![]()
![]()
![]()
3.1.1.1 - Hypothèses sur les processus et traitements mathématiques
Le modèle global met en relation la capture et le rendement, avec l'effort de pêche exercé sur un stock unitaire. Malgré ses limites, qui tiennent à sa simplicité en matière de données et de traitement, il reste un outil précieux de détermination des productions équilibrées et des efforts correspondant aux objectifs des politiques d'exploitation.
Comme tous les modèles d'évaluation des stocks halieutiques, il doit être appliqué à l'échelle d'un stock, c'est-à-dire à un ensemble discret de populations (une ou plusieurs formant
celui des hypothèses sur les processus dont le modèle est supposé rendre compte: modèle d'estimation sous hypothèse d'équilibre (par exemple Fox de PROFIT), modèle dynamique sans hypothèse d'équilibre (par exemple ASPIC), modèles prenant ou non en compte les fluctuations de l'environnement, modèles globaux enfin prenant en compte des variables biologiques autres que la prise et l'effort (modèle de Deriso);
celui du modèle mathématique supposé représenter la relation fonctionnelle entre l'effort et la production: courbe logistique, exponentielle ou autres.
Ces choix sont importants car d'eux dépendent largement les conclusions sur l'état des stocks.
Faute de temps, le Groupe de travail n'a pu réaliser des analyses comparatives complètes des différents modèles. Le modèle global généralisé a été le plus souvent utilisé. Malgré ses limites théoriques, ce modèle s'avère en effet souple et performant. Son application aux données de pêcheries simulées a souvent montré sa robustesse, c'est-à-dire sa capacité à rendre compte sans biais notables de déséquilibres résultant de modifications rapides de l'effort de pêche, et mê de fluctuations du recrutement ou de la capturabilité. On notera cependant que, si le modèle intègre implicitement les effets des fluctuations passées du recrutement ou de la capturabilité, il ne peut rendre compte des variations futures. Des biais significatifs dans les projections peuvent se produire lorsque les pêcheries subissent des changements ne figurant pas dans l'histoire passée de la pêcherie.
3.1.1.2 - Efforts de pêche nominaux et effectifsDans les pêcheries plurispécifiques, une difficulté sérieuse d'application du modèle réside dans l'estimation d'un effort “effectif” qui corresponde sans biais majeur à la mortalité par pêche exercée sur les différents populations composant un stock composite exploité par plusieurs flottilles. Un effort nominal global exercé sur l'ensemble des populations exploitées par une flottille ou un ensemble de flottilles peut être calculé à partir des données relatives aux navires qui la/les composent et leurs opérations de pêche: somme des temps de pêche des navires dont la puissance de pêche aura été préalablement calibrée par référence à des caractéristiques physiques appropriées. Un tel effort nominal sera commun à l'ensemble des populations, ou des espèces, simultanément exploitées par les flottilles composant la pêcherie considérée
Dans la réalité, les efforts de pêche effectivement exercés sur les différentes populations varient rarement parallèlement à l'effort nominal global, car l'intérêt des flottilles pour les différentes espèces change dans le temps en fonction de l'abondance relative des espèces, de leurs migrations, de leur disponibilité, du prix de vente des espèces, … Les pêcheurs modifient leurs stratégies de pêche (espèces cible) en conséquence. C'est le cas de beaucoup de pêcheries mauritaniennes, et notamment les plus intéressantes économiquement (céphalopodes, crevettes, …) ou les plus importantes en termes physiques (petits pélagiques).
Lorsque l'on dispose de statistiques de pêche très fines (par coups de filet par exemple, comme celles qui peuvent être recueillies par les observateurs), un effort “effectif”, proportionnel à la mortalité par pêche exercé sur chaque espèce ou population, peut être estimé mathématiquement. Les carences statistiques rendent actuellement difficile et insuffisante l'estimation d'efforts effectifs.
S'il pose des problèmes au niveau de l'analyse (mauvaise relation entre F et f au niveau de l'espèce ou de la population), le choix de l'effort nominal a également des implications importantes pour l'application des interventions: selon la mesure choisie, les flottilles disposeront d'une latitude plus ou moins grande, mais toujours appréciable, pour optimiser leurs efforts effectifs dans les limites de l'effort global nominal imposé par l'Administration. La somme des efforts effectifs des bateaux risque d'excéder graduellement les limites fixées par l'autorité chargée de l'aménagement.
Comme les précédents, les modèles analytiques de production par recrue doivent être appliqués au niveau d'une population ou, à défaut, d'un ensemble de populations dont les paramètres démographiques sont suffisamment proches.
Ils tirent profit, d'une part, de l'autonomie des cohortes par rapport à l'environnement après leur recrutement et, d'autre part, de leur construction qui intègre les processus démographiques dominants. Ils permettent d'analyser les effets du taux global d'exploitation sur la phase exploitée et la production des cohortes successives, mais aussi, à la différence des modèles globaux, les effets de modifications de la répartition de la mortalité globale par pêche en fonction de l'âge des cohortes (diagramme d'exploitation). Cela permet de comparer l'impact de mesures de protection des juvéniles sur la production des cohortes, ou les interactions entre des flottilles exploitant les mêmes populations selon des diagrammes d'exploitation différents. Ils ignorent par contre la dynamique des cohortes avant leur recrutement.
Leur capacité supérieure se paie par des besoins plus grands en matière de données (structure démographique des captures). Les différentes variantes du modèle (modèles de Beverton et Holt, de Ricker, analyse séquentielle de populations,…) diffèrent par les techniques de traitement des données, mais non par les processus démographiques qu'ils visent à représenter. Le Groupe a utilisé l'analyse séquentielle de populations qui permet d'estimer les stocks et la mortalité par pêche à partir des données de capture par âges. Elle est aussi appelée analyse des cohortes, parce qu'elle permet de simuler séparément la dynamique de chaque cohorte, à partir des relations entre les effectifs suivants:
| Nombre de poissons vivants au début de l'année t+1 | Nombre de poissons vivants au début de l'année t | Nombre de poissons capturés durant l'année t | Nombre de poissons morts naturellement durant l'année t |
Le niveau du recrutement annuel-c'est-à-dire de l'effectif (cohorte) venant chaque année renouveler le stock et compenser les pertes subies par mortalité naturelle et par pêchedépend a priori de la biomasse parentale-par l'intermédiaire du nombre d'ceufs émis par la population-et de la survie de ces ceufs au cours des phases précoces, jusqu'à ce que la cohorte soit devenue relativement autonome par rapport à son environnement physique et biologique.
Longtemps, les deux relations-recrutement/stock parental et recrutement/environnement-ont été mal distinguées. On réalise mieux maintenant que:
pour les densités couramment observées dans les populations exploitées par pêche, la première relation ne joue généralement - à court et moyen terme, mais peut-être pas à long terme-qu'un rôle secondaire par rapport à celui de la seconde;
dans cette dernière, les processus spatiaux (par exemple, la dispersion des ceufs et des larves dans la structure hydrodynamique colonisée par les phases précoces de la population) l'emportent souvent sur les phénomènes trophiques; pour cette raison, les structures hydrodynamiques sont déterminantes pour la localisation-et donc le nombre-des populations au sein d'une espèce, ainsi que pour leur abondance moyenne;
même si les processus biologiques (trophiques) sont les médiateurs des phénomènes spatiaux, les variations interannuelles du succès du recrutement - y compris l'établissement ou la disparition de populations - sont d'abord déterminées par les fluctuations des structures hydrodynamiques; il en résulte que les phénomènes de densité-dépendance-y compris la relation stock-recrutement-sont généralement moins importants qu'on a pu le penser.
Malgré leur comlexité, ces questions sont importantes pour la pêche mauritanienne. La distribution et la porduction des populations de petits pélagiques sont susceptibles de changer avec l'évolution du régime d'upwelling. Les populations de céphalopodes se sont fortement accrues, il y a une trentaine d'années, au cours d'une période de chalutage intense. Bien que cette question soit très importante pour l'avenir de la pêche mauritanienne, les causes de cet accroissement, et donc d'un déclin éventuel des populations de céphalopodes, ne sosnt pas élucidées.
3.2.1.1 - Données disponibles et traitement
Captures
Les statistiques de débarquement sont disponibles pour les trois groupes d'espèces suivants:
poulpe, avec une seule espèce, le ‘tako’ (Octopus vulgaris);
seiches, qui comprend le ‘choco’ ou ‘mongo’ (S. officinalis et S. berthelotti), le plus abondant, et la sépiole (Sepiola sp.);
calmar (Loligo vulgaris essentiellement).
Les rejets sont nuls ou négligeables; en pratique, les débarquements correspondent aux captures (tabl. 1).
Pour le poulpe, la série de captures 1966–1983 a été complétée par les données du CNROP pour la période 1984–1992. Pour les seiches et les calmars, les séries de capture de 1984 à 1992 sont tirées des statistiques du CNROP. Les données plus anciennes relatives à la seule zone mauritanienne n'ont pas été préparées. Elles devraient être compilées.
| Années | Congélateurs | Glaciers | Pêche artisanale | Autres | Total |
|---|---|---|---|---|---|
| poulpe | |||||
| 1984 | 25 970 | 802 | 18 | -* | 26 790** |
| 1985 | 30 325 | 716 | 54 | 6 874 | 37 969 |
| 1986 | 40 965 | 441 | 390 | 3 457 | 45 253 |
| 1987 | 38 416 | 1 361 | 3 300 | 2 655 | 45 732 |
| 1988 | 31 834 | 1 666 | 2 500 | 726 | 36 726 |
| 1989 | 29 558 | 812 | 3 000 | 163 | 33 533 |
| 1990 | 18 395 | 720 | 2 350 | 0 | 21 465 |
| 1991 | 24 277 | 1 734 | 4 621 | 0 | 30 632 |
| 1992 | 30 782 | 4 920 | 8 257 | 0 | 43 959 |
| Seiches | |||||
| 1984 | 6 870 | 280 | 0 | 0 | 7 150 |
| 1985 | 8 000 | 81 | 0 | 2 387 | 10 468 |
| 1986 | 9 752 | 217 | 0 | 829 | 10 798 |
| 1987 | 5 820 | 470 | 0 | 1 320 | 7 610 |
| 1988 | 4 303 | 393 | 0 | 945 | 5 641 |
| 1989 | 4 875 | 565 | 0 | 86 | 5 526 |
| 1990 | 6 620 | 609 | 0 | 0 | 7 229 |
| 1991 | 5 650 | 652 | 0 | 0 | 6 302 |
| 1992 | 5 050 | 565 | 0 | 0 | 5615 |
| Calmar | |||||
| 1984 | 456 | 300 | 0 | 0 | 756 |
| 1985 | 547 | 4 | 0 | 353 | 904 |
| 1986 | 399 | 198 | 0 | 41 | 638 |
| 1987 | 223 | 19 | 0 | 0 | 242 |
| 1988 | 80 | 19 | 0 | 0 | 99 |
| 1989 | 493 | 50 | 0 | 0 | 543 |
| 1990 | 1 124 | 145 | 0 | 0 | 1 269 |
| 1991 | 1 241 | 241 | 0 | 0 | 1 482 |
| 1992 | 1 226 | 475 | 0 | 0 | 1 701 |
Structure démographique
Les modèles analytiques d'évaluation nécessitent la connaissance de la structure démographique des captures (fréquences d'âge ou, à défaut, de taille ou de poids).
