 | 547 |
| 95–99 | 136 |
| TOTAL (№) | 43 058 846 |
Tabla VI - Composición por tallas de la captura española de merluza europea en la división 34.1.1 de CECAF. Año 1982.
| Grupoedad | O | I | II | III | IV | V | VI | VII | VIII | IX | >X | Numero |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| talla (cm) | ||||||||||||
| 5–9 | (100) | |||||||||||
| 10–14 | 63 | 37 | 8 | |||||||||
| 15–19 | 9 | 60 | 22 | 9 | 45 | |||||||
| 20–24 | 2 | 25 | 50 | 20 | 3 | 77 | ||||||
| 25–29 | 3 | 13 | 53 | 29 | 2 | 91 | ||||||
| 30–34 | 2 | 8 | 21 | 45 | 20 | 4 | 112 | |||||
| 35–39 | 1 | 16 | 30 | 36 | 15 | 2 | 115 | |||||
| 40–44 | 6 | 15 | 28 | 27 | 20 | 4 | 79 | |||||
| 45–49 | 4 | 15 | 40 | 27 | 10 | 4 | 55 | |||||
| 50–54 | 2 | 6 | 26 | 37 | 22 | 7 | 58 | |||||
| 55–59 | 6 | 6 | 24 | 14 | 50 | 34 | ||||||
| 60–64 | 6 | 25 | 69 | 16 | ||||||||
| 65–69 | 6 | 94 | 10 | |||||||||
| 70–74 | 100 | 7 | ||||||||||
| 75–79 | 100 | 2 | ||||||||||
| 80–84 | 100 | 3 | ||||||||||
| 85–89 | 100 | 1 | ||||||||||
| 90–94 | 100 | 1 | ||||||||||
| 95–99 | - | - | ||||||||||
| 100–104 | 100 | 1 | ||||||||||
| Total | 715 |
Tabla VII - Clave talla-edad de merluza europea de Africa Noroccidental. Año 1982
GRUPO DE EDAD No EJENPLRES
| O | 10 309 821 | 25.1 |
| I | 13 612 636 | 31.5 |
| II | 6 632 243 | 15.4 |
| III | 5 167 680 | 12.0 |
| IV | 2 923 023 | 6.8 |
| V | 1 480 493 | 3.4 |
| VI | 1 048 170 | 2.4 |
| VII | 651 730 | 1.5 |
| VIII | 804 716 | 0.7 |
| IX | 137 431 | 0.3 |
| >X | 288 051 | 0.8 |
| TOTAL | 43 058 846 | 100 |
Tabla VIII Composición por edad de la captura total española de menluza europea en la división 34.1.1. de CECAF. Añc 1982.
| TRINESTRES | ||||
|---|---|---|---|---|
| % EST. | ||||
| MADUREZ | 1 o | 2 o | 3 o | 4 o |
| I–II | 55 | 41 | 52 | 57 |
| III–IV | 35 | 47 | 28 | 35 |
| V–VI | 6 | 8 | 13 | 6 |
| VII–VIII | 3 | 4 | 7 | 2 |
| o ejemp. | 32 | 75 | 60 | 114 |
Tabala I X.-Porcentajes trimestrales de machos de merluza europea en cada estado de madurez. Individuos de talla ≥33 cm. Area de CECAF. Año 1982.
| TRINESTRES | ||||
|---|---|---|---|---|
| % EST. | ||||
| MADUREZ | 1 o | 2 o | 3 o | 4 o |
| I–II | 32 | 44 | 59 | 37 |
| III–IV | 24 | 48 | 23 | 34 |
| V–VI | 24 | 4 | 13 | 17 |
| VII–VIII | 20 | 4 | 6 | 11 |
| № ejemp. | 25 | 48 | 64 | 169 |
Tabla X.- Porcentajes trimestrales de numbras de merluza européa en cada estado de madurez. Individuos de tallas ≥40 cm. Area de CECAF. Año 1982.
| SIZE | № | ||
|---|---|---|---|
| LT (cm) | %♂ | %♀ | SPECIES |
| TALLA | № | ||
| LT(cm) | % ♂ | % ♀ | EJEMPLARES |
| 15–19 | - | - | 3 |
| 20–24 | 70 | 30 | 76 |
| 25–29 | 43 | 57 | 151 |
| 30–34 | 47 | 53 | 210 |
| 35–39 | 40 | 60 | 243 |
| 40–44 | 42 | 58 | 183 |
| 45–49 | 42 | 58 | 123 |
| 50–54 | 36 | 64 | 96 |
| 55–59 | 17 | 83 | 49 |
| 60–64 | 5 | 95 | 19 |
| 65–69 | 0 | 100 | 6 |
| 70–74 | 0 | 100 | 6 |
| 75–79 | 0 | 100 | 4 |
| 80–84 | 0 | 100 | 1 |
| № | 600 | 570 | 1170 |
| % CAPTURA | 51.2 | 48.8 | - |
Tabla XI .- Relación de sexos por intervalo de talla de merluza europea en el area de CECAF y sex-ratio en la captura española. Año 1982.
| Relación talla-peso vivo: W(g) = | .00737L2.99489 | .00752L2.99342 | .00595L3.05542 |
| F. conversión (vivo-eviscerado:f= | .9387 | .9127 | .9191 |
| Ecuación de crecimiento: Lt = | 100(1-e-.065(t+.82) | 110(1-e-.065(t+.82 | 110(1-e-.064(t+.76 |
Tabla XII .- Relaciones talla-peso vivo; peso vivo-peso eviscerado y ecuaciones de crecimiento por sexos para la merluza europea en la división 34.1.1. de CECAF. (Goñi, 1983; Goñi ycervantes, 1983).
