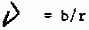![]()
![]()
![]()
4.1 Croissance
4.2 Croissance différentielle selon le sexe
4.3 Relation taille-poids
4.4 Rapport des sexes
4.5 Mortalités
4.6 Rendement par recrue
4.7. Modèles de production
4.8 Autocorrélation dans la série de cpue
Le tableau 1 et les figures 9, 10, 11 et 12 montrent les modes observés dans chaque campagne étudiée.
Dans la partie droite du tableau, on remarque qu’un facteur subjectif a influé de façon décisive. Nous avons néanmoins essayé de “deviner” la position des modes dans le tableau par homogénéisation des valeurs inscrites dans chaque colonne, réduisant ainsi les différences entre les rangs; nous devons de toute façon insister sur le fait que les deux derniers modes de chaque rang devraient être accompagnés de points d’interrogation, car leur valeur est pour le moment considérée comme guère plus qu’hypothétique.
En supposant l’existence de deux maximums de ponte bien définis chaque année, les campagnes réalisées en juillet-août devraient donner une distribution de tailles très semblable à celle réalisée en janvier-février 1976, et c’est effectivement le cas. Il convient de noter que, dans la distribution de tailles de Cort et Pérez-Gándaras, on ne voit pas apparaître le premier mode des autres distributions, qui est autour de 7-8 cm; toutefois, la maille utilisée à cette époque et dans cette zone par la flottille de pêche espagnole était de 35 mm (Cort, communication personnelle).
La concordance entre les premiers modes est très forte. Le mode qui apparaît en second pourrait correspondre aux individus ayant autour d’un an, dont les oeufs ont éclos en août/septembre. Il se situe en tout cas entre 11 et 12 cm (nous nous référons toujours a la longueur du manteau). Le troisième, qui correspondrait à un an et demi, se situerait entre 14 et 15 cm, sauf pour la campagne 1978 où, après ventilation de la distribution de tailles, aucun mode n’apparaît en ce point. Cette campagne a d’ailleurs été un peu plus difficile à analyser. Les modes suivants apparaissent à 18-19 cm, celui-ci toujours très net, un autre a 20-21 cm et un dernier à vrai dire douteux en raison du faible nombre d’exemplaires capturés à cette taille, à 24-25 cm.
Etant donné qu’Itami et ses collaborateurs ont trouvé que le poulpe, à 5 ou 6 mois de l’éclosion, avait en moyenne 5-6 cm de longueur de manteau, nous pouvons déduire que le second mode des distributions de tailles analysées, sauf celle de Cort et Pérez-Gándaras qui serait la première, correspond effectivement à des individus approchant de l’âge d’un an. Le troisième mode correspondrait pratiquement à un an et demi, et le quatrième à près de deux ans. Rappelons que l’équation de croissance de Guerra comme celle de Hatanaka prédisent pour des poulpes d’un an une taille moyenne se situant autour de 12 cm; la grande différence entre les deux équations apparaît à partir de la seconde année; d’après l’équation de Hatanaka, des exemplaires de deux ans mesureraient plus de 17 cm et des exemplaires de trois ans 20,3 cm. D’après l’équation de Guerra, en revanche, des exemplaires de deux ans mesureraient 26,8 cm, et ceux de 3 ans 36,8 cm.
Le choix d’une valeur K, tenté a maintes reprises, est une entreprise qu’il faudrait affronter avec plus de soin qu’on ne l’a fait jusqu’à présent, sans quoi, on ne fera que semer la confusion et perdre du temps. Voyons cette opération à travers un exemple: nos données semblent confirmer qu’à l’âge d’un an, la taille modale serait de quelque 12-13 cm et, à l’âge de 2 ans, dépasserait les 18 ou 19 cm. Si nous considérons un troisième mode annuel a 24 cm, nous obtenons simultanément K = 0,34 et L¥ = 36,2l cm, pour des valeurs de l1 = 12 cm, l2 = 19 cm et l3 = 24 cm.
Il suffirait de changer l1= 12 cm par l1 = 13 cm pour avoir K = 0,18 et L¥ = 109,53 cm; si nous considérons que les paramètres de croissance ont une signification biologique, ce deuxième résultat doit être repoussé comme absurde, vu la valeur obtenue pour L¥. Toute tentative, pour estimer la valeur de K à partir des données dont nous disposons, devrait donc passer par la réduction, a priori ou a posteriori, de la valeur de L¥. La valeur fixée pour L¥ déterminera à son tour les valeurs possibles de K.
