![]()
![]()
![]()
por
A.D. MacCall
(Presiente)1
| RESUMEN |
| En este informe se trazan algunas directrices generales para el estudio y monitoreo de recursos pesqueros variables y se facilita información que puede resultar útil para evaluar las necesidades y ayudar a elegir métodos y estudios apropiados. Se identifican los principales síntomas de variaciones adversas y colapso potencial de los recursos pesqueros y se examinan varias cuestiones relativas al monitoreo de recursos, operaciones de pesca y muestreos biológicos. |
Para ser plenamente eficaces, las investigaciones y el monitoreo biológicos han de hacerse teniendo en cuenta el contexto pesquero y los métodos alternativos. En este informe se examinan algunos de estos factores y se revisa su evolución reciente con la finalidad de que pueda servir de complemento a los numerosos manuales ya existentes. Es difícil hacer recomendaciones fuera del contexto ambiental, biológico y socioeconómico de una pesquería concreta. En algunos casos las necesidades generales son claras. Para los casos restantes, este documento ofrece información que puede ser útil para evaluar las necesidades y escoger entre los distintos métodos y sistemas de estudio existentes.
Este informe ha sido preparado por el Grupo de Trabajo sobre Investigación y Monitoreo de Recursos, que se reunió durante la Consulta de Expertos sobre Recursos Neríticos. La composición del grupo se modificó durante el transcurso de la Consulta. Aparte de sus aportes técnicos, el Sr. R. Jones y la Srta. I. Tsukayama prestaron un útil servicio actuando como relatores. Otras muchas personas participaron regularmente en las reuniones del grupo de trababjo o prepararon secciones importantes de este documento, en particular los Sres. J. Alheit, B. Brown, J. Carscadden, R. Crawford, P. Fréon, la Sra. M.L. García, el Sr. J. Lleonart, la Srta. J. Lopez, el Sr. A. Menz, las Srtas. S. Saccardo y H. Santander, los Sres R. Serra, G.D. Sharp, P. Shelton, E. Ursin y otros.
La ciencia pesquera ha surgido recientemente a partir de una tradicíon europea mucho más antigua, que conocemos con el nombre de “ciencia”. La ciencia tradicional se dedica casi exclusivamente a la búsqueda académica del saber y al establecimiento de la verdad. La ciencia pesquera, en cambio, trasciende esa tradición, porque ofrece una base de trabajo para modificar nuestro mundo: en otras palabras, es una ciencia aplicada. Con frecuencia, en los trabajos pesqueros no se tiene en cuenta las consecuencias de esta realidad que, en muchos aspectos, hace necesario un enfoque diverso del que se enseña en las universidades.
Además del criterio científico tradicional de ampliar el conocimiento, hay también otros criterios que deben guíar nuestros esfuerzos. Dado que los resultados de nuestros trabajos pueden contribuir a cambiar la sociedad y los ecosistemas, los científicos pesqueros llevan una carga de responsabilidad que generalmente no existe en las ciencias académicas normales. Un criterio importante es la oportunidad. Muchos análisis y recomendaciones en materia de pesca son específicos para las condiciones en curso de la pesquería y pierden todo valor si no se dan a conocer rápidamente. Otro criterio, estrechamente relacionado con el anterior, es que las recomendaciones han de ser aplicables. Este criterio es particularmente importante para los fines de esta consulta: las recomendaciones no sólo deben reflejar los conocimientos existentes y la interpretación y aplicación responsable de los mismos, éstas también deben responder a las necesidades de los destinatarios.
1 National Fisheries Services, c/o Southwest Fisheries Centre, P.O. Box 271, La Jolla, CA 92038, Estados Unidos
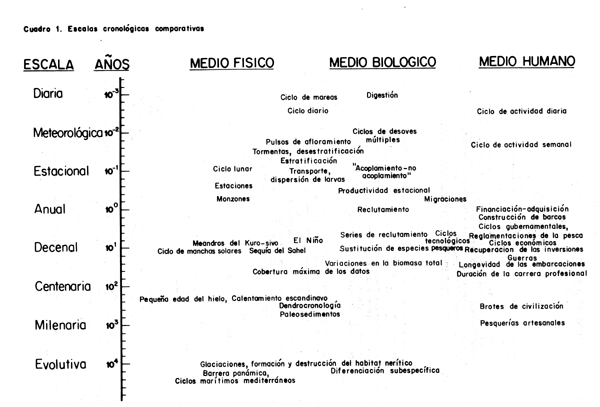
Cuadro 1. Escalas cronológicas comparativas
La ciencia pesquera consiste en aplicar principios ecológicos (en su sentido más amplio) a una pesquería determinada. Así pues, los modelos pesqueros en particular, y la ciencia pesquera en general, deben referirse a una situación concreta, como la ética situacional en sentido filosófico; las técnicas o mediciones, por tanto, no han de considerarse en abstracto, sino en relación con la pesquería particular que se esté examinando. Lamentablemente, por ser una revisión general, en este documento, no ha sido posible seguir este último criterio.
El uso de términos generales como pelágicos o demersales para agrupar a distintos tipos de peces crea una dicotomía que no existe en la naturaleza. Se dice por ejemplo que las especies pelágicas son muy variables, pero el eglefino del Banco Georges ha sido más variable que el arenque del Golfo de Maine, y la caballa, cuando ya se ha reclutado a la población, tiene una tasa de mortalidad natural inferior a la del bacalao. Por esto los científicos pesqueros deberian ser más especifícos al ocuparse de esos grupos, utilizando términos que se refieran a grupos de especies (como sardina/anchoveta) en lugar de términos genéricos como recursos pelágicos. Es preciso, además, indicar explícitamente las características que sirven de base para la formación de esos grupos.
Los recursos pesqueros experimentan variaciones en casi todas las escalas cronológicas. Además, en la variabilidad biológica influyen notablemente la variabilidad física o ambiental por un lado, y la variabilidad del sector humano por otro (Cuadro 1). Buena parte de la dinámica de las pesquerías y de las dificultades en la historia de su explotación y ordenación (por ejemplo, los auges y colapsos) pueden inferirse de la interacción de fuerzas que actúan en escalas de tiempo diferentes. Como en el caso de la ecología clásica de los ciclos predador/presa, los retrasos en la respuesta de la explotación pesquera a la abundancia de un recurso determinan un sistema inherentemente cíclico o inestable. Reconociendo el carácter de esas interacciones y adoptando normas apropiadas y eficaces, los responsables de la ordenación pueden controlar y reducir al mínimo esos ciclos de la tecnología pesquera dentro de los límites de la variabilidad natural del ambiente.
El monitoreo de las pesquerías y los recursos incluye a menudo una vasta selección de variables biológicas. El supuesto implícito en todos esos trabajos es que los cambios en las variables monitoreadas permiten disponer de información sobre el estado de los recursos, en particular sobre su productividad y abundancia. Para evaluar la utilidad de los trabajos de monitoreo pesquero es útil examinar más explícitamente la relación que existe entre las variables biológicas y las causas de cambios en la abundancia de peces.
Las causas de cambios en la abundancia de peces pueden agruparse en cinco categorías:
| - | Dinámica intra-específica, que incluye mecanismos de compensación, como relaciones población desovante-reclutamiento, crecimiento en función de la densidad y canibalismo. |
| - | Competencia entre especies, que reviste considerable interés, pero en la práctica es difícil de demonstrar en forma concluyente. |
| - | Predación, que en general se incluye en los análisis pesqueros dentro de la mortalidad natural. |
| - | Pesca, o explotación. |
| - | Fluctuaciones ambientales, en particular de factores abióticos y factores biológicos del nivel trófico inferior. |
Si son adecuadamente monitoreadas, las variables biológicas pueden dar indicación de las causas de cambio ya mencionadas. En el cuadro 2 se indican las variables biológicas que se suelen monitorear y las causas de variabilidad con que éstas están relacionadas en la pesquería de anchoveta (Engraulis). Casi todas las variables están relacionadas con variaciones causadas por el ambiente, y muchas de ellas están relacionadas (al menos indirectamente) con la pesca. La competencia es una posible causa de variabilidad en muchos casos, pero ninguna de las variables da pruebas sólidas de que así sea. Los mecanismos intra-específicos pueden observarse sobre todo mediante estudio directo de los peces, incluyendo las primeras fases de sus vidas. Estas últimas son las pocas variables que no están relacionadas con la pesca. Algunas variables biológicas proporcionan poca informacón útil sobre las causas de variabilidad, como sucede, por ejemplo, con la proporción de sexos, variable que se suele monitorear pero que dificilmente proporciona información útil sobre las causes de variabilidad.
| CAUSA DE VARIABILIDAD DE LA ABUNDANCIA | |||||
|---|---|---|---|---|---|
| VARIABLE MONITOREADA | |||||
| Tamaño de la población | x | ? | x | xx | i |
| Estructura por edades de los adultos | ? | xx | i | ||
| Tasa de mortalidad por pesca | xx | i | |||
| Tasa de mortalidad natural | xx | i | x | ||
| Indices de predación1 | X | X | |||
| Distribución (a largo plazo) | ? | X | X | ||
| Distribución (a corto plazo) | X | XX | |||
| Intensidad del reclutamiento | X | ? | i? | i | XX |
| Factor de condición/contenido de grasa | X | ? | i | XX | |
| Dieta | X | ? | i | X | |
| Tasa de crecimiento | X | ? | X | X | |
| Fecundidad | XX | ? | i | XX | |
| Edad/talla de madurez | XX | ? | i | XX | |
| Tasa de mortalidad de huevos/larvas | XX | ? | XX | XX | |
| Tasa de crecimiento de larvas | ? | X | |||
| Co-presencia de otras especies2 | ? | ? | ? | ? | X |
| Proporción de sexos | ? | ||||
| Estacionalidad del desove | i | X | |||
2 Incluidos cardúmenes mixtos y superposiciones geográficas/temporales
Naturalmente, el cuadro 2 será diferente para otras especies de peces. Dado que Engraulis se ha estudiado más que la mayoría de las especies, en el caso de otras especies habrá la tendencia a que los datos concluyentes disponibles sean menos. También en algunos casos puede ser oportuno utilizar otras variables biológicas. Por ejemplo, el arenque del noroeste del Atlántico se ha visto afectado en el pasado por enfermedades, por lo que el monitoreo de ese factor puede ser importante.
