![]()
![]()
![]()
3.2.4.1 - Crevettes côtières et profondes
Description de la pêcherie
a) Rappel historique
La pêche de la crevette a démarré en Mauritanie à la fin des années 1960 (FAO, 1990a; Chavance et Girardin, 1991; Diop, 1991; Anon., 1993). Ce n'est qu'à partir de 1987 que des statistiques ont été obtenues sur la pêche crevettière. Deux flottilles distinctes se sont consacrées à cette pêche:
une première, composée d'unités mauritaniennes disparates (affrétées et/ou sous licence) et débarquant en Mauritanie; elles ont cessé leur activité en 19902;
la seconde, plus importante, composée de chalutiers congélateurs espagnols, opérant depuis de nombreuses années dans la sous-région; pêchant sous licence dans la ZEE mauritanienne, cette flottille débarque sa production en Espagne.
Alors que la première ne pêchait que le stock de crevette côtière (Pencoeus notialis), la seconde flottille exploite alternativement ce stock et, sur le talus, celui de crevettes profondes. Cette dernière ne fournit pas de statistiques séparées des efforts qu'elle exerce sur les deux stocks.
b) La flottille espagnole et les espèces cible
La durée de validité des licences est d'un an (juillet à juin). La structure de la flottille pour la saison 1992/93 est donnée dans le tableau 14. On constate l'existence de deux groupes de bateaux dont il serait intéressant de comparer l'origine et les performances.
| Tonnage (TJB) | 50–100 | 100–150 | 150–200 | 200–250 | 250–300 |
|---|---|---|---|---|---|
| Pourcentage | 20 | 40 | 12 | 20 | 8 |
Au cours de la saison de pêche, les chalutiers recherchent alternativement trois groupes d'espèces:
les ‘langostinos’ (Penaeus notialis et P. kerathurus), sur les fonds de 12 à 60 mètres, entre 19 et 20°N;
la ‘gamba’ (Parapenaeus longirostris), sur des fonds de 170 à 300 mètres, entre 17°20 et 18°50'N;
l'alistado' (Aristeus varidens), sur le talus entre 350 et 700 m et les latitudes 16° et 20° N.
Des informations sur la biologie et l'écologie de ces espèces ont été synthétisées dans Chavance et Girardin (1991).
Statistiques disponibles
Elles portent sur la période 1987–1992 (tabl. 15). Les données de capture, d'effort et de rendement ont été ventilées par mois en suivant la procédure décrite dans le rapport de la deuxième recontre CNROP/IEO sur les crevettes (Anon., 1993).
1 -Cette partie du rapport final est basée sur la contribution de MM. M. Diop et U. Löwenberg.
2 -Il serait intéressant d'analyser les raions de l'arrêt de cette activité.
| Années | 1987 | 1988 | 1989 | 1990 | 1991 | 1992 | |
|---|---|---|---|---|---|---|---|
| Nombre de bateaux* | 42 | 44 | 37 | 32 | 21 | 20 | |
| Tonnage total (TJB)* | 9586,9 | 9767,3 | 8271,7 | 6387,3 | 3772,6 | 2958,4 | |
| Temps total de peche (jours) | 4060 | 9100 | 10691 | 9018 | 6570 | 5648 | |
| Parapenæneus | Capture (t) | 1002,6 | 2182,2 | 2382,4 | 1470,9 | 651,6 | 487,9 |
| longirostris | Rendement (kg/j) | 216,9 | 239,8 | 222,8 | 163,1 | 99,2 | 86,4 |
| Penæus | Capture (t) | 550,9 | 1 033,2 | 1 081,9 | 1 007,9 | 484,7 | 469,6 |
| notialis | Rendement(kg/j) | 135,7 | 113,5 | 101,2 | 111,8 | 73,8 | 83,1 |
| Aristeus | Capture(t) | 82,9 | 285,4 | 314,1 | 253,2 | 219,6 | 236,1 |
| varidens | Rendement(kg/j) | 20,4 | 31,4 | 29,4 | 28,1 | 33,4 | 41,8 |
| Autres | Capture (t) | 23,4 | 50,5 | 53,7 | 24,2 | 90,7 | 137,2 |
| crevettes | Rendement (kg/j) | 5,8 | 5,6 | 5,0 | 2,7 | 13,8 | 24,3 |
| Total | Capture(t) | 1659,8 | 3551,4 | 3832,2 | 2756,2 | 1446,6 | 1330,8 |
| crevettes | Rendement(kg/j) | 378,8 | 390,3 | 358,4 | 305,7 | 220,2 | 235,6 |
Captures
Les captures totales déclarées ont fortement diminué de 1989 à 1992 (fig. 11). Pendant la même période, la proportion de Penæus notialis est restée stable (entre 27 et 33 %); celle de Parapenæus longirostris a beaucoup décliné de 58% à 30%, tandis que celle d'Aristeus varidens montait de 8% à 14%, et celle de crabe rouge profond (Geryon maritae) de 5% à 18 %. Comme, simultanément, l'effort a nettement diminué (de près de moitié pour les jours de pêche, et des deux tiers pour le tonnage total; tabl. 15), ces évolutions traduisent une réduction de l'effort de pêche plus forte sur Parapenæus longirostris (partie haute du talus), que sur les espèces de la partie la plus profone du talus où l'effort aurait le moins chagé, et sur Penæus notialis (partie côtière du plateau).
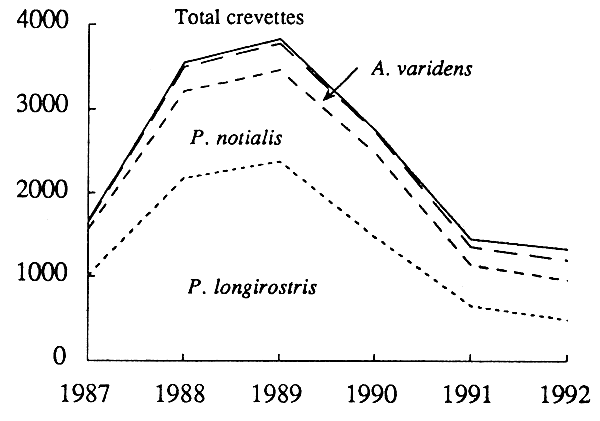
Figure 11 - Pêche espagnole de crevettes: évolution des captures cumulées
Prises accessoires d'espèces côtières par la flottille espagnole
Une estimation de la composition des captures des captures dans la zone côtière (% par grands groupes) est disponible pour la flottille espagnole durant la période 1991–93 (tabl. 16).
| Crevettes | Autres crustacés | Poissons démersaux | Céphalopodes | Poissons pélagiques |
|---|---|---|---|---|
| 11% | 6% | 58% | 10% | 15% |
Les captures des espèces cible sont inférieures ou équivalentes à celles d'autres groupes d'espèces spécialement recherchées par d'autres flottilles (poissons démersaux et céphalopodes). Les prises accessoires de flottilles spécialisées peuvent donc être respinsables d'une part significative du taux d'exploitation exercé sur des stocks accessoires, de poissons du plateau notamment. En outre, le fait que l'activité des flottilles n'est pas régulé par groupes d'espèces cible, donc par secteurs, laisse la porte ouverte à des pratiques illicites.
Effort de pêche
Pour l'ensemble des stocks de crevettes de la ZEE mauritanienne, une estimation groissière (compte tenu de l'hétérogénéeité de la pêcherie) de l'effort global annuel (tous flottilles) peut être calculée en divisant les captures totales de crevettes par le rendement moyen annuel de la flottille espagnole (tabl. 17). Après être montée de 5 500 à 11 000 jours de pêche entre 1987 et 1989, cette estimation est redescendue en 1992 à son niveau de 1987. La répartition de l'effort de pêche entre les diférents stocks n'est pas connue.
| Année | Capturres (tonnes) | Pue (Espagne, Kg/j. de pêche) | Effort estimé (jours de pêche) | |||
|---|---|---|---|---|---|---|
| Mauriranie | Espagne | Autres | Total | |||
| 1987 | 407 | 1 660 | 0 | 2 067 | 379 | 5 454 |
| 1988 | 214 | 3 551 | 0 | 3 765 | 390 | 9 654 |
| 1989 | 78 | 3 832 | 0 | 3 910 | 358 | 10 922 |
| 1090 | 0 | 2 750 | 0 | 2 750 | 306 | 8 997 |
| 1991 | 0 | 1 447 | 0 | 1 447 | 220 | 6 577 |
| 1992 | 0 | 1 331 | 29 | 1 360 | 236 | 5 763 |
Rendements
Le rendement global a fortement baissé (de 44 %) entre 1988 et 1992.