La sensibilité de K et de L¥ à des variations aussi légères des tailles modales - sensibilité qui dépend évidemment du petit nombre de modes utilises - devrait nous inciter à la prudence au moment d’utiliser une équation de von Bertalanffy aux fins d’évaluation. Un fait dont il faut tenir compte est que la valeur choisie par Guerra pour L¥ est sans doute trop forte. Probablement, après ce que nous venons d’exposer, il vaudrait mieux attribuer dès le début à L¥ une valeur comprise entre 30 et 40 cm de longueur de manteau, en écartant toutes les estimations qui ne donneraient pas comme résultat des valeurs de L¥ entrant dans cet intervalle. Nous pourrions par conséquent admettre comme possibles des valeurs K= 0,34 et L¥ = 36,41 que nous avons déjà signalées; K =0,29 et L¥ = 40 cm, ou K = 0,36, L¥ = 35 cm, etc.
Une fois cette décision prise, le choix de l’équation de croissance proprement dite n’a plus beaucoup d’importance. Si nous considérons des tailles moyennes de 13, 19, 23 et 26 cm correspondant à des classes d’âges successives, il en sort une équation de croissance dans laquelle K = 0,38 et L¥ =32,15 cm. Etant donné que dans cet essai on exploite mieux les données obtenues par analyse des distributions de tailles et que le résultat se maintient dans les limites considérées comme possibles, nous retiendrons cette équation pour les essais de rendement. L’équation de von Bertalanffy prend la forme ci-après:
lt = 32,15 (1 - e - 0,38 (t + 0,36))sans que cela veuille dire que nous écartons toute autre solution qui maintienne L¥ dans une valeur comprise entre 30 et 40 cm, comme nous l’avons expliqué.
Un second grand problème est représenté par la croissance différentielle selon le sexe. Le travail de Cort et Pérez-Gándaras nous a permis de distinguer la composition normale des distributions de tailles par sexe. On peut noter une grande concordance entre miles et femelles dans les deux premiers modes, qui correspondaient aux deuxième et troisième modes chez les autres auteurs. Le troisième mode correspond à 19 cm pour les mâles et 17 cm pour les femelles; le quatrième à 22 cm et 20 cm respectivement, et le cinquième à 24-25 et 22 cm. Les distributions de tailles auxquelles on se réfère sont représentées sur les figures 13 et 14 pour les mâles et les femelles respectivement.
Il faut se souvenir que, dans la gamme de tailles ou l’on note une croissance différentielle suivant le sexe, les femelles sont soumises à une forte mortalité, ce qui pourrait fausser considérablement nos estimations de la croissance si la mortalité s’avérait être spécifique de la taille. Si tel n’était pas le cas, nous inclinerions à penser que l’on trouverait une valeur satisfaisante de K pour les femelles autour de K = 0,5 avec L¥ autour de 25 cm. Souvenons-nous que la plus grande femelle mesurée au cours de la campagne susmentionnée avait un manteau de 24 cm, tandis que le mâle le plus grand atteint 30 cm.
Une équation de von Bertalanffy possible pour les femelles serait donc la suivante:
lt = 24,5 (1 - e - 0,51 (t + 0,32))Cette équation apparaît, en même temps que l’équation générale, sur la figure 15.
Aux fins d’évaluation et pour le moment, nous retiendrons l’équation générale déjà décrite, mais pour des travaux ultérieurs il serait plus exact d’utiliser des équations différentielles par sexe.
L’équation “générale” est en réalité conforme à la croissance des mâles, étant donné que les modes qui figuraient dans les distributions de tailles générales coïncident avec les modes caractéristiques des mâles; cette particularité tient à la mortalité des femelles dans la gamme de tailles dans laquelle la croissance est différentielle, de sorte qu’au moment de procéder à l’ajustement, trois des quatre points utilisés correspondent à des tailles qui ne se rencontrent pratiquement que chez les mâles.
Les données de taille et de poids des deux campagnes réalisées par l’équipe de pêche de l’Institut espagnol d’océanographie (laboratoire des Canaries) ont servi a approfondir l’étude de la relation taille-poids chez le poulpe de ces stocks.
