![]()
![]()
![]()
Les effets d'une amélioration du rapport P/E dans les nutriments dont disposent les ruminants qui se nourrissent de fourrages de qualité médiocre ont déjà été exposés plus haut. La recherche sur l'alimentation de complément est déjà bien avancée dans un certain nombre de pays, portant soit sur une optimalisation de la digestion fermentative (par l'emploi de pierres à lécher contenant de la mélasse et de l'urée), soit sur une optimalisation de l'efficacité d'utilisation des nutriments à partir de la panse (par l'emploi d'un complément protéique de dérivation).
On estime qu'une cinquantaine de pays soumettent actuellement à des essais les pierres à lécher contenant de la mélasse et de l'urée comme moyen de fournir un complément aux ruminants dont le régime alimentaire à base de fourrage accuse une carence critique en certains nutriments.
Le tableau 5.1 indique quelques résultats d'études entreprises en Indonésie sur les effets de l'introduction de pierres à lécher à base de mélasse et d'urée dans les systèmes de production de ruminants utilisant de l'herbe fraîchement coupée. De nombreux pays qui n'ont sur place aucune source de mélasse recherchent actuellement d'autres moyens d'administrer le même assortiment de nutriments à leur bétail indigène.
Il convient de souligner de nouveau qu'il est possible d'améliorer le rapport protéines/énergie dans les nutriments mis à la disposition d'un animal aussi bien en introduisant dans la panse les nutriments dont il y a carence (par exemple le phosphore, le soufre et l'azote non protéique) qu'en les administrant à l'animal (au moyen de protéines de dérivation). Cela permettra d'optimaliser la production à partir des ressources disponibles. Même quand la panse n'est pas optimalisée, l'utilisation d'une protéine de dérivation permet encore d'ajuster le rapport P/E de façon appréciable. Dans certains cas, quand une protéine de dérivation est relativement peu coûteuse ou quand il est difficile de se procurer des pierres à lécher à base de mélasse et d'urée ou d'obtenir des animaux qu'ils s'en servent, un simple complément de protéines de dérivation peut se révéler suffisant pour améliorer notablement l'efficacité d'utilisation du fourrage. D'autre part, la protéine soluble qui est toujours présente dans les compléments protéiques de dérivation, de même que le recyclage vers la panse de l'azote de l'urée, réduisent souvent l'efficacité des pierres à lécher à base d'urée et de mélasse, comme on a pu le constater lors de maints travaux de recherche appliquée; en particulier, quand les vaches laitières reçoivent un apport important de compléments protéiques de dérivation, la pierre à lécher à base de mélasse et d'urée peut n'avoir plus qu'un intérêt marginal. Le tableau 5.2 indique le calcul théorique des rapports P/E. Il va de soi que l'option la meilleure est le recours aux deux types d'alimentation de complément.
Pour le petit éleveur, il est souvent difficile et parfois trop coûteux de fournir au bétail des protéines de dérivation. Il manque souvent d'informations sur les sources de protéines disponibles sur place, et notamment sur le degré de protection des protéines dans la panse. D'une manière générale, les tourteaux d'oléagineux extraits par solvant, la farine de poisson qui a été séchée à la flamme (mais non la farine de poisson séchée au soleil ni l'ensilage de poisson) et les sources de protéines qui ont subi un traitement thermique bénéficient d'une protection non négligeable contre la dégradation dans la panse. Le degré de protection sera renforcé si l'on comprime le complément protéique en présence de xylose, de glucose ou de fructose libres (comme c'est le cas pour la mélasse), auquel cas il se produira une légère réaction de brunissement (observations inédites).
Les principales conditions qui doivent être remplies avant que ces technologies puissent être largement appliquées dans un pays sont les suivantes:
| Bétail | Lieu | Résultat mesuré | Traitement | Réaction à la pierre à lécher | |
| Témoin | Pierre à lécher | ||||
| Vaches laitières (frisonnes) | Java ouest | Rendement laitier (l/j) | 7,44 | 9,92 | |
| Java centre | 5,70 | 8,00 | |||
| Moyenne | 6,57 | 8,96 | + 36% | ||
| Boeufs de boucherie (Ongole) | Java ouest | 1 Gain de poids (kg/j) | 0,182 | 0,400 | |
| " | 2 | 0,333 | 0,526 | ||
| " | 3 | 0,277 | 0,439 | ||
| " | 4 | 0,478 | 0,465* | ||
| " | 5 | 0,200 | 0,278 | ||
| Java centre | 6 | 0,446 | 0,689 | ||
| Moyenne | 0,319 | 0,466 | + 46% | ||
| Ovins (Garut) | Java ouest | Gain de poids (kg/j) | 0,130 | 0,320 | + 46% |
| Caprins (Ettawa) | Java centre | Rendement laitier (l/j) | 0,795 | 1,091 | + 37% |
| Milieu dans la panse | Protéines
alimentaires de dérivation (g/j) | Cellules
microbiennes produites (g/j) | Protéines
microbiennes (g/j) | Protéines
totales (g/j)(P) | AGV
produits (MJ/j)(E) | P/E* (g/MJ) |
| Médiocre | 0 | 800 | 500 | 500 | 41 | 12 |
| Optimalisé | 0 | 1 680 | 1 010 | 1 070 | 30 | 33 |
| Médiocre** | 400 | 830 | 500 | 1 330 | 41 | 22 |
| Optimalisé | 1 680 | 1 010 | 2 690 | 30 | 47 |
* Rapport entre les protéines microbiennes plus les protéines alimentaires et l'énergie AGV.
** Bien qu'il soit admis que le milieu dans la panse n'est pas modifié par l'addition d'aliments protéiques, en fait il aura été amélioré mais le degré d'optimalisation ne pourra pas être le même qu'avec l'emploi d'une pierre à lécher à base de mélasse et d'urée. Le rapport P/E dans ce cas est sous-estimé.
il faut déterminer le degréde protection contre la dégradation dans la panse dont bénéficient les protéines contenues dans les ressources protéiques disponibles;
il faut recenser les sources de protéines potentielles et mettre au point des techniques permettant de neutraliser les composés qui nuisent à la qualité des protéines et de protéger celles-ci (voir plus loin la section sur la technologie alimentaire);
dans certains pays, il n'existe pas de cultures reconnues comme étant riches en protéines dans les régions à forte densité de ruminants. Il faut donc trouver de nouvelles sources de protéines dans ces régions. Il pourrait s'agir de planter des arbres ou des cultures fourragères pour la production de feuillage, de graines ou de gousses, puis de mettre au point des techniques appropriées pour récolter et protéger ces protéines quand elles ne bénéficient pas d'une protection naturelle;
il faut déterminer la réaction des ruminants à des niveaux croissants de compléments protéiques de dérivation afin de permettre une évaluation économique;
il faut mettre au point des techniques de commercialisation et de vulgarisation pour faire parvenir les produits jusqu'aux éleveurs.
L'efficacité de la croissance microbienne est très variable, pouvant aller d'un minimum de 10 g de cellules sèches/MJ de AGV produits dans la panse jusqu'à un maximum de 30 g de cellules sèches/MJ de AGV (voir tableau 5.2). Ces fluctuations s'expliquent par des différences en ce qui concerne l'assortiment de microbes dans une population et par une carence des nutriments spécifiques des micro-organismes qui se développent dans la panse. Dans la plupart des conditions d'élevage où les ruminants reçoivent une ration à base de fourrage de bonne qualité, le rapport P/E est théoriquement de l'ordre de 17 g de cellules/MJ de AGV; avec un fourrage de qualité médiocre, le rapport peut se situer entre 4 et 10 g de cellules/MJ de AGV; il existe donc de larges possibilités de manipuler ce rapport.
Si l'on corrige chez les ruminants une carence chronique de la panse en nutriments nécessaires à la croissance des micro-organismes, cela accroît généralement la digestibilité, l'apport fourrager et les quantités de cellules microbiennes produites par rapport aux AGV. Les techniques exposées ci-après ont toutes pour condition préalable que l'écosystème de la panse ait été amené à un haut degré d'efficacité par une alimentation de complément avant que l'on cherche à manipuler davantage le rapport P/E.
