![]()
![]()
![]()
Si analizamos las ecuaciones 2.3 y 2.11, donde:
Nt = No (e-F)t (e-M)t No (e-F+M)t;
encontraremos que para sobrevivir durante cualquier período de tiempo (t), los peces tienen que sobrevivir simultáneamente a todas las causas de mortalidad. Es decir, cada pez que sobreviva a la mortalidad por pesca tendrá que sobrevivir también a la mortalidad natural (y viceversa) para llegar con vida al final de cada período de tiempo.
Por otro lado, como ningún ser viviente puede morir más de una vez, ni tampoco puede morir por dos o más causas de mortalidad diferentes, tenemos que los peces que mueren por efecto de la pesca ya no pueden ser capturados por otros animales ni tampoco pueden morir por otras causas. Lo mismo sucede con los peces que mueren por causas naturales, al morir son removidos de la población y desaparecen, desapareciendo también la posibilidad de que puedan ser capturados por el hombre.
De esta forma se establece una competencia entre la mortalidad por pesca y las otras causas de mortalidad. Lo que coloca al hombre como un predador más, que trata de capturar a los peces que tienen importancia comercial antes que otros predadores lo hagan.
Desde que nacen, todos los seres vivientes comienzan a afrontar la posibilidad de mo– rir, lo que invariablemente ocurre tarde o temprano. Dependiendo de la longevidad de la especie, todos aquellos peces que al cabo de cierto tiempo no hubieran muerto por la pesca habrán muerto por las causas naturales y, por el contrario, todos aquellos que no hubieran muerto por causas naturales habrán sido capturados por el hombre.
Aun cuando la mortalidad natural y la mortalidad por pesca actúan en forma simultánea, está claro que cada una de estas causas de mortalidad actúa en forma independiente; y si tomamos períodos de tiempo infinitamente pequeños (tales que dt ∞0) podemos expresar sepa– radamente los cambios instantáneos que ocurren en el número de peces que mueren por cada una de estas causas de mortalidad. Lo que en el caso más simple podrá representarse por las ecuaciones diferenciales siguientes:

que representa el incremento instantáneo del número de peces muertos por las causas naturales;

que representa el incremento instantáneo del número de peces muertos por la pesca; y

que representa el incremento instantáneo del número de peces muertos por ambas causas en conjunto, donde:
M, F y Z representan a cada una de las tasas instantáneas de mortalidad natural, por pesca y total;
t = tiempo;
Nt = número de peces existentes en la población o en la clase anual en cada momento t;
C = captura en número, número de peces muertos por la pesca;
D = número total de peces muertos; y
DM = númuero de peces muertos por causas naturales.
De momento, la ecuación que más nos interesa es la que se refiere a los incrementos instantáneos en la captura o en la número de peces muertos por la pesca. Esto debido a que desde el punto de vista de la población, la captura es de por sí un factor importante; pero se hace más importante en cuanto es el único factor que puede ser controlado y regulado por el hombre. Además de que el dato de la captura o número de peces muertos por la pesca es uno de los pocos, y a veces el único dato de la población que puede ser medido y observado directamente y que puede ser relacionado posteriormente con el número de individuos vivos existentes.
Si partimos de la ecuación 4.2 que expresa el incremento instantáneo de la captura en número, y reemplazamos el número de individuos (Nt) por su expresión correspondiente de la ecuación 2.11, tenemos que:
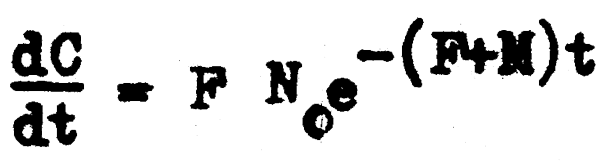
y si integramos para un intervalo de tiempo (t) cuyos límites son t = 0 y t = T, que corres– ponden a los momentos en que el número de individuos es No y NT, y los valores de captura son C = O y C = CT respectivamente, tenemos:

llegamos a la muy conocida ecuación de captura:

que expresa el número de peces muertos por la pesca (C) durante un período de tiempo (T), en función al número de peces (No) existentes al inicio de dicho período y a la mortalidad (M y F) a la que han estado expuestos estos peces en dicho período de tiempo.
Siguiendo este mismo procedimiento a partir de la ecuación diferencial 4.1 se llegará a expresar el número de peces muertos por las causas naturales (DM), en función a los mismos3arámetros:

Y si se hace lo mismo para el número total de muertos, con todas las causas de mortalidadincluidas (ecuación diferencial 4.3) llegaremos a la ecuación:
| DT = No (1 -e -(F+M)T); | (4.6) |
que es exactamente igual a la ecuación 2.12.
A partir de estas ecuaciones podemos determinar que la tasa de mortalidad o proporción total de peces muertos (AT) es:

que la tasa de explotación (u) es:

y que la relación entre el número de peces muertos por la pesca y el número total de peces muertos, también conocida como fracción de muerte por explotación (E) es:

A partir de estas ecuaciones podemos deducir que la pesca actúa sobre la población co– mo un factor de mortalidad más, que se adiciona a las otras causas de mortalidad y hace que las clases anuales disminuyan más rápidamente disminuyendo también la población que resulta de sumar a los sobrevivientes de cada una de las clases anuales existentes. Mientras que la mortalidad natural M puede considerarse siempre como un factor constante que afecta por igual y en forma permanente a todos los peces, la mortalidad por pesca F no siempre puede ser con– siderada como una constante, ya que en gran parte está sujeta a los deseos o habilidades del hombre.