Le tableau 2 montre le poids moyen par taille pour des exemplaires éviscérés de poulpes ayant de 11 à 19 cm, recueillis en juillet 1978 et en mars 1979 (campagnes 7807 et 7903 respectivement). On peut noter la grande différence de poids qui existe entre des exemplaires de même taille.
Au début, nous étions tentés de penser que c’est le facteur de condition qui est responsable de ces différences, mais la chose est beaucoup moins évidente si nous observons les résultats de campagnes antérieures. Ainsi, par exemple, Cort et Pérez-Gándaras publient une relation taille-poids qui annonce des poids très inférieurs à ceux qui ont été donnés bien que leur campagne ait été réalisée, a la même époque (juillet-août) que la campagne 7807.
Pour en revenir au tableau 2, nous pouvons constater que le poids moyen varie d’une campagne à l’autre d’un coefficient de 1,2-1,3 et si maintenant nous multiplions les poids ainsi obtenus par un coefficient de 1,4-1,5 nous “retrouvons” les poids de la campagne 7903.
Le fait que nous puissions passer de l’une à l’autre moyennant un coefficient pratiquement constant dans la gamme de tailles que nous avons dans la phase exploitée montre que, pour l’évaluation des rendements, le fait d’employer une relation ou une autre perd son importance, pour ce qui est des variations interannuelles. Il n’en est pas de même, malheureusement, de la condition d’une pêcherie saisonnière comme celle-ci.
En tout cas, il est nécessaire d’étudier d’urgence aussi bien la condition saisonnière du poulpe que les variations géographiques, et d’une année sur l’autre, de la relation taille-poids.
Quant à nous, nous retiendrons pour le moment la relation obtenue au cours de la campagne 7903 pour le poulpe éviscéré.
W = 1,27 L 2,62
Le tableau 3 analyse le rapport des sexes d’après les données présentées par Cort et Pérez-Gándaras (1973).
La figure 16 montre comment la proportion de femelles dans les captures va en diminuant; la courbe qui montre cette diminution a été tracée à la main.
Le segment A suppose le remplacement de la courbe par une disparition massive des femelles en “arête vive” qui se produirait au niveau d’une taille de 22,25 cm approximativement, correspondant à un âge de 2,74 ans d’après l’équation générale de von Bertalanffy retenue. Pour cela, nous supposerons plus loin, aux fins d’évaluation, qu’il se produit une mortalité massive des femelles a 2,75 ans, la biomasse du stock se trouvant à ce moment la réduite à la moitié.
La division des histogrammes des tailles en composantes normales a permis d’effectuer une première étude de la mortalité totale au moyen de courbes de capture (Ricker, 1958, 1975).
Les estimations de Z ainsi obtenues doivent être uniquement considérées comme des approximations provisoires et grossières de la valeur réelle; il n’est pas tout a fait licite d’employer des courbes de capture dans ce cas, étant donné que l’on suppose que chaque point apparaîtra comme le reflet de l’abondance d’une cohorte; les cohortes sont séparées par des semestres et l’effort (reflet de la mortalité) n’est pas stable d’un semestre à l’autre, du moins pour la flottille espagnole.
On ne dispose pas non plus de données sur l’évolution de recrutements successifs qui, s’ils manifestent une tendance, pourraient avoir une incidence sur les estimations obtenues; enfin, il est à peu près évident qu’il existe des différences de mortalité aussi bien d’une cohorte à une autre que d’une classe d’âge à la suivante.
Bien que conscients de tous ces facteurs, nous avons considéré qu’il valait la peine d’exploiter au maximum les renseignements dont nous disposons; nous avons donc analysé les courbes de capture, dont les distributions sont corrigées pour tenir compte de la sélectivité, car celle-ci couvre une grande partie de la gamme de tailles de la phase exploitée.
La figure 17 montre les résultats. Les estimations de la mortalité correspondent à des points séparés par des semestres, donc à des taux de mortalité dans cette échelle de temps.
On est frappe par la ressemblance de la pente des courbes correspondant aux années 1972, 1976 et 1978 - 1,27, 1,09 et 1,10 respectivement; si l’on fait la moyenne des différences des népériens consécutifs correspondant aux abondances des quatre premières classes d’âge, on obtient les chiffres de 1,49, 1,09 et 1,42 pour les trois dates citées.