La evaluación de las poblaciones ícticas tiene por objeto determinar la productividad actual de una población en relación con su productividad potencial. Esta última se calcula a partir de datos históricos sobre los recursos, estudios biológicos e información comparativa sobre especies o pesquerías similares. La experiencia muestra que las poblaciones de peces responden a la explotación en forma predecible, pero a menudo las fluctuaciones ambientales complican esos patrones de reacción. La similaridad fundamental entre las respuestas que se producen en distintas pesquerías ha favorecido el desarrollo de varios modelos que resumen concisamente la relación entre la explotación pesquera y los recursos. Por ejemplo, a medida que aumenta la intensidad de la explotación pesquera, disminuyen la abundancia, la edad o la talla media de los peces y la edad a la primera madurez. Estos cambios reflejan sencillamente la presión ejercida por la pesca y la respuesta compensatoria de la población. Para determinar en base a esos cambios que la población se encuentra en mal estado (es decir, que el rendimiento ha disminuido por debajo del rendimiento potencial debido a la explotación excesiva) es necesario tener en cuenta la historia de los niveles y las fluctuaciones anteriores así como otras características de la población, que también pueden haber variado. Por esta razón será de poca utilidad hacer una discusión generalizada de los indicadores o síntomas de cambios adversos en el estado de una población. De todas maneras, para facilitar el examen de la cuestión, en el cuadro 3 se enumeran algunos de los síntomas e indicadores más comunes, indicándose su interpretación más probable y algunas de las consideraciones que es preciso tener en cuenta.
Antes de llegar a la conclusión de que la presencia de un síntoma significa una situación adversa, es necesario conocer y comprender la naturaleza de la pesquería y de los recursos. Los riesgos de la explotación pesquera varían sustancialmente según la especie y las carácteristicas ambientales, como se indica en el cuadro 4. Teniendo esto presente, es preciso estudiar los síntomas indicados en el cuadro 3 para ver si existen otras causas alternativas, como cambios ambientales o variaciones en la demanda de mercado, que puedan dar respuestas aparentes similares. Una vez que el síntoma ha sido aceptado como indicador válido del estado de una población, es preciso juzgar la gravedad del síntoma sobre una base objetiva. De ordinario para ello se utiliza información histórica en un modelo de pesquerías que tiene métodos de interpretación bien establecidos. El modelo puede indicar además la naturaleza y extensión de las medidas que son necesaries para rehabilitar la población.
Las categorías en que pueden agruparse los síntomas que permiten hacer diagnósticos son pocas, y se refieren a la abundancia, el reclutamiento (peces jóvenes que se añaden a la población) y la intensidad relativa de explotación. Además, hay varios signos de alerta que son más difíciles de interpretar. La abundancia es de ordinario el indicador más importante del estado de una población y, por esa razón, una parte importante de los trabajos de investigación y de monitoreo han de concentrarse en ese aspecto. Si no se dispone de una estimación directa de la abundancia, pueden utilizarse otros indicadores, como la tasa de captura de la pesquería, la extensión geográfica de la población, o la situación de predadores más visibles que dependen de la población, como las aves marinas. Debido a la relativa imprecisión de esos indicadores auxiliares, es conveniente confirmar los datos con información de más de una fuente. Utilizando técnicas analíticas, como modelos de producción, se pueden obtener estimaciones de los niveles de abundancia que son “satisfactorios” (por ejemplo, el nivel de abundancia con el que se obtiene la productividad máxima). Dependiendo de la variabilidad natural del recurso en cuestión, se puede fijar el nivel de “alerta” cuando la población se reduce a la mitad de la abundancia que permite obtener la productividad media máxima. En forma similar, el nivel de “peligro” se puede fijar en una cifra algo menor.
El reclutamiento constituye la fuente principal de biomasa de peces para sustituir las pérdidas de la población debidas a la explotación y a la mortalidad natural. Cuanto mayor es la tasa total de mortalidad, más sensible es un recurso a la variabilidad y/o fallas del reclutamiento. Las fluctuaciones del reclutamiento debidas a la variabilidad del ambiente son, en su mayor parte, inevitables, aunque en el caso de muchos recursos las fluctuaciones del reclutamiento son menores cuando la población reproductura es mayor. Cuando el reclutamiento disminuye al mismo tiempo que decrece la abundancia de la población reproductora, aumentan las posibilidades de que el recurso se agote (“por sobrepesca del reclutamiento”). Cuando se dispone de datos históricos sobre el reclutamiento, es siempre útil graficar los datos de reclutamiento en función de la población reproductora, con o sin una línea ajustada de regresión. Esto permite identificar la relación media y además, lo que a menudo es más importante, permite determinar la existencia de series de reclutamientos por encima - o por debajo - de lo normal. La experiencia de Perú y Namibia ha mostrado que la presencia de un patrón anómalo en el contenido de grasa o de aceite en los peces advierte por anticipado de condiciones oceánicas también anómalas (por ejemplo “El Niño”) y de la consiguiente probabilidad de una falla futura en el reclutamiento.
| SINTOMAS E INDICADORES | CONSIDERACIONES | INTERPRETACION |
|---|---|---|
a) Disminución de la abundancia |
- Influencia del ambiente |
Alerta, establecer un nivel de referencia |
- Cambios en la disponibilidad y la vulnerabilidad |
||
Disminución de la tasa de captura
(CPUE) |
- Historia y definición del esfuerzo |
Alerta, peligro (la CPUE subestima a
menudo la magnitud de la disminución
real), probar con modelos de producción |
-Disponibilidad, vulnerabilidad |
||
Reducción del área de
la población de
distribución |
- Influencia del ambiente |
Alerta |
Variación en la
composición por especies |
- Influencia del ambiente |
Alerta |
- Cambios en otras especies |
||
- Cambios en el mercado, los reglamentos, la estructura de la pesca |
||
Cambio en los indices de predación |
- Influencia del ambiente |
Alerta |
- Disponibilidad para los predadores |
||
b) Disminución del reclutamiento |
- Influencia del ambiente |
Alerta, establecer un nivel de referencia,
estudiar la relación
poblaciónreclutas, comparar el nivel de
sustitución |
- Disponibilidad, vulnerabilidad |
||
Aumento de la edad media |
- Mercado, reglamentos, estructura de la pesca |
Peligro, falla del recluta miento |
Ciclo graso anómalo |
- Influencia del ambiente |
Peligro |
- Patrones normales |
||
c) La mortalidad por pesca se acerca a la mortalidad
natural |
- Influencia del ambiente |
Peligro, tratar rendimiento por recluta,
modelos de producción |
- Cambios en la disponibilidad, vulnerabilidad |
||
La edad o la talla media se acercan a la edad o talla
a a la primera madurez |
- Influencia del ambiente |
Peligro |
- Disponibilidad, vulnerabilidad |
||
- Cambios en los mercados, reglamentos, la estructura
de la pesca |
||
d) Captura variable (despues de un aumento de las
capturas |
- Influencia del ambiente |
Alerta |
- Mercado, reglamentos |
||
e) Desviación de los patrones
normales |
Alerta |
|
Cambios en los patrones de desove o de reclutamiento |
- Influencia del ambiente |
|
-Disponibilidad |
Alerta |
|
Cambios en la edad/ talla de madurez |
- Influencia del ambiente |
Alerta |
Cambios en la fecundidad |
- Influencia del ambiente |
Alerta |
Cambios en la composición por tallas
de las capturas |
- Influencia del ambiente |
|
- Cambios en la disponibilidad, vulnerabilidad |
Alerta |
|
- Cambios en los mercados, los reglamentos, la
estructura de la pesca |
| PLATIJA DEL MAR DEL NORTE | EGLEFINO DEL MAR DEL NORTE | ARENQUE DEL MAR DEL NORTE | ARENQUE ATLANTICO- ESCANDINAVO | ANCHOVTETA PERUANA | SARDINA DE CALIFORNIA | |
|---|---|---|---|---|---|---|
| AMBIENTE MARINO | Estable | Estable | Estable | Moderadamente estable | Inestable (afloramiento) | Inestable (afloramiento) |
| DINAMICA DE LA POBLACION | ||||||
- Grado de compensación del reclutamiento | alto | indeterminado | relativamente alto | moderado | bajo | muy bajo |
- Variabilidad del reclutamiento | baja | muy alta | moderada | espasmódicamente alta | baja-moderada (con fallas) | moderada |
- Longevidad (inversa a la tasa de mortalidad natural) | larga (20+años) | media (12+años) | media (12+años) | medio-larga (15+años) | corta (4+años) | media (10+años) |
- Duración de la fase de premadurez | larga (3-4 años) | media (2 años) | media (2 años) | media (2-3 años) | breve (< 1 año) | media (2 años) |
- Porcentaje del período de creci- miento (peso) que se extiende despues del reclutamiento | grande (90%) | grande (80%) | pequeño (40%) | medio (60%) | pequeño a medio (50%) | medio (50%) |
| COMPORTAMIENTO DE LOS PECES | ||||||
- Hábitos | demersales | demersales | pelágicos | pelágicos | pelágicos | pelágicos |
- Protección o refugios ambientales | parcial | parcial | ninguno | ninguno | ninguno | ninguno |
- Tendencia a formar cardúmenes | ligera | alguna | fuerte | muy fuerte | fuerte | fuerte |
- Facilidad de detección | indetectable | limitada | fácil | fácil | fácil | fácil |
- Dependencia (inversa) de la captu- rabilidad (q) respecto de la abundancia | ninguna | probablemente ninguna | probablemente fuerte | muy fuerte | fuerte | fuerte |
- Vulnerabilidad al aumento intenso de la explotación | elástica | elástica | vulnerable | muy vulnerable | muy vulnerable | muy vulnerable |
| PERSPECTIVAS PESQUERAS GENERALES | Continua y fiable a breve y largo plazo | Muy irregular a corto y a medio plazo; probable- mente fiable a largo plazo | Relativamente constante en el corto a medio plazo; fiabilidad incierta a largo plazo | Espasmódica; sospechosa fiabilidad a largo plazo | No fiable en el medio a largo plazo, con cambios repentinos | No fiable; inestable a largo p zo |
| RIESGO DE LA EXPLOTACION EN AUMENTO | ||||||
La actividad pesquera repercute en un recurso sobre todo a causa de la mortalidad adicional (mortalidad por pesca) que impone. Parece existir una relación coherente entre la magnitud de la tasa de mortalidad natural (M) y los niveles sostenibles de mortalidad por pesca (F). Esa relación constituye la base de la regla empírica popular según la cual F⋍M, pero la experiencia ha mostrado que a menudo esta regla sobrestima la tasa de mortalidad por pesca (F) óptima. Por ello, el acercarse de la tasa de mortalidad por pesca a la tasa de mortalidad natural constituye un síntoma de sobrepesca. Otro indicador alternativo es que la edad o talla media en la captura esté próxima a la edad o talla de primer desove. Además de indicar que la presión de pesca es relativamente alta, esa situación indica una inminente disminución del reclutamiento debido a falta de potencial de desove.