El hombre puede decidir pescar algo más en un año y algo menos en el otro; puede deci– dir pescar a las clases anuales desde que alcanzan el tamaño o la edad del reclutamiento o puede comenzar a pescarlas algo más tarde, cuando han alcanzado una edad o un tamaño prome– dio algo mayor. En fin, hay una serie de circunstancias bajo las cuales la mortalidad por pesca puede variar, pero en todo caso, gran parte de estas circunstancias pueden ser detec– tadas y tomadas en consideración al estudiar la dinámica de las poblaciones de peces.
Considerando la situación más simple, que se da cuando existen condiciones ideales de equilibrio, podemos seguir a una clase anual a través del tiempo introduciendo la ecuación de captura; y a modo de ejemplo podemos comenzar con la clase anual de una especie hipoté– tica que tenga una mortalidad natural equivalente a M = 0,35 por ano; que se recluta a la edad tr = 2 años; que comience a ser capturada a la edad de tc = 4 años; que experimente una mortalidad por pesca equivalente a F = 0,4 por año; y que al momento de pasar a la fase posrecluta en el año 1968 esté compuesta por R = 49 658 individuos, que constituyen el re– clutamiento de esta clase anual. Con estos datos, y mediante la aplicación de las ecuacio– nes 3.3, 4.4 y 4.5 se ha calculado el número inicial de individuos (No), el número de peces capturado (C), y el número total de peces muertos por otras causas (DM) en cada año, cuyos valores aparecen en la tabla 4.1 y en la Fig. 4.1. Además se ha calculado para períodos crecientes de tiempo el número de peces muertos por la pesca, el número de peces muertos por otras causas y el número total de muertos, cuyos valores juntos con los de sobrevivencia en número aparecen en la Fig. 4.2.
En este caso, de la aplicación de las ecuaciones 4.9 y 4.8, vemos que año tras año la fracción de muerte por explotación (E = 0,5333) y la tasa de explotación (u = 0,28140) se mantienen constantes. Circunstancia que tiene una gran aplicación práctica ya que si para cada año (i) la captura (Ci) es directamente propocional al número inicial de individuos (Ni), es decir, si:
| Ci = ui Ni; | (4.10) |
donde el grado de proporcionalidad está dado precisamente por ui, veremos que es posible seguir a través del tiempo las fluctuaciones y determinar la velocidad de cambio de las cla– ses anuales y de la población mediante el análisis de las capturas que de ellas se obtengan, bastando para esto que estemos en condiciones de asumir que M, F y Z son constantes aun cuan– do no sepamos qué valor tiene cada una.
Por ejemplo, si para un período de dos o más años bajo estas condiciones de equilibrio, obtenemos valores consecutivos de la captura, donde para cada año (i) de acuordo con la ecuación 4.10, la captura es:
Ci = ui Ni
Ci+1 = ui+1Ni+1
Ci+2 = ui+2 Ni+2
•
•
•Ci+n = ui+n Ni+n
y si se cumple que:
ui = ui+1 = ui+2 = ui+n
podemos fácilmente determinar las fluctuaciones relativas de la población o de la clase anual bajo estudio, ya que en todo momento el número de peces vivos al inicio de cada año es direotamenteproporcional a las capturas durante ese mismo año.