L’image que nous obtenons ne contredit pas celle qui découle de la série chronologique d’efforts publies par le Groupe de travail du COPACE, d’après laquelle la pêcherie est passée par deux étapes clairement différenciées: une période de faible mortalité, dans laquelle serait incluse l’année 1967, date de la campagne décrite par Cabrera (1967) et une autre de forte mortalité qui commencerait en 1971 et dans laquelle on trouverait les trois autres campagnes auxquelles nous nous référons.
Pour l’année 1967, la mortalité totale trouvée est de 0,75 sur une base semestrielle. La valeur de Z obtenue pour l’année 1978 doit, en fait, être surestimée car, à partir de 1977, la flottille espagnole a changé de maille de sorte qu’on comptabilise dans l’abondance les survivants non capturés, ce qui fait apparaître une période de forte transition.
D’après le Groupe de travail du COPACE (COPACE, 1968), l’effort exercé en 1976 a été 2,22 fois supérieur à celui de 1967; et celui de 1972, 2,16 fois supérieur.
En utilisant les pentes des courbes de capture respectives comme des estimations absolues de la mortalité totale chaque année, nous obtiendrions des valeurs M de 0,96 et 0,6 respectivement, sur une base annuelle, au moyen des relations:
(Zi - M)/(Zj - M) = Fi/Fj = fi/fjAux fins de notre évaluation, nous retiendrons par conséquent une limite inférieure M de 0,5 sur une base annuelle et une limite supérieure de 1,0.
Si l’on examine les distributions de tailles par sexe qui figurent dans le travail de Cort et Pérez-Gándaras, nous pouvons constater que la mortalité des mâles et des femelles est différente, différence qui va en augmentant avec l’âge et devient particulièrement importante à partir de la troisième classe d’âge (exemplaires ayant vécu un an et demi), comme il ressort de la figure 18 qui montre les courbes de capture des mâles et des femelles.
Toutes les figures des courbes de capture accusent une forte pente qui correspond aux premières classes d’âge et s’atténue à partir de la classe que nous avons appelée classe IV et qui correspondrait à des exemplaires âgés de 2 ans. Ce fait paraît démontrer que la pèche s’oriente de préférence sur les exemplaires jeunes ou, ce qui est peut-être plus exact, que de la manière dont l’activité de pêche se déroule, les exemplaires jeunes sont soumis à une mortalité plus forte. Effectivement, la mortalité totale diminue chez les individus plus âgés, bien que la mortalité naturelle des femelles augmente. Les valeurs obtenues pour la mortalité totale chez les individus âgés nous amènent à penser que la valeur réelle de M devrait se trouver plus près de la limite inférieure que nous avons choisie, M = 0,5 sur une base annuelle, que de la limite supérieure M = 1,0, ce dont nous devrions tenir compte dans les chapitres sur les évaluations.
Le calcul du rendement par recrue proposé par Ricker (1958, 1975) est un bon outil pour établir le cadre des rendements pouvant être obtenus compte tenu d’un recrutement constant de ce ou de ces stocks.
Nous avons utilisé dans tous les cas le modèle linéaire de Ricker, certains que le modèle exponentiel n’aurait pas donne de grandes différences.
La stratégie suivie a été celle-ci: on a supposé l’entrée dans la pêcherie de 1 000 kg de recrues ayant une taille de 7 cm, supposée être la taille de première capture avec une maille de 40 mm. Le premier intervalle prend fin quand le poulpe atteint 10,16 cm de taille de manteau, taille de la première capture correspondant à la maille qu’autorisé la réglementation du COPACE.
Les intervalles suivants prennent fin quand le poulpe atteint 1, 1,5, 2 et 2,75 ans. A ce dernier stade se produit la mortalité massive des femelles et la population se réduit de moitié. Les intervalles suivants prennent fin à 3,5, 4,5 et 5,5 ans, moment ou l’on suppose que les poulpes abandonnent la pêcherie.
On a utilisé l’équation de von Bertalanffy déjà citée:
lt = 32,15 (1 - e - 0,38 (t + 0,34))et une relation taille-poids tirée des données de la dernière campagne réalisée par le personnel de l’Institut espagnol d’océanographie (laboratoire des Canaries) et qui correspond a mars 1979
W = 1,27 L 2,62On a utilisé des valeurs de mortalité naturelle M sur une base annuelle de 0,5 et de 1 (= un), (M/K = 1,32 et 2,64 respectivement) et des valeurs de mortalité due à la pêche, F étant constante, de 0,5, 1, 1,5 et 2 pour une maille de 40 mm; de 0,5, 0,75, 1,0, 1,25, 1,5 et 2 pour une maille de 60 mm et M = 0,5; enfin, de 0,5, 0,75, 1, 1,25, 1,5, 1,75 et 2 et 2,5 pour une maille de 60 mm et M = 1.