En ce qui concerne les variations saisonnières, on observe deux pics de rendement pour la crevette rose (gamba): un entre mars et juin, l'autre secondaire entre octobre et décembre. Pour la crevette baanche (‘langostino’), il n'existe qu'une période d'abondance, de septembre à janvier; de mars à mai, le rendement est faible.
Potentiel exploitable
La répartition verticale des stocks au-dessus du fond étant supérieure à la hauteur du chalut, les campagnes de chalutage qui ne sont plantifiées pour tenir compte de ce comportement ne permettent pas d'estimer convenablement la biomasse des stocks profonds. Ce type de réparatition a été observé pour plusieurs stocks de crevettes profondes et de merlus.
Pour se faire une idée du potentiel de ces stocks, on peut tenter de comparer les chiffres dont on dispose à ceux d'autres stocks, a priori comparables, dont le potentiel est connu. Auparavant, on rappellera que, schématiquement, le potentiel de production de stocks d'une même espèce peut différer en fonction:
du niveau moyen du recrutement, lequel dépend principalement de la capacité biotique (mais pas nécessairement trophique) des structures hydrodynamiques colonisées pendant la (ou les) phase (s) critique (s) du recrutement (§3.1.3);
de la variabilité du recrutement, fonction elle-même de la variabilité interannuelle de cette capacité biotique;
des interactions biologiques (prédation des juvéniles par des espèces exploitées), dont les effets sont généralement plus faible que ceux des processus précédents;
des phénomènes de densité-dépendance affectant la phase recrutée; toutefois, ceux-ci ont généralement des effets encore moindres; à cause de la prépondérance des phénomènes déterminant le niveau de recrutement, on réalise maintenant que des populations adultes d'une même espèce peuvent avoir hors effets de la pêche, des densités moyennes différentes même lorsqu'elles habitent des environnements a priori comparables (par exemple, les stocks de gadidés de mer du Nord et de mer d'Irlande); ce constat limite la validité d'évaluations basées sur la comparasion des aires et des densités des populations adultes, qui admettent implicitement que le potentiel est principalement déterminé par la capacité trophique du milieu pour la fraction recrutée du stock;
du taux d'exploitation.
Un stock de crevette Parapenoeopsis longirostris est pêché, entre 150 et 400 m, dans les parties septentrionale et centrale de la ZEE sénégalaise. De ce stock, on connaît:
la flottille de pêche: il s'agit également de crevettiers espagnols dont les caractéristiques sont similaires à celles des navires exploitant le stock mauritanien (ce sont souvent les mêmes bateaux);
l'aire de la zone de pêche (1700 km2);
le rendement global moyen (toutes espèces de crevettes condondues): 40 kg/heure de pêche, soit kg/jour (pour 14 heures de pêche par jour), en 1988 et 1989;
une valuer minimale du potentiel de capture: les prises ayant varié entre 2 500 à 3 000 tonnes pendant plusieurs années, on peut, compte tenu de la durée de vie courte (2 ans) de l'espèce, admettre qu'à recrutement constant son potentiel est au moins égal à ce niveau de capture.
Des concentrations exploitées en Mauritanie, on connaît:
la superficie de l'aire de pêche: 3 200 km2 (N/O ‘Lublino’, 1990);
le rendement global moyen à la même époque: 380 – 390 kg/jour; une partie de cette différence (25 % inférieur) s'explique par le monde de mesure de l'effort de pêche: au Sénégal, il est égal au temps de pêche effectif, alors qu'en Mauritanie, il est égal au temps de mer qui ni tient pas compte des temps morts consacrés au transfert entre les fonds côtiers et les fonds hauturiers, et aux arrêts en mer (croches, mesure des funes, pannes, …).
On ignore, par contre, tout des autres sources d'erreur. Si l'on a affaire à deux populations distinctes1 et si le potentiel des deux stocks est dans le rapport de l'aire de distribution de leur phase recrutée - hypothèse que n'infirme pas la comparaison des rendements -, celui du stock mauritanien de Parapencoeopsis longirostris serait supérieur à celui du Sénégal et, donc, aux captures annuelles réalisées dans la ZEE mauritanienne pendant la période 1987 – 92 (<2 400 tonnes).
Sur la base de ces informations, on pourrait augmenter par paliers l'effort total annuel autorisé, en s'assurant que les chalutiers remplissent, comme ils en ont l'obligation, les journaux de pêche (statistiques journalières indiquant les secteurs - profondeur - de pêche, les captures par espèces et le temps de pêche effectif).
En ce qui concerne Pencoeis notialis, la crevette côtière, on remarque que les prises ont varié entre 500 et 1 000 tonnes pendant la période 1987–92. Cette période étant supérieure à la longévité de l'espèce (18 mois), et comme aucune baisse de la capture totale annuelle n'a été observée pendant cette période, on peut admettre que la biomasse accumulée a été épuisée, et que le potentiel est au moins égal, et pourrait être supérieur à ce niveau de production. Pour cette espèce également, on pourrait maintenir la pêche pendant quelques années à un niveau proche de cette limite supérieure (en en suivant les effects).
1 - Si l'espèce est pêchée tout ai long du plateau, la structure des populations n'est pas connue.
Priorités de recherche
Il n'est pas possible, avec les statistiques disponibles, d'aller au-delà de ces premières indications sur le potentiel des stocks de crevettes. La première priortié, parce qu'elle est plus facilement réalisable, porte sur la réalisation de modèles globaux pour les principaux stocks. Pour cela, la collecte de statisques de capture et d'effort de êche fines est indispensable. Les journaux de pêche des chalutiers donnent en principe la compostion des captures par esèces dans la ZEE mauritanienne. Ces journaux sont disponibles depuis 1991. Leur contenu devrait être analysé.
De façon générale, les statistiques doivent distingue:
les secteurs: côtier et talus (et dans ce cas par tranches de profondeur);
les captures: par espèces ou principaux groupes d'espèces;
l'effort de pêche effectif (par jour, sinon par marée) par secteurs et, si possible,par espèces cible pour la pêche sur le talus.
Même si les stocks ne peuvent être évalués avec la précision souhaitable par cette méthode, ces données apporteront une information précieuse et permettront de suivre l'évolution de la pêche sur les différents stocks.
Un progrès similaire est nécessaire dans la régulation de la pêche des différentes flottilles. Les capacités de capture et les activités des flottilles devraient être ajustées par principaux groupes d'espèces cible.
3.2.4.2 - Langoustes
Langouste rose (Palinurus mauritanicus)
a) Flottille
Jusqu'en 1991, ce stock faisait l'objet d'une pêche spécialisée de la part d'une flottille française de caseyeurs à viviers et de quelques glaciers pourtgais. les navires travaillaient par marée de 7 à 150 jours. Le pêche durait toute l'année. les statisques sur cette pêcherie sont donnés dans le tableau 18. On ignore si, depuis 1991, la pêche est interrompue (aucune licence), ou si les statistiques ne sont plus transmises.
| Année | Capture | Nombre de navires | Effort | Pue |
|---|---|---|---|---|
| (tonnes) | (jours de pêche) | (kg/jour de pêche) | ||
| 1983 | 840 | 10 | 1585 | 530 |
| 1984 | 737 | 10 | 1657 | 445 |
| 1985 | 791 | 10 | 1568 | 504 |
| 1986 | 853 | 11 | 1874 | 455 |
| 1987 | 943 | 23 | 2375* | 397** |
| 1988 | 696 | 24 | 3240* | 215** |
| 1989*** | 303 | 16 | 929 | 326 |
| 1990*** | 213 | 16 | 311 | 687 |
| 1991*** | ? | ? | ? | ? |
* : effort normalisé (capture totale/pue des caseyeurs);
** : pue normalisée;
*** : données incomplèes ou absentes.
b) Potentiel et niveau de capture
En 1990, le stock a été estimé à 800 tonnes (Diop et Kjoemiakine,1990). Cette estimation reste valide. Pour une exploitation exploitation convenable, la flottille ne devrait pas dépasser l'équivalent de 12 caseyeurs de type français (Diop et Kojemiakine, 1990). En effet, après plusieurs années de surexploitation, le stock ne s'est pas encore reconstitué. Si la pêche devait reprendreen supposant qu'elle ait cessé -, elle devrait se faire exclusivement au casier. En effet, étant peu sélectifs, les filets maillants utilisés jusqu'en 1989 capturaient des proportions élevées de juvéniles.