Además, si a continuación relacionamos la captura de un año (i) con la captura del año inmediato anterior (i-1); podemos calcular la tasa de sobrevivencia (S) para cada año (i) ya que, de acuerdo con la ecuación 2.4 y 2.5:
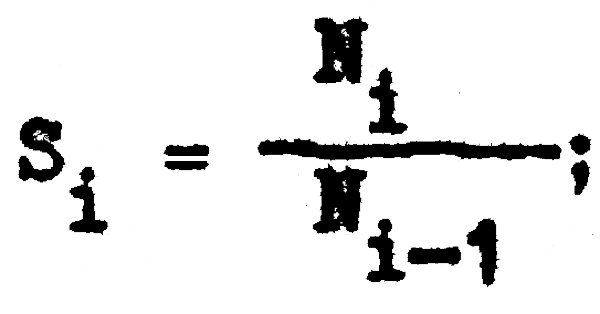
y por lo tanto:
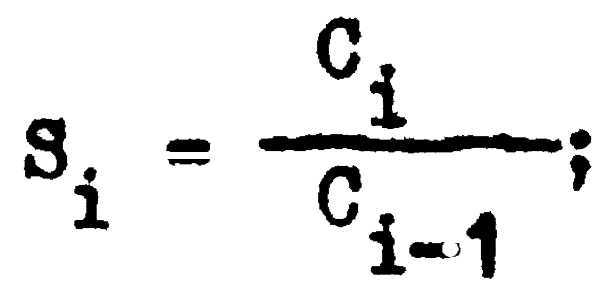
Al haber hallado el valor de Si podemos calcular el valor de Z, ya que como t = 1:
Si = e-Z;
y el valor de Z es:
Z = -ln (Si);
también podemos calcular la proporción de peces muertos (A), ya que:
Ai = 1-Si = 1-e-Z
Para comprobar numéricamente estas aseveraciones podemos tomar cualquier par consecutivo de datos de captura de la tabla 4.1, y si por ejemplo tomamos:
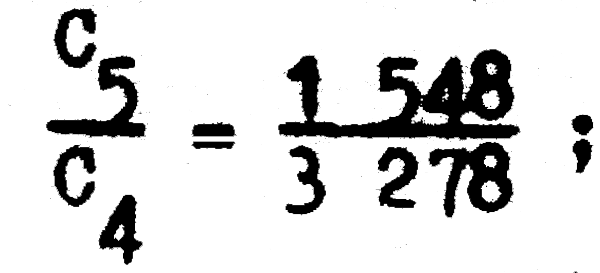
encontramos que la tasa de sobrevivencia es:
S = 0,4722 ;
Z = 0,75 ;
el coeficiente instantáneo de mortalidad total es:
Z = 0,75 ;
y la proporción total de peces muertos es:
A = 0,5278
Además, si por cualquier método alternativo podemos obtener el dato de M o el dato de F, podremos reconstruir íntegramente la historia de la clase anual, ya que teniendo uno de estos coeficientes de mortalidad podemos fácilmente calcular el otro a partir de:
Z = F+M;
y teniendo M, F, Z y Ci podemos calcular el número de individuos (Ni) existentes en cada año, ya que de la ecuación de captura:
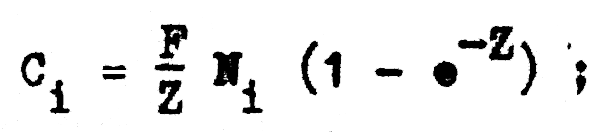
podemos despejar (Ni), de tal forma que:
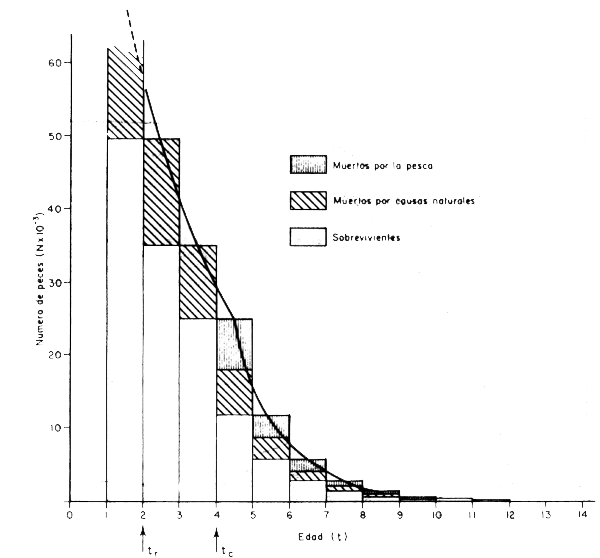
Figura 4.1 Número de sobrevivientes, número de muertos por causas naturales y número de muertos por la pesca por año en una clase anual
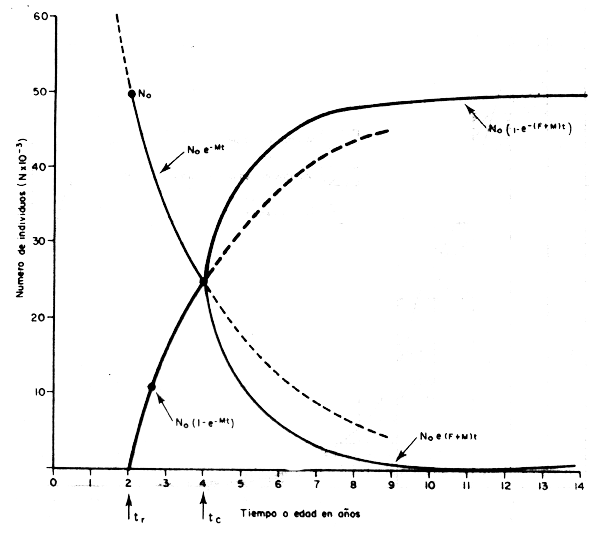
Figura 4.2 Sobrevivencia y mortalidad total de una clase anual a través del tiempo
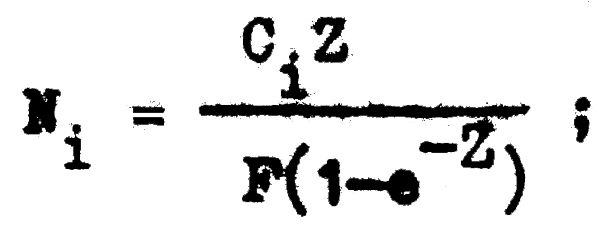
y a partir de este valor, el número de individuos existentes en cualquier otro año será:
Ni+n = Ni e-Zn;
o también:
Ni-n = NieZn;
En el caso que la mortalidad por pesca sea variable, lo que por cierto es más común en la vida real, es siempre posible hacer el mismo seguimiento y reconstrucción de cada clase anual y (sumando los resultados parciales obtenidos para cada clase anual) de toda la población.Pero, en este caso, además de la captura para cada año i, es necesario conocer la tasa instantánea de mortalidad natural M y además, para poder iniciar el seguimiento en uno u otro sentido es necesario tener un valor de F o de N para cualquiera de los años considerados.
De esta forma, si conocemos M que es constante, si conocemos la captura C obtenida en cada año, y además conocemos el valor de F para un año i en particular, podemos iniciar la reconstrucción de la historia de la clase anual a partir de ese mismo año. El número de individuos existentes al inicio del año lo podemos hallar despejando este valor de la ecuaciónde captura, despejando así nuestra primera incógnita:

Como la captura para el año anterior (i-1), de acuerdo con la ecuación de captura, es:

donde hay dos incógnitas: Fi-1 y Ni-1, recordamos que:
Ni = Ni - 1 e-(Fi-1+M));
de donde podemos despejar:
| Ni-1= Nie(Fi-1+M); | (4.13) |
que era una de nuestras incógnitas, la cual puede ser reemplazada por este nuevo valor en la ecuación 4.12 llegando así a la ecuación:
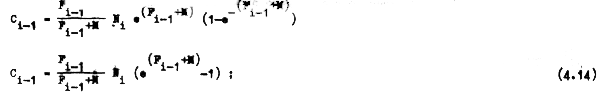
donde la única incógnita es Fi-1.