Dans certains essais, on a supposé que la mortalité due à la pêche commençait à se faire sentir à partir d’une taille de 7 cm, et dans d’autres essais, a partir de 10,16, tailles de première capture correspondant à des mailles de 40 et 60 mm comme nous avons déjà dit. Enfin, on a calculé le rendement par recrue pour les vecteurs du tableau 4, avec F variable, vecteur dont nous supposons qu’il reflétait approximativement la situation réelle en 1976.
La figure 19 montre l’évolution de la biomasse de la cohorte non exploitée pour les deux valeurs de M.
Les résultats présentés dans les tableaux 5 et 6 et les figures 20 et 21 font apparaître des conclusions très intéressantes; pour les deux valeurs de M, il se produit des augmentations importantes des rendements quand on change de maille à des taux d’exploitation élevés. Le changement de maille, dans le système actuel, supposerait en particulier une augmentation de 34 pour cent des rendements par recrue pour M = 0,5 et de 23 pour cent pour M = 1.
Quand on a M = 0,5, les rendements par recrue commenceraient a diminuer à partir de F = 1 pour l’une et l’autre mailles. Avec M = 1 et une maille de 40 mm, il se produirait une baisse de rendement à partir de F = 1. Avec la maille de 60 mm, le rendement se stabiliserait à partir de F = 1,5. Tout donne à penser par conséquent qu’une augmentation quelconque de l’effort de pèche au cours de la période 1972-1976 provoquerait une baisse des rendements ou, dans le meilleur des cas, aucun gain appréciable. On peut plutôt penser qu’on se trouve devant une situation de surpêche de croissance, dans laquelle il est possible d’obtenir des captures accrues tout en diminuant l’effort. Nous développerons cette question dans le chapitre des conclusions.
En ce qui concerne les fécondités relatives, elles ont été calculées comme rapport entre la biomasse moyenne des femelles de 1 an à 2 ans et demi, dans chaque système d’exploitation, et la biomasse moyenne des femelles fécondées dans le stock vierge, comme le montrent les tableaux 7 et 8 et les figures 22 et 23. Cette hypothèse équivaut à supposer que les femelles ont une fécondité qui dépend de leur poids.
Il est possible de vérifier la forte baisse de fécondité par recrue à de forts taux d’exploitation. Comme on ne sait rien de la relation stock/recrutement, on ne peut pas prévoir l’importance de ce phénomène pour la régulation du stock. Mais nous pouvons dire que le changement de maille doit avoir provoqué une augmentation importante de la fécondité, encore que celle-ci conserve des valeurs très basses.
Le Groupe de travail du COPACE susmentionné (1978) a procédé à l’adaptation de séries statistiques capture/effort à des modèles de production de Schaefer (Schaefer, 1954, 1957; Beverton et Schaefer, 1963) et Fox (1970) avec des modèles de régression fonctionnelle. Ce groupe a utilisé une unité d’effort arbitraire obtenue en combinant des données espagnoles et japonaises.
Nous avons pour notre part jugé sur la même série de données, en introduisant quelques variantes dans l’élaboration des modèles afin de connaître avec une précision plus rapide leur comportement. Dans tous ceux-ci, nous avons résolu le modèle de régression prospectif facilement transformable en régression fonctionnelle si l’on se souvient que:
Les différents modèles que nous avons ajustés ont été les suivants:
1) Avec la cpue et l’effort utilisé par le Groupe de travail, on a résolu graphiquement le modèle général de production afin de se rendre compte de la forme de la courbe et d’avoir une indication de la valeur possible du paramètre m. La figure 24 montre ce modèle résolu graphiquement. Sa forme est très semblable à celle du modèle de Fox (1971), m tend vers 1, est n’est que légèrement plus pessimiste, de sorte qu’on pourrait prévoir une valeur m légèrement supérieure à 1. Le modèle de Fox, que l’on peut soir superposé au modèle général sur la figure 25, prédit un RME (rendement maximum équilibré) de 110 000 t pour un effort de 1 258 unités du Groupe de travail (r = -0,94; = 0,0008456).Pour clarifier un peu la situation et établir un critère qui nous permette de choisir les essais susceptibles de s’ajuster avec plus de probabilité à la situation réelle, nous avons calculé les coefficients de corrélation entre les différentes séries de cpue. Les résultats ont été les suivants:2) Nous avons essayé un modèle de production avec la série de cpue espagnole. La raison de ce choix était liée I la prépondérance des captures espagnoles dans la zone nord, les captures japonaises se répartissant par contre entre deux zones où habitent probablement des stocks différents du point de vue de la dynamique (zones du cap Garnett et du cap Blanc), avec une évolution importante dans la proportion des captures d’une zone à l’autre ces dernières années.