Pour les poulpes, on dispose des fréquences de poids. Les quantités débarquées sont enregistrées par catégories commerciales. Lors des inspections de la SMCP, ces catégories commerciales font l'objet d'un échantillonnage biologique destiné à ventiler les captures de chaque catégorie commerciale en classes de poids de 100 g chacune. Cet échantillonnage a permis de vérifier que la composition des catégories commerciales variait très peu dans le temps. On peut donc raisonnablement appliquer les mêmes clés poids-longueur ou âge aux débarquements des années où les captures n'ont pas été échantillonnées. Les prises par âge des années 1991 et 1992 ont été calculées selon la méthode utilisée lors des réunions tenues précédemment au CNROP (CNROP, 1993).
On ne dispose pas des mêmes informations sur la structure démographique des captures de seiches et de calmar. Pour les premières, on pourrait tenter d'utiliser les données des campagnes scientifiques de prospection. Pour le calmar, un échantillonnage des catégories commerciales est nécessaire. Un tel programme devrait être lancé.
En 1991, et surtout 1992, les captures de petits poulpes (moins de 200 g) ont diminué par rapport à celles des tailles intermédiaires (autour de 1 kg); la taille moyenne des captures s'est accrue pour dépasser 1 kg en 1992. Cette évolution correspond au développement de la pêche piroguière au pot dont les captures, centrées sur les tailles moyennes (fig.2), ont atteint 10 000 tonnes en 1992. Les analyses antérieures de production par recrue ont montré que la distribution des tailles des captures de la pêche piroguière était, en termes de productivité, meilleure que celui de la pêche chalutière (CNROP, 1990). Toutes choses égales par ailleurs, cette meilleure productivité du stock de poulpe au cours des dernières années devrait apparaître sur les modèles globaux.
Le fait que la pêche piroguière capture des individus plus âgés ne peut être avancé à l'encontre de ce mode de pêche. En effet, si le recrutement est effectivement réduit par la pêche des reproducteurs-ce qui n'est pas observé-, il le sera davantage, à effectif capturé égal, si les individus sont capturés avant d'avoir pu pondre une première fois, c'est-à-dire si les captures comprennent plus d'immatures que d'adultes. En outre, pour un même tonnage total de capture, le nombre d'individus capturés est d'autant plus grand que la pêche porte sur des individus plus jeunes, donc plus petits.
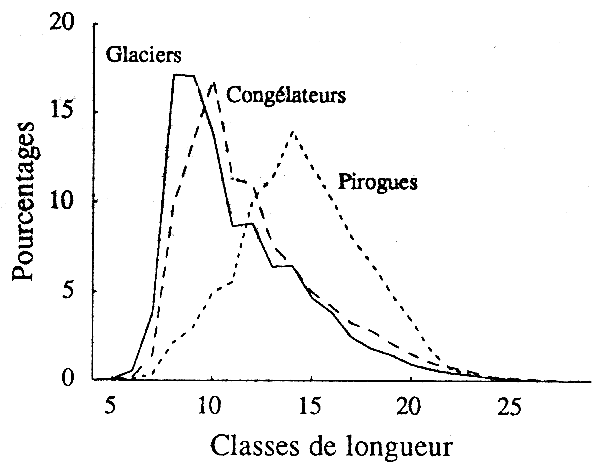
Figure 2- Distribution par taille des captures des flottilles de congélateurs, de glaciers et de pirogues (années 1989).
Effort de pêche
Différentes flottilles participent à l'exploitation des céphalopodes dans la ZEE mauritanienne (CNROP, 1992; Inejih, en préparation), à savoir:
flotte chalutière:
* congélateurs;
* glaciers;
flottille piroguière artisanale.
L'élaboration, pour chaque flottille, de séries d'effort de pêche, cohérentes avec celles de captures et cumulables, se heurte à deux difficultés:
les efforts de la flotte industrielle et de la flottille artisanale ne sont pas directement comparables; pour cumuler les efforts des deux flottilles, l'effort de l'une doit être calibré par rapport à celui de l'autre;
ces estimations d'effort se rapportent à l'ensemble des espèces démersales (céphalopodes et poissons) pêchées par les mêmes flottilles; comme leurs stratégies de pêche (choix des espèces cible) changent en cours d'annéee, et d'une année à l'autre de manière non cyclique, l'estimation d'efforts effectifs spécifiques (cf. § 3.1.1.2) demande une information dont on ne dispose pas; le poulpe étant recherché toute l'année, les écarts entre l'effort global et l'effort effectif pour cette espèce devraient rester faibles; compte tenu de la faiblesse de leurs captures de juin à octobre, période pendant laquelle l'abondance du poulpe est maximale, il n'en va vraisemblablement pas de même pour les seiches et le calmar (fig. 3).
Pour réduire ces biais, les estimations d'effort ont été intercalibrées de la façon suivante (tabl. 2):
on est parti des données brutes d'effort mensuel publiées par Inejih (1992) pour la période 1984 à 1990, et des données similaires préparées le même auteur pour les années 1991 et 1992;
certaines sources de données expriment l'effort en jours de mer, d'autres en heures de pêche: pour ;es combiner, on a admis que la durée de la pêche effective par jour de mer était de 18 heures;
pour estimer l'effort global déployé par toutes les flottilles, on a pris comme référence la flottille la plus importante, celle des congélateurs; les estimations de l'effrt des glaciers, de la pêche piroguière, et des aurtres flottilles pêchant accessoirement les céphalopodes ont été exprimées en équivalents ‘jours de mer congélateurs’; cela revient à admettre que leurs captures ont été réalisées avec des efforts égaux à ceux que les congélateurs auraient dû déployer pour réaliser les mêmes prises;
pour l'évaluationb du stock de poulpe, on a utilisé l'effort global fannuel (somme des efforts normalisés des quatre pêcheries);
pour les seiches et le calmar, on a aussi calcylé un effort “réduit”, obtenu en soustrauyant de l'effprt global annuel, la somme des efforts des mois de juin à octobre (inclus) correspondant à la période pendant laquelle les prises de calmar et seiches sont faibles, alors que l'effort global et les captures de poulpe sont élevées;
pour la pêche artisanale, on dispose de recensements de l'effectif des pirogues pêchant au pot en férvrier et en juillet de chaque année (enquêtes cadre); ces effectifs donnent une estimation de l'effort de pêche artisanal.
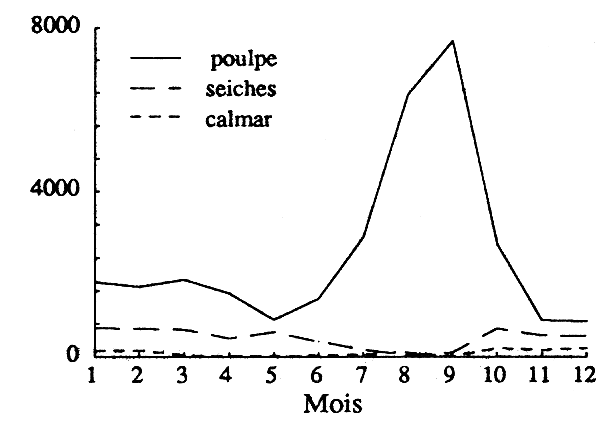
Figure 3 - Flottille de congélateurs: variation saisonnière de la composition dpécifique des captures de céphalopodes (1992).
| Congélateurs (jours de pêche) | Glaciers (jours de pêche) | Pêche artisanale (effectif des pirogues pêchant le poulpe) | |||
|---|---|---|---|---|---|
| Année | réduit(novembre à mai) | année | février | juillet | |
| 1984 | 20 113 | 12 549 | 2 033 | - | - |
| 1985 | 20 692 | 12 218 | 3 610 | - | - |
| 1986 | 23 559 | 13 267 | 2 877 | - | - |
| 1987 | 28 843 | 17 242 | 4 802 | - | 99 |
| 1988 | 21 659 | 12 325 | 2 546 | 177 | 142 |
| 1989 | 22 848 | 15 133 | 3 737 | 146 | 178 |
| 1990 | 19 960 | 11 882 | 2 376 | 277 | 343 |
| 1991 | 18 955 | 10 682 | 3 976 | 280 | 327 |
| 1992 | 14 428 | 8 487 | 3 807 | 281 | 385 |
Prises par unité d'effort et indices d'abondance
Des pue annuelles ont été calculées pour chaque groupe d'espèces et chaque flottille (tabl. 3). Pour la pêche artisanale, on a calculé deux séries d'abondance annuels en divisant la capture totale annuelle de poulpe réalisée par la pêche artisanale, par les effectifs de pirogues en février et en juillet.