Como Fi-1 no puede ser despejada de la ecuación 4.14, para calcular su valor se esta blece primero la relación (Vi) entre la captura Ci-1 (ecuación 4.14) y el número de individuosNi (ecuación 4.11), de forma que:

y luego, por aproximación numérica (cosa muy fácil con el uso de las modernas calculadoras programables) o mediante el uso de tablas, se procede a establecer el valor de Fi-1 que corresponde al valor calculado de Vi.
Con este nuevo valor de mortalidad Fi-1 se tienen nuevamente los valores iniciales (C, F, y M) para el año inmediato anterior (i-1), el cual se toma como dato inicial (i) y se repite el ciclo de las ecuaciones 4.11, 4.13, 4.14 y 4.15 para cada año, hasta completar y reconstruir la historia de toda la clase anual, y al repetir este mismo seguimiento para varias clases anuales podremos también reconstruir la historia de toda la población.
Ya en la sección anterior hemos visto que el número de individuos que son capturados o que mueren por la pesca se inorementa a cada instante de acuerdo con la función diferencial:
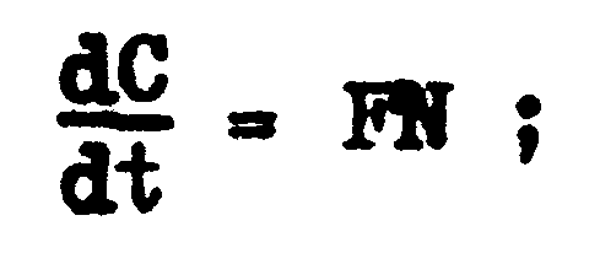
de donde se deriva la ecuación 4.4 que expresa que en cada unidad de tiempo la captura en número es directamente proporcional al número de individuos existentes, siendo este grado de proporcionalidad mayor o menor de acuerdo con el valor que tenga la tasa de explotación (u, ecuación 4.5) y en especial de acuerdo al valor que tenga la mortalidad por pesca representadapor F.
En el caso de la ecuación de captura vemos que la edad o el tamaño promedio de los individuosexistentes no tiene aparentemente ninguna importancia, ya que si se trata de individuos grandes o de individuos pequeños el resultado en número de piezas muertoas o capturadas por cada unidad de mortalidad será siempre el mismo.
En cambio el rendimiento o captura en peso, sí está fuertemente influenciado por la edad o el tamaño promedio de los ejemplares capturados ya que el rendimiento (Y) es en todo momento igual al número de peces capturados (C) y al peso promedio (ω) que tienen dichos individuos. De forma que a cada instante i:
Yi = Ci∞i
y en cada intervalo de tiempo el rendimiento se incrementa de acuerdo con el incremento del número de peces capturados y de acuerdo con el peso promedio que tengan los peces en ese preciso instante, de forma que:
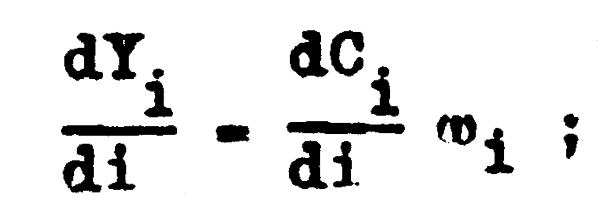
y como dCi/di = FNi tenemos que a cada instante el rendimiento se incrementa de acuerdo con la función diferencial:
| dYi = FNiwidi; | (4.16) |
pero como estamos interesados en lo que sucede en períodos de tiempo más largos, que permitandeterminar el rendimiento de una clase anual a través de toda su existencia o el randimiento de toda la población en un año en particular, debemos sumar todos los intervalos infinitesimalesde tiempo de la ecuación 4.16 integrándolos entre los límites tc que es la edad a la cual las clases anuales recién comienzan a rendir en las capturas, y una edad t siguiente, de forma que el rendimiento a la edad t (t > tc) es:

y, si recordamos las ecuaciones 3.3a y 2.21 podemos desarrollar esta expresión siguiendo la historia de una clase anual explotada, reemplazando Ni por su expresión:
Ni = Re-M(tc-tr) e-Z(I-tc)
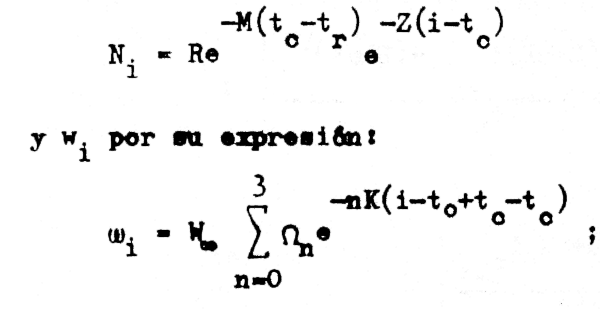
donde previamente hemos sumado y hemos restado un mismo valor de tc al valor de i. La ecuación4.17 es:
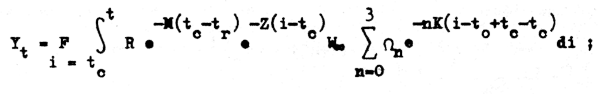
sacando parte de las expresiones que sean constantes fuera de la integral:
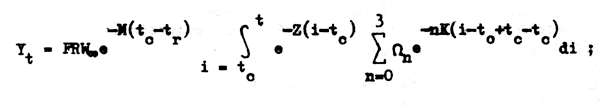
efectuamos parte de las operaciones indicadas y sacamos el signo sumatorio fuera de la iutograly así tenemos:
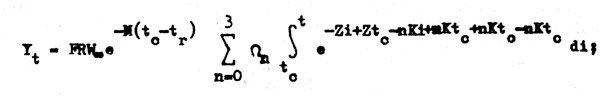
ordenando:
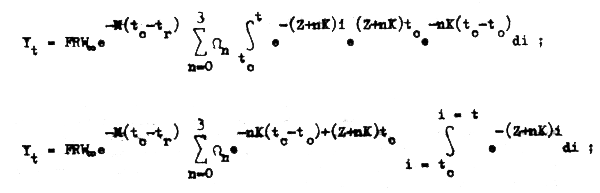
que ahora sí puede ser integrado fácilmente:
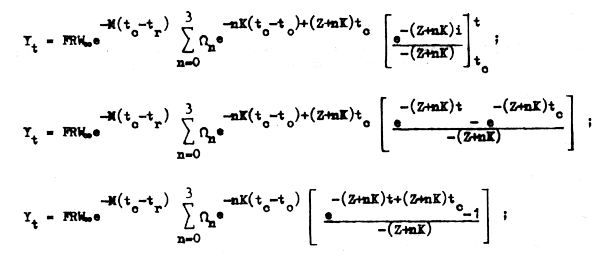
y así finalmente tenemos la ecuación:
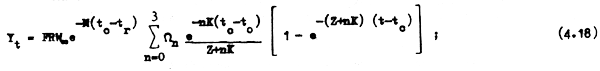
que nos permite conocer la captura en peso o el rendimiento (Y) de cualquier clase anual desde el momento en que apareció en la pesquería y comenzó a ser capturada (tc) hasta cualquierotra edad t posterior.