Otras varias condiciones de la población pueden tomarse como signo de alerta, pero no ofrecen diagnósticos específicos. Por ejemplo, el aumento de las fluctuaciones de la captura sugiere la posibilidad de que haya una pérdida de capacidad de compensación y un mayor riesgo de colapso. De igual forma, las desviaciones respecto de los patrones fisiológicos o de comportamiento normales pueden deberse a fluctuaciones ambientales o pueden ser respuestas normales a la presión de pesca, pero no han de desecharse sin antes haber considerado el posible deterioramento del estado de la población.
Es importante observar que el cuadro 3 no es exhaustivo. Pueden emplearse también otros muchos síntomas e indicadores, algunos de los cuales pueden tener utilidad general y otros utilidad específica en pesquerías determinadas. Una vez más hay que subrayar que cada pesquería es, en ciertos aspectos, una realidad única y que, por tanto, los síntomas y consideraciones pertinentes varían de un caso a otro.
La información necesaria para el monitoreo de una pesquería puede obtenerse directamente de las operaciones de pesca, por muestreo o mediante estudios biológicos. Existe una notable interacción entre esos tipos de información, ya que todos ellos están en relación con las actividades de la pesquería.
5.1.1 Documentación histórica
Los cambios secuenciales que se producen en las condiciones de mercado, la capacidad de elaboración, la legislación o los reglamentos y toda una serie de acontecimientos o procesos condicionados por el ambiente tienen efectos directos en las faenas de pesca. Por tanto, es preciso documentarlos para disponer de informentación sobre las razones del paso de una situación a otra en la pesquería. Este trabajo de documentación no cuesta mucho (basta un simple diario de crisis o acontecimientos), pero raramente se hace. En las series largas de datos pesqueros se encontrarán a menudo variaciones repentinas debidas a problemas de ese tipo y quienes han de analizar los datos necesitan esta información, para evitar interpretaciones equivocadas y errores sistemáticos en sus estudios.
Los datos sobre las fases de desarrollo histórico de una pesquería han de incluir información sobre: cambios en los métodos o artes de pesca; evolución de los mercados (es decir, nuevos productos, nuevos métodos de elaboración, etc.); perturbaciones de los mercados (por ejemplo, huelgas de mano de obra o de transporte, colapso de los recursos, legislación en materia de contaminantes, saturación del mercado debida a exceso de producción, etc.); reglamentos de ordenación y mercadéo; y perturbaciones condicionadas por el ambiente (como el fenómeno “El Niño”) y cambios a largo plazo en la distribución, composición y abundancia de especies.
5.1.2 CapturaLa base de toda actividad de producción y mercadeo de pescado es la especie que constituye el recurso. Incluso en el caso de especies muy afines existen claras diferencias en la aceptabilidad del producto y, por tanto, en su valor en los mercados locales y regionales. La elaboración y distribución de los productos varía según sus características de mercado. Incluso dentro de una misma especie son posibles utilizaciones y valores diversos, como en el caso de las huevas de arenque y capelán, o en el uso de pescado de calidad selecta para sashimi y de otras calidades para la fabricación de conservas o de harina.
En las pesquerías menos complejas, donde sólo se explotan y descargan una o pocas especies, el problema de identificación de las especies es mínimo. En cambio, las pesquerías multiespecíficas o las pesquerías de gran volumen (para fabricación de harina, por ejemplo), a menudo no dan la posibilidad de obtener una estratificación por especies a la hora de recoger los datos. En otros casos, las capturas de pesquerías complejas de especies múltiples se clasifican en el mercado. La interpretación de los datos de captura y descarga procedentes de pesquerías complejas es potencialmente importante para determinar los cambios en la distribución y abundancia y en la composición por especies.
En la mayoría de los casos (alugunas pesquerías artesanales podrían constituir una excepción) la captura es el dato pesquero más importante. Si bien la abundancia de los recursos puede estimarse por vías independientes, el volumen de la captura constituye una indicación directa del impacto de la pesca en los recursos, y es poco lo que puede hacerse en materia de planificación o evaluación pesquera sin disponer de información sobre la captura.
5.1.3 Registros de pesca
Un sistema bien diseñado y aplicado de cuadernos o registros de pesca puede ofrecer valiosos datos cuantitativos y cualitativos sobre una pesquería. La información facilitada por esos registros puede incluir capturas, descargas, esfuerzo, ubicación de las capturas, identificación de poblaciones, distribución, co-presencia de especies diferentes, y descartes. Como la colección de la información depende de la cooperación de personal no cientifico, existe el peligro de que los datos no sean colectados correctamente y, por lo tanto, resulten inútiles o, peor aún, conduzcan a error.
Además de facilitar información cuantitativa y cualitativa sobre la pesquería, los registros de pesca tienen la ventaja de ser relativamente poco costosos en términos de equipo, facilitando además índices a largo plazo de la abundancia. Sin embargo, la complación y estudio de los registros de pesca depende de la cooperación de personal no técnico, cuya actitud puede variar si consideran que la información contenida en esos registros puede influir en la reglamentación de la pesca (en el establecimiento de cuotas por ejemplo). Se trata además de una procedimiento que exige relativamente mucho personal y que da información que es dificil de verificar.
La poca fiabilidad de la captura por unidad de esfuerzo (CPUE) como índice de la abundancia está bien documentada, y por ello se recomienda que los sistemas de registros de pesca se complementen con estimaciones independientes de la abundancia. El diseño de los registros de pesca variará probablemente de una pesquería otra, según factores tales como las actividades de los pescadores, el grado de alfabetización de los mismos y la cantidad deseada de datos. Lo fundamental es que el registro de pesca sea simple y que, al mismo tiempo, proporcione información suficiente. Es claro que el diseño de los registros de pesca irá evolucionando a medida que los cientificos vayan viendo lo que los pescadores pueden o quieren hacer y a medida que se vaya obteniendo información más o menos valiosa. Ha de evitarse el uso de varios registros de pesca (por ejemplo, uno para cada especie o para cada institución); para evitar confusiones, conviene combinarlos en uno solo. La información recogida variará según la pesquería y sus productos. A la hora de diseñar un registro de pesca es preciso consultar la bibliografía existente sobre ese tema y sobre determinación del esfuerzo de pesca (véase también la sección siguiente) y conviene ponerse en contacto con especialistas en ciencias pesqueras que tengan experiencia en el uso de sistemas similares.
Son varios los factores que hay que tener en cuenta para la adecuada aplicación de un buen sistema de registros de pesca. Es imperativo que el científico mantenga estrechos contactos con los pescadores, preferiblemente con visitas personales para recoger los registros y cambiar impresiones sobre ellos y sobre la información que contienen. Hay que asegurar el carácter confidencial de la información. Dado que, para la recogida de los datos, el científico depende de la colaboración de personal no técnico (es decir, los pescadores), es necesario disponer de algún método para verificar la información. Probablemente no existe ningún método perfectamente válido para confirmar todos los datos, pero el problema es capital y debe ser planteado. Puede ser necesario utilizar varias técnicas de comprobación. Una vez que los datos hayan sido analizados, hay que poner los resultados en conocimiento de los pescadores, sirviéndose para ello de términos no técnicos.
5.1.4 Determinación del esfuerzo de pesca
Existe una vasta bibliografía sobre la determinación del esfuerzo, pero subsisten aún graves problemas. Uno es el de la terminología. Beverton y Holt han utilizado el término “esfuerzo” para referirse a un índice de la tasa instantánea de mortalidad por pesca (suponiendo la proporcionalidad). Gulland y otros, más tarde, han utilizado el término “esfuerzo” para indicar una medida de la actividad nominal de pesca. Esta evolución fue desafortunada, ya que primero se dió por sentado lo que estaba en discusión (la proporcionalidad) y luego se generalizó el uso del término. Probablemente convendría descartar el término “esfuerzo”, pero su uso está hoy firmemente afianzado. La mejor solución, por tanto, parece ser reservar la palabra “esfuerzo” para referirse a la actividad nominal de pesca, y para referirse a la “tasa de mortalidad por pesca” usar estas mismas palabras.
El uso de los índices de captura por unidad de esfuerzo (CPUE) ha ido dando lugar a un número creciente de resultados sesgados y equívocos. Una de las principales razones de ello es el contenido informativo que entraña la unidad nominal de actividad de pesca. En términos de teoría de la información, los pescadores proceden a una búsqueda en los caladeros para reducir la incertidumbre de conseguir capturas. Si la información conseguida está en estrecha relación con el tiempo o distancia de esa búsqueda, puede constituir una unidad nominal útil de medición del esfuerzo. Sin embargo, la tecnología moderna proporciona a los pescadores otras informaciones importantes y a menudo preponderantes, a través de la radio y de reconocimientos aéreos; información que de ordinario no se incluye en la unidad nominal del esfuerzo. La consecuencia es que el índice de CPUE tiende a ser insensible a los cambios en la abundancia de los peces. A este propósito, vale la pena hacer tres recomendaciones: (1) puede ser más conveniente definir el esfuerzo nominal en términos del mínimo contenido de información (por ejemplo, viajes que se hacen el lunes o después de la luna nueva, cuando los pescadores tienen menos datos sobre la ubicación de los peces); (2) es preciso ir directamente, siempre que sea posible, a la fuente de la información, consultando, por ejemplo, los registros de las observaciones aéreas, anotando el trayecto de vuelo y las cantidades estimadas de peces observados; y (3) puede ser más útil definir una medida precisa de la CPUE, con un sesgo conocido, que una CPUE imprecisa que quizás esté menos sesgada (que proporcionalmente esté más cercana a la abundancia). Si se conoce el sesgo, es posible tenerlo en cuenta explícitamente.
Se recomienda proceder a dos estudios complementarios del esfuerzo. El primero consiste en un análisis de los componentes biológicos y de comportamiento humano que intervienen en la operación de pesca. En algunos casos puede prepararse un modelo simulado y evaluar el rendimiento de varios índices de CPUE. Los primeros en hacer esto fueron Paloheimo y Dickie (1964) y posteriormente se ha intentado en varias pesquerías (por ejemplo, la Comisión Interamericana del Atún Tropical ha preparado recientemente un modelo de este tipo para las pesquerías de atún del Pacífico tropical oriental). La utilidad de esos modelos ha sido muy variada y no está dicho que esos trabajos resulten productivos.
El segundo estudio sería una comparación mundial de los índices de CPUE de donde se disponga de estimaciones fidedignas independientes de la abundancia. Podrían obtenerse para cada pesquería varios índices de CPUE y evaluar su desempeño. Podrían establecerse patrones de semejanza entre medidas similares de la actividad nominal de pesca y entre recursos de biología similar y/o sometidos a operaciones de pesca semejantes. Un estudio de ese tipo, además de ser útil para ayudar a elegir la definición del esfuerzo nominal, contribuiría a documentar la fiabilidad general y la validez de las medidas de la captura por unidad de esfuerzo.