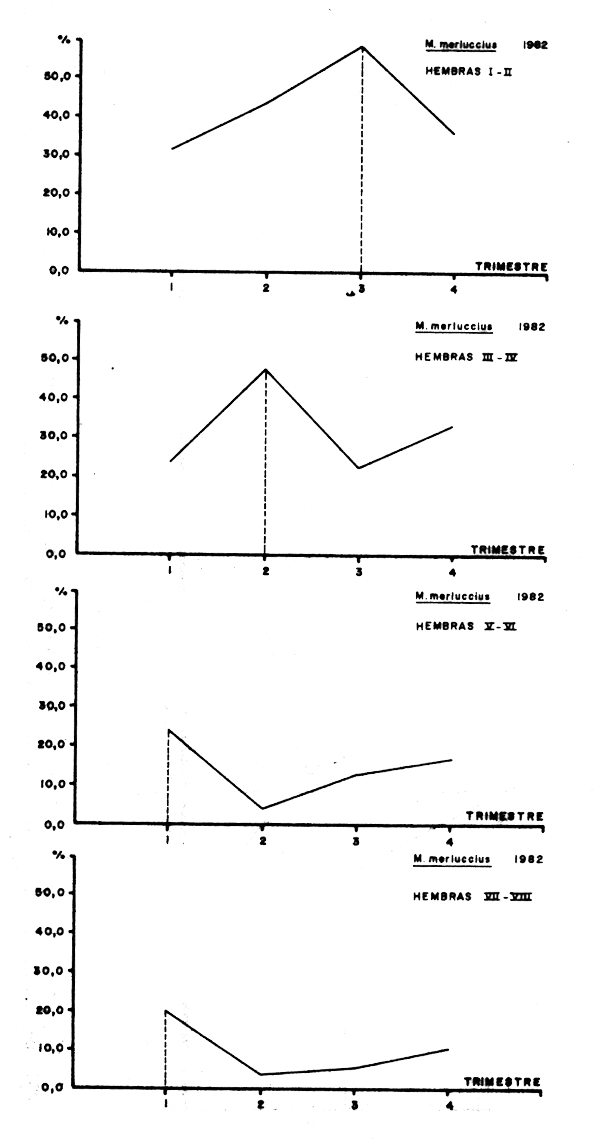
Figura 1 - Evolució anual de los porcentajes trimestrales de cada estado de madurez en hembras de merluza europea. División 34.1.1. de CECAF. Año 1982.
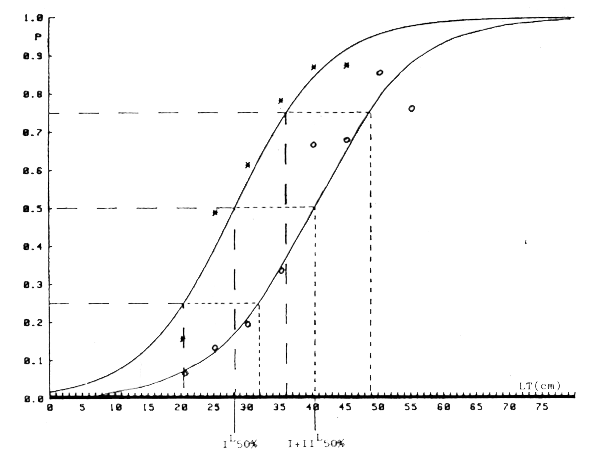
Figura 2.- Ajuste a la logística de la curva de maduración de machos de merluza europea en Africa Moroccidental. Año 1982. Dos supuestos: I - Inmaduros solo estado I. * = valores observados I + II - Inmaduros estados I y II. o = valores observados.
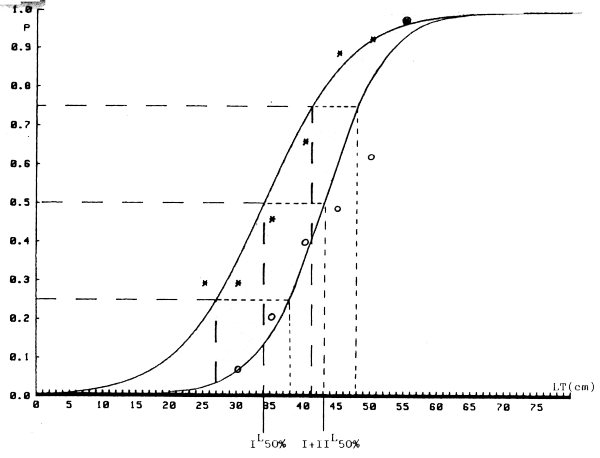
Figura 3.- Ajuste logístico de la curva de maduración de hembras de merluza europea en Africa Moroccidental. año 1982. Dos supuestos: I - Inmaduros solo estado I. * = valores observados. I + II - Inmaduros estados I y II. o = valores observados.
![]()
![]()
![]()