Il existe un certain nombre de substances chimiques qui, lorsqu'on les incorpore au régime alimentaire, améliorent l'efficacité d'utilisation du fourrage de 5 à 20 pour cent (voir Chalupa, 1980). Les principales sont la monensine (ou rumensine), la lasalocide et l'avoparcine. Cependant, l'amélioration qui résulte si les disponibilités d'acides aminés sont équilibrées dans une ration à base fourragère peut accroître de plusieurs fois cette valeur l'efficacité de conversion du fourrage en poids vif chez une vache laitière (voir Leng, 1990).
Bien que le coût soit élevé et les avantages assez faibles, la monensine est largement utilisée dans le monde entier pour l'élevage dans des parcs d'engraissement où les bêtes sont nourries de céréales. On a pensé que ces substances chimiques favorisaient la production de propionate (dans la panse), mais il est plus probable qu'elles améliorent le rapport P/E en réduisant la dégradation des peptides et des acides aminés (voir Russell et al. 1989) dans la panse et peut-être aussi en empêchant le développement des protozoaires, ce qui améliore le rapport P/E dans les nutriments absorbés.
Du fait que ces produits chimiques ont parfois des effets secondaires en ce sens qu'ils réduisent l'apport tout au moins au départ, ce qui supprime souvent tout avantage global lorsqu'ils sont incorporés au régime alimentaire, ils n'ont pas été acceptés pour les ruminants nourris de fourrage.
Les méthodes qui réduisent la dégradation des peptides et des acides aminés dans la panse pourraient constituer un moyen très efficace de stimuler le rapport P/E avec des rations protéiques modérées; elles seraient extrêmement efficaces lorsqu'on dispose de sources de protéines solubles et elles éviteraient la nécessité de protéger les protéines par des moyens chimiques. En résumé, il faudrait rechercher des substances chimiques actives contre les micro-organismes qui décomposent les peptides et les acides aminés car ces organismes sont différents, semble-t-il, de ceux qui provoquent la dégradation des protéines en peptides et en acides aminés (Russell et al., 1989).
Manipulation de l'assortiment microbien dans la panse
Les populations microbiennes de la panse se composent de protozoaires, de bactéries et de champignons anaérobies, ainsi que de virus et de phages. La densité de peuplement dans chaque catégorie peut être extrêmement variable selon le régime alimentaire mais, d'une manière générale, les produits finals de la digestion fermentative sont les mêmes du point de vue qualitatif.
Bien qu'on ait déployé de grands efforts pour manipuler les populations de protozoaires (voir Leng, 1984), ils n'ont guère été couronnés de succès jusqu'à ce qu'on ait inventé des systèmes de laboratoire satisfaisants pour la défaunation afin de pouvoir étudier des effectifs nombreux de ruminants (voir Bird et al., 1990).
Il est hors de doute que la défaunation accroît le débit des cellules microbiennes vers l'intestin (la défaunation améliore le rapport P/E dans les nutriments absorbés) (voir à ce sujet Nolan et al., 1989). Jouany et Ushida (1990) ont clairement démontré par ailleurs qu'avec un régime contenant une forte proportion de protéines vraies, une plus grande quantité de protéines alimentaires parvient à l'intestin outre les protéines microbiennes supplémentaires chez les animaux qui sont exempts de faune par rapport à ceux qui hébergent une population normale de protozoaires.
A condition que la quantité de protéines de dérivation dans la ration soit inférieure aux besoins de l'animal hébergeant une faune, l'élimination des protozoaires de la panse permet une utilisation plus efficace du fourrage. La défaunation a parfois augmenté de 50% les niveaux de production des ruminants (voir Bird et Leng, 1978).
Avec des aliments concentrés de haute qualité contenant une forte proportion de protéines de dérivation, la meilleure efficacité résultant de la défaunation peut ne pas être exprimée, mais le niveau de protéines alimentaires doit pouvoir être réduit de 20 à 50% avec la même efficacité et le même taux de production.
Le potentiel d'application de la défaunation est mis en lumière par la meilleure efficacité d'utilisation de la ressource fourragère de base chez les animaux ayant subi une défaunation ou qui sont exempts de faune. La défaunation ou le maintien de l'absence de faune a permis d'accroître le taux de croissance de 30 à 60% chez des ovins, des bovins et des buffles (Bird et al., 1990) et les résultats d'une étude préliminaire permettent d'espérer un supplément de lait de 2 litres par jour chez les frisonnes (Moate, 1989). Les problèmes consistent à:
trouver des traitements convenables de défaunation ou d'élimination des protozoaires qui soient simples et économiques;
trouver des mécanismes d'administration des composés antiprotozoaires aux ruminants avec les systèmes d'exploitation actuels.
Agents de défaunation
Les protozoaires de la panse sont sensibles à tout un éventail de substances chimiques, notamment celles qui perturbent les membranes celluaires (c'est-à-dire qui exercent une action superficielle). A la suite d'une enquête très restreinte sur des produits naturels (l'enquête a porté sur moins de 100 fourrages), on a pu identifier deux sources d'agents antiprotozoaires qui peuvent être utilisés à faibles doses pour supprimer ou éliminer les protozoaires dans la panse (Assafa, G., Klieve, A. et Leng, R.A., données inédites).
Il se peut que la riche flore tropicale de beaucoup de pays en développement soit une source de nombreux autres composés ou fourrages antiprotozoaires.
Un domaine de recherche prioritaire serait de mettre au point une épreuve antiprotozoaire simple dans un certain nombre de laboratoires dans des zones climatiques différentes, puis de recenser rapidement les sources potentielles de substances antiprotozoaires.
Au départ, le plus simple serait d'observer les effets de quantités croissantes d'un extrait soluble d'un fourrage sur la viabilité des protozoaires placés en étuve avec les substances en question pendant 2 heures par exemple. Il faudrait manifestement que cette épreuve fût suivie d'une expérimentation sur l'animal, puis d'études de croissance pour déterminer si les autres effets de ces composés antiprotozoaires pourraient être nocifs pour l'animal.
Mécanismes de la défaunation
Il semble bien que le seul moyen pratique de manipuler la population de protozoaires de la panse consiste à administrer quotidiennement ou à intervalles réguliers des suppléments stratégiques contenant un agent de défaunation.
Comme indiqué précédemment, la défaunation a un très net effet de stimulation sur le rapport P/E et sur l'efficacité de la conversion du fourrage.
Des recherches seront nécessaires pour examiner l'administration de tout agent potentiel de défaunation par l'intermédiaire du fourrage de base, soit dans un aliment protéique de complément, soit dans une pierre à lécher à nutriments multiples. Ce dernier moyen serait potentiellement le plus pratique, mais la posologie serait nécessairement réduite à des quantités très faibles car l'apport provenant de ces pierres ne représente généralement qu'une fraction infime de l'apport total.
Peut-être serait-il possible d'administrer un agent antiprotozoaire extrêmement actif au moyen d'un dispositif à libération lente placé dans la panse (voir Ellis et Costigan, 1989), mais là encore le coût risque de militer contre l'adoption de cette méthode.
Par un heureux hasard, des travaux de recherche récents ont montré que l'incorporation dans la ration des ovins de petites quantités d'un minéral argileux (la bentonite, une argile de montmorillonite) améliorait la croissance de la laine de 20% chez les animaux hébergeant des protozoaires, mais seulement de 5% chez les ovins exempts de faune (Fenn et Leng, 1989; Fenn et Leng, 1990). On ignore le mécanisme d'action de la bentonite, mais ou bien elle améliore le rapport P/E, ou bien elle améliore expressément le débit d'acides aminés soufrés à partir de la panse, car ceux-ci sont les principaux nutriments contrôlant la croissance de la laine. Les problèmes consistent:
à comprendere les mécanismes d'action de la bentonite car c'est un aspect capital pour déterminer les sources de bentonite à utiliser et les systèmes dans lesquels elle améliorera la productivité des animaux;
à trouver un mécanisme d'administration de la bentonite par l'intermédiaire d'aliments de complément.