5.1.5 Selectividad
El problema de la selectividad es bien conocido en el caso de las especies demersales y se han propuesto ya soluciones apropiadas. En las pesquerías pelágicas, especialmente cuando se usan redes de cerco de jareta, se suele suponer que no hay selectividad. De hecho, pueden darse tres tipos de selectividad según la talla. En primer lugar, existe una selectividad indirecta de la luz de la malla que raramente se observa porque el tamaño de la malla se adapta usualmente a la talla de los peces que se desea capturar, y los pescadores no tienen ningún interés en tener peces enmallados en la red. En segundo lugar, los pescadores a menudo son capaces de estimar la talla de los peces antes de calar las redes y dejan libres a los peces que consideran demasiado pequeños para usos comerciales. Así pues, la selección, en este caso, no la hace el arte sino el pescador. Esta forma de selectividad tiende a variar según la abundancia de la población y las condiciones económicas, lo que complica los índices de CPUE. En tercer lugar, el efecto más importante de la selectividad depende del comportamiento de los peces. Los peces se juntan en cardúmenes según la talla y las concentraciones de cardúmenes suelen estar formadas por grupos de individuos de talla media similar. Por ello, se recomienda estratificar el muestreo todo lo posible, para tener en cuenta la estructura de toda la población. En algunos casos se ha observado una selectividad por sexos en especies pelágicas: por ejemplo, en algunos cardúmenes predominan significativamente los machos o las hembras, con diferente vulnerabilidad a los artes de pesca, particularmente durante el ciclo de desove.
5.2.1 Especies y poblaciones: consideraciones en relación con la multiplicidad de especies
Los planes de monitoreo y muestreo actualmente utilizados en la mayor parte de las pesquerías facilitan información razonablemente útil por especies o categorías genéricas, pero, en muchos casos, debido al carácter integrativo de algunas pesquerías o a la superposición de especies muy afines, no se han obtenido datos que hagan posible estratificar las capturas por especies o, más defícil aún, por grupos subespecíficos o “poblaciones”, como es necesario para la preparación de modelos y los trabajos de monitoreo.
En la mayoría de los casos, es posible establecer categorías mediante identificación visual, ayudándose quizás con informaciones sobre la procedencia de las capturas. Pero hay casos en que la identificación de la población es extremamente difícil, como, por ejemplo, en el caso de peces anádromos, como es el salmón, o de desovadores locales, como el arenque. En la sección 5.3.3 se indican algunos métodos para distinguir las distintas poblaciones.
Los muestreos destinados a monitorear las capturas de especies importantes han de diseñarse teniendo en cuenta la multiplicidad de especies presentes en la pesquería. A menudo sólo se han tomado muestras de la especie predominante. Posteriormente, a medida que la pesquería cambia y otras especies aparecen con mayor abundancia en las capturas, sucede que no se dispone de información anterior sobre esas especies que ahora predominan en las capturas. Por ello, es preciso determinar la estructura por especies de toda la captura, procediendo luego a determinar la estructura de edad y talla de cada especie. Por último, es preciso determinar las características biológicas de la especie seleccionada: talla, peso, madurez, etc.
El plan de muestreo ha de ser representativo, tanto desde el punto de vista biológico como desde el punto de vista estadístico. El muestreo, además, ha de ser consistente para que sea posible hacer comparaciones entre los datos correspondientes a distintos años o diferentes lugares. Para preparar el plan de muestreo y determinar el tamaño de las muestras han de usarse consideraciones estadísticas, pero ni el diseno ni el tamaño de las muestras han de modificarse de un año para otro. Cuando se cambien los métodos de muestreo, es preciso anotar en detalle tales cambios, para que los datos puedan interpretarse adecuadamente en el futuro.
5.2.2 Muestreo en el mar
Por razón de los costos, conviene de ordinario obtener todos los parámetros a partir de datos del pescado descargado (muestreo en mercado). Cuando ello no sea posible, porque el pescado se elabora a bordo, se pueden embarcar técnicos para que hagan el muestreo a bordo. Cuando no es posible monitorear las embarcaciones pesqueras mientras se hallan en el mar, resulta necesario utilizar un buque de investigación. Dos sistemas son posibles en ese caso. Se pueden establecer factores de corrección, para estandarizar la información obtenida en los mercados con la obtenida a bordo (es decir, factores de transformación, categorías comerciales, talla media, etc.). Como última solución, se puede proceder a hacer todas las estimaciones con buques de investigación. En todo caso, se recomienda controlar la validez de los datos obtenidos en los mercados mediante trabajos a bordo.
5.2.3 Producción industrial
A veces es útil monitorear los productos ya elaborados del pescado descargado. Como ejemplos pueden citarse la producción de harina de pescado, aceite de pescado, filetes de pescado o productos enlatados. Esta información es de ordinario exacta y no deberia ser difícil de obtener de las empresas pesqueras y las plantas de elaboración. Una serie cronológica de datos permite a los responsables de administrar los recursos, disponer de una perspectiva inmediata de las tendencias en la calidad y/o valor del pescado descargado. Por ejemplo, en las pesquerías de cerco del Africa austral, el agotamiento de los recursos de sardina (Sardinops ocellata), jurel (Trachurus trachurus) y caballa (Scomber japoncius) ha determinado una considerable disminución de la producción de pescado enlatado, a pesar de que las capturas totales (incluyendo todas las especies) se han mantenido relativamente constantes.
El contenido de grasa de los peces refleja a menudo su estado fisiológico y de desove y puede constituir un indicador útil de dificultades en el desove. Algunas veces es fácil y poco costoso obtener información sobre el contenido de grasa de los peces a partir de los productos elaborados. Este sistema, sin embargo, resulta menos válido cuando en la pesquería se explota más de una especie, dado que puede haber diferencias en el contenido de grasa. En ese caso, lo mejor es determinar el contenido de grasa de cada especie a partir de muestreos en los puntos de descarga.
5.2.4 Edad y talla
Los métodos de determinación de la edad (particularmente las claves edad-talla), una vez establecidos, tienden a considerarse como un elemento estable del sistema de monitoreo. Ello puede llevar a errores (Kimura, 1977) y debe evitarse incluyendo en el sistema de monitoreo una reevaluación periódica de los métodos y de la interpretación. Dado que cada vez se recurre más a menudo a comparaciones de distintas pesquerías a nivel internacional y regional y dado que muchos recursos se extienden a través de las fronteras políticas, es muy conveniente uniformar los métodos de determinación de la edad y medición de la talla de los peces. Cuando se procede a trabajos de monitoreo de un recursos en dos o más países, es esencial llegar a un acuerdo sobre las técnicas de medición (por ejemplo, grupos de talla y fecha nominal de nacimiento).
Por último, el desarrollo reciente de métodos que son atrayentes y que se basan exclusivamente en la composición por tallas para el análisis de las poblaciones de peces no debe impedir que se hagan esfuerzos serios para utilizar métodos más tradicionales para la atribución de edades absolutas a los peces. Es necesario además examinar atentamente los métodos de frecuencia de tallas para determinar su precisión y los posibles sesgos.
5.2.5 Fecundidad
La determinación de la fecundidad por puesta solía constituir un problema en el caso de desovantes múltiples. De ordinario se establecían diagramas de volumen-frecuencia de los ovocitos y se suponía que el grupo modal más avanzado representaba el grupo de la próxima puesta. Este método carece de precisión y requiere muchísimo tiempo. Recientemente se ha desarrollado un método nuevo (Alheit et al., 1983), en el que sólo se cuentan los ovocitos hidratados, dado que son éstos los que representan exactamente la próxima puesta. Para este método se requiere colectar hembras en el momento justo del día, pocas horas antes de que desoven.
5.2.6 Madurez
La madurez se determina de ordinario mediante criterios macroscópicos, pero este método tiene la desventaja de ser subjetivo y carecer de precisión. Además, su utilidad es dudosa, porque reciéntemente se ha descubierto que los peces con desoves múltiples (por ejemplo, las anchovetas) pueden proceder a una nueva puesta cada seis u ocho días durante el período de desove. Ello significa que en una sola semana atraviesan de tres a cinco fases de madurez, según los criterios macroscópicos que se utilicen. Probablemente, algunas de esas fases no duran más que pocas horas. Los criterios macroscópicos para la determinación de la madurez son, por tanto, de valor limitado en el caso de especies con un desarrollo tan rápido de los ovocitos.
5.2.7 Proporción sexual
Reciéntemente se han encontrado algunos problemas en la determinación de la proporción sexual en anchovetas (Engraulis). Como parte de su comportamiento reproductor, los machos y las hembras tienden a segregarse de los otros cardúmenes cuando se aproxima el momento del desove. Estos “cardúmenes de desovantes” son muy vulnerables a la captura y de ordinario tienen una preponderancia de machos. También al acercarse el momento del desove, las hembras son más vulnerables a la captura que en otros momentos del ciclo de desarrollo de los ovocitos.
5.2.8 Factor de condición
Algunas investigaciones recientes han mostrado que la hidratación de las gónadas de las anchovetas puede hacer aumentar el peso de las hembras hasta en un 30 por ciento en el plazo de pocas horas. Además, si en un cardúmen se encuentran hembras con las gónadas hidratadas, el porcentaje de éstas suele ser muy elevado. Es preciso, pues, tomar precauciones para modificar el muestreo adecuadamente y/o preparar métodos de conversión para esos casos.
5.3.1 Crecimiento
Hay dos tipos de estudios (excluyendo los trabajos fisiológicos) que han proporcionado información útil sobre el crecimiento; estos son el marcado y el examen de los anillos de crecimiento diario. El marcado puede proporcionar información sobre el crecimiento cuando no se dispone de otras fuentes, etales como marcas de crecimiento anual, y es también útil para verificar otros métodos de estimación de la edad y del crecimiento, como el análisis modal de las frecuencias de tallas. El examen de los anillos de crecimiento diario es potencialmente muy útil, pero aún no se han resuelto algunos problemas que plantea. Algunos trabajos recientes sugieren que la frecuencia de los anillos de crecimiento puede estar relacionada con la tasa de crecimiento, pero otros estudios han mostrado que hay discontinuidades.