Langouste verte (Panulirus regius)
a) Flottile
La langouste verte est pêchée par des pirogues de type sénégalanis et, secondairement, par des piorgues plastique opérant surtout à partir de Nouadhibou. Le nombre de pirogues et de filets employés est difficile à estimer. En 1987,86 embarcations péchant exclusivement la langouste, et 6 autres la pêchant avec d'autres espèces, ont été recensées. La pêche se fait à l'aide d'un filet dormant de 400 m de longueur, de 8 m de chute, et de maille (étirée) de 100 mm (Chavance et Girardin,1991).
b) Statistiques
Les statistiques disponibles sont présentées dans le tableau 19.
| Année | Capture(tonnes) | Effort(sorties) | Pue(kg/sortie) | ||
|---|---|---|---|---|---|
| Stock nord | Stock sud | Total | Stock sud | ||
| 1984 | 194,7 | 35,9 | 230,5 | ||
| 1985 | 146,0 | 21, | 167,1 | ||
| 1986 | 249,1 | 8-12 | 257-261 | ||
| 1987 | 201,7 | 23,2 | 224,7 | 2489 | 9,32 |
| 1988 | 98,8 | 7,3 | 104,1 | 712 | 10,25 |
| 1989 | 116,7 | 2,7 | 119,4 | 121 | 22,31 |
| 1990 | 4,2 | 38,0 | 42,2 | ||
| 1991 | 49,0 | 26,7 | 75,7 | 1050 | 25,4 |
| 1992 | 39,7 | ||||
c) Potentiel et niveau de capture
On distingue deux stocks: un au nord, l'autre au sud. Les données sont insuffisantes pour estimer leur potentiel, car on ne dispose de mesure d'effort que pour le seul stock sud et sur cinq années seulement. Une estimation ancienne du potentiel du stock nord (cap Blanc - Villa Cisneros) donnait une valeur de 300 tonnes pour une longueur de côte de 333 km. Ce stock a vraisemblablement été surexploité par le passé (prises comprises entre 100 et 250 tonnes). Bien que les prises aient baissé de 1990 à 1992, ce stock montre toujours des signes de surexploitation. La taille modale a aussi diminué pendant la même toujours des signes de 1991: 22–23 cm; 1992: 18–21 cm). Pour la fraction comprise le long du littoral mauritanien, il est prudent de limiter les captures à 50 tonnes.
Par analogie avec le stock nord, le potentiel du stock sud pourrait être de l'ordre de 175 tonnes. Ce stock a été surexploité pendant longtemps. L'augmentaiton des pue (tabl. 19) doit être interprétée avec prudence, d'une part, parce que cette hausse ne signifie pas que le stock soit déjà reconstitué et, d'autre part, parce que les tailles moyennes des captures sont toujours inférieures àce qu'elles étaient lorsque le stock n'était pas surexploité.
3.2.4.3 - Crabe profond (Geryon maritae)
Pêche
Les connaissances disponibles sur l'écologie et la biologie de cette espèce ont été présentées (Chavance et Girardin, 1991; Diop et Kojemiakine, sous presse). En Mauritanie, le crabe profond ne fait pas l'object d'une pêche spécialisée. Il est pêché en même temps que les crevettes profondes par la flottille crevettière espagnole (§ 3.2.4.1). Les débarquements de Geryon maritae- déclarés par l'Espagne pour la période 1987 – 1992 sont donnés dans le tableau 20. Les déclaration doivent être multipliées par 1,6 car elles portent sur les pattes de crabe. L'augmentaiton du rendement à partir de 1990 s'explique par une concentration de l'effort dans la zone plus profonde (§ 3.2.4.1).
| Année | Capture(tonnes) | Effort(jours de pêche) | Pue(kg/jp) |
|---|---|---|---|
| 1987* | 64,9 | 4 060 | 16,0 |
| 1988 | 174,5 | 9 100 | 19,2 |
| 1989 | 187,2 | 10 691 | 17,5 |
| 1990 | 234,2 | 9 018 | 26,0 |
| 1991 | 288,4 | 6 570 | 43,9 |
| 1992 | 289,4 | 5 648 | 51,2 |
* :deuxième semestre seulement.
Potentiel et niveau de capture
En admettant que le crabe profond représenterait entre 4 et 6 % du potentiel halieutique du talus, cette espèce pourrait produire entre 1 800 et 4 000 tonnes de façon continue (Diop et Kojemiakine, sous presse). Ce chiffre doit être réduit de la fraction juvé nile qui est très importante (on l'estime à 90 %). La part de gros crabes actuellement exploitable serait donc plus faible (200–400 tonnes peut-être). Parce que son taux de renouvellement est faible, l'esèce ne peut supporter qu'un taux d'exploitation modéré. Les captures actuelles sont proches de cette première estimation. Sa pêche devrait se faire au casier, plus approprié que le chalut.
3.2.4.4 - Bivalves(praires)
Il existe deux espèces de praires dans les eaux mauritaniennes: Venus verrucosa dans la baie du Lévrier, et Venus rosalina au sud du cap Blanc, devant le banc d'Arguin. La biologie et l'écologie de ces espèce sont décrites dans Chavance et Giradin (1991).
Le potentiel de ces stocks a été estimé entre 500 et 1000 tonnes pour Venus verrucosa, et à 300 000 tonnes pour Venus rosalina. Le stock de Venus verrucosa est actuellement exploité par un navire de 16 m et les prises déclarées ces dernières années tournent autour de 500 tonnes. Compte-tenu de la grande longévité de cette espèce (environ 10 ans), son taux de renouvellement est faible. Il est donc recommandé de ne pas accroître l'effort de pêche au dessus du niveau actuel (1 bateau).
Le stock de Venus rosalina n'est pas encore exploité. Son exploitation se heurte à des problèmes de traitement et de commercialisation. En outre,sa pêche se fait à la drague. En grattant le fond, cet engin peut altérer les frayères. Les conquillages entrent aussi dans l'alimentation du poulpe. Ces effets de l'exploitation des praires sur la productivité des stocks halieutiques, le poulpe notamment, ne sont pas connus. La détermination de l'effort de peche à ne pas dépasser pour l'exploitation des praires doit donc tenir compte d'aspects autres que le potentiel de renouvellement de ce stock.
3.3.1.1 - Stocks
L'aire de répartition des stocks de petits pélagiques côtiers pêchés dans la ZEE mauritanienne couvre approximativement les deux divisions COPACE 34.1.3 (19°N à 26°N) et 34.3.1 (9°N à 19°N),c'est-à-dire les secteurs sénégalais, mauritanien et saharien.
Cinq groupes d'espèces intéressent principalement la Mauritanie: les chinchards européen (Trachurus trachurus), africain (T.trecoe) et jaune (Decapterus punctatus); les sardinelles ronde (Sardinella aurita) et plate (S.maderensis); la sardine (Sardina pilchardus); et le maquereau (Scomber japonicus). Elles représentent jusqu'à 85 % des captures. Le sabre (Trichiurus lepturus) et l'anchois (Engraulis encrasicolus) sont les principales espèces secondaires.
Par leur affinité écologique, ces espèce peuvent être séparées en deux groupes. La sardine, le chinchard européen et le maquereau, à affinité tempérée, intéressent les ecteurs mauritanien et saharien. Le chinchard africain, le chinchard jaune et les sardinelles, à affinité tropicale, sont concentrées dans les secteurs mauritanien et sénégalais. Ces deux ensembles occupent saisonnièrement le secteur mauritanien, où alternent dans l'année des conditions subtempérées et troicales. La structure,les aires de ponte et les schémea de migration des polulations des différentes espèce sont mal connus.
3.3.1.2 - Flottilles
Dans leur aire de répartition, ces espèces font,ou ont fait,l'objet d'une pêche dirigée de la de deux ensembles de flottilles:
les flottilles piroguières artisanales: leur activité est localisée, dans la bande littorale, d'une part, et au Sénégal et au sud de la Mauritanie, d'autre part; elles utilisent la senne de plage, la senne tournante,et des filets maillants;
les flottilles industrielles pêchent au chalut pélagique et à la senne tournante,généralement plus au large, dans les secteurs mauritanien et saharien.