Il semble que la composition en acides aminés des protéines microbiennes de la panse soit relativement constante quel que soit l'assortiment de microbes dans la population totale (Czerkawski, 1986). Pour la croissance de la laine, les principaux acides aminés limitants chez les ovins sont la méthionine et la cystéine et, pour la croissance de l'animal, ce sont peut-être ces mêmes acides aminés plus la lysine et la thréonine.
Il est hors de doute que si l'on pouvait manipuler les microbes de la panse pour produire des protéines protoplasmiques riches en acides aminés carentiels, il en résulterait une amélioration de la production de laine. La méthionine est décomposée assez lentement dans la panse et il semble que les peptides et les protéines riches en acides aminés contenant du soufre soient également décomposés assez lentement par les microbes (Nugent et Mangan, 1981). Par conséquent, l'établissement dans la panse des micro-organismes qui produisent ces acides aminés pourrait accroître le débit des acides aminés contenant du soufre et, partant, l'efficacité de la croissance de la laine.
Les problèmes consisteront à isoler et à identifier l'ADN codé pour les enzymes nécessaires, à mettre au point des vecteurs et à transformer les microbes de la panse. Ensuite, les microbes de la panse transformés devront pouvoir s'y développer en grandes quantités et y rester, notamment pendant les périodes où ces microbes de la panse subissent une contrainte nutritionnelle.
Il est hors de doute qu'une alimentation de complément destinée à rendre plus efficace la digestion dans la panse des bovins nourris de fourrage médiocre accroît le rendement des ressources fourragères locales. Les recherches menées par l'Office national indien de développement de l'industrie laitière (NDDB) l'ont amplement démontré. Le NDDB a mis au point des pierres à lécher multinutriments à base de mélasse et d'urée qui sont livrées aux éleveurs villageois à prix coûtant. Il en va de même actuellement dans une cinquantaine de pays tropicaux en développement. Toutefois, rares sont les pays qui exploitent la possibilité de mettre au point et d'utiliser des concentrés protéiques de dérivation. Un exemple de l'application de ces techniques alimentaires en Inde est fourni ci-après.
Dans leurs fabriques de produits d'alimentation animale, deux enterprises laitières (à Kedah et à Baroda) produisent et vendent un concentré riche en protéines (30%) qui contient une forte proportion de protéines de dérivation (une centaine de tonnes par jour rien qu'à Kedah). Les farines protéiques sélectionnées proviennent toutes de fabriques d'olégineux et elles ont été soumises à une technique de protection comportant une extraction au solvant et un traitement thermique. En outre, le concentré protéique final est aggloméré avec environ 8% de mélasse, ce qui semble lui conférer une protection supplémentaire (sans doute par suite de la chaleur dans la presse et d'une certaine réaction de brunissement du sucre avec la lysine provenant des protéines superficielles).
Pour une opération commerciale avec une usine produisant 100 tonnes par jour, il est nécessaire d'utiliser des assortiments de protéines de sources diverses et le contrôle de la qualité est indispensable. Il est largement admis que la mesure dans laquelle les protéines échappent à la panse est variable selon leur origine et il semble que la confection d'aggloméré confère une meilleure protection.
Le tableau 5.3 compare quelques méthodes d'utilisation de protéines de dérivation dans des systèmes de production laitière.
| Fourrage de base | Compléments | Production laitière |
| 1. Ensilage de graminées tropicales/maïs ou autres cultures plus 1-2 kg de paille de riz par jour | Libre choix de pierres à lécher à mélasse/ urée plus agglomérés protéiques (30% AC) (350 g/kg de lait) | 5.000-6.500 kg/305 j |
| 2. Paille de riz/paille millet (8 kg/jour) | Agglomérés de protéines de dérivation 300 g/kg de lait | 25 1/jour (à 3 mois) |
| 3. A volonté mélange de téguments de graines de coton (46%); mélasse (17%); farine de graines de coton (18 %); farine de graines de sésame (15 %); lécithine brute (4 %); et 10 kg d'herbe kikuyu fraîchement coupée (2 kg MS par jour) | 6.200 kg/300 jours (vaches avec 2ème veau) 5.700 kg/300 jours (vaches avec ler veau) | |
| 4. Sommités de cannes (50 % de plus que l'apport journalier plus herbe fraîchement coupée selon disponibilité | Farine de graines de coton 250 g/kg de lait | 2.800 kg/lactation/an(4) |
Références:
1) NDDB Anand.
2) Observations personnelles - dans le système villageois un animal seulement.
3) C.E. Payon, V. (communication personnelle).
4) Boodoo et al., 1988.
Dans beaucoup de pays en développement, les protéines pour le bétail sont rares. Des protéines hautement solubles et aisément dégradables sont parfois disponibles (par exemple le lupin et les graines de coton complètes), mais leur efficacité pour modifier le rapport P/E ne dépasse pas celle d'un complément d'urée. Il est grandement nécessaire de mettre au point des techniques bon marché pour la protection des farines protéiques.
Les protéines peuvent être protégées contre la décomposition dans la panse par des procédés d'insolubilisation, à savoir:
un traitement thermique (selon l'origine des protéines); les températures se situant entre 120 et 180°C dénatureront la protéine et l'insolubilité qui en résulte confère à celle-ci une protection non négligeable contre les atteintes microbiennes quand elle est dans la panse;
des réactions chimiques:
des réactions avec des aldéhydes tels que le formaldéhyde et le glutéraldéhyde, ou
le mélange avec des tanins ou la sélection de plantes riches en tanins.
une faible application de chaleur avec la farine protéique et un ose; il se produit alors une légère réaction de brunissement (voir Lewis et al., 1988).
la protection des protéines par enrobage avec un polymère résistant ou un savon calcique à acides gras à chaîne longue.
Technologies les plus probables
Il est hautement probable qu'à l'avenir pour traiter les protéines en vue de garantir qu'elles contournent la panse, la technologie alimentaire aura recours à la “réaction de brunissement” ou à l'emploi de savons calciques à acides gras à chaîne longue (notamment pour les vaches laitières où les acides gras à chaîne longue peuvent être utilisés dans la synthèse du lait et où la ration est pauvre en lipides) pour enrober les protéines.
Il est certain que l'emploi d'aldéhydes pour protéger les protéines a donné des résultats satisfaisants, mais cette méthode est souvent peu économique et elle n'est pas aisément appliquée dans la plupart des pays en développement. On y a renoncé progressivement dans certains pays en raison de l'effet potentiellement cancérogène d'un composé volatil qui est produit lors de la réaction entre le formaldéhyde et les acides.
Depuis quelques années, il s'est révélé possible de mettre au point un traitement léger des farines protéiques au xylose (un pentose) à basse température. Il ressort clairement des résultats obtenus par le Professeur T. Klopfenstein dans son laboratoire de l'Université du Nébraska (Lewis et al., 1988) que de faibles quantités de xylose chauffées avec de la farine de soja évitent nettement aux protéines de se décomposer dans la panse et améliorent l'efficacité de l'utilisation des aliments chez les bovins nourris de fourrage et de concentrés.
Cette technologie a été sélectionnée parce qu'elle est simple, que le processus offre de grandes possibilités de perfectionnement par des scientifiques qualifiés et que les sources de xylose sont aisément accessibles dans la plupart des pays en développement.
Cependant, avant d'appliquer ce système il faut mettre au point des techniques de production du xylose. Le xylose est l'un des composants majeurs de l'hémicellulose et, parmi les sources utiles de xylose, figurent la bagasse de canne à sucre et les téguments de graines de coton. Le xylose peut être libéré dans un mélange de sucres par simple hydrolyse de l'acide ou par application de vapeur sous pression. Le xylose est également présent en quantités non négligeables dans la liqueur noire (par exemple la liqueur de sulfite) issue du blanchiment à l'acide dans les papeteries. Quand le blanchiment se fait à l'alcali, les sucres sont probablement détruits et la liqueur noire n'en contient pas. Malheureusement, c'est ce second procédé qui est le plus souvent utilisé dans les pays en développement.