| Années | Poulpe | Seiches | Calmar | |||||
|---|---|---|---|---|---|---|---|---|
| Congélateurs | Glaciers | Pêche artisanale (Capture totale annuelle par effectif pirogues) | Congélateurs | Glaciers | Congélateurs | Glaciers | ||
| (t/j) | (t/j) | février | juillet | (t/j) | (t/j) | (t/j) | (t/j) | |
| 1984 | 1, 29 | 0, 39 | - | - | 0, 34 | 0, 14 | 0, 02 | 0, 15 |
| 1985 | 1, 47 | 0, 20 | - | - | 0, 39 | 0, 02 | 0, 03 | 0, 00 |
| 1986 | 1, 74 | 0, 15 | - | - | 0, 41 | 0, 08 | 0, 02 | 0, 07 |
| 1987 | 1, 33 | 0, 28 | - | 33,3 | 0,20 | 0,10 | 0,01 | 0,00 |
| 1988 | 1, 47 | 0, 65 | 14, 2 | 17, 6 | 0, 20 | 0, 15 | 0, 00 | 0, 01 |
| 1989 | 1, 29 | 0, 22 | 20, 6 | 16, 9 | 0, 21 | 0, 15 | 0, 02 | 0, 01 |
| 1990 | 0, 92 | 0, 30 | 8, 5 | 6, 9 | 0, 33 | 0, 26 | 0, 06 | 0, 06 |
| 1991 | 1, 28 | 0, 44 | 16, 5 | 14, 1 | 0, 30 | 0, 16 | 0, 07 | 0, 06 |
| 1992 | 2, 13 | 1, 29 | 29, 4 | 21, 4 | 0, 35 | 0, 15 | 0, 08 | 0, 12 |
Depuis 1987, les pue des glaciers et des congélateurs évoluent assez parallèlement au sein de chaque groupe d'espèces (fig.4). Ceci s'expliquerait par le fait que les glaciers affrétés ciblent les céphalopodes, le poulpe particulièrement. Ils auraient done une stratégie de pêche voisine de celle des congélateurs. Les indices d'abondance de poulpe de la pêche artisanale sui vent la même évolution que celles des chalutiers. Les pue calculées refléteraient done bien la variation d'abondance, après 1987, des trois groupes d'espèces, et particulièrement celle du poulpe. Par contre, les pue de chaque groupe d'espèces ne varient pas parallèlement. Cette divergence peut provenir d'une évolution différente de l'abondance des trois ensembles d'espè ou/et de changements dans les stratgies de pêche des chalutiers, pricipalement les glaciers (on sait qu'avant 1987 les glaciers recherchaient relativement plus le poisson).
Le tableau l de l'annexe 4 donne, pour les trois groupes d'espèces et par zone, et la période 1982-92, les résultats des campagnes de chalutage du N/O ‘N'Diago’ (kg/demi-heure de trait). Le tableau 4 donne des indices d'abondance moyens annuels (moyennes annuellesdés désaisonnalisées), calculés à partir de ces données. Les années où les prospections ont été nombreuses et bien réparties dans le temps ont servi pour estimer un schéma de variation saisonnière et la moyenne de l'and'indices d'abondance saisonniers, rapports entre la valeur saisonnière et la moyenne de l'an née). Ce schéma a été utilisé pour désaisonnaliser les estimations des années où les prospections étaient moins fréquentes et mal réparties dans le temps. Les indices d'abondance du poulpe, des seiches et du calmar ainsi calculés (fig. 5) suivent approximativement, mais avec une amplitude moins forte, l'évolution donnée par les statistiques commerciales (fig.4). Ces données confirment, pour les seiches et le calmar-mais pas pour le poulpe-, l'amélioration des rendements durant la seconde moitié des années 80. Une certaine inversion des tendances des rendements de poulpe et de seiches apparaît, tant au niveau des indices d'abondance des campagnes de prospection du ‘N'Diago’, que de ceux déduits des statistiques commerciales. Ceci pourrait résulter d'un intérêt plus grand pour la pêche des seiches et des seiches et des calmars les années où le poulpe est relativement moins abondant.
En résumé, les différences d'évolution des indices d'abondance des mêmes groupes d'espèces (pue des congélateurs, des chalutiers, et du navire de recherche) peuvent provenir:
de différences dans le volume des échantillons, celui des flottilles commerciales étant supérieur à celui des prospections scientifiques;
du temps nécessaire aux pêcheurs pour détecter les zones de forte concentration des espèces;
de changements dans le choix des espèces cibles, qui se traduisent par la concentration des opérations de pêche sur certaines secteurs, variables selon les saisons.
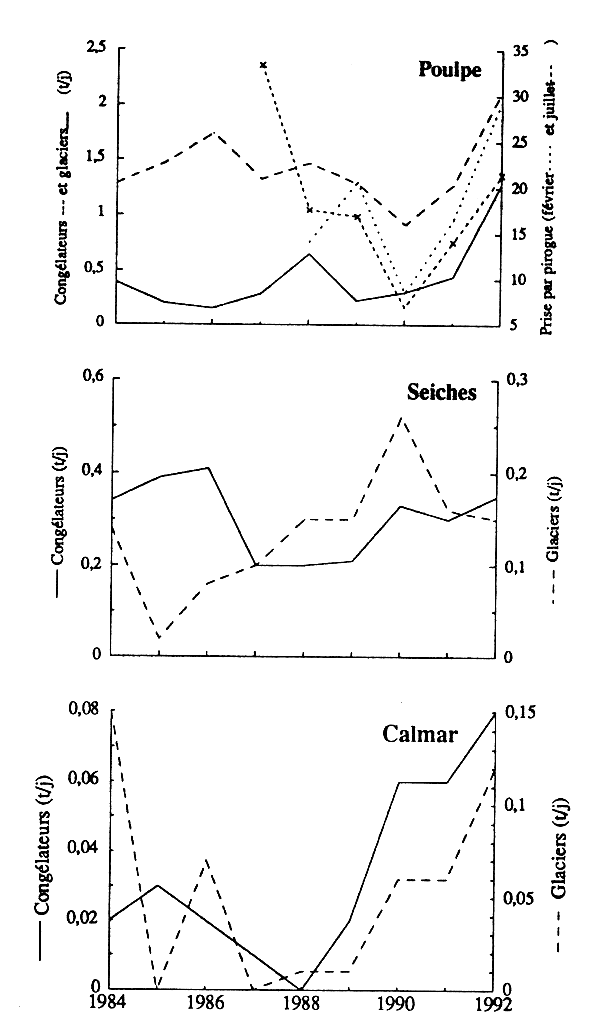
Figure 4 - Évolutin des pue des trois groupes d'espèces dans les différentes flottilles.
| 1982 | 1983 | 1984 | 1985 | 1986 | 1987 | 1988 | 1989 | 1990 | 1991 | 1992 | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Moyenne annuelle | |||||||||||
| Poulpe | 11,4 | 6,4 | 9,6 | 4,4 | 5,85 | 8,95 | 10,0 | 6,0 | 11,3 | 8,6 | |
| Seiches | 3,3 | 3,95 | 2,1 | 1,7 | 1,55 | 1,4 | 4,2 | 5,3 | 3,8 | 2,1 | |
| Calmar | 0,9 | 0,85 | 0,6 | 0,7 | 0,3 | 0,67 | 1,33 | 0,9 | 1,6 | 2,2 | |
| Facteur de désaisonnalisation | |||||||||||
| Poulpe | 0,78 | 1,0 | 0,93 | 1,86 | 1,0 | 1,0 | 1,0 | 0,93 | 0,93 | 0,73 | |
| Seiches | 0,86 | 1,0 | 0,67 | 0,93 | 1,0 | 1,0 | 1,0 | 0,67 | 0,67 | 0,82 | |
| Calmar | 0,85 | 1,0 | 1,11 | 0,37 | 1,0 | 1,0 | 1,0 | 1,11 | 1,11 | 0,67 | |
| Indices annuels désaisaonnalisés | |||||||||||
| Poulpe | 8,90 | 6,40 | 8,95 | 8,20 | 5,85 | 8,95 | 10,00 | 5,59 | 10,50 | 6,26 | |
| Seiches | 2,83 | 3,95 | 1,40 | 1,59 | 1,55 | 1,40 | 4,20 | 3,53 | 2,53 | 1,73 | |
| Calmar | 0,77 | 0,85 | 0,67 | 0,26 | 0,30 | 0,67 | 1,33 | 1,00 | 1,78 | 1,40 |
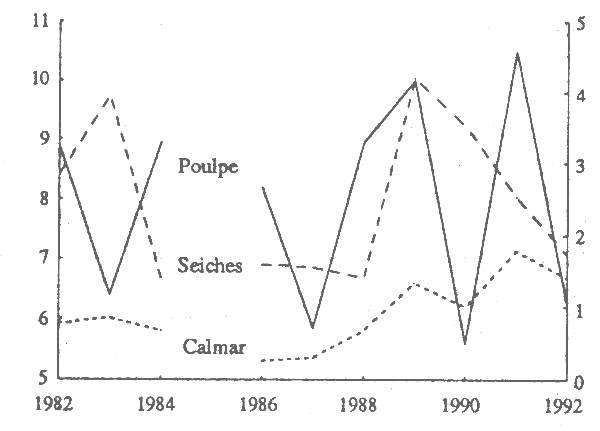
Figure 5 - Évolution des indices d'abondance tirés des campagnes de prospection (moyennes annuelles désaisonnalisées).
3.2.1.2 - Évaluation des stocks
Modèle global
Poulpe
La série historique des données de capture et d'effort a étéanalysée à l'aide d'un modèle généralisé (logiciel Prodfit, trois classes d'âge dans les captures, stock en état de transition, m = 1 et m = 2). La figure 6 représente l'evolution de la capture moyenne anuelle en fonction de l'effort, pour le stock en situation d'équilibre. La courbe convexe (m=2; r=0,82) représente mieux la distribution des données d'observation, que la courbe asymptotique (m = 1). Elle passe par un maximum de 49 000 tonnes pour un effort total annuel de l'ordre de 250 000 heures de pêche. En 1992, l'effort déployé était e 370 000 heures de pêche, soit 50% au-dessus de celui correspondant au maximum de production.