Si queremos determinar el rendimiento de una clase anual ya extinguida o si queremos proyectar el rendimiento total de una clase anual basta reemplazar t por tλ en la ecuación 4.18:
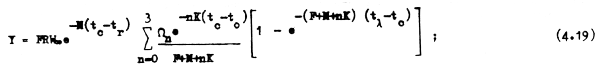
y así tenemos el rendimiento de una clase anual a lo largo de toda su vida, desde que se reclutao ingresa a formar parte de la población explotable hasta que se extingue.
Si las condiciones mencionadas para la ecuación 3.7 se cumplen, es decir si el reclutamientoes constante y si la población se encuentra en equilibrio, la ecuación 4.19 puede representar también el rendimiento anual de toda la población.
La ecuación 4.19 puede todavía simplificarse si es que la edad máxima tλ es muy grande o si es que de acuerdo con la definición de tλ hacemos que la tasa de sobrevivencia e-Ztλ se aproxime a cero. En cualquiera de estos casos el último término de la ecuación 4.19 se hace infinitamente pequeño, pudiéndose descartar de forma que:
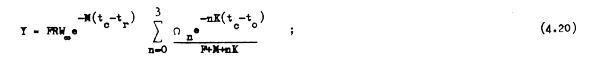
Como vemos, en esta ecuación hay dos tipos de parámetros:
1) aquellos que describen alguna característica inherente a la especie, que están controlados por factores genéticos o del medio ambiente, y sobre los cuales el hombre no tiene ningún dominio, y
2) aquellos que pueden ser controlados por el hombre.
La mortalidad natural (M), el crecimiento (K), la edad a la cual se produce el reclutamiento(tr), la edad a la cual el pez comienza teóricamente a crecer (to), el peso máximo que puede alcanzar (W∞) y la longevidad (tλ) son todos ellos parámetros poblacionales del primer tipo, y a menos que ocurran cambios notables en el ecosistema, suelen mantenerse estables,incluyéndoseles casi siempre como constantes en todos los modelos de dinámica de poblaciones.
En cambio la mortalidad por pesca (F) y la edad a la primera captura (tc) sí son parámetrospoblacionales que el hombre puede controlar. Pudiendo aumentarlos o disminuirlos dentro de ciertos límites cuando aumenta o disminuye su capacidad de captura u orienta sus faenas de pesca hacia la captura de ejemplares de mayor o de menor tamaño.
El reclutamiento (R), que es uno de los factores más importantes en la dinámica de la población, y el cual analizaremos más en detalle en el capítulo 6, puede considerarse muchas veces como constante. Pero no siempre es ese el caso, razón por la cual resulta más realistay se facilitan las comparaciones cuando la ecuación de rendimiento se expresa en términos del rendimiento por recluta (Y/R); y así, de la ecuación 4.19 tenemos:
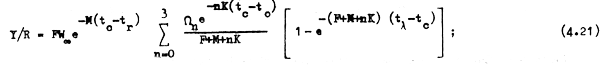
Donde todos los parámetros son constantes, a excepción de la mortalidad por pesca F y la edad a la primera captura tc, que pueden ser controlados por el hombre y permiten ir aumentandoir disminuyendo el rendimiento por recluta.
Por ejemplo, si en la ecuación 4.21 dejamos a la mortalidad por pesca como única variable,hacemos tc = 3,0 años y mantenemos a todos los otros valores constantes, donde W∞ = 82,6 kg; M = 0,2; tr = 3 años; K = 0,18, vemos (fig. 4.3) que el rendimiento por recluta(Y/R) aumenta con rapidez con los primeros incrementos de la mortalidad por pesca (F), llega a un máximo que en este caso es Y/R = 11,377 cuando F = 0,337, y luego comienza a disminuir lentamente, acercándose a medida que F tiende a infinito a un valor límite de Y/R que resulta ser igual al peso promedio que tiene cada recluta cuando alcanza la edad a la primera captura, es decir Y/R = ωtc cuando F→∞. En este caso, como tc = 3 años, ωtc = 7,176 kg; ya que:
wtc = W∞ (1 - e -K(tc - tc))3;
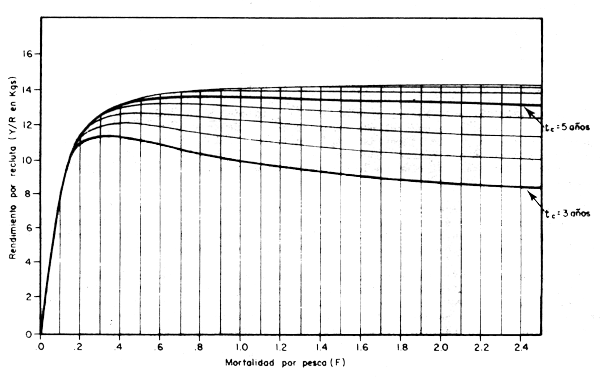
Figura 4.3 Curvas de rendimiento por reculta (Y/R) en función a la mortalidad por pesca (F) considerando distintos valores de edad a la primera captura (tc) y condiciones idealsde equilibrio
Si cambiamos la edad a la primera captura la relación entre Y/R y F cambia también; por ejemplo si hacemos tc = 5 años encontramos que cambia en algo la forma de la curva, cambia el nivel de rendimiento máximo posible, que en este caso es Y/R = 13,50 kg y para el cual se requiere un esfuerzo o mortalidad por pesca bastante mayor, equivalente a F = 0,82, para alcanzar este máximo.