Les captures totales de petits pélagiques dans la sous-région ont fluctuéentre 700 000 tonnes en 1970 et 1 900 000 tonnes en 1990. Dans la ZEE mauritanienne, les captures ont varié au cours des 15 dernières années entre 210 et 470 000 tonnes, autour d'une moyenne de 380 000 tonnes (tabl.21). Depuis la fin de l'année 1992, l'activitédes flottilles hauturieères des pays de l'Europe de l'est est fortement perturbée par les changements survenus dans ces pays ainsi que par l'effet de la dévaluation du france CFA sur la demande solvable des pays du golfe de Guinée de la zone franc4. Des inenrtitudes sur l'avenir de cette pêche en résultenet.
3.3.1.3 - Statistiques de pêche
Dans leur ventilation comme dans leur couverture, les données disponibles ne reflètent pas correctement la structure de la pêche des petits pélagiques. Ces insuffisances empêchent d'estimer les efforts effectivement exercés par les diverses flottilles sur les différentes espèces et leurs populations (§3.1.1.2). Les évaluations doivent donc porter sur des ensembles plus vastes que la ZEE mauritanienne.
2 - Cette partie du rapport reprend pour l'essentiel le raport de M.M'Bereck S.
3 - Cette partie est basée pour l'essentiel sur la contribution de MM.S. Bairane et O/Taleb Sidi.
| Espèces | 1979 | 1980 | 1981 | 1982 | 1983 | 1984 | 1985 | 1986 | 1987 | 1988 | 1989 | 1990 | 1991 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S.pilchardus | 31 | 72 | 8 | 0 | 75 | 48 | 58 | 84 | 35 | 37 | 40 | 34 | 70 |
| S.aurita | 71 | 129 | 118 | 41 | 54 | 53 | 69 | 39 | 47 | 43 | 51 | 79 | 69 |
| S.maderensis | 2 | 15 | 4 | 2 | 9 | 13 | 25 | 22 | 17 | 15 | 15 | 29 | 15 |
| T.trachurus | 53 | 201 | 34 | 38 | 19 | 41 | 54 | 45 | 85 | 50 | 47 | 33 | 11 |
| T.treca | 45 | 47 | 86 | 74 | 109 | 72 | 71 | 145 | 129 | 132 | 104 | 57 | 101 |
| D.ronchus | 0 | 10 | 22 | 38 | 75 | 58 | 72 | 14 | 13 | 15 | 19 | 22 | 16 |
| S.japonicus | 2 | 9 | 5 | 11 | 45 | 14 | 18 | 29 | 24 | 31 | 19 | 20 | 14 |
| Autres | 4 | 13 | 9 | 70 | 84 | 68 | 88 | 77 | 119 | 80 | 87 | 22 | 86 |
| Total | 207 | 495 | 286 | 274 | 469 | 373 | 454 | 456 | 470 | 403 | 383 | 295 | 381 |
Pour les évaluations, les statistiques ont été regroupées en trois grands ensembles:
ensemble ‘URSS’, qui a opéré dans les zones ‘Mauritanie’ et ‘Sahara’;
ensemble ‘Roumanie’, qui comprend la flottille roumaine et les autres flottilles ayant opérédans la zone ‘Mauritanie’;
ensemble ‘Sénégal’, qui regroupe les flottilles asrtisanale et industrielle opérant dans la zone ‘Sénégal’.
Les statistiques disponibles ont été ventilées par espèces et par flottilles, pour le secteur allant du Sahara au sud de Sénégal, et la période 1979 – 91 (tabl.22).
| Années | Effort nominal | Prises par unité d'effort | ||||||
|---|---|---|---|---|---|---|---|---|
| S.aurita | S.maderensis | T.trachurus | T.treca | D.ronchus | Sc.japonicus | Total | ||
| 1979 | 15 479 | 8,25 | 2,82 | 9,75 | 5,34 | 0,79 | 4,60 | 46,11 |
| 1980 | 26 238 | 7,36 | 1,80 | 12,84 | 4,27 | 0,68 | 2,38 | 42,06 |
| 1981 | 28 386 | 6,38 | 1,95 | 6,02 | 4,66 | 1,09 | 3,25 | 36,03 |
| 1982 | 30 280 | 2,15 | 1,49 | 5,23 | 3,88 | 1,67 | 3,27 | 30,40 |
| 1983 | 34798 | 2,53 | 2,01 | 2,70 | 3,85 | 2,39 | 2,20 | 25,96 |
| 1984 | 30 059 | 2,49 | 2,32 | 3,96 | 3,10 | 2,13 | 2,76 | 29,68 |
| 1985 | 20 818 | 5,23 | 3,73 | 6,72 | 4,76 | 3,71 | 3,01 | 42,95 |
| 1986 | 23 353 | 5,39 | 3,81 | 4,92 | 7,77 | 0,82 | 4,03 | 43,47 |
| 1987 | 30 699 | 5,62 | 3,38 | 4,90 | 5,73 | 0,56 | 2,68 | 42,10 |
| 1988 | 38 570 | 4,76 | 2,65 | 2,59 | 4,67 | 0,50 | 7,19 | 41,09 |
| 1989 | 38 659 | 5,73 | 2,82 | 2,03 | 4,33 | 0,58 | 6,77 | 47,53 |
| 1990 | 50 178 | 5,50 | 2,83 | 1,65 | 2,64 | 0,52 | 2,63 | 37,54 |
| 1991 | 16 015 | 11,53 | 5,88 | 0,94 | 6,40 | 1,17 | 0,93 | 36,69 |
Pour l'application de modèles globaux, seuls des efforts nominaux ont été utilisés. Ce choix est imposé par les incertitudes qui affectent l'estimation d'efforts effectifs, et l'unicité de la pêche de petits pélagiques qui rend difficile l'ajustement séparé des activités des flottilles au potentiel des différentes populations. S'il est insuffisant pour l'évaluation fine de potentiel des populations, le choix d'un effort nominal reste toutefois intéressant tant que, pour des raisons opérationnelles, la pêcherie de petits pélagiques ne peut être aménagée par ensembles plus petits. Parce que la série est la plus complète, les efforts de la flottille roumaine ontété utilisés pour calculer des efforts standardisés.
Des efforts effectifs ont aussi été soumis au Groupe de travail par les chercheurs russes, mais pour la flottille soviétique et les deux espèces de chinchard seulement (tabl. 23).
| Années | Trachurus trachurus | Trachurus trecæe | ||||
|---|---|---|---|---|---|---|
| Moyenne | Erreur standard | Effort effectif | Moyenne | Erreur standard | Effort effectif | |
| 1979 | 87,10 | 16,08 | 1918 | 10,62 | 2,99 | 561 |
| 1980 | 63,82 | 8,43 | 4683 | 11,00 | 2,41 | 1068 |
| 1981 | 81,17 | 10,72 | 2485 | 14,53 | 3,04 | 845 |
| 1982 | 57,17 | 7,59 | 3412 | 9,14 | 2,07 | 683 |
| 1983 | 51,74 | 6,68 | 3145 | 8,25 | 1,62 | 570 |
| 1984 | 55,48 | 7,14 | 3054 | 8,04 | 1,68 | 493 |
| 1985 | 50,32 | 6,30 | 3553 | 8,90 | 1,75 | 626 |
| 1986 | 48,08 | 6,04 | 3246 | 11,81 | 2,26 | 1109 |
| 1987 | 41,80 | 5,25 | 4604 | 18,16 | 3,54 | 705 |
| 1988 | 58,37 | 7,23 | 3329 | 22,60 | 4,49 | 474 |
| 1989 | 53,08 | 6,97 | 2626 | 21,06 | 4,11 | 531 |
| 1990 | 48,68 | 6,32 | 2594 | 23,15 | 4,75 | 447 |
Plusieurs campagnes d'évaluation par écho-intégration ont été réalisées dans la sousrégion: campagnes du‘Louis Sauger’ dans le secteur sénégalais, du ‘N'Diago’ dans le secteur mauritanien, de l'‘Ibn Sina’ dans la zone ‘Sahara’, du ‘Capricorne’ et du ‘Dr Fridtjorf Nansen’ sur l'ensemble de la sour-région.
Si l'on excepte les campagnes de février 1982 et de mars 1989, les estimations de biomasse des petits pélagiques présents dans la zone sénégalaise sont, malgré leur dispersion saisonnière, assez homogènes. Leur moyenne est de 588 000 tonnes (avec un coefficient de variation de 14 %). Les estimations tirées des campagnes du ‘Capricorne’ (1975 – 1977) sont du même ordre.