Les méthodes de protection des protéines doivent être un domaine de recherche prioritaire dans les pays où les protéines contenues dans les farines disponibles sont hautement solubles, par exemple où l'on utilise des tourteaux d'oléagineux non transformés, des graines comme celles du lupin, des tourteaux d'oléagineux extraits sous pression, des fourrages ou du feuilage.
D'autre part, l'application de la chaleur élimine les facteurs qui nuisent à la qualité et élargit donc la base fourragère protéique susceptible d'être utilisée (par exemple la graine de canavalia peut devenir plus utilisable). Le filage semble être un système de fabrication susceptible d'être employé à l'avenir.
Utilisation de banques de protéines comme source de protéines de dérivation
La création de “banques de protéines” qui offrent aux bovins au pacage une alimentation stratégique de complément quand les pâturages sont secs et pauvres en protéines représente un perfectionnement important dans certains pays où les vastes terrains de parcours sont une source non négligeable de produits d'origine animale. Ces banques de protéines constituent une source aisément accessible d'azote et de minéraux pour les micro-organismes de la panse et l'animal. La stratégie consiste à semer une légumineuse riche en protéines sur une certaine surface d'un terrain de parcours et d'y laisser paître les ruminants pendant la saison sèche.
Il est hors de doute que l'incorporation de petites quantités de légumineuses dans une ration composée d'herbage de qualité médiocre est bénéfique car elle fournit des minéraux et de l'azote aux micro-organismes de la panse et stimule la digestibilité du fourrage de mauvaise qualité qui constitue le gros du régime. Toutefois, il est peu probable qu'un tel complément modifie le rapport P/E, si ce n'est par son effet direct sur le milieu ambiant dans la panse, de sorte que sa valeur ne dépasse guère celle d'une pierre à lécher contenant de la mélasse et de l'urée. Une manièrer beaucoup plus efficace d'utiliser ces banques de protéines consisterait à récolter et protéger les protéines et à développer éventuellement d'autres sources d'azote non protéique, telles que les pierres à lécher contenant de la mélasse et de l'urée, pour les ajouter à ces protéines. D'un autre côté, une légumineuse contenant des tanins pourrait fournir une quantité non négligeable de protéines de dérivation et une pierre à lécher contenant de la mélasse et de l'urée serait alors un véritable aliment de complément.
Les banques de protéines peuvent se composer de légumineuses fourragères, de feuilles d'arbres, de graines ou de gousses provenant des cultures ou d'arbres (par exemple Enterolobium, Prosopis et Leucaena) ou de toute autre phylloprotéine.
Il est nécessaire de mettre au point des techniques appropriées pour tirer profit de ces sources de protéines dans les pays où les protéines sont coûteuses ou inexistantes. Peut-être faudrait-il commencer par des travaux de botanique et d'agronomie pour déterminer les plantes qui conviennent puis mettre au point la méthode de récolte correcte, suivie de techniques pour protéger la protéine.
Les arbres comme source potentielle de protéines de dérivation
On a grandement sous-estimé le potentiel des feuilles d'arbres, des graines d'arbres et des gousses. Il a été démontré, par exemple, que des arbres Prosopis juliflora parvenus à maturité produisent de 400 à 660 kg de gousses par arbre et par dans les régions où la pluviométrie annuelle est de 400 mm (fourchette: 200–600 mm) (F. Riveros, communication personnelle), avec 16 à 20% de protéine brute. Beaucoup de terrains de parcours dans les régions semi-arides produisent moins de 2.500 kg par an de biomasse herbacée, dont 200 à 400 kg seulement sont récoltés par les bovins (voir Ellis et Swift, 1988). Par conséquent, quelques arbres à l'hectare n'auront guère d'effets sur la biomasse herbacée disponible, mais en revanche un effet énorme sur la biomasse globale disponible et, de plus, ils constitueront une source d'azote soluble et offriront l'option de produire de la farine contenant des protéines de dérivation.
Les arbres cultivés comme source de protéines représentent un potentiel énorme sur les terrains de parcours. Les advantages des plantations de Prosopis spp. sont évidents et ont été démontrés (voir Riveros, 1988). Il est nécessaire de concilier cette approche avec les progrès récemment accomplis en ce qui concerne l'alimentation protéique des ruminants et de mettre au point:
des techniques de récolte pour les gousses et les graines;
des techniques de séchage, de broyage et de protection de la protéine (s'il y a lieu);
des techniques pour concentrer les protéines et les protéger;
des méthodes de commercialisation du fourrage;
des travaux de recherche pour déterminer la réaction du bétail à l'utilisation de ces fourrages.
Il existe un grand nombre d'arbres dotés d'attributs potentiellement analogues; toutefois, dans les zones semi-arides la préférence devrait être donnée à l'emploi de Prosopis comme source de protéines.
Le fractionnement de la gousse pour produire un composé riche en protéines serait une première étape utile avant de mettre au point une technique de protection.
Il convient de signaler que, dans de nombreuses régions, Prosopis est considéré comme une plante adventice potentiellement dangereuse. Dans ces régions, il ne doit pas être brouté pendant la chute des grousses car les graines seraient propagées dans la bouse des bovines et l'arbre prendrait ainsi racine sur de vastes superficies. Cependant, avec un système de plantation, il serait aisé de remédier à ce problème.
Possibilités d'utiliser la biotechnologie végétale
La biotechnologie végétale pourrait se révéler très importante pour utiliser les arbres comme composants du fourrage; du fait que la production de semences est très variable parmi les arbres, le clonage et la culture tissulaire des jeunes plants provenant de variétés à haut rendement représenteraient un progrès notable. Cependant, les essais tentés avec le cocotier et le palmier à huile n'ont about qu'à une inflorescence médiocre ou nulle chez les sujects produits par culture tissulaire: il faut donc procéder avec prudence.
Prosopis spp. n'est cité ici qu'à titre d'exemple et il existe à cette fin beaucoup d'autres sources potentielles de protéines qui ne sont encore qu'insuffisamment utilisées.
Le développement des légumineuses comportant une proportion appropriée de tanins est un objectif valable puisque Barry et ses collaborateurs ont montré que les feuilles de Lotus sp. (3 à 6% de tanins) contiennent apparement plus de protéines de dérivation que des espèces semblables sans tanins (Barry et Blaney, 1987). Les recherches à ce sujet pourraient être poursuivies dans le contexte de la banque de légumineuses. Là encore, le problème pourrait être résolu par expression génique puisqu'un excédent de tanins est tout aussi désavantageux qu'une insuffisance. Il arrive souvent que la teneur en tanins soit directement liée au type de sol et à la pluviosité (G. Blair, communication personnelle).
Toute carence d'un nutriment quelconque dans la panse affectera d'abord cet écosystème en réduisant l'efficacité de la croissance microblienne; à mesure que la carence revêt plus d'acuité, la dimension du réservoir microbien diminuera à la longue et il en sera inévitablement de même pour la digestibilité et l'absorption des fourrages grossiers par les ruminants. La diminution de la digestibilité n'apparaîtra qu'au bout d'un certain temps à mesure que la carence du nutriment s'accroît progressivement. L'administration de nutriments de complément pour la panse à un animal accusant une carence critique augmentera l'efficacité de la croissance microbienne, la dimension du réservoir microbien et la digestibilité du fourrage, ce qui aure presque toujours pour conséquence un accroissement de l'apport fourrager. Toutefois, ce dernier effet dépend souvent du degré de carence du nutriment limitant.
Les nutriments dont il y aura le plus souvent carence sont l'azote non protéique, le phosphore, le soufre, le magnésium, le sodium et des oligo-éléments tels que le cobalt, le cuivre et le zinc. Une alimentation de complément comportant ces différents nutriments réussit généralement à accroître l'apport fourrager et la digestibilité, mais c'est l'association de ces compléments avec une source de protéines de dérivation qui permettra d'augmenter la productivité dans de fortes proportions.