5.3.2 Contenidos estómacales
La interpretación de los contenidos del estómago es notoriamente difícil, por dos razones. Primeramente, la falta de tiempo o experiencia suficientes hacen que muy a menudo no sea posible determinar a qué especie pertenecen los alimentos presentes en el estómago. Sin embargo, la mayoría de los índices de similaridad ecológica utilizados para comparar los espetros alimentarios de diferentes especies de peces sólo son significativos si se aplican a nivel de la especie que sirve de presa. El segundo problema es que resulta extremamente difícil determinar las tasas de digestión. Sin embargo la cuantificación del consumo de alimentos depende íntimamente de una estimación razonable de las tasas de digestión, las que probablemente varían incluso para diferentes tipos de presas.
5.3.3 Identificación de poblaciones
En último término, por población, “stock” o más precisamente por población explotable, se entiende sencillamente aquella parte de la población que constituye la base de los trabajos de análisis y ordenación. Por tanto, un stock o población explotable puede definirse de muchas maneras, algunas de las cuales resultan más útiles que otras. Son varias la cuestiones que han de tenerse en cuenta, y no todas tienen una base biológica.
Reciéntemente, en el Simposio Internacional sobre el Concepto de Stock, Booke (1981) dió una definición del término “stock” o población para uso en las ciencias pesqueras. según su difinición general (es decir, vaga), “un stock es un grupo de individuos de una misma especie, o una población de peces, que se mantiene y sustenta a lo largo del tiempo en un área definible. Con más precisión, cuando se conoce el genotipo, se puede definir idealmente un stock de peces que conserva y mantiene el equilibrio de Castle-Hardy Weinberg. Cuando no se dispone de una base genética para caracterizar a una población, han de examinarse las características fenotípicas, para ver si una colectividad de peces las mantiene y si, por tanto, puede considerarse como un stock o población.” Esta última perspectiva es la que generalmente se aplica en la actualidad. Considerando el rol tutelar de la ordenación pesquera, Booke insiste también en la necesidad de tener presente la variabilidad genética de las poblaciones en los trabajos de ordenación.
Probablemente quienes se dedican a la evaluación de poblaciones estén más familiarizados con la asignación convencional de la etiqueta de “stock”, basadas en los artes de pesca, la geografía (por ejemplo, por hemisferios oceánicos) o la simple agrupación por especies (por ejemplo, albacora del sur del Atlántico, arenque del Báltico, etc.). En el caso de la mayoría de las especies de peces, las hipótesis para determinar la estructura poblacional son numerosas y muy tenues y esto ha llevado a que para fines prácticos se tiendan a definir los “stocks” o poblaciones en base a consideraciones hemisféricas o regionales.
Como punto de partida para delimitar un “stock” o población, es evidente que el lugar geográfico donde se verifica la reproducción constituye un criterio importante, pero hay que tener presente que una misma especie puede presentar patrones de reproducción variables y estratificados geográfica y temporalmente; lo que puede dar lugar, dentro de un contexto geográfico dado, no a poblaciones panmícticas simples sino a poblaciones más complejas. Esa complejidad en las estructuras de edad entraña complejos patrones temporales y espaciales de reproducción.
| TECNICA | METODO | COSTO | TIPO DE DATOS | DETERMINACION DE LAS CARACTERISTICAS DE LA POBLACION | DEFINITIVO O NO PARA LA DIFERENCIACION | ASPECTOS: 1. EN CONTRA, Y 2. A FAVOR |
|---|---|---|---|---|---|---|
| A1 Caracterización de proteínas | a. electroforesis | bajo | no paramétricos | dependiente del tamaño de la muestra | si | 1. dificultad de muestreo |
| 2. base genética | ||||||
| b. electroenfoque | alto | no paramétricos | dependiente del tamaño de la muestra | si | 1. procedimiento lento | |
| 2. elevada resolución | ||||||
| c. purificación y funcional | muy alto | paramétricos | dependiente del tamaño de la muestra | puede serlo | 1. técnica difícil | |
| 2. gran sensibilidad | ||||||
| A2 Comparaciones de cromosomas | a. determinación del cariotipo | bajo | subjetivos | puede ser útil | puede serlo | 1. requiere grandes diferencias |
| 2. base genética | ||||||
| b. estudios de las bandas | moderado | subjetivos | puede ser útil | si | 1. requiere cromosomas grandes | |
| 2. base genética | ||||||
| A3 ADN mitocondrial | a. aislamiento y fracciona- miento | moderado | no paramétricos | dependiente del tamaño de la muestra | puede serlo para estudios de familias o diferenciación por áreas | 1.
procedimiento complicado |
| 2. herencia de la madre | ||||||
| A4 Estructuras de color | a. estructuras de pigmenta- ción | bajo | no paramétricos | puede ser útil | puede serlo | 1. es necesario deter- minar la base: es decir, ontogenéticas o hereditarias |
| 2. de ordinario, base genética | ||||||
| A5 Inmunología | a. tipificación de los tejidos: de la sangre | bajo | no paramétricos | dependiente del tamaño de la muestra | si | 1. sensible al ambiente |
| 2. base genética | ||||||
| b. fijación del microcomplemento | moderado | no paramétricos | demasiado sensible | si | 1. muy sensible | |
| 2. útil a nivel de especie | ||||||
| A6 Estudios numéricos o de metrificación | a. dimensiones de las partes duras | bajo | paramétricos | puede ser útil | si | 1. es necesario determinar la base; las muestras deben tener el mismo valor medio. |
| 2. de ordinario específico regionalmente, a menudo con base genética | ||||||
| b. morfometría de las dimensiones corporales | bajo | paramétricos | puede ser útil | puede serlo | 1. es necesario determinar la base; las muestras deben tener la misma probabilidad de ser tomadas | |
| 2. de ordinario específico a nivel regional a menudo con base genética | ||||||
| B1 Parámetros de crecimiento y ciclo vital | a.
edad-crecimiento por: 1. anillos anuales en partes duras |
puede ser | paramétricos | a menudo útil | confirmativo | 1. subjetivo |
| 2. fácil de obtener | ||||||
| 2. tasa de diario | elevado | paramétricos | definitivo | puede ser útil | 1. requiere calibración | |
| 2. da tasas de crecimiento a intervalos breves | ||||||
| 3. recuperación de marcas | alto | estamacón | no ordinariamente | confirmativo | 1. de ordinario, sesgos en la liberación y recaptura | |
| 2. el marcado modifica los patrones naturales a breve plazo | ||||||
| b. inicio de la madurez | bajo | paramétricos | útil | confirmativo | 1. puede haber diferencias debido a cambios ambientales | |
| 2. útil para evaluar los cambios | ||||||
| c. fecundidad | moderado | paramétricos | útil | confirmativo | 1. el número real de huevos producidos o incubados es difícil de evaluar | |
| 2. útil para evaluar cambios a menudo menos importante que saber donde son viables los huevos | ||||||
| B2 Estudios de distribución | a. marcado y recaptura 1. marcas y marbetes |
alto | desplazamiento de un punto (a) a otro (b) | puede ser útil | confirmativo | 1. necesita apoyarse con datos de las técnicas del tipo A |
| 2. con peligro de errores de estimación pero útil para evaluar los movimientos | ||||||
| 2. anzuelos | bajo | de un punto (a) a otro (b) | puede ser útil | confirmativo | ||
| B3 Marcas naturales | a. parásitos | moderado | no paramétricos | puede ser útil | confirmativo | 1. pueden perderse o transferirse |
| 2. específicos según la zona o el huesped | ||||||
| b. químico/partes duras | alto | paramétricos | puede ser útil | confirmativo | 1. pueden transferirse o la fuente puede estar distribuida irregularmente, e.g. con las corrientes | |
| 2. mucha información si la fuente tiene localización especie | ||||||
| B4 Recuentos merísticos | a. branquispines o vértebras | moderado | paramétrico | no útil en los túnidos | no | 1. muy variable en algunas especies, debido al ambiente |
| 2. puede indicar algunas diferencias dentro de la especie |
Los recientes progresos en el conocimiento de las exigencias de las formas larvales de teleósteos marinos han proporcionado una base considerable para modificar las premisas usuales sobre el éxito reproductivo de los peces. El concepto de “ventana de supervivencia” en el tiempo y en el espacio, derivado de los estudios de las exigencias energéticas críticas de las formas larvales y de los procesos que las promueven, dan una clave para entender mejor el fenómeno de las cohortes múltiples de reclutas que entran en la mayoría de las pesquerías. Todo esto sugiere que la composición genética compleja puede ser un tema confuso y que se necesitan estudios más definitivos.
En el cuadro 5 se da una sinopsis de los métodos y datos que son útiles para la descriminación de poblaciones.
Los métodos basados en las características de las poblaciones, que se consideran relativamente sensibles, y por tanto definitivos, pero que al mismo tiempo tienen carácter discontinuo y pueden considerarse útiles para la diferenciación y el establecimiento de la identidad son los siguientes: características de las proteínas o las enzimas, comparaciones del ácido deoxiribonucleico (ADN) mitocondrial y reacciones inmunológicas. Los métodos basados en comparaciones cromosomáticas, estructuras de color y estudios numéricos o biométricos están expuestos a interpretaciones subjetivas y son, por tanto, integrativos y menos definitivos, excepto para la diferenciación, donde pueden constituir instrumentos adecuados para hacer inferencias.
Otros métodos útiles para discriminar poblaciones de peces son los que se basan en: estudios de los parámetros de crecimiento y ciclo vital, estudios de distribución mediante marcado y recaptura, marcas naturales, y recuentos merísticos. Estos métodos permiten varios tipos de inferencias, pero por sí solos no pueden considerarse definitivos. Son útiles sobre todo para hacer estimaciones, a menos que se recojan más datos o se conozcan y estudien ampliamente las poblaciones de desovantes y/o los aspectos hereditarios, ya que las conclusiones sobre la estructura de la población derivadas exclusivamente de este tipo de datos son siempre frágiles.
Para el análisis y ordenación de una pesquería normalmente se necesita conocer la abundancia de los recursos. El rendimiento sostenible tiende a estar estrechamente relacionado con la abundancia absoluta, y la mejor manera de monitorizar el impacto que tiene la explotación es a través de los cambios relativos en la abundancia. En la mayoría de los casos, la preparación, calibración y monitoreo de un índice de la abundancia debe recibir una alta prioridad, y en general, al hacer esto, no es conveniente depender de un sólo método debido a las incertidumbres asociadas con todos los métodos de estimación de la abundancia. En el cuadro 6 se hace una comparación y evaluación de diversos métodos para monitorizar la abundancia.