Dans la zone mauritanienne, en revanche, les estimations du ‘N'Diago’ (tabl. 24) sont plus dispersées. Pour la période 1986–89, l'estimation la plus faible était de 937 000 tonnes, et la plus forte de 2 238 000 tonnes. Ces variations peuvent avoir pour origine les migrations saisonnières que les stocks à affinités sub-tempérée ou tropicale effectuent entre les secteurs voisins et la zone de balancement que constitue le secteur mauritanien.
Au cours de la saison froide 1992, le ‘Dr Fridtjorf Nansen’ a couvert la zone allant du Maroc à la Guinée Bissau (tabl. 24). La biomasse présente dans les ZEE mauritanienne, sénégalaise et gambienne fut évaluèe à 4 millions de tonnes. Cette estimation est supérieure à la somme des plus fortes biomasses mesurées séparément au Sénégal et en Mauritanie. Cette valeur élevée résultait de l'abondance particulièrement forte des sardinelles (ronde en Mauritanie, plate dans la zone sénégambienne); les carangidés, par contre, étaient moins abondants que lors des prospections réalisées par le même navire en 1981, 1982 et 1986.
Malgré leur dispersion, ces estimations donnent l'ordre de grandeur des biomasses présentes dans la sous-région.
Si, à l'avenir, les prospections pouvaient couvrir l'ensemble de l'aire de distribution des populations, les erreurs liées aux migrations saisonnières entre secteurs seraient moindres, et la connaissance de la répartition spatio-temporelle des biomasses améliorée. En permettant une couverture plus rapide de la zone, ces campagnes réduiraient encore les biais causés par le déplacement des concentrations au cours des prospections. Cette condition peut être satisfaite par des campagnes coordonnées à plusieurs navires, nationaux notamment.
| Navires | Espèces | Dates | Secteurs | ||
|---|---|---|---|---|---|
| Sénégal-Gambie | Mauritanie | Sahara | |||
| ‘F.Nansen’ | S. pilchardus | 1992 | 20 | 4 050 | |
| S.aurita | 480 | 1410 | |||
| S.maderensis | 1050 | 560 | |||
| T.trachurus | 440 | 70 | |||
| T.trecæ | 90 | ||||
| D.ronchus | 182 | 30 | |||
| Autres, dont anchois | 240 | ||||
| ‘N'Diago’ | Total petits | Août 86 | 1037 | ||
| pélagiques | Janv.87 | 1855 | |||
| Oct.87 | 1147 | ||||
| 1988 | 2238 | ||||
| 1989 | 937 | ||||
3.3.3.1 - Ajustement et résultats du modèle
Chinchards
Des analyses ont été effectuées sur les deux espèces de chinchard (T. trachurus et T. trecae), séparément d'abord, ensemble ensuite. On rendra seulement compte des résultats, plus cohérents, de la deuxième analyse (fig. 12).
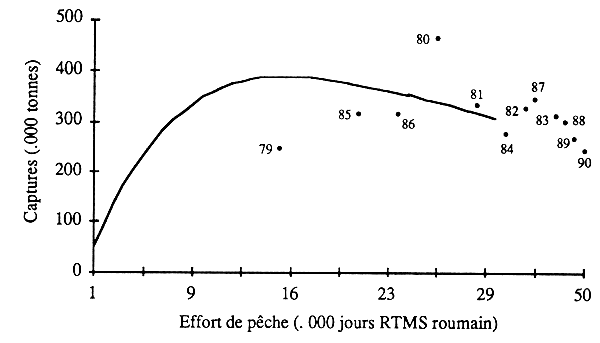
Figure 12 - Modèle global appliquéau stock de chinchards (Trachurus spp.) de la sous-région (période 1977–90; les points représentent les captures annueles réelles).
Ajusté à ces données, le modèle PRODFIT ne donne pas de résultat satisfaisant. L'accroissement apparemment régulier de l'effort de pêche nominal ne se traduit pas par une augmentation de la prise de chinchards. Celle-ci fluctue entre 300 et 400 000 tonnes, sans tendance apparente. L'effort correspondant à la prise maximale équilibrée serait largement dépassé: d'après ce traitement, le stock de chinchards serait surexploité.
II est toutefois possible que l'effort nominal reflète mal la mortalité effectivement exercée par la pêche. Si les navires se sont moins intéressé à ces espèces, leur taux d'exploitation peut être resté plus stable que ne l'indique l'effort nominal. Cette hypothèse est en accord avec l'évolution des efforts effectifs calculés pour les deux espèces à partir des données fines des chalutiers soviétiques (tabl. 23). Celles-ci ne montrent pas de tendance nette à la hausse. Avec des fluctuations importantes de courte période, l'effort effectif appliqué à T. trachurus a eu tendance à monter légèrement jusqu'en 1987, pour baisser ensuite, tandis que l'effort appliqué à T. trecæ pendant la même période est passé par deux maxima et trois minima dans un même intervalle de variation. Si la pue (effort effectif) de T. trachurus a eu plutôt tendance à baisser pendant la période d'observation, celle de T. trecæ a nettement augmenté dans la seconde moitié des années 80. D'apprès ces données, le stock n'aurait pas été surexploité. Dans les conditions présentes d'application, le modèle global ne permet pas de déterminer son potentiel.
Sardinelles
Les deux espèces de sardinelles ont été analyseées ensemble. Ce choix est justifié par la relative stabilité de l'importance relative des deux espèces dans les captures.
La relation entre la prise totale de sardinelles et l'effort nominal révèle un accroissement régulier des prises totales annuelles lorsque l'effort croît (fig. 13). Le modèle PRODFIT estime à 750 000 tonnes la prise maximale équilibrée de sardinelles. Elle serait atteinte pour un effort quatre fois supérieur à celui de 1990. Cette estimation souffre des mêmes faiblesses que l'estimation précédente. Toutefois, vu l'écart entre le taux d'exploitation actuel et celui de l'estimation du maximum de production équilibrée, on peut penser que le stock de sardinelles ne serait encore que modérément exploité, et que la prise équilibrée pourrait être significativement accure par une intensification de la pêche.
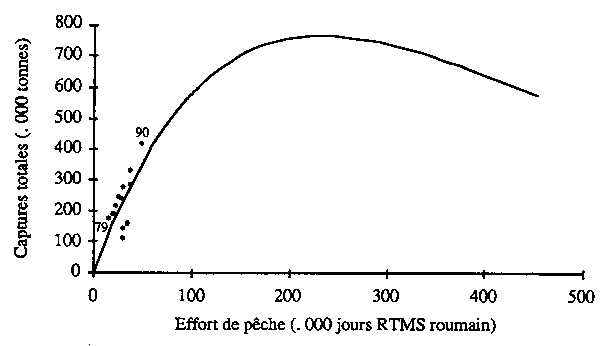
Figure 13 - Modèle global appliqué au stock de sardinelles (Sardinella spp.) de la sous-région (période 1979–90; les points représentent aux captures annuelles réelles).
(Tous pélagiques côtiers (sept espèces principales, sauf la sardine)
Le Groupe de travail a réalisé la même analyse sur l'ensemble des espèces principales de petits pélagiques, à, l'exception de la sardine (fig. 14). Celle-ci n'a pas été inclue, car elle n'est pas régulièrement pêchée dans la ZEE mauritanienne, et l'on ne disposait pas de statisques sur les captures réalsées dans les zones Sahara et Maroc.
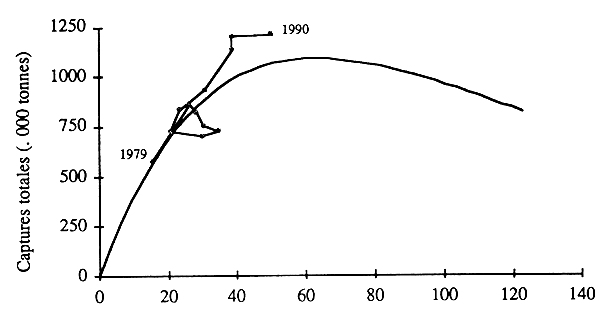
Figure 14 - Modèle global appliqué à l'ensemble des sept espèces principales de petits pélagiques (moins la sardine): période 1979–90; les points représentent les captures annuelles.
L'application du modèle PRODFIT montre un accroissement net des prises avec l'effort de pêche. Selon ce modèle, la prise maximale équilibrée du stock de petits pélagiques serait de 1, 1 million de tonnes. Jusqu'en 1990, l'effort est resté inférieur à celui correspondant à cette production. A cette date, il en était néanmoins proche. Apparemment, dans les années 80, les stocks pélagiques côtiers de la sous-région n'auraient pas été pleinement exploités.