Sources de nutriments minéraux et pierres à lécher composées de mélasse et d'urée
Dans les pays en développement, un suc végétal concentré tel que la mélasse ou le résidu de l'huile de palme constitue l'une des meilleures sources d'oligo-éléments minéraux. Les pierres à lécher comportant plusieurs nutriments n'ont pour seul avantage que le mélange de substances qui les composent. Elles sont utiles parce qu'elles constituent un module facile à manipuler, à commercialiser et à utiliser par l'éleveur sans que cela lui prenne du temps sur ses autres activités. Ces pierres à lécher posent surtout un problème dans les régions d'herbages où il faut trouver un moyen d'encourager le bétail à les consommer en quantités suffisantes. Ces régions sont souvent dépourvues de mélasse, ou bien ce produit coûte trop cher. Des travaux de recherche plus poussés sont nécessaires pour produire pour ces régions des modules appropriés (par exemple des pierres à lécher sans mélasse).
L'administration d'une alimentation de complément sous forme de pierres à lécher est surtout importante parce qu'elle donne accès au bétail des petits éleveurs. Peut-être sera-t-il possible à l'avenir d'y incorporer des innovations, par exemple en introduisant dans les pierres des produits chimiques et d'autres substances manipulant la panse (par exemple des produits antiprotozoaires) ou manipulant l'animal (par exemple des produits chimiques gastrorésistants qui sont absorbés à partir de l'intestin), ou même combattant les maladies et les parasitoses (anthelminthiques). A ce propos, la production et l'essai de pierres à lécher contenant des anthelminthiques sont tout à fait appropriés et sont actuellement enterpris par l'Organisation scientifique et industrielle du Commonwealth en collaboration avec le Centre australien pour la recherche agronomique internationale (ACIAR).
C'est un domaine qui a fait l'objet d'études approfondies et a suscité beaucoup d'espérances. Le principe est simple: si l'on parvient à mettre au point des microbes de la panse doués d'une meilleure aptitude à décomposer les fibres ou leurs composantes, ces microbes pourront extraire du fourrage consommé une plus forte proportion des nutriments disponibles.
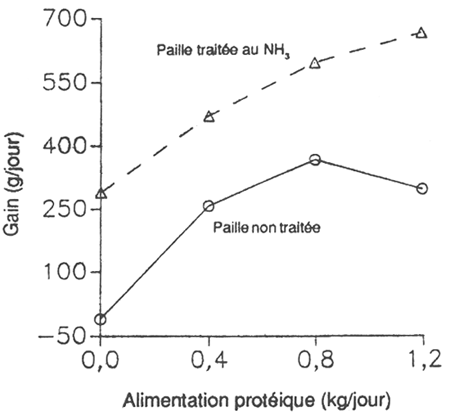
Cette filière de recherche bénéficie souvent d'une haute priorité parce qu'on pense que les animaux élevés sur des pâturages de qualité médiocre ont avant tout une carence énergétique. En fait, le plus souvent ces animaux utilisent les nutriments disponibles d'une manière peu efficace. Il convient de souligner que la première des priorités est une alimentation équilibrée et, pour éviter toute autre perturbation du système, l'animal doit avoir une panse et un métabolisme efficaces. Chez un animal nourri de fourrage grossier mais dont la panse est efficace et l'alimentation protéique équilibrée, il est possible que pour toute augmentation de cinq unités de la digestibilité du fourrage grossier le gain de poids vif s'accroisse de 50% avec la même ressource fourragère de base (voir figure 5.1, Perdok et Leng, 1990).
Silva et Ørskov (1988b) ont démontré que le degré de décomposition d'un fourrage fibreux dépend du milieu ambiant dans la panse. De la paille broyée non traitée placée dans des sacs en nylon dans la panse d'ovins nourris de paille ammoniacale fermentait davantage que la même paille broyée non traitée placée dans la panse d'ovins nourris de paille non traitée. Il se pourrait que cette différence soit due à une plus forte concentration d'enzymes dans la panse des ovins nourris de paille ammoniacale par rapport à la paille non traitée.
Ces deux observations, considérées ensemble, indiquent la possibilité d'améliorer considérablement la productivité en introduisant dans la panse un micro-organisme génétiquement manipulé en vue de produire davantage d'enzymes pour la digestion des fibres ou des enzymes d'une plus grande diversité.
Il existe en principe trois moyens d'accroître l'aptitude des microbes de la panse à décomposer les fourrages fibreux:
sélectionner des microbes doués d'une forte aptitude à sécréter des enzymes fibrolytiques ou qui sécrètent des enzymes fibrolytiques ayant une forte activité spécifique;
créer des microbes dotés d'un plus large spectre de sécrétions enzymatiques grâce à la technologie de l'ADN recombinant, par exemple en dotant des microorganismes xylolytiques d'aptitudes cellulolytiques et inversement;
créer des microbes doués d'une plus forte activité fibrolytique grâce à la technologie de l'ADN recombinant.
Dans tous les cas, la condition préalable est que ces micro-organismes soient capables de se développer dans la panse et d'y conserver leur espace. Cette condition préalable s'applique également à la production par les micro-organismes de la panse, grâce aux techniques de génie génétique, de protéines d'une grande valeur biologique, d'autres sécrétions de protéines, d'acides aminés et de peptides (voir exposé plus haut).
Il existe un certain nombre de moyens bien connus d'incorporer le codage de l'ADN pour certaines enzymes dans les bactéries anaérobies grâce à la technologie de l'ADN recombinant. La démarche suivie est partout la même et l'on en trouvera la description à l'annexe A; il suffira de signaler l'incertitude de ces travaux de recherche et leurs complications.
Les progrès ont été relativement rapides, quatre groupes de chercheurs au moins ayant déjà réussi à incorporer un ADN étranger dans des microbes de la panse. Presque toutes ces transformations ont été obtenues par intégration de plasmides. Aucun des microorganismes ainsi manipulés n'a été réintroduit dans la panse pour en vérifier la stabilité et dans un cas seulement le microbe traité avait été récemment isolé de la panse. Le fait qu'on ait utilisé des souches de laboratoire de micro-organismes de la panse ayant nettement subi des mutations pendant leur culture montre clairement qu'il s'agit de travaux de recherche fondamentale dont l'objet principal était la transformation et non l'amélioration de la digestibilité qui pourrait en résulter.
L'un des obstacles majeurs était l'absence de systèmes de surveillance appropriés pour repérer dans la panse le micro-organisme manipulé. L'emploi de sondes d'ADN n'apparaît pas très satisfaisant en raison de la diversité génétique des souches bactériennes à l'intérieur d'une même espèce (Hudman et Gregg, 1989).
L'exposé qui précède a pour but de souligner que des travaux de recherche fondamentale très poussés s'imposent avant qu'on puisse mettre au point des microorganismes modifiés capables de survivre dans la panse. Il faut également insister sur le fait qu'il s'écoulera probablement un long délai entre le début des travaux, lorsqu'on ne disposera pratiquement d'aucune information, et le stade de l'application.
Les structures filamenteuses des champignons anaérobies de la panse pénètrent dans les membranes des cellules ligneuses des fourrages dans la panse et les affaiblissent onsidérablement, alors que les bactéries n'adhèrent guère à ces tissus végétaux extrêmement résistants, pas plus qu'elles ne les décomposent. A la différence des bactéries, les champignons de la panse percent la cuticule qui protège la surface de la feuille. Des travaux de recherche ont montré que ces champignons possèdent des enzymes particulières, ou des enzymes dotées d'une plus forte activité spécifique, qui les rendent capables d'affaiblir et de détruire les obstacles structurels qui s'opposent le plus à la décomposition.
Il ressort de ces observations que les recherches visant à développer la biomasse fongique dans la panse ou à y établir des champignons extrêmement actifs (par exemple par sélection) représenteraient un grand pas en avant pour accroître la digestibilité des fourrages de qualité médiocre chez les ruminants. L'identification des souches de champignons anaérobies qui décomposent le plus rapidement les membranes des cellules des plantes fourragères est la condition préalable de toute inoculation réussie de champignons dans la panse qui soit profitable pour l'animal sur le plan nutritionnel.
Il y a tout intérêt à développer les filières de recherche axées sur la culture, la sélection des souches et l'identification de nouvelles espèces de champignons anaérobies et sur la vérification de leur aptitude à affaiblir les fibres végétales et à améliorer leur digestion.