Normalmente, la misma pesquería puede proporcionar información útil para calcular índices de CPUE y para hacer estimaciones mediante análisis de la población virtual. Para obtener índices de CPUE es necesario monitorear el esfuerzo nominal de pesca y las capturas respectivas en un segmento representativo de la pesquería, mientras para el análisis de población virtual se requieren estimaciones de las capturas por edades en todos los segmentos de la pesquería. En algunos casos, las operaciones de la pesca pueden ofrecer además un índice de la abundancia basado en los resultados de los reconocimientos aéreos para localizar las concentraciones de peces. Debido al comportamiento de los peces y a la naturaleza de las operaciones de pesca, la captura por unidad de esfuerzo tiende a subestimar las variaciones de la abundancia y, por tanto, existe el riesgo de que se subestime la magnitud de las posibles disminuciones de la población. Por el análisis de población virtual no se pueden obtener estimados confiables de la abundancia actual, ya que con este método los estimados de la población son reconstruidos a partir de las capturas en los años subsiguientes. Por lo general, el análisis de población virtual da resultados con unos tres años de atraso.
Debido a las deficiencias de estos índices de la abundancia basados en los resultados de la pesca misma, es conveniente utilizar estimàciones independientes, las que por desgracia tienden a ser costosas. Una forma de reducir los costos es recurrir a reconocimientos sinópticos cooperativos, utilizando barcos de la flota pesquera. Este método se ha utilizado durante 20 años en el Perú, donde se conoce con el nombre de reconocimientos “Eureka”. La utilidad y fiabilidad de esos reconocimientos depende de la coordinación, la navegación y la calibración. La información obtenida de la calibración también puede ser útil para documentar los cambios en la capacidad de pesca o en el coeficiente de capturabilidad debidos a la modificación y mejora de los artes, y puede representar una útil adición al monitoreo normal de la CPUE. Con frecuencia, en un reconocimiento cooperativo de tipo “Eureka” se utilizará el equipo acústico de los barcos de pesca. Dado que es posible movilizar a gran número de embarcaciones, con un reconocimiento de ese tipo se puede cubrir toda la zona de distribución de la población. Además, ese método puede dar a los pescadores la sensación de participar en los trabajos de ordenación y puede contribuir a reducir el alejamiento que de ordinario existe entre los pescadores y los responsables de la ordenación.
Los métodos acústicos están aún en vías de desarrollo y todavía no han dado los frutos que de ellos se esperaban en el último decenio. La relación entre la biomasa y las propiedades acústicas es aún incierta en muchos casos y, en general, para la identificación de las especies es necesario un muestreo biológico directo, usualmente con redes. Los reconocimientos acústicos son útiles cuando se usan dentro de un programa de monitoreo a largo plazo, pero pueden conducir a error si se utilizan una sola vez para estimar la biomasa absoluta en un momento dado. Cuando se usan barcos y equipos diferentes, es necesario proceder a una intercalibración para poder comparar los resultados de los reconocimientos, ya que cada uno puede dar mediciones acústicas diferentes.
Para algunas especies pelágicas que se agrupan en la superficie se han utilizado reconocimientos aéreos sistemáticos. Los problemas de estimar la composición por especies y estimar la fracción visible de la población se resuelven de ordinario coordinando estos reconocimientos con un buque de investigación. En general, este último, además de contribuir a la estimación de la abundancia, puede ofrecer muestras biológicas para determinar la composición por edades, identificar la población, etc. En el caso de algunas especies, la pesca experimental constituye de por sl un método útil.
Los reconocimientos de huevos y larvas han sido utilizados ampliamente y, en general, son específicos para cada especie. Los principales problemas con las estimaciones de los censos de larvas (para estimar la biomasa instantánea) son: (1) la dificultad de calibración, (2) la variabilidad de la fecundidad y de la edad de madurez, y (3) la variabilidad de la tasa de mortalidad de las larvas. Los últimos dos factores actuan en forma compensatoria y hacen que las estimaciones de los censos de larvas no sean sensibles a los cambios en la abundancia de la población, ocurriendo así algo similar a lo que ocurre con los resultados de la captura por unidad de esfuerzo, aunque por razones diferentes. Un método introducido recientemente, conocido como “método de producción de huevos” (Parker, 1980; Alheit et al, 1983), resuelve la mayoría de los problemas de los censos de larvas. El método se basa en una estimación de la producción diaria de huevos (evitando el problema de la población instantánea) y recurre al examen histológico de las gónadas de los adultos para determinar la tasa de desove y la fecundidad por unidad de biomasa. Combinando ambas fuentes de información se obtiene una estimación directa de la biomasa desovante, con límites estadísticos de confianza. El método de producción de huevos resulta especialmente adecuado en el caso de desovantes seriales múltiples, cuando es facil muestrear los huevos o larvas y los adultos desovantes. Se ha utilizado una variante de este método para estimar el volumen de las poblaciones locales de arenque del Mar del Norte. En este caso, el arenque desova en una sola puesta y la fecundidad se determina directamente.
También se ha intentado utilizar métodos de marcado y recaptura para estimar la abundancia de peces neríticos, pero en general los resultados no son satisfactorios. Quedan aún muchos problemas por resolver en lo relativo a la mortalidad causada por las marcas, el comportamiento no aleatorio y la cantidad de peces marcados que se requiere para alcanzar una proporción adecuada entre peces marcados y no marcados. La marcación de peces parece ser más útil en los estudios de crecimiento y/o migración.
En algunas áreas, puede ser posible utilizar “índices de predadores” para estimar la abundancia de las presas. Por ejemplo, la producción de guano de las aves marinas de Sudáfrica parece estar relacionada con la biomasa de peces pelágicos que forman cardúmenes en superficie, como la anchoveta y la sardina. Hasta ahora, no se sabe de ningún caso en que los índices de predación sean suficientemente exactos para utilizarlos directamente en la ordenación de pesquerías pero, por otro lado, son un método poco costoso y puede proporcionar información útil sobre abundancia relativa, migraciones y relaciones ecológicas. En general se necesitan más estudios de las especies depredadoras para interpretar los “indices de predadores”, y en particular se necesita conocer más sobre las causas de variabilidad debidas al comportamiento y al medio ambiente.
Tanto la naturaleza de los pronósticos del reclutamiento como su utilidad dependen de los objetivos de ordenación que se persigan (es decir, el carácter de optimalidad que se desee). Cuanto más claramente definidos estén los objetivos, mejor se podrá evaluar la utilidad de un pronóstico del reclutamiento. Los pronósticos del reclutamiento pueden adoptar varias formas, cada una de las cuales entraña costos y niveles de precisión diversos. En cada caso, la forma más adecuada y los costos y precisión asociados dependarán de los objetivos que se persigan con la ordenación. Tres formas útiles de pronóstico son las siguientes:
Forma continua. El reclutamiento se estima como una variable continua, en general con límites estadísticos de confianza. La estimación puede hacerse en forma de un índice (reclutamiento relativo) o, si es posible establecer la proporcionalidad, puede calcularse el reclutamiento absoluto.
| METODO | CONSIDERACIONES BIOLOGICAS | CONSIDERACIONES AMBIENTALES | ESPECIFICIDAD |
|---|---|---|---|
| Captura por unidad de esfuerzo (CPUE) | Sensible al comporta- miento: tamaño de los cardúmenes, distri- bución, disponibilidad, mezcla de especies. Difícil definir el “esfuerzo” | Sensible a influen- cias físicas en la distribución y la disponibilidad | De regular a buena |
| Analisis de cohortes y población virtual | Es necesario disponer de edades exactas, estimación de la morta- lidad natural. Puede ser sensible a estruc- tura de la población, mezclas, mortalidad natural no constante | Cambios en la tasa de mortalidad natural | Buena |
| Reconocimientos cooperativos de tipo “eureka” con barcos de pesca Reconocimientos acústicos | Depende del método de reconocimiento Sensibles al comporta- miento: profundidad, movimientos diurnos, mezcla de especies, densidad de los cardú- menes, evasión | Depende del método de reconocimiento Pueden ser sensibles a la estructura térmica de la colum- na de agua y al estado del mar | Depende del método de reconocimiento De mala a buena (no resuelto) |
| Reconomientos aéreos (o registros de pesca) | Sensibles a: comporta- miento de la parte de la población visible en superficie, migra- ciones diurnas, mezcla de especies | Sensibles a: estruc- tura térmica, bio- luminiscencia, visi- bilidad atmosférica y acuática, estado del mar, fase lunar | De mala a buena |
| Pesca experimental | Evitación al arte de pesca, selectividad | Estructura térmica, estado del mar, bio- luminiscencia, fase lunar | Buena |
| Censos de huevos y larvas | Distribución diurna y evasión, irregularidad en la distribución (en parches), cambios com- pensatorios en los procesos vitales | Temperatura, luz estado del mar | Buena |
| Método de producción de huevos | Estacionalidad, distri- bución irregular, com- portamiento reproduc- tivo y segregación de estadios maduros | Temperatura, estado del mar | Buena, si los huevos son identificables |
| Marcado | Dispersión, mortalidad por marcado, comporta- miento anómalo, efi- ciencia e intensidad de recaptura | Buena | |
| Indices de predación | Sensibles a: comporta- miento, mezcla de presas, especificidad y cambios de la dieta | Influencias ambien- tales en los preda- dores, precipitacio- nes sobre el guano | De mala a buena |
| USO APROPIADO | COSTOS CAPITAL/HUMANO (equipo) | OPORTUNIDAD DE LA INFORMACION | MEJORAS NECESARIAS |
|---|---|---|---|
| Segmento explotado, aspectos económicos; malo para especies que forman cardúmenes fáciles de detectar; mejor para algu- nas especies demersales | Bajo/bajo a medio | Despues de una temporada de pesca | Determinar los sesgos, definición del esfuerzo nominal |
| Para análisis histórico, no actuales. Bueno para calibrar otros índices | (Costo de los datos)/bajo | Con varios años de atraso | Parámetros de entrada (supuesots), es necesario examinar la robustez |
| Reconocimientos programados regularmente, los pescadores participan constructivamente | Bajo/bajo a medio | Sinópticos | Calibración, coordinación navegación |
| Series de reconocimientos a largo plazo: mejor para recursos monoespecíficos y especies pelágicas. Utilidad no comprobada, especialmente en los reconocimientos a breve plazo | Alto/medio | Sinópticos | Calibración, identificación de especies, evitación |
| Evaluación rápida de los peces en superficie | Bajo a medio/ bajo | Sinópticos para una estación de pesca (con los registros de pesca | Fraccción visible, calibra- ción, identifica- ción de especies |
| Usado a menudo junto con otros métodos, permite cono- cer estructura de edad, etc., de recursos inexplotados | Alto/medio | Sinópticos | Diseño del muestreo, evita- ción del arte de pesca, selecti vidad |
| Indice de la biomasa de desovantes | Alto/alto | 6 o más meses | Calibración, automatización, determinar los sesgos |
| Estimación de la biomasa total o de desovantes, ictioplancton fácil de muestrear y abundante, bueno también para zonas localizadas de desove No recomendado para estima- ción de la abundancia | Alto/alto Bajo a medio/ alto | 4 o más meses De una temporada de pesca al ciclo vital de los peces | Conocimiento del comporta- miento de desove dispersión y predación Reducir y estimar la mortalidad por marcado, entender el comportamiento |
| Indice de abundancia suplementario y para antes de empezar la pesquería | Bajo/bajo | Variable | Entender el comportamiento, se necesitan más ejemplos |
Forma discontinua. Se puede pronosticar que el reclutamiento estará comprendido en una de varias categorías. En el caso de una estimación relativa, se puede decir que el reclutamiento será “superior”, “aproximadamente igual”, o “inferior a” lo normal; y, cuando se disponga de una estimación absoluta, se puede decir que será “mayor que R2”, “entre R1 y R2”, o “menor que R1”. Se puede calcular la probabilidad de incurrir en error al establecer la categoría, probabilidad que aumentará a medida que las cifras se acerquen al punto divisorio entre categorías. Por otro lado, el esfuerzo de muestreo y los costos correspondientes pueden resultar mucho menores que en el caso de los pronósticos de forma continua.