3.3.3.2 - Effets des fluctuations de l'hydroclimat sur les potentiels
Les stocks de petits pélagiques sont influencés par les fluctuations de leur environnement, et notamment du régime d'upwelling. Cette influence se manifeste au niveau de l'abondance moyenne des populations, de la variabilité du recrutement, et parfois par des changements dans la localisation des populations. Au cors des trois dernières décennies, des changements se sont manifesté dans le régime d'upwelling ouest-africain, avec trois périodes plus intensives tous les dix ans et, peut-être, une certaine tendance à la baisse sur les trois décennies (fig. 15). Si les modèles précédents intègrent indirectement les effets des fluctuations passées de l'hydroclimat sur la productivité des stocks, ils ne le font pas explicitement. Les estimations précédentes peuvent perdre une partie de leur validité si le régime d'upwelling venait à s'écarter significtivement de celui qui a prévalu pendant la période d'observation des stocks.
Une telle éventualité n'est pas à exclure. Au cours des deux décennies passées, les captures de sardine ont eu tendance à croître dans la zone Sahara, et l'espèce est également pêchée en quantités appréciables, périodiquement dans le secteur mauritanien et épisodiquement dans le secteur sénégalais. On ignore toutefois si ces captures proviennent du stock marocain méridional (zone Sahara) dont l'aire de distribution s'étendrait temporairement vers le sud, ou d'une nouvelle population qui s'établirait de temps en temps en relation avec les fluctuations de l'upwelling.
Toutefois, à la différence des stocks de petits pélagiques de beacoup d'autres régions, ceux du secteur étudié ont globalement fait preuve jusqu'à présent d'une bonne stabilité.
Les raisons de ce comportement - notamment le taux plus modéré de leur exploitation - mériteraient des investigations particulières. Des enseignements intéressants ont été obtenus en comparant le comportement de populations de mêmes espèces dans des systèmes d'upwelling différents. Pour cette raison, de telles recherches gagneraient à être menées en coopération avec celles conduites dans d'autres régions. Même si ces recherches ne pouvaient être lancées, un programme minimum de suivi d'indicateurs du régime d'upwelling et de l'état des stocks (abondance) est justifié. Si la pêche des petits pélagiques s'intensifiait, la variabilité d'origine environnementale aurait des effets plus grands. Des recherches sur l'identité des principales populations (localisation des frayères et des nourriceries) permettraient d'améliorer l'application des modèlesd'évaluation. Elles constituent, en outre, un préalable à l'étude de la variabilité du recrutement.
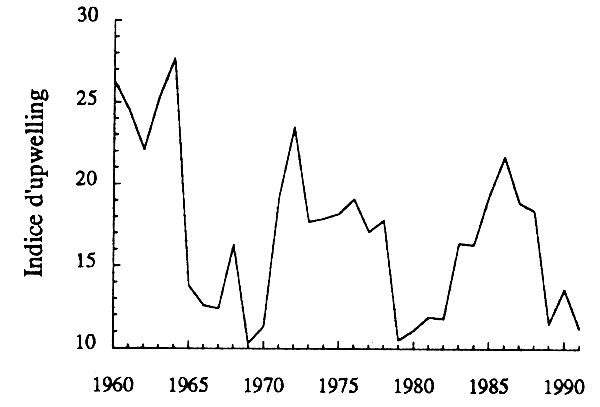
Figure 15 - Évolution de l'indice d'upwelling devant le littoral mauritanien au cours de la période 1960–1991 (données CNROP).
3.3.4.1 - Données et méthode
Données brutes
Les stocks des deux espèces de chinchard de la sous-région sont exploités principalement par la flottille de l'ex-URSS et l'ensemble roumain et, secondairement, par l'ensemble sénégalais. Des données de captures sont disponibles pour ces flottilles. On dispose également de fréquences de taille pour les captures de la flottille de l'ex-URSS. Ces données ont servi à calculer les distributions de taille des captures totales.
Méthode ADAPT
La méthode ADAPT (Gavaris, 1988) a été utilisée pour estimer, à partir des données précédentes, les mortalités par pêche exerceées sur les groupes d'âge des différentes cohortes, et les abondances annuelles (effectifs et biomasses) du stock. La méthode ADAPT est une analyse de populations véritables, basée sur les prises par âge. Elle estime abondances à la dernière année de pêche par la minimisation d'une fonction ‘critère’, en l'occurence, la différence de la moyenne des carrés entre Log (CPUE observées) et Log (CPUE calculées) par âges et par années. Les CPUE sont calculées par analyse de régression pour chaque groupe d'âge entre les valeurs observées et les abondances estimées par analyse séquentielle de populations. L'analyse de régression appliquée ici utilise l'hypothèse de l'erreur multiplicative.
La méthode ADAPT ajuste les biomasses à des d'abondance (pue) jugés représentatifs. Son application repose sur les données suivantes:
matrice des prises par âge;
poids moyen par âge;
effort standatd;
clé(s) taille/âge;
taux de mortalité mortalité naturelle.
Prise par âge
Les prise par âges (tabl. 25) ont été estimées en appliquant, aux prises par taille, des clés taille/âge établies par les chercheurs russes pour deux espèces. Pour les années 1986 à 1992, on a utilisé une clé par année éhantillonnée. Pour la période 1979 - 1985, une clé combinée des années 1988 et 1990 (à l'aide de l'algorithme de Kimura, 1987) a été utilisée.
Tableau 25 - Effectifs (en millions) de chinchards capturés par âges et par années dans la sousrégion.
| Age | Année | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1979 | 1980 | 1981 | 1982 | 1983 | 1984 | 1985 | 1986 | 1987 | 1988 | 1989 | 1990 | |
| a) T. trachurus | ||||||||||||
| 1 | 1,4 | 67,7 | 266,9 | 223,5 | 1384,6 | 35,6 | 528,2 | 289,2 | 113,3 | 54,2 | 0,4 | 0,0 |
| 2 | 10,2 | 28,5 | 516,2 | 583,2 | 185,8 | 88,5 | 584,3 | 166,1 | 361,1 | 124,5 | 16,5 | 21,2 |
| 3 | 64,5 | 360,2 | 64,9 | 119,6 | 6,7 | 84,9 | 179,4 | 94,5 | 164,6 | 307,3 | 136,6 | 105,6 |
| 4 | 57,4 | 139,4 | 197,9 | 27,3 | 11,1 | 18,2 | 62,4 | 94,9 | 90,6 | 128,7 | 214,1 | 225,1 |
| 5 | 450,9 | 583,9 | 62,0 | 37,6 | 6,7 | 18,9 | 9,5 | 50,7 | 57,9 | 45,9 | 71,9 | 82,4 |
| 6 | 0,0 | 3,6 | 32,7 | 26,3 | 3,2 | 179,2 | 36,4 | 51,6 | 47,6 | 34,8 | 26,1 | 27,8 |
| 7 | 0,1 | 35,2 | 2,6 | 39,3 | 0,0 | 78,0 | 12,4 | 27,0 | 20,8 | 24,8 | 20,4 | 17,2 |
| 8 | 1,0 | 10,2 | 0,0 | 20,6 | 0,1 | 1,1 | 0,4 | 18,5 | 4,6 | 10,3 | 2,5 | 0,1 |
| b) T. trecoe | ||||||||||||
| 1 | 31,9 | 23,2 | 0,0 | 110,4 | 15,4 | 0,1 | 41,1 | 106,8 | 156,5 | 54,1 | 0,4 | 0,1 |
| 2 | 123,4 | 264,1 | 141,9 | 58,3 | 121,6 | 387,6 | 238,8 | 87,1 | 308,3 | 128,2 | 4,7 | 3,5 |
| 3 | 5,3 | 102,7 | 211,9 | 65,8 | 4,5 | 225,2 | 89,0 | 82,5 | 96,3 | 192,6 | 88,2 | 110,5 |
| 4 | 380,4 | 687,6 | 37,6 | 206,7 | 61,4 | 32,8 | 149,3 | 92,5 | 64,2 | 75,4 | 190,5 | 158,6 |
| 5 | 34,6 | 182,7 | 79,9 | 42,7 | 125,8 | 6,4 | 68,1 | 191,7 | 49,8 | 41,0 | 125,4 | 108,2 |
| 6 | 5,4 | 10,4 | 40,1 | 49,1 | 89,0 | 1,4 | 20,4 | 83,1 | 48,5 | 26,3 | 60,2 | 39,3 |
| 7 | 0,8 | 0,1 | 6,0 | 0,4 | 17,4 | 0,3 | 1,8 | 8,3 | 5,7 | 4,1 | 29,8 | 3,9 |
| 8 | 0,0 | 2,9 | 0,0 | 0,0 | 0,5 | 1,0 | 0,0 | 10,8 | 1,3 | 0,1 | 5,1 | 0,7 |
Paramètres d'application
Pour appliquer la méthode, les paramètres suivants ont été retenus:
âges: 1 an à 8+;
années de pêche: 1979 à 1990;
âge de plein recrutement: 3 ans;
poids moyens par âge et par an: données de M'Bareck (1992);
mortalités naturelles: M = 0,5 pour T. trachurus; M = 0,6 pour T. treace (M'Bareck, 1992).