Il se peut que les pays en développement soient bien placés pour entreprendre et développer les recherches dans ce sens puisqu'ils disposent d'animaux vivant dans des conditions d'alimentation extrêmes, qui ont subi une longue période d'adaptation et qui ont donc peut-être déjà sélectionné des champignons doués d'une forte activité fibrolytique.
Les travaux de recherche doivent comporter des aspects qui permettent aux souches sélectionnées de se multiplier dans la panse et de résister à la concurrence des souches sauvages. Etant donné que les champignons de la panse sont propagés par des spores résistantes (Ho et al., 1990) qui sont expulsées dans les fèces et qui, une fois absorbés par l'animal dans le fourrage, se multiplient par sporulation dans la panse, le développement d'espèces spécifiques pour assurer la survie est un axe de recherche qui doit bénéficier d'une haute priorité.
Les critères de sélection comprendront la détermination du taux de solubilisation des glucides fibreux par les différents champignons et l'affaiblissement des tiges des plantes fourragères après une période d'incubation avec les fourrages.
La sélection de souches bactériennes douées d'une meilleure activité cellulolytique semble être un domaine de recherche moins vital car ce sont les champignons qui jouent un rôle relativement plus important dans la décomposition des éléments les plus résistants des fourrages, en particulier des fourrages de qualité médiocre.
La lignine se trouve dans les membranes cellulaires des plantes fourragères où elle est étroitement associée à l'hémicellulose, formant une matrice qui entoure les microfibrilles de cellulose. Il existe aussi de fortes concentrations de lignine dans la lamelle médiane entre les fibres où elle sert de liant pour les cellules contiguës. Les lignines polymères ne sont décomposées par aucun micro-organisme anaérobie connu.
Le Basidiomycetes, champignon aérobie de la pourriture blanche, décompose la lignine plus rapidement et d'une manière plus poussée que toute autre espèce microbienne étudiée. Il est généralement admis que la délignification d'une ligno-cellulose augmentera le taux d'atteinte des composés cellulose/hémicellulose quand ils pénètrent dans la panse.
En aérobiose, il existe deux mécanismes de délignification biologique, à savoir:
la minéralisation, la lignine étant alors convertie directement en dioxyde de carbone;
la solubilisation, la lignine étant alors libérée (solubilisée) à partir de la lignocellulose, jointe à une quantité variable d'hémicellulose (voir Paterson, 1989).
La solubilisation de la lignine des graminées est particulièrement importante pour sa décomposition dans les sols. Il semble se produire dans la panse un phénomène assez proche de la solubilisation car une quantité assez considérable de lignine est précipitée dans la caillette et, par conséquent, est soluble dans le suc de la panse (cette lignine est souvent appelée lignine polymère à précipitation dans l'acide). Quand elles sont présentes dans les micro-organismes de la panse, les enzymes de solubilisation de la lignine peuvent être renforcées par des moyens génétiques ou par sélection.
Si elles ne sont pas présentes dans les micro-organismes de la panse, la possibilité de produire des enzymes solubilisant la lignine serait un moyen important d'améliorer la digestibilité des fibres dans l'écosystème anaérobie de la panse.
Pour résister aux atteintes des insectes, des champignons et des bactéries, les plantes produisent des composés secondaires qui empêchent ces organismes de coloniser les tissus foliaires. Pour empêcher les lésions des feuilles dues au broutement, certains arbres et arbustes produisent des quantités plus fortes de composés secondaires qui les rendent souvent incomestibles. Il a été démontré récemment que le broutement d'un acacia déclenchait chez ce dernier la libération d'éthylène qui, en quelques minutes, poussait les acacias voisins dans le même bosquet à commencer à synthétiser des tanins qui incitaient les cervidés qui les broutaient à cesser de consommer les feuilles de ces arbres.
Beaucoup de composés secondaires des végétaux sont toxiques pour les microbes de la panse ou directement ou indirectement toxiques pour l'animal par suite du métabolisme microbien dans la panse. Ce phénomène limite la base fourragère des ruminants et empêche parfois l'élevage sur certains terrains. Les caprins, et probablement d'autres ruminants aussi, sont capables de détoxifier beaucoup de ces composés secondaires des végétaux dans le foie.
Les travaux de recherche portant sur la possibilité de détoxifier les composés secondaires des végétaux ont connu un nouvel essor quand on a découvert que des micro-organismes dans la panse de caprins à Hawaï étaient capables de dégrader le produit de décomposition de la mimosine (c'est-à-dire la dihydroxypyridine) dans Leucaena Leucocephala, mais des ruminants en Australie ne pouvaient pas le faire parce qu'ils n'hébergeaient plus dans la panse le microbe porteur des enzymes nécessaires (Jones, 1981). Le microbe isolé dans la panse des caprins de Hawaï dégradait la dihydroxypyridine, produit toxique de décomposition de la mimosine (voir Hegarty, 1982). L'introduction dans les ruminants australiens du microbe provenant des caprins de Hawaï a supprimé les symptômes de la toxicité due à la mimosine chez le bétail précédemment sensible.
Cette découverte a permis d'envisager la possibilité de conférer aux microorganismes de la panse, par des techniques de génie génétique, l'aptitude à dégrader ces composés toxiques. Des recherches sont en cours à la University of New England pour tenter d'introduire dans des bactéries des gènes favorisant la défluoration du fluoroacétate, composé mortel présent dans Acacia gidyea et qui tue souvent les bovins en Australie septentrionale quand cette plante est broutée pendant la saison sèche (voir Barry et Blaney, 1987).
La mutation réussie de bactéries de la panse avec un gène unique permettant la défluoration du fluoroacétate ouvrirait la perspective d'utiliser tout un éventail d'autres végétaux, soit des plantes adventices (par exemple la lantana), soit de gros producteurs potentiels de biomasse (quelques légumineuses dont certaines arborescentes), soit encore des cultures riches en protéines (par exemple la canavalia).
Les problèmes sont partout les mêmes. Le premier consiste à identifier l'ADN bactérien codé pour l'enzyme qui convient, après quoi il faut mettre au point des méthodes pour introduire cette enzyme dans une bactérie appropriée et stabiliser l'ADN dans la cellule, le dernier problème étant de déterminer si le micro-organisme modifié pourra survivre dans la panse et sera capable d'exprimer le gène.
Les recherches devront viser principalement à découvrir, isoler et traiter l'ADN codé pour l'enzyme appropriée qui doit être capable d'être exprimée et de fonctionner aisément dans les micro-organismes de l'écosystème de la panse.
Si l'on confère aux ruminants l'aptitude à dégrader des composés toxiques, cela peut avoir des conséquences dangereuses en ce sens que ces composés constituent souvent l'ultime moyen de défense des végétaux consommés, de sorte que dans les régions arides cela pourrait favoriser le déboisement et l'érosion du sol.
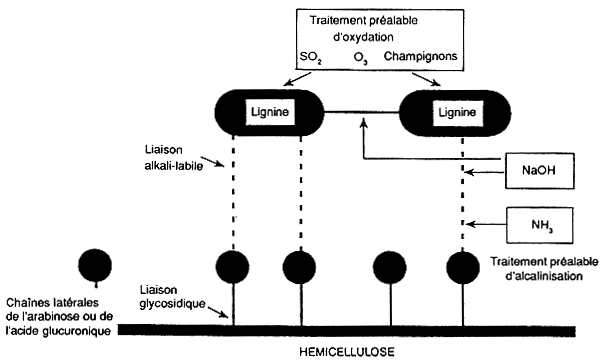
Il existe un certain nombre de procédés d'hydrolyse des substances acides et alcalines qui solubiliseront la lignine, provoqueront des perturbations dans les fibres par gonflement ou amélioreront par d'autres moyens la digestibilité potentielle des fourrages grossiers fibreux. La figure 5.2 indique la localisation de l'hydrolyse de plusieurs produits chimiques dans une fibre qui en améliorent la digestibilité.