Forma decisoria. Se trata de una variante de la forma discontinua y puede ser la mejor solución cuando el objetivo sea impedir que se produzca una catástrofe. Se clasificará el reclutamiento considerándolo superior o inferior a un valor crítico determinado, y podrá ser tratado estadisticamente mediante una prueba de verificación de las hipótesis. Sin entrar muy a fondo en cuestiones de teoría estadística, es importante examinar los tipos de error que pueden producirse en este caso. Considérense las siguientes posibilidades:
| Decisión | |||
| R es inferior al nivel ciítico | Nivel satisfactorio de R | ||
| Situación real | R es inferior al nivel ciítico | correcto | error (error de primer tipo) |
| Nivel satisfactorio de R | error (error de segundo tipo) | correcto | |
Si consideramos como hipótesis nula que “R es inferior al nivel crítico”, podemos cometer dos tipos de error. En un caso, podemos dejar que la pesquería prosiga sin tomar las medidas preventivas necesarias (error de primer tipo), y en el otro, podemos tomar medidas preventivas cuando de hecho no son necesarias (error de segundo tipo). Si a igual intensidad de muestreo (es decir, precisión) elegimos criterios que hagan menos probable un tipo de error, aumentará la probabilidad de cometer un error del otro tipo. El responsable de la ordenación debe decidir cuál es la gravedad relativa de ambos tipos de errores; de todas maneras, si el objetivo es prevenir catástrofes, es claro que conviene que la probabilidad de cometer un error de primer tipo sea pequeña.
Si bien es deseable llegar a un pronóstico del reclutamiento que se base sólo en variables ambientales causales o correlacionadas, son pocas (o ninguna) las relaciones empíricas conocidas suficientemente exactas como para ser utilizadas con este fin. En la actualidad, lo que ofrece mejores posibilidades para un pronóstico útil del reclutamiento es el muestreo directo o indirecto de las formas juveniles (pre-reclutas). Pueden utilizarse redes, pero los peces jóvenes son muy evasivos y es necesario majorar los artes y técnicas de muestreo. Entre los métodos de muestreo indirectos pueden citarse los reconocimientos acústicos, que pueden basarse en la resonancia de la vejiga natoria para identificar la especie y determinar la talla. En algunos casos, los reconocimientos de pre-reclutas pueden concentrarse en zonas indicadoras, donde se sabe que esos peces se concentran.
En general, cuanto menor es la edad a la que un pez entra en la pesquería, más difícil es obtener un pronóstico útil y oportuno del reclutamiento. Ironicamente, es precisamente en el caso de las especies de vida corta y de reclutamiento temprano donde más necesarios son los pronósticos de reclutamiento.
Para obtener estimados más actualizados, se pueden combinar los pronósticos del reclutamiento con indicaciones de la intensidad del reclutamiento (por ejemplo, tasas de capturas de individuos juveniles) obtenidas en el periodo inicial de la pesquería. De esa manera se consigue una mayor precisión a costa de un retraso en el tiempo; sin embargo, la ordenación puede incorporar fácilmente un mecanismo de respuesta progresiva en dos fases, a medida que la estimación del reclutamiento gana en precisión.
Si para los recursos de que se trate se dispone de una relación entre población y reclutamiento (incluida la amplitud de la variabilidad natural), los métodos estadísticos bayesianos pueden permitir mayor precisión y menores riesgos con poco costo. La relación población-reclutamiento ofrece una distribución de probabilidad a priori para el reclutamiento y se puede considerar que el reclutamiento pronosticado es una muestra de esa distribución a priori. Otra característica interesante de este método es que puede introducirse en la estimación del reclutamiento una función de riesgo (que defina la gravedad de los distintos errores), con lo que es posible incorporar explícitamente la sensibilidad a fallas catastróficas del reclutamiento (u otras condiciones).
La intensidad de reclutamiento muestra a menudo una correlación serial positiva, lo que quiere decir que reclutamientos sucesivos tienden a ser en cierta formas similares. Ello se debe a la relación población desovante-reclutamiento (dado que la misma población desovante muestra una correlación serial, debido a la persistencia de las clases anuales) y a la tendencia de las condiciones ambientales a perdurar por más de un año. La existencia de una elevada correlación serial puede servir como argumento para que se dé una respuesta desproporcionada ante un reclutamiento de intensidad desusual, ya que es probable que al año siguiente se produzcan condiciones similares.
En los últimos años se ha insistido mucho en la necesidad de abordar el análisis y ordenación de las pesquerías desde el punto de vista del “ecosistema” y con un enfoque “multiespecífico”. Aunque el valor teórico de esos enfoques es evidente, por lo general los conocimientos prácticos que se tienen son insuficientes como para poder hacer esto satisfactoriamente. En realidad, nuestros conocimientos de la dinámica de una sóla especie son apenas insuficientes como para abordar la ordenación de una pesquería monoespecífica y son totalmente inadecuados como base para modelos multiespecíficos.
Por ejemplo, especies de peces pequeños que se encuentran en la base de la cadena alimenticia, como la anchoveta, forman parte de la dieta de peces mayores y de aves y mamíferos marinos. Para una ordenación multiespecífica sería preciso tener en cuenta el valor trófico de las anchovetas no explotadas y el valor de los peces explotados. Por otro lado, dado que es planctívora, es indudable que la anchoveta consume huevos y larvas de peces depredadores mayores y, desde ese punto de vista, puede tener un valor negativo. El equilibrio entre esos aspectos podría estudiarse preparando modelos multiespecíficos, pero la información necesaria para aplicar un modelo de este tipo exigiría un costoso programa de investigaciones que sea independiente de la pesquería.
Los ecosistemas neríticos son muy variados, según la latitud y el hábitat. En los cuadros 7(a), (b) y (c), se examina la relación entre los hábitat neríticos y sus pesquerías en zonas tropicales, subtropicales y templadas.
Como consecuencia de profundos cambios en el ambiente físico y en la abundancia de peces neríticos, las principales pesquerías neríticas se desarrollan en un ambiente dinámico. Entender y predecir algunas de las respuestas de los peces a las variaciones del medio físico y las respuestas de los ecosistemas a los cambios en la abundancia de los peces puede ser útil a la hora de hacer recomendaciones sobre cómo ordenar la pesquería.
En primer lugar, los cambios del medio físico, como “El Niño”, se caracterizan por desplazamientos importantes de las masas de agua. Los límites entre las masas de agua cambian de posición geográfica. Muchos peces neríticos, como la anchoveta, la sardina, la caballa, etc., son pelágicos y responden a esos cambios como organismos nectónicos volitivos, redistribuyéndose geográficamente en relación con la temperatura y otras características del agua. Los peces tienen preferencias bien definidas, que guían esos movimientos en los gradientes oceánicos. Las estimaciones hechas en laboratorio sobre las preferencias en materia de temperatura y otras características, junto con información sinóptica sobre la oceanografía física, pueden “ser útiles para explicar los desplazamientos norte-sur de peces que viven en aguas más cálidas o más frías, así como las restricciones de la superficie total de los ambientes preferidos y la sustitución de una especie por otra en un área geográfica determinada. Además, si el alimento es abundante, los peces alcanzan de ordinario las tasas más elevadas de crecimiento cuando la temperatura del agua es la preferida. Hay algunas evidencias de que, si los alimentos son menos abundantes, los peces pueden desplazarse, y de hecho se desplazan hacia temperaturas más frías para maximizar su crecimiento. Crowder y Magnuson (1983) han preparado una hipótesis sobre la distribución de los peces en relación con los alimentos y la temperatura, en un ambiente espacialmente heterogéneo.