F terminal pour l'âge le plus vieux: moyenne pondérale (sur les abondances) des mortalités par pêche sur les groupes d'âge pleinement recrutés (3 à 7 ans);
abondance du premier groupe d'âge (recrutement) à l'année terminale (1990): moyenne arithmétique de ce groupe pour la période 1979–1989.
Effort standard
L'effort standard exercé sur T. trachurus et T. trecae est estimé en divisant les captures totales de chaque espèce par la pue standardisée de la flottille de RTMS russes ayant opéré dans la ZEE mauritanienne de 1979 à 1990. Les pue standardisées sont estimées à l'aide du modèle multiplicatif de Robson et Gavaris (Gavaris, 1980), programmé par Gavaris et Gascon (1985). Les captures journalières par bateau, d'une partie de la flottille de l'URSS (ZAPRIBA) opérant dans les zones COPACE 34.1.3 et 34.3.1 pendant la période 1979–1991, ont été utilisées. L'effort est aloué à l'espèce dont les captures sont égales ou supérieures à 50 % des captures journalières. La répartition des captures entre les deux espèces est basée sur l'échantillonnage réalisé par les observateurs du CNROP/AtlanNIRO pour la période 1986–1990, et à partir d'une combinaison d'informations fournies par les observateurs, les campagnes de chalutage, et les connaissances sur la bioécologie des espèces pour la période 1979–1985. Pour les périodes où l'information sur la composition spécifique faisait défaut, les proportions observées pendant la même période (mois) pour l'année la plus proche, ou leur moyenne pour la même période sur plusieurs années, ont été utilisées.
L'unité standard utilisée est:
pour T. trachurus:
* type de bateau: RTMS,
* période de pêche: janvier,
* secteur: zone Sahara (34.1.3);
pour T. trecae:
* type de bateau: RTMS,
* période de pêche: janvier,
* secteur: zone Mauritanie (34.3.1).
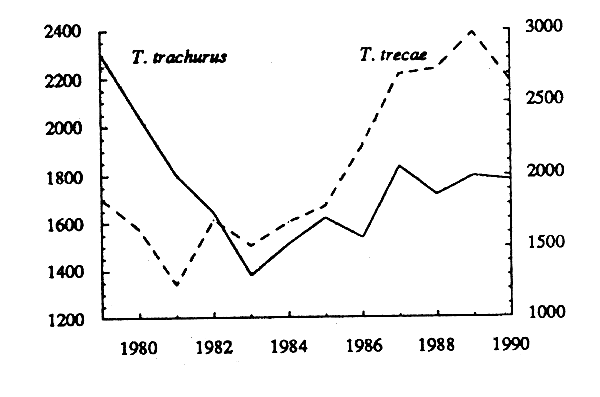
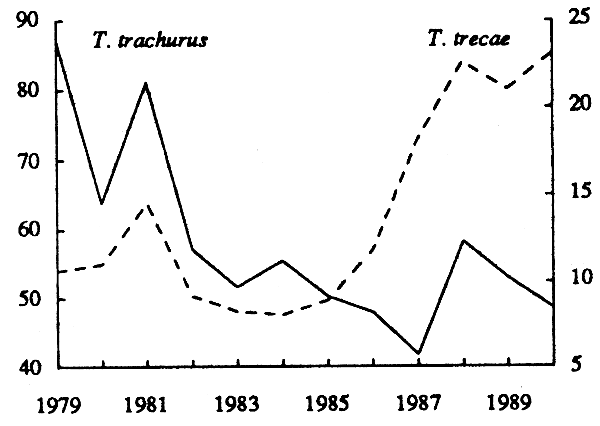
Les captures par effort (effectif), les carrés moyens des erreurs et les efforts effectifs ont été données dans le tableau 21. L'évolution des pue correspondantes est représentée sur la figure 16 b.
3.3.4.2- Résultats
Les résultats de l'analyse (effectifs, biomasses et coefficients de mortalité par pêche) sont donnés pour les deux espèces dans les tableaux 4 de l'annexe 4.
Chinchard européen (T.trachurus)
L' évolution de la biomasse de T. trachurus estimée par la méthode ADAPT est représentée, avec celle de T. trecae, sur la figure 16a. A partir d'un maximum en 1979, la biomasse a baissé graduellement jusqu'à un minimum en 1983, pour remonter ensuite jusqu'en 1987. A partit de cette date, elle est restée stable, à celui de 1979. A titre de comparaison, on a représenté sur la figure 16b lévolution de la prise par unité d'effort effectif (tabl. 23). L'évolution de la pue de T. trachurus diffère en partie de celle de la biomasse. La baisse initiale de cet indice d'abondance se poursuit plus longtemps (jusqu'en 1987 au lieu de 1983), et le redressement qui suit est à peine marqué.
Le recrutement (en effectif à l'âge l; ann. 4, tabl. 4.a.l) est très variable: les cohortes 1978 (âge l en 1979), 1984, 1985 et 1986 sont les plus abondantes. Ces forts recrutements expliquent la remontée de la biomasse estimée à partir de 1984. La cohorte née en 1988 est la plus faible. Ceci est peut-être dû à un artefact de calcul (cf. la question de la convergence et du degré de cumul de F; Pope, 1972). Cette anomalie possible de l'estimation du recrutement de 1989 a été corrigée en utilisant la moyenne arithmétique des recrutements des années 1979 à 1988.
La mortalité par pêche F (annexe 4, tabl. 4.a.3) est très faible pour les individus d'un an (en général<0,1), et faible pour ceux de 2 à 4 ans (≥0,1). Elle n'atteint des valeurs appréciables, mais pas excessives (≥ 0,2–0,3), que sur les classes de 4 ans et plus. La mortalité par pêche reste modérée sur l'ensemble des cohortes pleinement recrutées: la moyenne pondérale de F est inférieure à 0,2; une moyenne pondérale de 0,6 6 n'a été observée qu'une seule année (1980). Ce diagramme d'exploitation suggère que, pendant les années 80, le taux d'exploitation du stock est resté globalement modéré, et faible pour les classes d'âge inférieur à 4 ans.
a) biomasses (. 000 tonnes, âges≥l,F=0,15)
b) pue (effectif; tonnes/jour de mer)
Figure 16 - Biomasses estimées par la méthode ADAPT, et pue (effectif) des deux stocks de chinchard(T. trachurus T. treace) dans la sous-région.
Chinchard africain (T. treacae)
Après un léger déclin de 1979 à 1981, la biomasse de T. trecae estimée par la méthode ADAPT (fig. 16 a) a régulièrement augmenté jusqu'en 1989. La prise par unité d'effort effectif (fig. 16 b) suit une évolution similaire, à la différence près que le redressement du stock se manifeste un peu plus tardivement (en 1984 au lieu de 1981).
T. trecae a un recrutement plus variable que T. trachurus (annexe 4, tabl. 4.b.1). Les bons recrutements de 1982, 1984 et 1985 sont suivis par un redressement marqué et prolongé de la biomasse. Le recrutement de l'année 1989 montre la même anomalie que celle signalée pour T. trachurus. Cette estimation a été ajustée selon la même procédure.
La matrice de mortalité par pêche (annexe 4, tabl. 4.b.3) est semblable à celle décrite pour T. trachurus (avec des valeurs d'ensemble sensiblement inférieures et une variablité supérieure). La mortalité par pêche est très faible jusqu'à l'âge de trois ans compris, et modérée les deux années suivantes.