Il semble qu'on dispose d'informations suffisantes sur les techniques de traitement et sur les avantages qui en résultent pour les fourrages de qualité médiocre (voir Sundstol et Owen, 1984). Il n'y a donc pas beaucoup d'incitation à poursuivre la recherche d'autres méthodes. Bien qu'une démarche novatrice puisse justifier parfois de nouvelles recherches, on peut affirmer pour l'heure que le traitement qui est probablement le mieux susceptible d'être appliqué dans les pays en développement est l'hydrolyse alcaline de la paille mouillée par l'ammoniac produit par l'urée (généralement 4% de la matière sèche) provenant des microbes dans la paille. Il devrait aussi être possible de recourir à l'application d'ammoniac anhydre (voir Sundstøl et Owen, 1984).
L'ammoniac dans la paille a un double effet:
il rend la paille mieux fermentescible dans la panse par rupture de certaines des liaisons ligno-hémicellulose;
il fournit de l'azote non protéique pour la croissance des microbes dans la panse quand le fourrage est consommé.
Bien que l'ensilage de la paille à l'urée puisse être économique et efficace, cette technique n'a pas été largement adoptée par les petits exploitants des pays en développement en général; toutefois, quelques rapports signalent que des éleveurs individuels utilisent régulièrement cette technologie.
Il est nécessaire que les chercheurs déterminent les raisons pour lesquelles une telle technologie n'est pas acceptée; ce refus est presque toujours dû à la longue durée de l'amortissement et à des contraintes d'ordre sociologique (voir Sansoucy, 1989).
Depuis quelques années, on a mis l'accent sur une stratégie d'amélioration des cultures fibreuses par l'incorporation de champignons non toxiques dans la paille. On a utilisé en particulier les basidiomycètes, champignons de la pourriture blanche, à cause de leur aptitude à délignifier la matière végétale. Le principal problème qui a surgi est que cette technologie, quoique relativement simple, est plus complexe que celle du traitement des fourrages grossiers à l'urée et, bien que des protéines soient produites et que les résidus soient plus digestibles, une proportion non négligeable de la biomasse totale est perdue, notamment quand on laisse les basidiomycètes passer au stade de formation des champignons. On peut éviter la perte de biomasse dans une certaine mesure en diminuant la durée d'incubation de la paille et des fourrages; la perte de biomasse est alors ramenée à moins de 10% (voir Rai et al., 1989).
Le Dr B.N. Gupta et ses collaborateurs de l'Institut national de recherche laitière (NDRI) de Karnal (Inde) ont mis au point un procédé à deux étapes utilisant un champignon coprin (Coprinus fimetorius). Cette technologie à deux étapes comporte 1) un traitement à l'alcali (ensilage à l'urée) et 2) une deuxième période d'incubation pendant laquelle les champignons sont cultivés sur la paille. Cette technique est relativement simple, encore que les deux étapes la rendent plus complexe que le simple traitement à l'urée quand elle doit être appliquée par des petits éleveurs.
Cependant, les traitements chimiques et microbiologiques peuvent être très efficaces quand il faut emmagasiner de grandes quantités de fourrage et les transférer dans des régions où la sécheresse a provoqué une pénurie. Là encore, certaines précautions s'imposent car, pour que ces fourrages soient profitables au maximum, il faudra donner aux bovins un complément de protéines de dérivation (voir Leng, 1986).
Les caprins par rapport aux ovins et les buffles par rapport aux bovins ont tendance à retenir les matières fibreuses plus longtemps dans la panse et à en extraire par la même occasion quelques unités de plus du point de vue de la digestibilité. A cet égard, les caprins et les buffles recyclent vers la panse une plus grande quantité d'azote d'urée. De même, les bovins Bos indicus recyclent vers la panse plus d'azote d'urée que les bovins Bos taurus (voir Leng, 1990). Par conséquent, les mécanismes qui ralentissent le renouvellement dans la panse, accroissent le volume de la panse et permettent de recycler davantage d'azote d'urée améliorent l'utilisation des fourrages de qualité médiocre.
Il a été prouvé récemment que l'immunisation contre le facteur d'inhibition de la libération de somatotropine accroît le débit à travers l'intestin, d'où une meilleure digestibilité et des disponibilités de protéines plus abondantes pour les ovins nourris de concentrés et de fourrages (Sun et al., 1990). Il serait extrêmement intéressant d'observer les effets de l'immunisation contre ce facteur chez les bovins dont la ration est composée de fourrage et de protéines de dérivation. Toutefois, la technologie ne peut pas être appliquée aisément par les éleveurs villageois en raison du nombre d'injections actuellement nécessaires pour assurer l'immunité.
Il existe tout un éventail de composés injectables qui permettront au bétail d'utiliser les nutriments d'une manière plus efficace, notamment en répartissant les nutriments en faveur de la production de viande maigre ou la production de lait. Ces composés sont les stéroïdes anaboliques, les hormones de croissance et d'autres facteurs hormonaux de croissance, et les bêta-agonistes, outre des techniques telles que la neutralisation des hormones par immunisation. En règle générale, ces composés agissent par manipulation de l'état hormonal; ils améliorent la qualité de la viande maigre dans la carcasse et permettent une conversion plus efficace du fourrage en produits finals. Le lecteur trouvera un exposé récent sur la question dans la publication de Buttery et Dawson (1988).
Les stéroïdes naturels sécrétés par les glandes génitales sont utilisés pour favoriser la croissance et réduire le rapport graisses/protéines dans la viande dans de nombreux pays du monde, mais leur emploi a été récemment interdit en Europe. L'hormone injectée modifie l'influence des hormones de sécrétion endogène (les injections d'oestradiol chez les bovins provoquent habituellement un relèvement du taux sérique des hormones de croissance) et les hormones de croissance sont liées plus étroitement aux sites dans le foie.
Comme c'est habituellement le cas, il existe une interaction très nette avec la nutrition et, avec les régimes mal équilibrés en général, l'effet des hormones exogènes est moins évident que lorsqu'une ration d'ensilage, par exemple, est équilibrée par une protéine de dérivation (farine de poisson) (voir Gill et al., 1987).
Ces observations mettent de nouveau en lumière les différences qui existent et, par conséquent, la nécessité d'une nouvelle approche de la biotechnologie dans les pays en développement où le régime alimentaire des ruminants ne contient habituellement que peu de protéines vraies.
Grâce aux progrès récents de la technologie de la fermentation ainsi que de la technologie de l'ADN recombinant, il est désormais possible de produire de grandes quantités de protéines peptidiques qui semblent être analogues aux hormones de croissance bovines ou porcines (BSt. ou PSt.). Injectées dans les animaux, ces hormones stimulent la croissance et réduisent de façon spectaculaire la teneur en graisses de la carcasse (voir Buttery et Dawson, 1988). De plus, elles stimulent la production de lait de 20 à 25% chez le cheptel laitier.
Lors des études initiales sur l'injection d'hormones de croissance, l'augmentation de la production de lait semblait résulter d'une nouvelle répartition des nutriments et de la mobilisation des nutriments à partir des dépôts tissulaires (voir McCutcheon et Bauman, 1985); plus récemment, on a signalé un effet à long terme sur l'apport de fourrage (Chilliard, 1988). Par ailleurs, la stimulation de la production laitière dépendait des conditions d'élevage du cheptel et elle ne se manifestait pas dans les troupeaux exploités de façon médiocre (Bauman, D., 1990, communication personnelle). En Italie et en Inde, des hormones de croissance exogènes ont permis d'accroître de 25% le rendement laitier de bufflonnes élevées dans de bonnes conditions (voir, par exemple, Ferrara et al., 1989), mais il est possible que cette application soit limitée par le coût des injections.
Administrés par voie buccale, les bêta-agonistes augmentent le dépôt de viande maigre et réduisent la teneur en graisses de la carcasse chez les porcins et les ruminants (Hanrahan, 1987), mais le gain de poids vif est assez faible. Là encore, il est hautement probable que la réaction dépend de la nutrition de l'animal.