Este rápido aumento en el rendimiento por recluta (Y/R) que se aprecia en la fig. 4.3 con los primeros incrementos en la mortalidad por pesca (F) se debe a que con cada incrementode F hay una menor parte de la biomasa total que es tomada por la mortalidad natural (M, que es constante) ya que los peces comienzan a ser tomados por la pesca antes que las causas naturales los maten. La reducción que se aprecia a valores mayores de F (con tc = 3,0) se debe a que si bien la pesca logra obtener más peces que la mortalidad natural, la misma pesoa se encarga de capturar a estos peces rápidamente, no dejándoles crecer lo suficiente. Nótese que si se aplican altas mortalidades por pesca con una edad tc mayor (es decir cuando los peces ya han crecido más) esta disminución no se produce, o en todo caso no es tan marcada.
Si por el contrario, en la ecuación 4.21 hacemos que tc sea la variable y le damos un valor determinado a la mortalidad por pesca, digamos F = 0,3, podemos construir una curva de rendimiento por recluta en función a tc como la de la fig. 4.4, donde apreciamos que si se comienza a capturar a los peces desde muy pequeños, cuando recién se han reclutado, el rendimiento por individuo es relativamente bajo. Esto debido al bajo peso promedio que tienen los peces a esta edad. Pero a medida que se va aumentando la edad a la primera capturael rendimiento por individuo también va aumentando. Esto significa que se está aprovechando mejor la capacidad natural de incremento en peso de cada individuo, pero si se sigue aumentando la edad tc, llegará un momento en que el rendimiento por recluta comenzará a disminuir, luego de haber alcanzado su valor máximo (Y/R = 12,55, cuando tc = 5,0). En esta parte el Y/R disminuye debido a que a esas edades ya el crecimiento no sólo no es tan rápido, sino que, además, es superado por la mortalidad natural, que en este caso es la que se lleva una mayor parte de la población. A medida que tc se aproxima a tλ vemos que el rendimiento por recluta disminuye hasta llegar a cero. Esto es lógico, ya que para permitirque los peces lleguen a viejos sin haber sido capturados ha habido que hacer que el tiempo durante el cual los peces han estado expuestos a la mortalidad por pesca (tλ -tc en la ecuación 4.21) haya sido cada vez menos, aproximándose a cero a medida que tc - tλ, creándose así una situación semejante a que si F se fuera reduciendo hasta llegar a cero.
Una idea más precisa de las múltiples alternativas que presenta la posibilidad de combinarla edad a la primera captura (tc) y la mortalidad por pesca (F) se da con la fig. 4.5. En forma tridimensional se presenta una combinación de las dos figuras anteriores: en los ejes horizontales tenemos a la mortalidad por pesca (F, que varia entre 0 y 2,5) y a la edad a la primera captura (tc, que varia entre tr = 3 años y tλ = 25 años); y en el eje vertical tenemos el rendimiento por recluta (Y/R) correspondiente a cada combinación posible de F y tc.
Apreciaremos que los rendimientos por recluta máximos se obtienen a edades tc próximas a la edad crítica (ecuación 2.28). Es decir, a la edad a la que la clase anual alcanza su biomasa máxima cuando sólo está expuesta a la mortalidad natural. Si a esta edad (tc = t* = 7,02 años en este caso) capturamos a todos los peces existentes, se obtiene el rendimiento por recluta máximo posible (Y/R = 14,37 kg), pero para esto es necesario que F se haga infinitamente grande. Cosa muy dificil y muy costosa de hacer en la práctica, y que en la figura generará un punto en el espacio por fuera del gráfico (Y/R = 14,37, tc = 7,02, F = ∞).