Dans les années 80, le stock de chinchard africain aurait donc été exploité, avec un taux global apparement encore plus modéré que pour le chinchard européen, et un diagramme d'exploitation encore plus satisfaisant (en termes de protection des jeunes). En 1990, la biomasse du stock aurait été nettement plus forte que dix ans auparavant.
Projections
Pour projeter l'évolution future des stocks de chinchards, on a admis que les cohortes étaient pleinement recrutées à partir de trois ans. Le recrutement partiel (RPa) des classes d'âge plus jeunes (1 et 2 ans) est exprimé par des coefficients de sélectivité calculés par la méthode ADAPT:
RPa=qa/ q3
dans laquelle qa est le coefficient de sélectivité pour la classe d'âge considérée (l ou 2 ans).
Les captures par recrue sont estimées par la méthode de Thompson et Bell (Ricker, 1975) et le logiciel Rivard (1982). Les paramètres suivants ont été utilisés:
coefficient de mortalité naturelle: M = 0,5 pour T. trachurus, et 0, 6 pour T. trecae;
poids moyens des groupes d'âge (en début d'année) pour la période de 1979–90;
valeurs de Fo, 1 égales à 0,62 pour T. trachurus, et à 0,72 pour T. trecae.
Les principaux résultats sont donnés dans le tableau 26.
a) Trachurus trachurus
| Mortalité par pêche | Prise en | Rendement par | poids moyen des | Prise par unité | |
|---|---|---|---|---|---|
| nombre | recrue | individus | d'effort | ||
| 0,1 | 0,112 | 0,024 | 0,219 | 2,199 | |
| 0,2 | 0,194 | 0,040 | 0,204 | 1,783 | |
| 0,3 | 0,256 | 0,049 | 0,193 | 1,483 | |
| 0,4 | 0,305 | 0,056 | 0,183 | 1,260 | |
| 0,5 | 0,345 | 0,061 | 0,176 | 1,090 | |
| F0, 1 | 0,565 | 0,367 | 0,063 | 0,171 | 1,000 |
| 0,6 | 0,378 | 0,064 | 0,169 | 0,957 | |
| 0,7 | 0,406 | 0,066 | 0,163 | 0,851 | |
| 0,8 | 0,429 | 0,068 | 0,159 | 0,766 | |
| 0,9 | 0,450 | 0,069 | 0,154 | 0,695 | |
| 1,0 | 0,468 | 0,071 | 0,151 | 0,636 | |
| 1,1 | 0,485 | 0,072 | 0,148 | 0,585 | |
| 1,2 | 0,489 | 0,072 | 0,145 | 0,542 | |
| 1,3 | 0,512 | 0,073 | 0,142 | 0,505 | |
| 1,4 | 0,524 | 0,073 | 0,140 | 0,472 | |
| 1,5 | 0,536 | 0,074 | 0,138 | 0,443 | |
| Fmax* | 3,229 | 0,649 | 0,076 | 0,117 | 0,212 |
* attention: critère de convergence non satisfait.
b) Trachurus trecae
| Mortalité par pêche | Prise en nombre | Rendement par | Poids moyen des | Prise par unité | |
|---|---|---|---|---|---|
| recrue | individus | d'effort | |||
| 0,1 | 0,074 | 0,019 | 0,252 | 2,348 | |
| 0,2 | 0,130 | 0,031 | 0,239 | 1,960 | |
| 0,3 | 0,174 | 0,040 | 0,228 | 1,669 | |
| 0,4 | 0,210 | 0,046 | 0,218 | 1,446 | |
| 0,5 | 0,239 | 0,050 | 0,210 | 1,272 | |
| 0,6 | 0,264 | 0,054 | 0,204 | 1,132 | |
| 0,7 | 0,285 | 0,057 | 0,198 | 1,019 | |
| F0, 1 | 0,719 | 0,289 | 0,057 | 0,197 | 1,000 |
| 0,8 | 0,304 | 0,059 | 0,193 | 0,925 | |
| 0,9 | 0,320 | 0,060 | 0,189 | 0,847 | |
| 1,0 | 0,334 | 0,062 | 0,185 | 0,780 | |
| 1,1 | 0,347 | 0,063 | 0,182 | 0,723 | |
| 1,2 | 0,358 | 0,064 | 0,179 | 0,674 | |
| 1,3 | 0,368 | 0,065 | 0,176 | 0,631 | |
| 1,4 | 0,378 | 0,066 | 0,174 | 0,593 | |
| Fmax | 1,500 | 0,386 | 0,066 | 0,172 | 0,559 |
Si le choix de F0,1comme objectif d' exploitation permettrait d'accroître de trois à quatre fois les captures par rapport à celles correspondant au régime de 1990, cette augmentation s' accompagnerait d'une forte chute de la biomasse. Il est probable que la folttille ne puisse supporter une telle baisse de rendement. Des projections ont donc été réalisées pour deux tauxd d'exploitation inférieurs, plus comparables avec les performances récentes de la flottille. Pour cela, les paramètres suivants ont été utilisés:
poids moyen des groupes d'âge sur la période 1979–90;
niveau de recrutement: moyenne arthmétuque des recrutements annuels 1979–88;
même coefficients de mortalité naturelle et de recrutement partiel que ceux déjà utilisés.
Les résultats de ces projections, en termes de conséquences pour la pêche par rapport au régime de 1990, sont résumés dans le tableau 27.
| Mortalité par pêche | F = 0,15 | F = 0,25 | ||
|---|---|---|---|---|
| Espèces | T. trachurus | T. trecae | T. trachurus | T. trecae |
| Modification du rendement | négligeable | négligeable | -10 %env. | -10 %env. |
| Variabilité du rendement | comparable | comparable | comparable | comparable |
| Capture annuelle(.000 t) | 185–192 | 220 | 290–320 | 320 |
Pour les deux espèces, une mortalité par pêche de F = 0, 15 ne modifierait significativement ni le rendement, ni sa variabilité. Les captures moyennes annuelles des deux espèces augmenteraient alors d'un tiers environ par rapport au niveau qu'elles avaient à la fin des années 1990 (300 000 tonnes environ), pour se rapprocher du maximum observé pendant la période d'observation. Une mortalité de F = 0, 25 permettrait de doubler, en moyenne, la production annuelle des deux espèces (par rapport au niveau 1990), sans changement appréciable de la variabilité du rendement, mais celui-ci baisserait alors d'environ 10 %.
Compte tenu des incertitudes d'origine diverse (carences statistiques, méconnaissance de l'état actuel de la pêche des pays de l'Europe de l'est, variations futures du recrutement), les présentes évaluations doivent être considérées comme préliminaires. Cependant, globalement, le stock de petits pélagiques apparaît comme modérément exploité. Des estimations plus précises de leur potentiel deviendront plus urgentes leur exploitation deviendra plus intensive.
Les principales conclusions que l'on peut avancer de leur état actuel d'exploitation sont les suivantes:
le potentiel maximal de capture de l'ensemble des espèces principales de la sous-région (moins la sardine) serait de l'ordre de 1, 1 million de tonnes; ce n'est qu'en 1990 que le taux d'exploitation se serait approché du niveau correspondant; si les informations sur les difficultés de la pêche des pays de l'Europe de l'est sont fondées, le taux actuel d'exploitation aurait baissé depuis; cet ensemble de ressources n'aurait donc pas été surpêché en termes biologiques, et il est possible qu'il ne soit actuellement que modérément exploité;
à cause des insuffisances statistiques (converture et ventilation par flottilles et populations), les évaluations restent imprécises au niveau tant des pêcheries (stocks unitaires, flottilles) que de la ZEE mauritanienne; globalement, par rapport à celle de 1990, la production des stocks de sardinelles pourrait sans doute croître très significativement (de 4 fois); celle de chinchards également, mais dans une proportion moindre; si le modèle global et le modèle analytique conduisent, pour les chinchards, à des conclusions différentes, on doit accorder plus de confiance aux résultats du modèle analytique car il permet une analyse plus approfondie et il porte sur des données plus variées; dans leurs grandes lignes, ces premières conclusions sont vraisemblablement valables pour les autres espèces;
actuellement, la productivité des stocks de petits pélagiques n'est apparemment pas la contrainte majeure de la pêcherie; les contraintes au développement de la pêche artisanale et les incretitudes sur l'avenir de la pêche des flottilles de l'Europe de l'est sont avant tout d'order économique;
l'analyse séquetielle des populations de chinchards illustre les rensignememts que l'on puet tirer, à condition de disposer des statistiques nécessarires, de simulations des contraintes propres aux différentes pêcheires.
![]()
![]()
![]()