On a obtenu un RME de 136 000 t, correspondant à un effort de 1 165 103 unités espagnoles pour le modèle de Schaefer, et un RME de 124 000 t pour un effort de 1 216 x 103 unités d’après le modèle de Fox, qui a donné un ajustement de r = - 0,98 (ajustement réalisé sur neuf paires de valeurs).
3) Etant donné le changement intervenu dans les stratégies de pêche a partir de 1976, nous avons décidé de réaliser un nouveau modèle fondé sur les données de capture et d’effort du Groupe de travail du COPACE (1978), mais sans tenir compte des données de 1976 et 1977.
Le modèle de Fox a donné comme résultat un RME de 111 000 t, correspondant a un effort de 1 259 unités du Groupe de travail (r =0,93). Le modèle de Schaefer a donné un RME de 114 000 t pour un effort de 1 127 unités.
4) On a soumis à normalisation la cpue espagnole d’une part et la cpue japonaise et coréenne, supposées homogènes, d’autre part, en divisant les séries respectives de cpue par leur valeur moyenne. On a obtenu l’effort total après avoir détermine l’effort conjoint de ces trois pays et l’avoir extrapolé à l’effort correspondant à la capture totale.
Dans ce cas, le RME prédit par le modèle de Fox a été de 105 000 t pour un effort normalisé de 135 unités, tandis que le modèle de Schaefer annonçait un RME de 113 000 t pour un effort de 131 unités.
5) On a répété l’opération en éliminant les années 1976 et 1977, et l’on a obtenu un RME de quelque 102 000 t pour le modèle de Fox et de quelque 110 000 t pour celui de Schaefer.
Les valeurs des différents essais sont données, pour plus de clarté dans le tableau 9.
a) Entre les séries de cpue japonaise et espagnole pour la période 1969-1977, la corrélation a été de 0,0325; tandis que pour la période 1969-1975, elle s’élève à 0,92, ce qui confirme le “bruit” introduit par ces deux dernières années dans les analyses et que l’on peut attribuer à deux principales causes possibles: le changement de stratégie des chalutiers orientaux dont nous avons déjà parlé, et une rupture de la série de cpue espagnole, qui prend pour base à un moment donné l’effort des chalutiers congélateurs.Nous en concluons que le cas (5) devrait être considéré comme le plus apte à refléter la situation actuelle, de sorte que le RME se trouverait entre 100 000 et 110 000 t. Le cas (3) pourrait aussi être considéré comme acceptable, avec des valeurs de RME comprises entre 110 000 et 115 000 t. Les figures 25 et 26 montrent la représentation graphique de ces modèles pour des régressions prospectives.b) La corrélation entre la série de cpue japonaise et celle du Groupe de travail a été de 0,75.
c) La corrélation entre la cpue espagnole et celle du Groupe de travail a été de 0,96.
Au cours d’analyses ultérieures, faudrait faire très attention à l’absence de corrélation qui se manifeste à partir de 1976 entre les deux séries de cpue, japonaise et espagnole, et tenir compte également de la possibilité de manipuler des données d’effort coréennes.
Les derniers chiffres que nous avons cités montrent, de toute manière, la bonne concordance des séries capture/effort avec les modèles globaux, en particulier avec celui de Fox. Divers points se dégagent de ces figures, dont il faut tenir compte:
1) En 1975, le stock a été soumis à une forte surexploitation; nous ne savons pas avec certitude si cet effort s’est ralenti au cours des années 1976 et 1977, car c’est à ce moment là que change la mesure de l’effort du Groupe de travail du COPACE et de l’effort normalisé utilisé par nous-mêmes dans la présente communication.2) De toute manière, il aurait fallu réduire de 30 pour cent l’effort de pêche réalisé en 1975 pour nous trouver autour de l’effort qui aurait provoqué des captures voisines du RME.