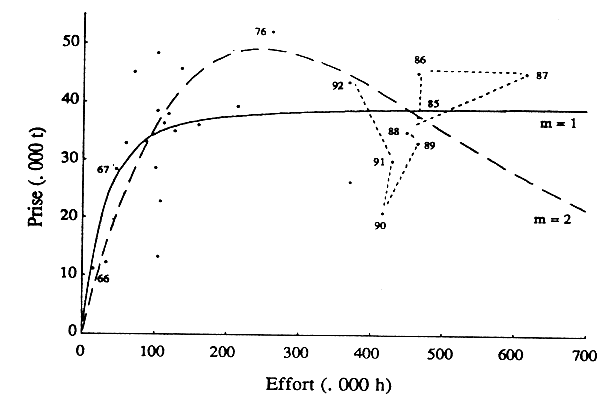
Figure 6 - Stock de poulpe: modèle global généralise (pour m=1 et 2), ajusté sur les données de 1966 à 1992
La courbe pour m=1 donne un maximum de production moyenne légèrement supérieur à 39 000 tonnes. Ce maximum correspond à un effort de 900 000 heures de pêche. Le coefficient de corrélation est de 0,87. Le maximum de production équilibrée obtenu par ce traitecontre, obtenu pour un niveau d'effort de pêche considérablement plus élevé.
Les estimations du potentiel de capture obtenues à l'aide des deux modèles sont légèrement supérieures-ce qui est conforme à l'amélioration du profil de capture survenue ces dernières années -, mais du même ordre que celles obtenues précédemment. L'evaluation du Groupe de travail de 1985 donnait une production maximale équilibrée de 35 000 tonnes, pour un effort de 230 000 heures de pêche. Celle de Gilly et Maucorps (1988) concluait que le potentiel maximum de capture était de 40 000 tonnes, pour un effort 250 000 heures.
Selon le profil de capture (qui dépend notamment de l'importance relative de la pêche artisanale et de la pêche industrielle), et les fluctuations naturelles du recrutement (le modèle global rend compete implicitement des variations passées, mais pas des veriations futures), le potentiel du stock de poulpe est compris entre 40 et 50 000 tonnes (peut-être plus proche de la limite supérieure). Bien qu'il se soit réduit ces dernières années, l'effort déployé par l'ensemble des flottilles sur le stock de poulpe dépasse largement celui qui correspond aux objectifs de la politique sectorielle (vior § 5.2.1).
La différence des estimations de l'effort correspondant au maximum de production, obtenues avec m=2 et m=1, n'est pas critique pour déterminer l'objectif d'aménagement. Le niveau d'exploitation correspondant aux objectifs de la politique sectorielle se situe, en effet, à gauche du maximum physique de production. Or, les deux modèles diffèrent surout sur leur partie droite. L'effort correspondant au maximum de production avec m = 1 ne constitue pas, en termes économiques, un objectif d'aménagement rationnel. En effet. entre 100 000 et 90 000 heures de pêches, la production - donc le revenu total - augemente peu comparativement à l'effort - donc au coût total de production.
seiches et calmar
L'analyse des pue en fonction de l'effort corrigé n'améliore pas sensiblement la cohérence des résultats obtenus en utilisant les efforts de pêches non corrigé. Dans ces conditions, l'utilisation des données corrigées ne se justifie pas. Le Groupe a done utilise, selon les mêmes procédures que pour poulpe, les séries statisques 1984–1982 correspondant aux captures des deux groupes d'esèces. Le traitement de ces données par un modèle global ne donne pas de résultat chhérent. Cela est vraisemblablement dûàla mauvaise estimation des efforts cible dominante, et où les straégies des flottilles ont changé.
Avec les données disponibles, les stocks de calmar et de seiches ne peuvent pas être évalués. Au cours des dernières années (89–91), tous les indices d'abondance disponibles pour ces espèces ont augmenté par rapport à ceux de la période 86–88, et lus nettement pour le calmar. Ce redressement peut être dû à des causes naturelles, ou à des changements dans le régime d'exploitation. On remarque que les indices tirés des campagnes (fig.5), s'ils augmentent bien pour le calmar après 1987, diminuent pour les seiches après 1989.
Du fait de la prédominance du poulpe dans la pêche des céphalopodes, l'aménagement de la pêcherie peut rester basé sur l'évaluation du stock de poulpe. En 1992, le poulpe représentait 85 % des prises de céphalopodes. Dans les conditions actuelles, l'état des stocks de seiches et de calmar, ou les incertitudes concernant cet état, ne peuveant être avancés comme arguments en faveur d'une intensification de la pêche des céphalopodes. Si elle existe, cette possibilité devrait être utilisée pour réduire l'effort exercé sur le poulpe. Cette possibilité dépend de la capacité à modifier la répartition de l'effor exercé total entre le poulpe, d'une part, et les autres espèces, d'autre part. Ceci peut être recherché par l'emploi d'engins sélectifs (casiers ou turluttes), ou par l'incitation des pêcheurs à modifier leurs stratégies de pêche-en modulant par exemple le taux par espèces de la taxe d'exportation. Si ces conditions pouvaient être réunies, un transfert graduel de l'effort exercé sur le poulpe vers les seiches, et surtout le calmar, est envisageable. L'évaluation des stocks de seiches et de calmar deviendrait alors urgente.
traitement complémentaire des statistiques
L'analyse des statistiques commerciales (y compris de la pêche artisanale) et des campagnes de prospection a montré la bonne qualité et la sensibilité générales des données. Le traitement de ces données devrait être raffiné, en accordant une attention particulière aux données relatives aux seiches et au calmar. On partira pour cela de l'analyse des statistiques individuelles des bateaux (petites strates espace-temps où les rendements sont les plus élevés), de façon à classer ces derniers sur la base de leurs stratégies de pêche (maximisation indifférente de la capture totale de céphalopodes, recherche préférentielle des seiches ou calmar pendant certaines saisons, notamment celles où les rendements de poulpe sont minima). Cette analyse peut être étayée par une enquête auprès des patrons de pêche. Les résultats des campagnes de prospecpection ne sont pas à négliger. Si les statistiques commerciales représentent un volume d'échantillonnage supérieur, on ne peut exclure que des strates bien choisies des campagnes de prospection pouissent donner des indices plus sensibles, ou moins biaisés dans le temps. Ensuite, l'évolution des séries les plus homogènes de pue ainsi indentifiées sera comparée à la même échelle, l'objectif étant de sélectionner les séries susceptibles de fournir les estimations les plus vraisemblables de l'abondance relative des différentes espèces, ou de strates particulières (âge ou tailles de celles-ci.Si l'on dispose de plusieurs séries apparemment fiables, elles pourment être combinées.
Le CNROP devrait donc entreprendre dès maintenant les traitements suivants:
reconstitution de séries longues de capture et d'abondance (données antérieures à 1984 de la pêche japonaise);
analyse fine des statistiques commerciales - industrielles et artisanale - et des campagnes de prospection, pour le poulpe, les seiches et le calmar;
comparaison des séries d'abondance des trois groupes d'espèces, afin de calculer, pour chaque groupe, la meilleure série;
reprise des évaluations à l'aide du modèle global, pour chaque groupe d'espèces;
mise en place éventuelle d'un programme d'échantillonnage biologique des prises commerciales de seiches et de calmar.
Modè le analytique 1
traitement des données
Les données disponibles ne permettent d'appliquer ce modèle qu'au stok de poulpe. Comme au cours des groupes de travail précédents, des analyses de pseudochoretes ont été conduites sur des séries d'effectifs moyen capturés par classes de taille, corespondant à différentes périodes de l'histoire de la pêcherie mauritanienne, à savior:
période 1966–1970, correspondant à la pêche extrême-orientale décrite par Hatanake (CNROP, 1990);
période 1985–1988;
période 1989–1991;
année 1992.
Les effectifs capturés par trailles sont donnés dans le tableau 2 de l'annexe 3. La figure 7 représente les effectifs moyens par âges pour les quatre périodes considérées.
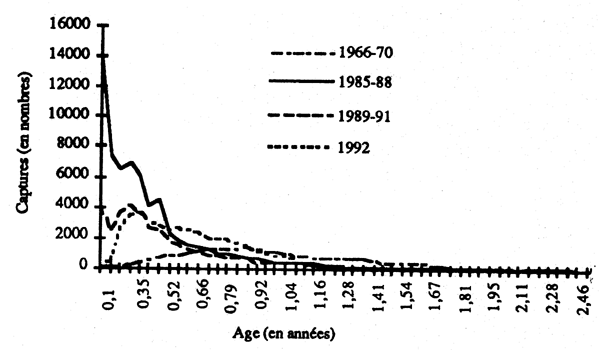
Figure 7 - Distributions de fréquence des captures de poulpe (moyennes correspondant aux quatre périodes retenues dans l'histoire de la pêcherie).
Ces analyses par pêriodes visaient àmettre en êvidence des changements dans le régime d'exploitation (effort global et distribution en fonction de l'âge), et d'en tirer des indications sur les interventions justifiées par l'état actuel de la pêcherie.
L'état des connaissances de la biologie du poulpe et les statisques disponibles obligent à reconsidérer l'hypothèse d'invariance de la mortalite naturelle pendant la phase exploitée. Cette hypothèse soulève un réel problème: les analyses démographiques confirement de plus en plus clairement les acquis de la biologie, à savoir que la mortalité naturelle s'élève après la ponte, brutalement chez les femelles, et probablement fortement chez les mâles. La prise en compte de ce phénomène dans les analyses de chortes ne devrait pas poser de problème méthodologique. Elle modifierait pourtant de façon significative les conclusions. La difficulté majeure réside dans la détermination du diagramme de mortalité naturelle après la ponte, ainsi que celle de l'âge de première reproduction (âge moyen et variabilité de cet âge). Ceci nécessite un travail important d'expérimentation et d'observations directes.