On observe de très fortes similitudes entre la réaction à l'administration d'hormones de croissance, de bêta-agonistes, etc. et la réaction aux mesures visant à corriger le rapport P/E dans les nutriments absorbés. Une communication au moins signale une interaction entre l'administration de protéines de dérivation et celle d'hormones exogènes (voir Gill et al., 1987). Il est donc possible que l'optimalisation du rapport P/E dans les nutriments absorbés déclenche une libération d'hormones équivalant à l'injection d'une hormone de croissance. Réciproquement, et si tel est le cas, cela signifie que les effets des hormones exogènes ne seront peut-être pas si évidents quand le rapport P/E dans les nutriments absorbés est optimalisé. En outre, chez les animaux issus d'une mutation génique qui expriment une hormone de croissance, les besoins de protéines par rapport aux besoins énergétiques seront considérablement accrus, de même que les besoins de minéraux et de vitamines.
La conclusion importante de la communication de Buttery et Dawson (1988) est la suivante:
“La technologie actuelle ouvre de nombreuses perspectives très stimulantes, mais notre connaissance des principaux processus métaboliques associés à la croissance comporte encore de grandes lacunes qu'il faudra combler avant de pouvoir mettre au point une méthode de promotion de la croissance pouvant être partout acceptée”.
Cette réserve est constamment formulée par les spécialistes de la biotechnologie et elle traduit la nécessité de poursuivre en permanence les travaux de recherche fondamentale portant sur les mécanismes du métabolisme.
De l'avis de maints spécialistes scientifiques, la sélection effectuée parmi les différentes races laitières est trop lente pour aboutir aux fortes augmentations de la production de lait qui sont nécessaires pour faire face à l'expansion démographique et à l'accroissement de la demande dans les pays en développement. Il est partout admis que la voie à suivre est le croisement d'animaux de race indigène avec des vaches laitières à haut rendement provenant des pays à climat tempéré. On estime que la vache laitière à haut rendement est capable d'utiliser les nutriments plus efficacement que les animaux indigènes en vue de la production de lait. Ce principe s'est révélé exact quand les conditions d'exploitation sont optimales, l'alimentation bien équilibrée et le fourrage extrêmement digestible. Pour que la production laitière soit efficace chez les bovins des régions tropicales, il est absolument indispensable de combattre la maladie et de compléter les fourrages de mauvaise qualité afin d'équilibrer la nutrition, faute de quoi le bas taux de reproduction, joint à un taux de mortalité élevé, réduisent souvent à néant les avantages du croisement.
Cependant, l'expérience récemment acquise en Inde donne à penser que tel n'est pas nécessairement le scénario du croisement.
Etude de cas (Inde). Le rendement laitier des frisonnes importées à la station du NDDB de Anand (Inde) constitue un exemple frappant de l'amélioration de la production de lait qu'il est possible d'obtenir en appliquant un ensemble de techniques. Ces vaches sont élevées dans un établissement, la maladie est parfaitement circonscrite et la nourriture est constituée de fourrage frais (jusqu'à 40 kg par jour) complété par de l'ensilage de maïs et de la paille de riz pendant la saison sèche, plus 350 g d'un concentré protéique de dérivation (30% de CP) pour chaque litre de lait produit. Le rendement moyen des vaches en première lactation était supérieur à 5.000 litres pendant 305 jours, certaines bêtes exceptionnelles en fournissant même 6.500. Ces vaches en sont maintenant toutes à leur deuxième lactation, avec les indices d'un rendement laitier moyen supérieur à 6.000 litres. On a pu obtenir ce niveau de production en utilisant des pierres à lécher composées de mélasse et d'urée et une alimen-tation de complément comprenant des protéines de dérivation. Le seul concentré céréalier, qui représente 10% du mélange de protéines de dérivation, a pour but d'assurer au stade de la fabrication la formation d'agglomérés satisfaisants. Si nous citons cet exemple, c'est parce qu'il indique que même dans les régions climatiques les plus difficiles d'Asie, où la tempé-rature dépasse souvent 40°C et atteint parfois 50°C, les frisonnes peuvent produire un volume de lait correspndant à au moins les deux tiers de leur capacité génétique. Il met aussi en évidence un principe très important sur lequel il convient d'insister dans le présent rapport, à savoir que l'amélioration génétique n'est pas une panacée pour l'élevage dans les pays en développement et que les programmes de sélection ne seront couronnés de succès que si l'on parvient à optimaliser l'état nutritionnel et la lutte contre les maladies (voir tableau 5.3).
Bien que le transfert d'embryons n'ait que peu d'importance dans la manipulation nutritionnelle, c'est une technologie qui s'impose comme précurseur de la production d'animaux issus d'une mutation génique.
La mise au point de techniques pour isoler, manipuler et transférer des brins bien définis d'ADN dans des embryons du premier stade élimine l'obstacle qui aurait pu s'opposer au transfert d'informations génétiques entre des organismes vivants totalement différents. Par exemple, l'ADN codé pour des enzymes qui sont normalement produites par des bactéries peut être inséré dans le génome d'un animal et l'on peut s'attendre à ce qu'il exprime la production de ces enzymes chez l'animal après la naissance.
La technologie comporte la digestion et la liaison de sources appropriées d'ADN avec des enzymes spécifiques afin de définir avec exactitude et redisposer des brins d'ADN particuliers, le clonage moléculaire de ces brins dans des bactéries et des phages ou sous forme de plasmides, le séquençage nucléotidique de l'ADN cloné et la technologie de transfert de ce dernier dans les cellules cultivées et les embryons du premier stade.
La recherche sur la mutation génique vise essentiellement à mettre au point de nouveaux génomes (le patrimoine génique total d'un animal) au-delà de l'assortiment aléatoire des gènes lors de la reproduction sexuelle, complétée par la mutation, laquelle est limitée par la variation génétique possible et par des incompatibilités spécifiques. D'autre part, un sujet choisi pour la sélection trasmet souvent à sa descendance certains de ses gènes indésirables. La mutation génique, qui regroupe les techniques de recombinaison de l'ADN, de manipulation de l'embryon et de transfert de l'embryon, offre des possibilités supplémentaires d'élaborer de nouveaux génomes.
Les possibilités d'exploiter utilement de nouveaux génomes chez les animaux ont pour seule limite l'imagination du chercheur ainsi que la définition, l'isolement et l'encapsulation de l'ADN avec des méthodes appropriées pour l'insérer dans l'embryon du premier stade. Comme exemple d'une démarche témoignant d'une grande imagination, on peut citer l'introduction chez des ovins de gènes codés pour la synthèse des acides aminés soufrés, mais, bien qu'on ait produit ainsi des animaux “transgéniques”, la production de laine n'a pas augmenté pour autant et le problème le plus important que soulève cette filière de recherche est l'expression des gènes.
Jusqu' à présent, on a surtout insisté sur la mise au point d'une technologie pour introduire l'ADN en vue de l'expression d'une hormone de croissance. Ces travaux ont été couronnés de succès chez de nombreux types d'animaux. Néanmoins, lors d'une étude récente à ce sujet, le Dr C. Polge (1990, communication personnelle) a conclu que le principal problème auquel se heurte la recherche sur la mutation génique est l'incapacité de contrôler le niveau d'expression d'un gène donné; de plus, le taux de réussite a été très faible en ce qui concerne aussi bien la mutation génique que l'expression du gène. La conclusion générale est qu'il faut en revenir à l'étude des facteurs qui permettent ou contrôlent l'expression des gènes avant que toute percée nouvelle puisse être réalisée. Il faudra en tenir compte lors de toute allocation de crédits dans ce domaine.
Archibald (1989) a récemment conclu un article sur la mutation génique par la déclaration suivante:
“La mutation génique offre de nouvelles possibilités de produire des génomes nouveaux. A court terme, les animaux issus de la mutation génique auront plus de chances de nous faire mieux comprendre comment contrôler les performances par des moyens génétiques que de contribuer à la production agricole.”
Il ressort clairement de cette affirmation qu'à l'heure actuelle la recherche visant à mettre au point des génomes nouveaux chez les bactéries, chez d'autres organismes ou chez des animaux doit viser manifestement l'acquisition de connaissances fondamentales. Cela ne signifie nullement que les travaux en question ne puissent pas avoir quelque application pratique à l'avenir, mais ce pourrait être seulement dans cinquante ou cent ans.
![]()
![]()
![]()