3) Il ne semble pas qu’il y ait lieu de penser 1 une transition qui aurait duré plus d’une ou deux années. Nous pouvons constater, sur les modèles de production réalisés par le Groupe de travail ou sur ceux qui figurent dans cette même communication, que les déviations de l’équilibre ne sont pas considérables, malgré les énormes variations de l’effort d’une année sur l’autre. C’est pourquoi nous croyons qu’il faudrait raisonner en fonction de la cpue des années antérieures, comme le fait le rapport du Groupe de travail du COPACE (Tableau 10), au moment de penser aux contingents de capture.
4) Le Comité du COPACE a conseillé d’utiliser une maille de 60 mm a la place de la maille de 35 mm traditionnellement employée dans cette zone (recommandation CECAF/VI/2, sixième session, Agadir, décembre 1979). Ce fait devrait modifier le modèle de production, dans le sens d’une augmentation du RME, tandis que l’échelle de l’axe des abscisses devrait varier. L’image actuelle par conséquent est plus optimiste que celles que nous montrent les figures présentées.
5) Le RME que montrent les figures est, de ce fait, une sous-estimation du RME réel; cette estimation pourrait servir de marge de sécurité au moment d’attribuer les contingents de capture par exemple.
6) L’absence de corrélation des cpue japonaise et espagnole pour les années 1976 et 1977 pourrait être due a des causes que nous avons déjà mentionnées: changement de zone de pêche de la part de la flottille japonaise, son remplacement par la flottille coréenne, et le changement de la base de mesure de l’effort de la part de la flottille espagnole.
Tous ces éléments permettent de constater la nécessité d’une étude approfondie de cette pêcherie dans les prochaines années.
7) Le fait qu’il se soit produit une expansion progressive de la zone de pèche au cours de la période d’exploitation peut nous donner par contre une vue optimiste de sa situation actuelle, qui se traduirait par une valeur M estimée plus basse que la valeur réelle. Nous pensons pour notre part qu’il est possible que dans ce stock, du moins à de faibles niveaux d’abondance, les recrutements se soient réduits de façon presque proportionnelle à l’augmentation de la mortalité due a la pèche, avec une forte chute dans la partie droite des courbe? de production; c’est-à-dire que, si le modèle concordait mieux avec une courbe de Fox qu’avec une courbe de Schaefer, cela serait dû au fait que nous n’aurions pas tenu compte de ladite expansion de la zone de pèche.
8) Il faudrait savoir exactement à quoi est due l’augmentation de la cpue de la flottille japonaise, car cela pourrait être lié à un changement de fond de pèche. S’il en était ainsi, il faudrait à l’avenir étudier plus 5 fond la structure du stock, du point de vue de la dynamique.
La figure 27 montre des paires de valeurs de la cpue du Groupe de travail du COPACE au cours d’années successives, selon une échelle logarithmique.
L’interprétation de cette figure n’est pas simple, car il y a deux facteurs qui doivent intervenir fortement. D’une part, l’effort exercé qui fait apparaître une corrélation bâtarde entre la cpue (i) et la cpue (i + 1).
Effectivement, les deux nuages de points correspondent aux deux grandes périodes de la pêcherie; une période qui va de 1964 à 1971, avec une faible intensité d’effort, et une autre qui va de 1972 à 1977 avec une haute intensité.
Mais il est un autre facteur qui doit sans aucun doute avoir une influence dans cette figure, à savoir le recrutement qui se produit chaque année. Ce que nous voyons, c’est que les “oscillations” qui se produisent entre deux paires successives de valeurs sont faibles; disons que la figure est trop régulière par rapport à ce que nous escomptions ou, ce qui revient au même, que les nouveaux recrutements ne paraissent guère influer sur la régularité de la figure.
Etant donné que nous ne sommes pas en mesure, par manque de données, d’établir une relation stock/recrutement, nous ne poursuivrons pas la discussion de ce point, mais il faut se souvenir que, en raison du type de croissance du poulpe telle que nous l’avons décrite, on ne trouve pas dans la pêcherie, en nombres abondants, plus de deux classes annuelles. Si l’affirmation qui précède s’avérait exacte, il serait nécessaire de surveiller strictement la dimension du stock reproducteur qui représente un élément important du recrutement ultérieur.
![]()
![]()
![]()