A cette incertitude s'ajoutent la méconnaissance de l'âge des poulpes, qui empêche de suivre convenablement l'évolution des cohortes dans la pêcherie, et l'imprécision sur l'estimation du taux annuel effectif d'exploitation. Pour ces raisons, seules des méthodes grossières, dont les limitations se répercutent sur la validité des conclusions, peuvent être employées.
taux de mortalité par pêche
Ces incertitudes se répercutent nécessairement sur les estimations de mortalité par pêche en fonction de l'âge déduites de ce traitement (fig. 8), et il n'est pas possible d'apprécier l'ampleur des erreurs résultantes. Néanmoins, quelles que soient les analyses effectuées, les solutions les plus raisonnables conduisent à des conclusions comparables.
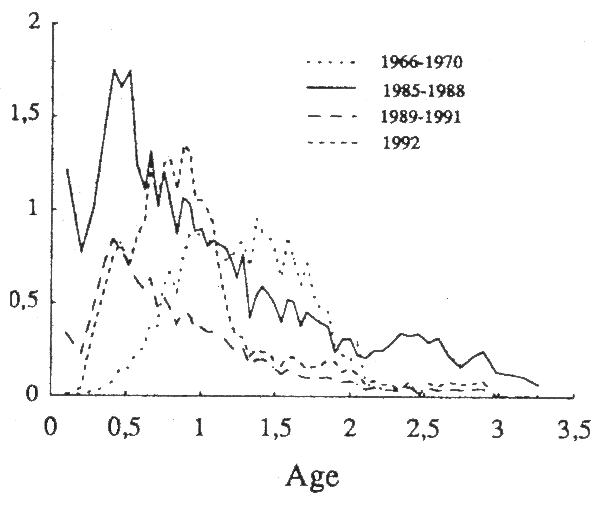
Figure 8 - Analyse de pseudocohortes du stock de poulpe: profils de mortalité par pêche au cours de quatre périodes de l'histoire de la pêcherie.
Le schéma d'exploitation aurait évolué de façon sensible. Il semble que l'on puisse distinguer trois périodes dans l'histoire de la pêcherie:
initialement, la pêche extrême-orientale visait les tailles intermédiaires; très peu de jeunes individus étaient capturés; le profil de mortalité par pêche en fonction de l'âge était caractéristique d'une exploitation qui protégeait les juvéniles; on a là l'indication qu'une surexploitation des juvéniles n'est peut-être pas inévitable dans la pêche au chalut;
pendant la période intermédiaire (85–88, puis 89–91), au cours de laquelle la proportion de juvéniles dans les prises était élevée, la mortalité par pêche s'est élevée de façon marquée; le stock aurait été à la fois surexploité et mésexploité;
au cours de la dernière année d'observation (1992), la mortalité par pêche a diminué à la fois globalement et relativement; les jeunes seraient mieux épargnés; le stock serait moins et mieux exploité.
production par recrue
Dans la mesure où elles sont représentatives, les estimations précédentes de mortalité par pêche peuvent être utilisées pour affiner ce scénario. Pour cela, on peut les introduire dans un modèle de production par recrue - de Ricker par exemple -, qui intègre les effets de la croissance, de la mortalité naturelle, et de la mortalité par pêche en fonction de l'âge. On peut ainsi apprécier l'efficacité technique de la pêche à exploiter convenablement les cohortes (indépendamment des niveaux annuels du recrutement). Si l'on admet une mortalité naturelle (M) constante et égale à 1, 0 pendant toute la phase exploitée, le recrutement annuel moyen serait de l'ordre de 11 à 14 millions de poulpes de 100 grammes, et serait resté relativement stable au cours des dernières années. Si l'on admet un recrutement moyen et stable de 12 millions d'individus de 100 g chaque année, un tel traitement conduit aux conclusions suivantes (annexe 4, tabl. 3a et 3b):
en 1992, le taux d'exploitation a diminué par rapport au niveau de la fin des années 1980; en termes biologiques, la surexploitation aurait diminué;
avec la taille de première capture observée en 1992, la production par recrue est élevée: une augmentation de la taille de première capture par rapport à cette valeur n'augmenterait pas de façon significative la productivité du stock; l'amélioration récente du régime d'exploitation du stock porte donc à la fois sur la réduction de la mortalité par peche et sur sa distribution par âge; la distribution de taille des captures s'est rapprochée de celle de la pêche japonaise au cours des années 60 (CNROP, 1991); si l'on savait que la réduction de la pêche des poulpes de petite taille est bénéfique, l'évolution récente de la pêcherie montre qu'elle est possible.
Une analyse approfondie de l'évolution des diagrammes de capture depuis l'origine de la pêche, ainsi que leur comparaison entre les différentes flottilles actuellement en activité, permettraient d'approfondir ces premières conclusions. Cette analyse devrait porter à la fois sur les aspects démographiques (évaluation) et sur les aspects réglementaires (aménagement). Il s'agit de vérifier si la réduction relative des captures de petite taille est due principalement à l'augmentation de l'activité de la pêche piroguière, ou si elle s'est produite également dans la pêche chalutière. Si c'était bien le cas, il faudrait en examiner les raisons pour en tirer des enseignements sur les possibilités pratiques de mieux protéger les juvéniles dans ce type de pêche. Malgré les incertitudes qui affectent l'évaluation par un modèle analytique, les résultats que l'on obtient sont très comparables à ceux obtenus avec le modèle global. Sans être une preuve, cette similitude renforce la confiance que l'on peut accorder aux premières conclusions.
3.2.1.3 - Besoins de recherche sur l'écologie et la dynamique des populations de poulpe
Programme de recherche régional sur la biologie et l'écologie du poulpe
Les stocks de poulpe de la région sont les plus importants du monde. Les exportations de céphalopodes sont pour les pays riverains une source majeure de devises. Les recettes substantielles que certains Etats tirent déjà de leurs pêcheries pourraient être significativement accrues si ces dernières étaient convenablement gérées. Si, actuellement, l'amélioration des régimes d'exploitation dépend d'abord de l'adoption de mécanismes nécessaires à l'allocation des droits de pêche (§ 5.2.1), l'évaluation et le suivi de l'état des stocks seront essentiels pour l'aménagement. En fait, la demande en matière d'évaluation et de suivi des stocks croîtra avec le progrès de l'aménagement.
Actuellement, l'obstacle majeur à l'amélioration des évaluations de stocks réside dans l'insuffisance des connaissances sur la l'écologie du poulpe. Vu la similitude des stocks, des conditions écologiques et des pêcheries, et le coût de recherches nouvelles, les pays de la sous-région ont grand intérêt à coopérer à l'exécution de telles investigations. Les sujets prioritaires de recherche sont les suivants:
identité et schémas de distribution et de déplacement des populations de poulpe:
* les poulpes ‘noirs’ et les ‘poulpes d'eau’ appartiennent-ils à une population distincte ou à un stade physiologique particulier (sénescence postérieure à la ponte)?
* la connaissance de l'identité des populations et de leurs schémas de distribution entre les ZEEs de la sous-région fournirait des informations sur les besoins et les possibilités d'aménagement des stocks partagés;
* celle des variations et du déterminisme du recrutement en fonction des conditions écologiques et halieutiques (apparition, propagation et déclin des populations de poulpe ouest-africaines), et de l'écologie et de la dynamique des phases critiques permettrait de comprendre les conditions du maintien d'un recrutement élevé;
mortalité naturelle: évolution, en fonction de l'âge des femelles et des mâles, de la mortalité naturelle après la reproduction;
croissance du poulpe: courbe moyenne et variabilité individuelle;
confirmation ou invalidation de l'hypothèse d'une fraction cryptique des populations de poulpe;
implications des particularités de la biologie et de l'écologie du poulpe sur la dynamique des populations, les techniques d'évaluation des stocks, et les stratégies d'exploitation.
En 1992, des chercheurs spécialistes de ces questions ont élaboré un programme de recherche intensif, étalé sur trois ans, portant sur la biologie, le comportement, l'écologie, l'identité (par marquage et méthodes génétiques) des populations de poulpe, et la modélisation bio-économique des pêcheries. La réalisation de ce programme devrait apporter des réponses à la plupart des questions évoquées ci-dessus. Vu son importance pour l'avenir des pêcheries de poulpe de la sous-région, le Groupe recommande vivement que les centres de recherche des pays riverains et les instituts de recherche et de développement étrangers qui s'intéressent à leurs travaux collaborent à sa réalisation.
Simulation des pêcheries
Le CNROP devrait entreprendre sans tarder des analyses de simulation de la pêcherie de céphalopodes, en vue d'évaluer:
les effets de différentes mesures de protection des juvéniles;
les interactions entre les pêches artisanale et chalutière.
Les statistiques de prise par âge et par saison doivent permettre de simuler les effets, à court, moyen et long termes, sur le stock et la pêcherie de diverses mesures de protection des juvéniles (périodes de repos biologique, maillage). Ces simulations fourniraient l'information nécessaire pour évaluer l'effet, sur le chiffre d'affaire, de la variation du prix du poulpe en fonction de la taille des captures.
L'aménagement devra définir aussi l'équilibre entre les deux composantes - artisanale et industrielle - de la pêcherie. La simulation des interactions techniques entre pêches piroguière et chalutière fournirait aux économistes l'information sur la ressource dont ils ont besoin pour évaluer les conséquences en termes de création de richesses et de devises, de réalisation de la rente halieutique, ou d'emploi, de différents schémas de partage du potentiel du stock de poulpe.
3.2.2.1 - Évolution du stock et de la pêche
Les captures mauritaniennes annuelles de poissons démersaux par flottilles sont données dans le tableau 5 et la figure 9.