Para cada edad tc hay siempre un valor de F que permite obtener un máximo de rendimiento,sin cambiar la edad de la primera captura (lo cual en la práctica significa no cambiar el tamaño de malla o la selectividad de las artes de pesca existentes). Así, por ejemplo, a la edad tc = 3 años el rendimiento por recluta es máximo (Y/R = 11,375) con una F = 0,33; y a la edad tc = 3,5 el rendimiento es máximo (Y/R = 12,034) con una F = 0,4 años; y a la edad tc = 4,0 años este máximo (Y/R = 12,640 kg) se obtiene con un F = 0,49 y así sucesivamente.Con estos valores de Y/R máximos para cada edad tc (lo que significa ir variando F
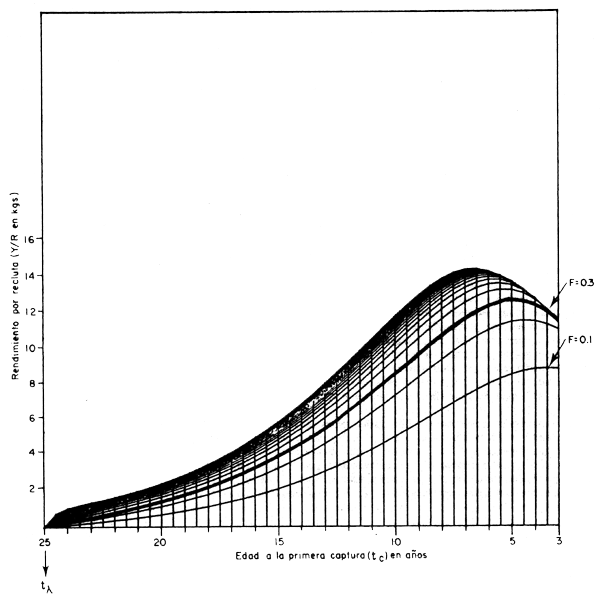
Figura 4.4 Curvas de rendimiento por recluta (Y/R) en función a la edad a la primera captura (tc) considerando distintos valores de mortalidad por pesca (F) y condiciones ideales de equilibrio
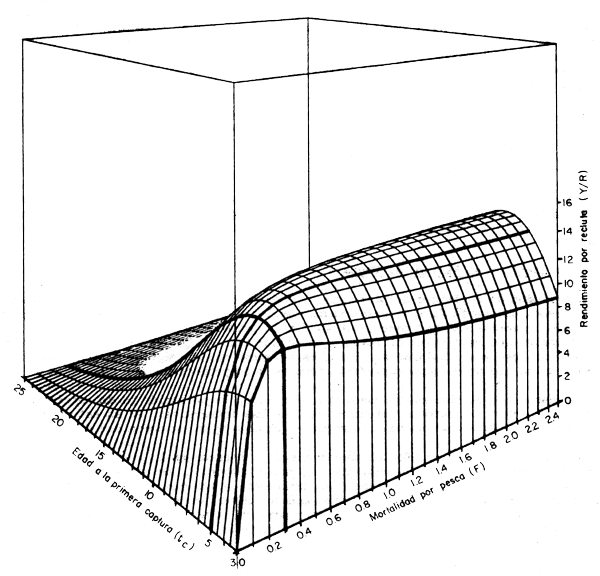
Figura 4.5 Curvas de rendimiento por recluta (Y/R) en función a la edad a la primera captura (tc) y a la mortalidad por pesca (F) considerando condiciones ideales de equilibrio
dentro de cada tc hasta encontrar el Y/R indicado) se puede construir una curva (curva AA' en la fig. 4.6) que permite encontrar el esfuerzo de pesca más adecuado para cada tamaño a la primera captura.
La fig. 4.6 no es sino la proyección en un solo plano de la vista superior de la fig. 4.5, lo que permite trazar un contorno que une valores iguales de rendimiento por recluta (en este caso Y/R = 2,0; 4,0; 6,0; 8,0; 10,0; 12,0 y 14,0 kg) dando por resultado un diagramade isopletas de rendimiento. En esta figura se ha trazado también la línea BB', que se llama línea o curva de “pesca eumétrica” (del griego eu = bien y metros = medida) que correspondea los valores máximos de rendimiento por recluta (Y/R) que se pueden obtener con cada valor de esfuerzo de pesca (F), cambiando en este caso la edad a la primera captura (lo que en práctica equivale a ir cambiando la selectividad de las artes de pesca) hasta encontrar el máximo posible para el valor de F dado.
Así, por ejemplo, para una pesca equivalente a una F = 0,3, se obtiene un valor máximo de Y/R = 12,552 fijando la edad a la primera captura en tc = 5,0 años. Si la actividad pesqueraes equivalente a una F = 0,5, el máximo de Y/R = 13,491 kg se obtiene cuando tc = 5,5 años; para una F = 0,9 el máximo Y/R = 14,033 kg se obtiene con una tc = 6,1 años y así sucesivamente.
Nótese que en ningún caso los valores máximos de rendimiento por recluta que se obtienen caen dentro de un rango de tamaño superior al de la edad o tamaño crítico, ya que a partir de este momento la mortalidad natural produce más pérdidas que las ganancias que produce el crecimiento.
La variedad de regímenes de pesca que se pueden adoptar al explotar a una población de peces aparecen en cierta forma representados por las diversas combinaciones que se pueden hacer entre la mortalidad por pesca y la edad a la primera captura. En las fig. 4.5 y 4.6 se aprecian los resultados de estas combinaciones en lo que se refiere a la cantidad o peso total de los peces capturados. Pero, es evidente que el régimen de pesca afecta no solamente a las capturas, sino que afecta también a la población.
De acuerdo con el régimen de pesca que se adopte, la población puede sufrir cambios mayores o menores en el número y biomasa total de sus integrantes, en su estructura por edad o por tamaños, en su velocidad de crecimiento e inclusive en su capacidad de reproducción.
Por ejemplo, uno de los efectos más fáoiles de observar es la reducción del tamaño o edad promedio de los peces en la población (y en la captura). Es ampliamente conocido que las poblaciones vírgenes que nunca han sido explotadas o las que están expuestas a un nivel de explotación muy bajo, están constituídas por una mayor proporción de peces viejos. Pero a medida que aumenta el nivel de explotación y aumenta la mortalidad por pesca, la proporciónde peces viejos disminuye rápidamente, disminuyendo también el tamaño promedio de los individuos en toda la población.
Las tallas de peces que más rápidamente desaparecen de una población expuesta a altos niveles de explotación son las correspondientes a los individuos más viejos. Esto debido a que están más tiempo expuestos a mortalidades más altas. En términos relativos los individuosjóvenes tienden en cambio a aumentar, aun cuando en términos absolutos éstos también disminuyen, pero menos.