Tableau 5 - Captures annuelles (en tonnes) de poissons démersaux des flottilles mauritaniennes.
| Années | Congélateurs | ∑ Glaciers* | Pêche artisabale | Total Mauritanie |
|---|---|---|---|---|
| 1984 | 20 656 | 5 422 | 10 203 | 36 281 |
| 1985 | 24 751 | 6 477 | 8 972 | 40 200 |
| 1986 | 15 564 | 8 446 | 8 012 | 32 021 |
| 1987 | 22 455 | 4 857 | 11 254 | 38 565 |
| 1988 | 8 491 | 3 584 | 17 441 | 29 517 |
| 1989 | 8 077 | 4 902 | 8 332 | 21 311 |
| 1990 | 11300 | 3636 | 4377 | 19313 |
| 1991 | 5822 | 3793 | 4395 | 14010 |
| 1992 | 4 930 | 1 373 | 6 518 | 12 821 |
* : chalutiers mauritaniens et affrétés
La figure 9 fait apparaître une baisse régulière de la production totale de poissons à partir de 1987. Après avoir atteint 40 000 tonnes en 1985, les débarquements totaux ont décliné pour descendre en dessous de 13 000 tonnes en 1992. Avec des différences, cette baisse s'observe dans les trois flottilles. Elle est particulièrement marquée pour les congélateurs, dont les captures déclarées sont tombées de 24 700 tonnes en 1985 à 5 000 tonnes en 1992.
1 - Cette partie du rapport final est basée sur la contribution de M. G. O/M. Fall et D. Leung Tack.
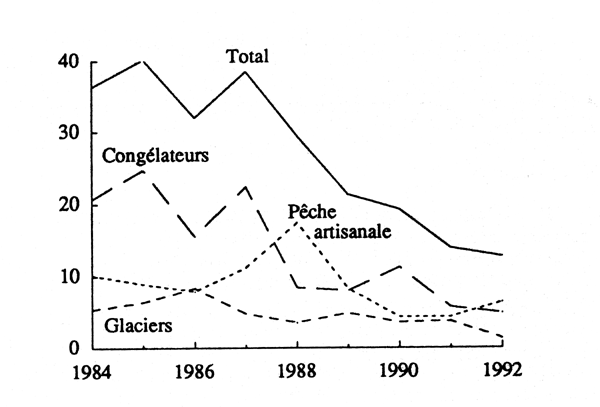
Figure 9 - Captures annuelles (en milliers de tonnes) de poissons par flottilles et totales
Cette évolution reflète des processus complexes qui compliquent l'analyse des données (CNROP, 1992):
le poids respectifs des différentes flottilles dans l'effort de pêche total déployé sur les poissons: sur l'ensemble de la période, les prises des congélateurs dominent dans les débarquements; elles sont suivies par celles de la pêche artisanale, puis par celles des glaciers (fig. 9);
l'intérêt de chaque flottille pour la pêche des céphalopodes et des poissons: par rappot aux congélateurs et aux glaciers étrangers, les glaciers mauritaniens s'intéressent davantage à la pêche des poissons démersaux; la proportion de poissons dans ses captures est restée plus élevée et plus stable sur la période d'observation; ainsi, en 1992, les captures de poissons des glaciers mauritaniens représentaient encore 80 % de ses débarquements totaux;
des changements dans les stratégies de pêche des flottilles: l'intérêt pour la pêche des poissons augmente lorsque l'abondance des céphalopodes décline (Inejih, sous presse); ainsi, après 1987, les congélateurs et les glaciers étrangers se sont intéressés encore plus à la pêche des céphalopodes; en 1992, leurs captures de poissons ne représentaient plus respectivement que 14 et 2 % de leurs débarquements totaux.
Compte tenu de l'intérêt plus grand et plus constant manifesté par les glaciers mauritaniens pour la pêche des poissons, leur pue devrait fournir un meilleur indice de l'abondance du stock. Le tableau 6 donne le nombre et les caractéristiques physiques moyennes des bateaux de cette flottille, et leur évolution au cours de la dernière décennie. La structure de la flottille est restée homogène. Par contre, son effectif a fluctué, pour décliner à partir de 1989. Le tableau 7 donne pour la même période:
les quantités totales de poissons démersaux débarquées;
celles des glaciers mauritaniens;
les rendements des glaciers mauritaniens,
les indices d'abondance tirés des campagnes de prospection en saison froide et saison chaude.
L'évolution des différents indices d'abondance (fig. 10) révèle:
une assez bonne concordance entre les indices indices tirés des statistques commerciales et des prospections scientifiques;
des fluctuations assez fortes des pue des glaciers;
une tendance modérée apparente à la baisse.
| Année | Nombre | Tonnage moyen (tonneaux de jauge brute) | Puissance moyenne (CV) | Longueur moyenne (HT,m) |
|---|---|---|---|---|
| 1984 | 24 | 88 | 473 | 19 |
| 1985 | 18 | 72 | 352 | 19 |
| 1986 | 29 | 76 | 377 | 19 |
| 1987 | 29 | 76 | 369 | 19 |
| 1988 | 34 | 74 | 385 | 19 |
| 1989 | 28 | 61 | 364 | 19 |
| 1990 | 26 * | 76 | 283 | 20 |
| 1991 | 12 ** | 92 | 296 | 20 |
| 1992 | 16 *** | 91 | 310 | 20 |
* :pour un total de 34 glaciers;
** :pour un total de 26 glaciers; arrêt des glaciers de l'armement ALMAP;
*** :pour un total de 36 glaciers.
| Années | Débarquements (tonnes) | Pue (tonnes/jour de mer) | Prospections scientifiques (tonnes/demi-heure) | ||
|---|---|---|---|---|---|
| ∑ Mauritanie * | Glaciers mauritaniens | saison froide | saison chaude | ||
| 1982 | 211,7 | ||||
| 1983 | 231,4 | ||||
| 1984 | 36 281 | 2 830 | 1,04 | 363,6 | |
| 1985 | 40 200 | 3 164 | 1,60 | ||
| 1986 | 32 022 | 5 074 | 1,40 | ||
| 1987 | 38 565 | 4 387 | 1,05 | 174,8 | 198,4 |
| 1988 | 29 516 | 3 487 | 1,00 | 225,8 | 230,4 |
| 1989 | 21 311 | 4 857 | 1,20 | 201,3 | 144,9 |
| 1990 | 19 313 | 3 394 | 1,95 | 249,0 | |
| 1991 | 14 010 | 3 177 | 1,06 ** | 97,3 | 184,7 |
| 1992 | 12 821 | 1840 | 0,60 *** | ||
* :Congélateurs + glaciers + pêche artisanale.
** :Année où le nombre de glaciers mauritaniens a fortement diminué;
*** :Année où le stock de poulpe s'est redressé.
Les débarquements annuels des glaciers par principaux groupes d'espèces (tabl. 8) ne montrent pas une évolution homogène. On observe bien, à partir de la fin des années 80, une baisse des débarquements pour plusieurs groupes (sparidés, serranidés, sélaciens), mais le déclin apparaît à des dates différentes et est plus ou moins prononcé selon les groupes. Si les haemulidés (hors Plectorhynchus mediterraneum) connaissent une baisse similaire, celle-ci ne se manifeste qu'en 1992 chez cette dernière espèce. Pour les serranidés, la baisse s'observe à partir de 1989 dans les statistiques commerciales, et aussi dans les indices d'abondance tirés des campagnes de prospection (surtout de saison froide).
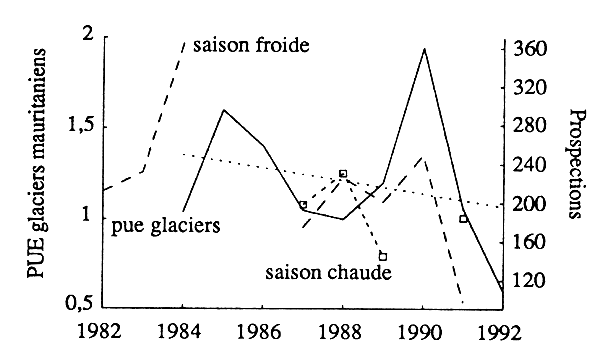
Figure 10 - Poissons démersaux: évolution des pue des glaciers mauritaniens et des indices d'abondance (saisons chaude et froide) des campagnes de prospection.
| 1984 | 1985 | 1986 | 1987 | 1988 | 1989 | 1990 | 1991 | 1992 | |
|---|---|---|---|---|---|---|---|---|---|
| Sparidés | 336 | 860 | 1582 | 2075 | 1751 | 2990 | 1398 | 313 | |
| Serranidés | 47 | 58 | 80 | 150 | 219 | 99 | 75 | 2 | |
| Sciaenidés | 188 | 309 | 365 | 326 | 448 | 539 | 314 | 7 | |
| Haemulidés* | 16 | 134 | 40 | 77 | 83 | 82 | 42 | 1 | |
| Plectorhynchus | |||||||||
| mediterraneum | 432 | 445 | 311 | 46 | 127 | 175 | 455 | 0 | |
| Pseudupaeneus | |||||||||
| prayensis | 2 | 26 | 71 | 81 | 153 | 151 | 76 | 0 | |
| Pleuronectiformes | 80 | 135 | 243 | 370 | 452 | 474 | 556 | 259 | |
| Sélaciens | 444 | 579 | 265 | 273 | 86 | 82 | 45 | 2 | |
| Autres poissons | |||||||||
| démersaux | 1284 | 619 | 2117 | 990 | 171 | 203 | 401 | 478 | |
| Total poissons | |||||||||
| démersaux | 2830 | 3164 | 5074 | 4387 | 3489 | 4805 | 3394 | 3362 | 1062 |
| Effort (j.de mer) | 2 720 | 1 975 | 3 676 | 4 159 | 3 626 | 3 975 | 1 737 | 3 177 | 1 840 |
* :non compris Plectorhynchus mediterraneum.
L'évolution des pue (débarquements)montre des évolutions différentes selon les groupes d'espèces (tabl.9): ils passent par un pic en 1988 pour les serranidés et les rougets, en 1989 pour les sparidés et les sciaenidés, alors qu'ils augmentent régulièrement chez les poissons plats jusqu'en 1991, et que pour les sélaciens, ils déclinent depuis 1985. Ces variations paraissent refléter au moins autant des changements dans le choix des espèces cible que dans l'abondance absolue des espèces. Ces changements de stratégie dans l'exploitation des poissons (y compris dans les taux de rejet) réduisent beaucoup la signification des indices d'abondance et des efforts effectifs tirés des statistiques commerciales. On sait que ces dernières années les glaciers étrangers et les congélateurs ont eu tendance à s'intéresser davantage à la pêche du poulpe (Girardin, 1988), et à accroître en conséquence leurs rejets de poissons. Une observation conforte cette interprétation: en 1992, l'effort des congélateurs s'est déplace vers la zone centrale de la ZEE (Inejih, en prép.); alora que, dans cette zone, le poissons est relativement plus abondant, cette abondance n'apparaît pas dans les débarquements. La réduction des liaisons aériennes par lesquelles une partie importante du poisson noble débarqué à Nouadhibou était exportée en frais vers l'Europe peut étre la cause de ce changement de comportement, Si c'était le cas, le stock de poisson devrait être moins fortement exploité que ne l'indiquent la baisse des débarquements par jour de pêche des glaciers, et l'estimation de l'effort de pêche tirée des mêmes statistiques commerciales.
| Années | Sparidés | Diagramme | Serranidés | Sciaenidés | Rouget | Sélaciens | Poissons plats |
|---|---|---|---|---|---|---|---|
| 1984 | 0,12 | 0,16 | 0,02 | 0,07 | 0,00 | 0,16 | 0,03 |
| 1985 | 0,32 | 0,23 | 0,03 | 0,16 | 0,01 | 0,29 | 0,07 |
| 1986 | 0,58 | 0,09 | 0,02 | 0,10 | 0,02 | 0,07 | 0,07 |
| 1987 | 0,76 | 0,01 | 0,04 | 0,08 | 0,02 | 0,07 | 0,09 |
| 1988 | 0,64 | 0,04 | 0,06 | 0,12 | 0,04 | 0,02 | 0,13 |
| 1989 | 1,10 | 0,04 | 0,03 | 0,14 | 0,04 | 0,02 | 0,12 |
| 1990 | 0,51 | 0,14 | 0,02 | 0,10 | 0,02 | 0,01 | 0,18 |
| 1991 | - | - | - | - | - | - | - |
| 1992 | 0,12 | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 | 0,14 |
Toutefois, la prédominance de la pêche des céphalopodes (51 300 tonnes de céphalopodes contre 12 000 tonnes de poissons débarquées en 1992) fait que les flottilles qui les flottilles qui recherchent ces espèces peuvent contribuer à maintenir un effort de pêche élevé sur les stocks de poisson. Il est peu probable, par ailleurs, que le recrutement soit la cause de la baisse apparente des rendements de poissons: d'une part, parce que le recrutement des poissons démersaux est souvent relativement stable (par rapport à celui des petits pélagiques ou de diverses espèces de bivalves par exemple); d'autre part, parce, parce que les fluctutation du recrutement au niveau des espèces devraient ê tre amorties au niveau de leur ensemble.
Deux évaluations de la biomasse minimale de poissons démersaux du plateau continental, basées sur les résultats de prospections, on été publiées (Josse et Garcia, 1985). La première correspond à la période 1982-1984 (campagnes du `N'Diago' pour le plateau continental entre 0 et 200 m,et de l"Almoravide' pour le secteur du banc d'Arguin et de la baie du Lévrier). Les biomasses minimales estimées de poisson démersal sont de 57 500 tonnes pour le plateau continental et de 27 500 tonnes pour la baie du Lévrier et le banc d'Arguin, soit 85 000 tonnes au total.
La seconde correspond à la période 1989-92 (campagnes du ‘N'Diago’); elle n'inclut pas le secteur de la baie du Lévrier et du banc d'Arguin. Les estimations de biomasse par saisons et par zones, tirées des campagnes du ‘N'Diago’ (tabl. 10) montrent une grande variabilité spatio-temporelle. Les biomasse les plus fortes ont été successivement recontrées dans les secteurs centre, sud, et nord. On sait néanmoins, de par la répartition des activités de chalutage, que la partie nord du plateau mauritanien est exploitée plus intensivement que la partie sud (de la latitude de Nouakchott à la frontière sénégalaise). La biomasse minimale moyenne pour l'ensemble du plateau est de 91 400 tonnes.
Ces deux estimations de biomasse instantanée pour l'ensemble du plateau continental (57 500 tonnes pour la période 1982–84 et 91 400 pour la période 1989–92) diffèrent d'autant plus significativement que la différence est de sens opposé à l'évolution des rendements des glaciers. Cette différence ne peut vraisemblablement s'expliquer par une meilleure efficacité du chalut du ‘N'Diago’ (obtenu par des modification du train de pêche en 1989), car des corrections ont été apportées pour tenir compete de ces modifications. Cette différence incite, néanmoins, a penser que la baisse des quantités de poissons débarquées par les glaciers proviendrait en grande partie de leur intérêt plus grand pour la pêche des céphalopodes, et de problèmes de gestion et de commercialisation rencontrée par les sociétés qui arment ces bateaux (SALIMAUREM et ALMAP).
Les données disponibles ne permettent donc pas d'établir un diagnostic clair de l'état du stock. Néanmonins, le stock de poissons du plateau serait moins intensément exploité que ne l'indique la baisse des rendements apparents (débarquements) des chalutiers glaciers.
| Saisons | Années | Secteurs | |||
|---|---|---|---|---|---|
| nord | centre | sud | total | ||
| Saison froide | 1989 | 32,4(84%) | 41,0(84%) | 35,3(89%) | 108,7 |
| 1990 | 18,6(76%) | 25,9(79%) | 33,7(83%) | 78.2 | |
| 1991 | 13,1(72%) | 21,5(84%) | 15,0(84%) | 49,6 | |
| Saison | 1989 | 24,7(68%) | 34,2(86%) | 19,8(89%) | 78,7 |
| 1991 | 36,9(92%) | 33,5(82%) | 25,2(90%) | 95,6 | |
| Inter- Saisons | 1989 | 14,9(67%) | 65,3(91%) | 38,0(86%) | 118,2 |
| 1992 | 34,6(92%) | 33,5(70%) | 42,7(84%) | 110,8 | |
| Moyennes annuellles | 1989 | 24,0(74%) | 46,8(88%) | 31,0(88%) | 101,8 |
| 1990 | 18,6(76 %) | 25,9(79 %) | 33,7(83 %) | 78,2 | |
| 1991 | 25,0(86 %) | 27,3(82 %) | 20,1(88 %) | 72,5 | |
| 1992 | 34,6(92 %) | 33,5(70 %) | 42,7(84 %) | 110,8 | |
| Moyenne | 1989–92 | 25,1(80 %) | 36,4(83 %) | 30,0(86 %) | 91,4 |
| Densité moy.(t/m2) | 10,9 | 13,8 | 12,5 | 12,5 | |
| N.B.: Les pourcentages indiquent la proportion de poissons démersaux dans les prises totales. | |||||
3.2.2.2 - État du stock et potentiel de capture
Josse et Garcis (1985) estimaient à 50 000 tonnes le potentiel de sparidés du plateau continental (prise moyenne de 25000 tonnes doublée pour tenir compte des rejets importants par les flottilles ne s'intéressant pas directement à la pêche du poisson). Le potentiel des autres espèces était évalué à 34 000 tonnes: considérées comme moins surexploitées, la prise moyenne courante de ces espèces n'a été multipliée que par un facteur de 1,5. Le potentiel total de poisson démersal ainsi calculé était donc de 84 000 tonnes, soit 67 000 tonnes pour la fraction disponible sur le plateau continental.
Pendant la période 1984-92, les débarquements moyens annuels ont été de 8 000 tonnes pour les sparidés, et de 20 000 tonnes pour les autres espèces, soit 28 000 tonnes au total. C'est approximativement le tiers de l'estmation précédente. Cette moyenne est tombée à 13 000 vu, d'un changement dans la stratégie des flottilles se traduisant par un intérêt moindre pour le poisson et par une augmentation du taux de rejet. A partir de 1988, le taux d'exploitation a peutêtre diminué avec la baisse de l'effort de pêche exercé sur les céphalopodes.
Avec les données disponibles, on ne peut évaluer le potentiel de stock. Malgré les conclusions en partie divergentes auxquelles conduit l'analyse des différentes informations, il paraît peu probable que l'exploitation se soit intensifiée au cours de la dernière décennie. Toutefois, aucun signe d'amélioration de l'état du stock n'a été décelé dans les données étudiées.
3.2.2.3 - Besoins de recherche
Même si les stocks de poisson ont actuellement une importance moindre dans la pêche mauritanienne, cette part n'est pas négligeable. Elle est susceptible de croître dans l'avenir, notamment si l'état du stock de céphalopode se dégrade. Il est donc important de vérifier rapidement la part d'un changement dans les stratégies de pêche de commercialisation, et de l'intensité de la pêche, dans le déclin des débarquements. Pour cela, les investigations ou améliorations suivantes sont nécessaires:
entreprendre une enquês de la profession, afin de vérifier les changements dans les stratégies de pêche, dans la nature (espèces) et le volume des rejets de poissons dans la pêche chalutière, et si des pêches significatives n'échappent pas au système statistique (captures secondaires des flottilles spécialisées et des chalutiers pélagiques);
analyser finement les statistiques commerciales, par groupes d'espèces et espèces, par flottilles (y compris la pêche artisanle), et secteurs;
traiter en temps réel les campagnes du ‘N’Diago' (campagnes 1993);
comparer systématiquement sur une même échelle l'évolution des indices d'abondance ainsi calculés (comme cela a été indiqué pourles céphalopodes); cette analyse pourrait déboucher sur la définition d'une mailleure stratégie de suivi de l'état des stocks démersaux;
compléter éventuellement cette analyse par un travail ponctuel d'échantillonnage et de mensuration de quelques espèces choisies (prospections scientifiques et débarquements commerciaux), afin d'obtenir rapidement des indications sur le taux d'exploitation des principaux stocks de poisson à partir de l'examen de leurs distributions de fréquence;
consulter un spécialiste des estimations de biomasse par prospection pour évaluer l'efficacité de la méthode et, si la conclusion est positive, définir les procédures d'amélioration des plans de campagne;
mettre en place un programme de suivi de l'activité de bateaux appartenant aux principales flottilles distunguées selon leur stratégie de pêche, et d'échantillonnage biologique visant à suivre la structure de taille des principales espèces exploitées; l'échantillonnage biologique fournirait des renseignements précieux sur l'état des principaux stocks.