En la tabla 4.2 se ilustra numéricamente lo que puede suceder con la parte explotable de una población en equilibrio según el régimen de pesca al que se le someta. En el primer casola población no ha sido explotada, y como toda población en equilibrio presenta un predominio de individuos jóvenes. Donde por ejemplo los peces entre 2 y 4 años representan el 45,81% del total (que es 327 107 individuos), los peces entre 8 y 10 años hacen el 7,57% y los peces más viejos, que tienen entre 14 y 16 años, hacen sólo el 1,25% del total.
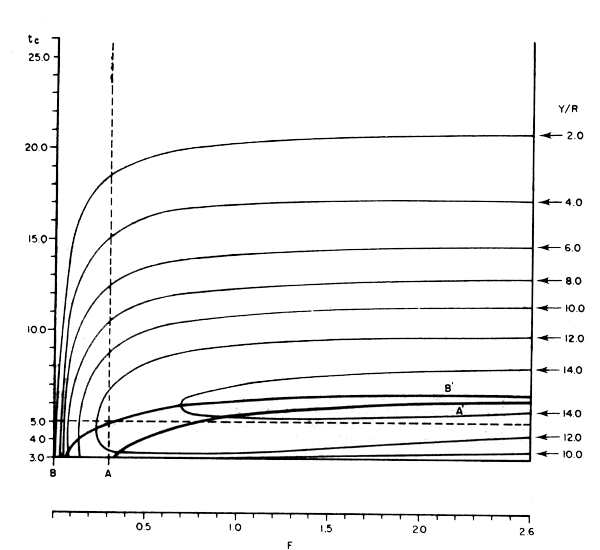
Figura 4.6 Isopletas de rendimiento
En el segundo caso, con una tasa de explotación más o menos baja (u = Ct/Nt = 0,121) vemosque hay una disminución de individuos en todos los grupos de tamaño, pero esta disminución es más marcada en los individuos más viejos. E1 último rango de tamaños (de 15 a 16 años) se reduce hasta 0,132 veces su tamaño previo en la población no explotada mientras que el primer rango (2 a 3 años) sólo se reduce en 0,928 veces. Esta disminución diferencialhace aumentar considerablemente la proporción de individuos jóvenes en la población (59,45% de ejemplares entre 2 y 4 años contra 4,00% de ejemplares entre 8 y 10 años y sólo 0,27% de los ejemplares entre 14 y 16 años de edad) aun cuando el total de individuos ha disminuido a 219,954 ejemplares.
En el tercer caso este cambio es más marcado, y en el último caso, donde la tasa de explotación es más o menos alta, vemos que los peoes más viejos (entre 14–16 años) prácticamentedesaparecen, los otros, que tienen edades entre 8 y 10 años no llegan ni al 1,0%, y los más jóvenes son los que definitivamente dominan. Los peces que tienen entre 2 y 4 años llegan a representar hasta el 77,69% de la población en este caso, mientras que en la poblaciónno explotada sólo llegaban al 45,81%.
Además de los cambios en la composición por tamaños, en la tabla 4.2 es posible apreciarque en cada caso hay una reducción del número total de individuos con una disminución de su edad o tamaño promedio. Esto trae como consecuencia que la biomasa total de la poblacióntambién disminuya, y en este caso disminuye por doble motivo, primero por la reducción del número total de individuos y segundo por la reducción del peso promedio de estos individuos.
Sin embargo, si para las poblaciones de este ejemplo asumimos los mismos parámetros de
crecimiento dados en la tabla 2.2 y utilizados en la fig. 2.3, y si observamos las edades
promedio resultantes en cada caso de la tabla 4.2 (donde ![]() = 5,1 años en el caso de la poblaciónno explotada;
= 5,1 años en el caso de la poblaciónno explotada; ![]() = 4,2 años en la población levemente explotada;
= 4,2 años en la población levemente explotada; ![]() = 3,7 en la población
medianamente explotada y
= 3,7 en la población
medianamente explotada y ![]() = 3,4 años en el caso de la población altamente explotada) encontraremosque a medida que aumenta la tasa de explotación la edad promedio de la población
(
= 3,4 años en el caso de la población altamente explotada) encontraremosque a medida que aumenta la tasa de explotación la edad promedio de la población
(![]() ) se va acercando a la edad a la primera captura (tc), pasando en este caso por la edad a
la cual el individuo desarrolla su máxima velocidad de crecimiento, que es cuando wt=0.296 W∞
y que en este caso corresponde a una edad t = 3,43 años.
) se va acercando a la edad a la primera captura (tc), pasando en este caso por la edad a
la cual el individuo desarrolla su máxima velocidad de crecimiento, que es cuando wt=0.296 W∞
y que en este caso corresponde a una edad t = 3,43 años.
Con una edad promedio de t = 3,4 años, los peces que sobrevivan hasta la edad siguiente(es decir, hasta t = 4,4 años) tienen la posibilidad de incrementar su peso de 126 g a 181 g lo que equivale a un incremento en peso del 43%. Los que tienen una edad promedio algo mayor, t = 3,7 años, sólo incrementarán su peso en un 37% (de 143 a 196 g) en el lapso de un año, y los de la población no explotada que tienen una edad promedio t = 5,15 pueden incrementar su peso si sobreviven en sólo un 19%.
Según esto la pesca tiene el efecto de acelerar la velocidad de crecimiento en peso de la población. Al reducir la edad promedio de los indivuos de la población se hace que ésta crezca más rápidamente, lo que no significa que el tamaño de la población aumente, si no que por el contrario el tamaño o biomasa total disminuye a conçecuencia de la mayor mortalidad introducida.
![]()
![]()
![]()