3.2.3.1 - Merlus
Information sur la pêcherie
Depuis plusieurs dizaines d'zaines d'années, une flottille de chalutiers étrangers, essentiellement espagnols, exploite troiis espèces de merlu: le merlu européen, le merlu sénégalais et le merlu noir sur le plateau et le talus continental (Bravo de Laguna, 1985; Wysokinski, 1986; Chavance et Girardin, 1991). Les espèces ne sont pas distinguées dans les statistiques. Les merlutiers espagnols opèrent à partir de Cadiz. Avant 1990, la flottille était composée de chalutiers de pêche fraîche (“bous”). A partir de 1990, des congélateurs sont venus renforcer la flottille, la rendant ce faisant plus hétérogène (tabl.11). En 1990, les marées étaient de 17 jours environ, et l'activité annuelle totale de chaque bateau de 135 jours de pêche. Le maillage des chaluts est de 60 mm.
| Classe | 150-200 | 200-250 | 250-300 | 300-350 | 350-400 | 400-450 | 450-500 | Total |
|---|---|---|---|---|---|---|---|---|
| Poucentage | 7,7 | 10,2 | 33,3 | 20,6 | 15,4 | 2,6 | 10,2 | 100,0 |
La pêche s'effectue tout le long de la côte mauritanienne, entre 140 et 750 m. On considère qu'entre le cap Blanc et le cap Timiris, les trois quarts des captures sont constituées de merlu sénégalais et, qu'entre le cap Timiris et Saint-Louis cette espèce serait un peu abondante. Durant la période froide, la flottille concentre ses activités dans le secteur nord où le merlu sénégalais est saisonnièrement plus abondant (en relation probablement avec le déplacement du front thermique vers l'équateur).
1 - Cette partie du rapport final est pour l'essentiel basée sur le manuscrit de MM.M. Diop et bU.Lowenberg..
Statistique de pêche
Jusqu'en 1986, les statistiques espagnoles n'étaient pas déclarées par ZEE, mais par divisions statistiques du COPACE (Ramos y Fernandez,1992). La ZEE mauritanienne ne couvre qu'une partie des divisions COPACE 34.3.1 et 34.1.3. Le Groupe a analysé les données relatives à la période s'étendant de la fin de la période des “Accords Bilatéraux” (1983–1987) et couvrant toute cell des “Accors Communautaires” (1987–1991). Pendant la période 1983– 1992, les captures annuelles de merlus (Espagne et Mauritanie) ont oscillé entre 6 580 et 12 750 tonnes (tabl.12). Les rendements ont varié, sans tendance nette, entre 2,2 et 4,0 tonnes/jour de pêche. Les informations disponibles sur la composition spécifique des captures de la flottille espagnole (Ramos y Fernandez, 1992) sont données dans le tableau 13.
| Année | Captures annuelles(tonnes) | Rendement (t/jour pêche) merlutiers | Effort corrgé (jour pêche) | |||
|---|---|---|---|---|---|---|
| Merlutiers | Chalutiers pélagiques | Chautiers de fond | Total | |||
| 1983 | 12754 | 12754 | 3,99 | 3196 | ||
| 1984 | 8 295 | 8 295 | 2,38 | 3 485 | ||
| 1985 | 6 580 | 6 580 | 2,17 | 3 032 | ||
| 1986 | 6522 | 3287 | 148 | 9957 | 3,15 | 3161 |
| 1987 | 8141 | 546 | 100 | 8787 | 3,23 | 2720 |
| 1988 | 9474 | 1007 | 100 | 10581 | 3,29 | 3216 |
| 1989 | 10163 | 247 | 10410 | 2,90 | 3590 | |
| 1990 | 11210 | 139 | 32,1 | 11381,1 | 2,24 | 5081 |
| 1991 | 9335 | 506,5 | 9841,5 | 3,36 | 2935 | |
| 1992 | 9086 | 7,3 | 9093,3 | 2,48 | 3667 | |
| Espèces | 1984 | 1985 | 1986 | 1987 | 1988 | 1989 | 1990 | 1991 | 1992 |
|---|---|---|---|---|---|---|---|---|---|
| Merlus | 8295 | 6580 | 6522 | 8141 | 9474 | 10163 | 11210 | 9335 | 9079 |
| Dentés profonds | 697 | 257 | 407 | 203 | 305 | 262 | 222 | 11 | - |
| Diagramme | - | - | - | - | - | - | - | 3 | - |
| Pageot | - | - | - | - | - | - | - | 15 | - |
| Congre | - | - | - | - | - | - | - | 100 | - |
| Baudroies | - | - | - | - | - | - | - | 2 | 2 |
| Langouste rose | 348 | 266 | 86 | 55 | 22 | 23 | 13 | 0 | 0 |
| Geryon | - | - | - | - | - | - | - | - | 16 |
| Crevettes | |||||||||
| - alistado | - | - | - | - | - | - | - | - | 18 |
| - gamba | - | - | - | - | - | - | - | - | 2 |
| - langostino | - | - | - | - | - | - | - | 0 | |
| - carabinero | - | - | - | - | - | - | - | - | 0 |
| Divers crustacés | - | - | - | - | - | - | - | 12 | 153 |
| Céphalopodes | - | - | - | - | - | - | - | - | 3 |
| Divers | 1203 | 1731 | 806 | 832 | 1078 | 958 | 1620 | 1810 | 1006 |
Biomasses minimales et potentiel de capture
Sur la base de prospections réalisées par un navire allemand, un Groupe de travail précédent avait estimé en 1984 la biomasse absolue de merlus présente sur le talus contiental mauritanien à 13 000 tonnes en 1982, et à 23 000 tonnes en 1983 (FAO, 1986). A partir des prospections du ‘N'Diago’, la biomasse de la fraction juvénile de merlus a été estimée à 8 600 tonnes pour l'année 1982, et à 2 200 tonnes l'année suivante. La biomasse totale instantanée de merlus (adultes + juvéniles) présente dans la ZEE mauritanienne aurait donc été de 21 600 tonnes en 1982, et de 25 200 tonnes l'année suivante. La convergence des estimations globales de ces deux prospections n'est qu'apparente, puisque les deux estimations diffèrent sur la part relative des juvéniles et des adultes.
Deux estimations du potentiel de merlus de la ZEE mauritanienne ont été publiées:
13 200 tonnes, soit le tiers du potentiel de 40 000 tonnes estimé pour l'ensemble des zones COPACE 34.3.1 et 34.3.3 (Josse et Garcia, 1986);
10 000 tonnes, déduit de l'examen des statisques espagnoles et mauritaniennes sur la pêche commerciale pour la période 1983–1992 (Ramos y Fernandez, 1992).
Un réexamen des données relatives à période 1983–92 conduit aux observations suivantes:
la stabilité de l'effort de pêche espagnol pendant cette période (tabl. 12) empêche d'évaluer le stock à l'aide du modèle global; le mélange d'espèces à croissance différente dans les statistiques ne permet pas non plus l'utilisation d'un modèle analytique; enfin, les déplacements verticaux des merlus entraînent une sous-estimation des biomasses mesurées par prospection;
au cours de la période, les captures ont oscillé entre 6 600 et 12 800 tonnes, et les pue de la flotte espagnole ont fluctué sans tendance nette;
même si on ne peut les expliquer avec l'information disponible (recrutement exceptionnel, données erronées), les fortes prises déclarées par l'URSS au milieu des années 70 (80 000 à 100 000 tonnes) ne peuvent être ignorées.
Compte tenu de la stabilité des pue, des bonnes prises (9 000–11 000 tonnes) maintenues au cours des dernières années (1989–92), et de la faible mortalité exercée sur les juvéniles par les chalutiers spécialisés (Ramos y Fernandez, 1992), on peut estimer que les concentrations de merlus présentes dans la ZEE mauritanienne pourraient supporter de façon régulière un prélèvement au moins égal à la première estimation (13 000 tonnes). Ce chiffre n'est pas un potentiel maximal de capture, lequel ne peut pas être évalué avec l'information disponible. Il ne correspond pas non plus à la production d'un ensemble déterminé de populations car la répartition des populations exploitées dans la ZEE mauritanienne déborde ce secteur (la structure et la distribution des populations ouest-africaines de merlus restent mal connues).
A condition d'être accompagné de mesures garantissant que les données nécessaires aux évaluations soient collectées, déclarées et analysées, un accroissement contrôlé de l'effort de pêche est envisageable. Simultanément, les prises accessoires dont on soupçonne la pêche pélagique, et les captures de juvéniles effectuées accessoirement par les flottilles de chalutiers opérant sur le plateau, devraient être estimées. Une intensification de la pêche par paliers permettrait de préciser ces premières conclusions.
3.2.3.2 - Autres espèces
Le Groupe de travail de 1985 avait estimé à 71 000 tonnes le potentiel de l'ensemble “autres espèces profondes”, constitué par les poissons du talus à l'exclusion apparemment des merlus. Il avait admis en première approximation que ce potentiel était égal à la moitié du potentiel démersal du plateau (incluant les céphalopodes). La meilleure connaissance du peuplement halieutique du talus apportée par les campagnes de chalutage expérimental effectuées plus au sud dans le golfe de Guinée (superficie, nature et abondance relative des espèces) indique que ces hypothèses étaient trop optimistes. En effet, l'importance des espèces actuellement commercialisées (sparidés profonds essentiellement) par rapport aux espèces de valeur commerciale faible ou nulle (munidae, poissons de petite taille, crevettes non commercialisables,…) est nettement plus faible sur le talus que le plateau. L'évaluation de leur potentiel devrait être reprise. Dans l'attente, les captures de poissons commercialisables (hors merlus) du talus pourraient être limitées à 10 à 15 000 tonnes.
![]()
![]()
![]()