| ESTUARINOS VARIOS | FONDOS DUROS, ARRECIFES | FONDOS BLANDOS, DELTAS FLUVIALES | ESPECIES PELAGICAS EN AFLORAMIENTOS COSTEROS | PREDADORES ASOCIADOS A ARRECIFES O ISLAS | |
|---|---|---|---|---|---|
| Base de recursos | Camarones, peces marinos juveniles, adultos reproduc- tores, especies de aguas dulces | Especies herbívoras y piscívoras | Numerosas especies de todos los niveles tróficos | De una a tres especies prin- pales y preda- dores | Túnidos, agujas, jureles grandes |
| Principales grupos taxonó- micos explotados | Peneidos, Mugílidos, Perciformes Selacios | Perciformes | Perciformes | Clupeidos, Escóm- bridos, Carángidos | Thunnus, Katsuwonus, Istioforidade, Perciformes Carángidos, Serránidos |
| Densidad de población en estado virgen | Variable, media, estacional | Alta, aparentemente estable | De media a alta | Muy variable: de baja a alta | De baja a media |
| Principales artes empleados | De cerco y artesanales | Trampas, Iíneas de mano, algunos artes destructivos | Arrastre demersal y artesanales | Cerco | Curricán, Caña y línea, líneas de mano y artesanales |
| Profundidad de los estratos explotados | De la superficie al fondo | Hasta unos 100 m | De la superficie al fondo ⋍ 100 m | Capa de mezcla | De la superficie al fondo, según las especies |
| Pesquerías artesanales importantes | Si | Si | Si | Algunas | Si |
| Uso de las capturas | Alimentación y exportación | Alimentación, fotografía | Alimentación y exportación | Alimentación y fabricación de harina | Alimentación y exportación |
| Calidad y valor de los productos | Por lo general bueno y medio | Por lo general muy alta | Varía según el tratamiento | Por lo general pobre y bajo | Altos, pero con proble- mas de mercadeo |
| Patrón de abun- dancia de las clases anuales | Relativamente estables | Posibilidad de inestabilidad a nivel de especie, estables a nivel de comunidad | Poca información, pero parecen ser variables en pequeña escala | Inestables, pero pueden ser altas por periodos largos | Reclutamiento continuo, con ciertos cambios en la disponibilidad estacional |
| Conocimiento de la biología de los recursos | Bueno | Como recursos los peces de arrecife no se conocen bien | Pobre | bueno | Variable |
| Explotación por parte de | Pescadores locales | Pescadores locales, turismo | Flotas locales y algu- nas de gran altura | Flotas locales y a menudo de gran altura | Sobre todo pescadores locales |
| Perturbaciones principales | Cambios ambienta- les causados por el hombre y de origen biológico-oceánico | Destrucciones de arrecifes, algunos predadores de arrecife (por ejemplo, la estrella de mar Acanthaster planci | Inducidas por la pesquería y bioló- gicas localizadas | Tendencias climá- ticas, eventos meteorológicos | No se conoce |
| Posibilidades de expansión | Pocas | Pocas | De ordinario ninguna pero en algunas áreas uede haberlas | Varían según el flujo de los recursos | No grandes |
| Limitaciones estacionales | Inundaciones | Pocas | Monzones o tormentas tropicales | Vientos fuertes o tormentas tropicales | Monzones o tormentas tropicales |
| COSTERO/LAGUNAR | AGREGACIONES EN LA PLATAFORMA E ISLAS | ESPECIES PELAGICAS | CAPAS SUB- SUPERFICIALES | |
|---|---|---|---|---|
| Base de recursos | Especies pelágicas pequeñas, predadores pequeñ, peces marinos juveniles | Jureles, bonitos, túnidos, calamares, estornino | Anchovetas, sardinas, caballas, jureles, bonito | Jureles adultos, merluza, galli- neta, bonito |
| Principales grupos taxonómicos explotados | Engraulis, Opistonema, Sphyraenidae, Caranx, Serranidae | Seriola, Sarda, Thunnus, Loligo, Scomberomorus, Acanthocybium | Sardina, Sardinella, Engraulis, Sardinops, Scomber, Trachurus, Sarda | Trachurus, Merluccius, Sebastes, etc. |
| Densidad de población en estado virgen | De variable a alta | Estacionalmente alta | De variable a alta | Alta |
| Principales artes empleados | Cerco, artesanales | Curricanes, líneas de mano, cerco | Cerco | Arrastre |
| Profundidad de los estratos explotados | De la superficie al fondo | Superficie y fondo | Superficie | En aguas medias a profundas, es decir: 25–1 000 m |
| Pesquerías artesanales | Si | Si | por lo general | No |
| Uso de las capturas | Alimentación | Alimentación y exportación | Alimentación, fabri- cación de harina, | Exportación exportación |
| Calidad y valor de los productos | Casi siempre baja | Alta | Mejormente baja a media | Media |
| Patrón de abundancia de las clases anuales | Muy variables | Variables | Erupciones y fallas | Erupciones y fallas |
| Conocimiento de la bio- logía de los recursos | Pobre | pobre a razonable | En algunos casos muy bueno | Razonable |
| Explotación por parte de | Pescadores locales | Pescadores locales y extranjeros | Pescadores locales y extranjeros en algunas áreas | A menudo pesca- dores extranjeros |
| Perturbaciones principales | Cambios del ecosistema cíclicos o por épocas (por ejemplo, aves y predadores) | Ciclos poblacionales de largo período, debidos a eventos climáticos oceánicos | Tendencias esporádicas y por épocas modifican aparentemente el sistema (por ejemplo, ciclos de El Niño y del Sahel) | Tendencias esporádicas y por épocas modifican aparentemente el sistema (por ejemplo, ciclos de El Niño y del Sahel) |
| Posibilidades de expansión | Ninguna | En algunas áreas, muchas | Por lo general, ninguna | En algunas áreas, muchas |
| Limitaciones estacionales | Cambios cíclicos en las influencias tropicales/ templadas | Migración y distribución asociadas con cambios en la ubicación de la zona de transición | Pocas | pocas |
| ESPECIES PELAGICAS | CAPAS SUB-SUPERFICIALES | ESPECIES ASOCIADAS A ISLAS O MESETAS SUBMARINAS | |
|---|---|---|---|
| Base de recursos | Capelán, arenque, espadín, caballa, sardina, osmeroides | Carbonero, bacalao y afines, peces planos, merluza, gallineta, jureles grandes | Bacalao, merluza, arenque, carbonero, peces planos |
| Principales grupos taconómicos explotados | clupes, Sprattus, Scomber, Sardinops, Mallotus | Theragra, Gadidos, Merluccius, Sebastes, Trachurus, Pleuronectidos | Gádidos, merluccius, Clupes Theragra, Pleuronectidos |
| Densidad de la población | De variable a alta | Generalmente alta | De variable a alta |
| Principales artes empleados | Cerco | Arrastre, enmalle, palangre | Arrastre, palangre, cerco |
| Profundidad de los estratos explotados | Capa de mezcla | Medios a profundos | De la superficie al fondo |
| Pesquerías artesanales importantes | Ninguna | Ninguna | Ningna |
| Uso de las capturas | Alimentación y exportación | Alimentación y exportación | Alimentación y exportación |
| Calidad y valor de los productos | Media-alta | Media-alta | Media-alta |
| Patrones de abundancia de las clases anuales | Fallas frecuentes | Fallas ocasionales | Fallas frecuentes |
| Conocimiento de la biología de los recursos | Bueno | De ordinario bueno | De ordinario bueno |
| Explotación por | Pescadores locales y extranjeros | Pescadores locales y extranjeros | Pescadores locales y extranjeros |
| Perturbaciones principales | Variaciones climáticas | Tendencias climáticas y en la formación de los hielos | tendencias climáticas |
| Posibilidades de expansión | Varían según la abundancia de los recursos | Ninguna en el hemisferio septentrional | Ninguna de ordinario |
| Limitaciones estacionales | Ciclo migratorio y reproductivo | Migraciones y tormentas | migraciones y tormentas |
En segundo lugar, los cambios importantes en la abundancia de algunos peces neríticos, como anchoveta y sardina, repercuten inevitablemente en su ecosistema. La forma más directa en que los peces, en cuanto consumidores, modifican su ambiente, especialmente cuando la densidad de población es alta, es a través del consumo de los organismos que les sirven de presa. Los peces consumidores pueden reducir la abundancia de los que les sirven de presa y si, por ejemplo, tienden a elegir zooplancton de mayor tamaño, pueden modificar la distribución por tallas de sus presas. Se ha observado que, cuando los organismos planctónicos de mayor tamaño se hacen escasos, los peces crecen rápidamente hasta alcanzar un tamaño determinado y luego se produce una notable disminución de la tasa de crecimiento. Si el crecimiento se detiene en esa talla, es porque ya no encuentran partículas de mayor tamaño para alimentarse con menor costo energético. Por tanto, el mayor costo energético que supone depredar organismos menores reduce la ingesta neta de energía y el crecimiento resulta frenado o se detiene. Los cambios así inducidos en el plancton pueden modificar la gama de tamaños de las presas haciéndola menos adecuada para otros zooplanctívoros y determinar, de esa manera, una disminución de la abundancia de otras especies. Cuando disminuye la abundancia de los zooplanctívoros predominates, la presión que ejercen sobre el ambiente se elimina y otras especies pueden prosperar mejor. También, si la distribución por tamaños de las partículas de zooplancton se modifica, la depredación de huevos puede ser un factor más crítico durante los períodos de elevada abundancia de peces.
Otros cambios determinados por los peces en cuanto consumidores son la modificación de las tasas de consumo de las especies depredadas, que se alimentan de organismos situados más abajo en la cadena trófica, y la alteración de los patrones de remineralización de los nutrientes (es decir, los compuestos nitrogenados pueden concentrarse o mantenerse en aguas próximas a la superficie). Al mismo tiempo, la producción primaria puede resultar modificada como consecuencia de las concentraciones de compuestos nitrógenados inducidas por los peces y de la modificación de la biomasa fitoplanctónica a causa de su consumo por los peces o por los organismos de que éstos se alimentan. Estos efectos se han observado y documentado en ecosistemas pelágicos lacustres y algunos se han observado también en ambientes marinos (Stroud y Clepper, 1979).
Durante mucho tiempo se ha especulado sobre una posible competencia alimentaria entre la sardina y la anchoveta adulta como medio para explicar la sustitución recíproca de estas dos especies, pero hasta la fecha no se ha dado prueba alguna que demuestre esa competencia alimentaria entre ambas especies. Una de las razones de ello es la evidente dificultad de cuantificar los hábitos alimentarios de muchas especies de peces planctívoros, debido a la gran diversidad taxonómica de los alimentos que consumen. Además, tanto la sardina como la anchoveta pueden modificar su forma de alimentación según el tamaño de las presas, recurriendo al filtrado o al mordisco. En segundo lugar, aún en el caso de que se mostrara que la gama de alimentos de ambos géneros coincide en gran medida, ello no significaría que existe una competencia alimentaria. Si la capacidad de carga del ambiente es elevada (en este caso, si el plancton es muy abundante), las dos especies podrían ocupar exactamente el mismo nicho alimentario sin competir entre sí. Sería un caso de “coexistencia alimentaria”, en el sentido en que lo entiende Jones (1982). De todas maneras, aunque nunca se han dado pruebas sólidas de que exista competencia alimentaria entre la sardina y la anchoveta, algunos científicos ignoran la debilidad de los datos disponibles y señalan que la competencia alimentaria es uno de los mecanismos que determinan la sustitución entre estas dos especies. Es necesario estudiar atentamente los hábitos de ambas especies en un mismo ambiente y proceder a un análisis paralelo de sus fuentes de alimentación y de los aspectos energéticos, para aclarar la relación entre ambas especies y poner fin a la larga serie de especulaciones sobre este punto.
En el mismo estudio podrían examinarse también los efectos de la predación y el canibalismo en los huevos y las larvas. Este fenómeno ha sido ya documentado (por ejemplo, Santander et al., 1983), y su influencia en las relaciones entre la anchoveta y la sardina es semejante a la que se supone tendría la competencia, aunque el mecanismo es diferente.
Conviene tener presente que un modelo no es más que la formulación racional de las ideas de quien lo prepara. El modelo, una vez aplicado, muestra si la información disponible es coherente con dichas ideas. Cuando lo es, los resultados obtenidos con el modelo indican a menudo qué nueva información podría servir para someter esas ideas o otra verificación más rigurosa.
Los modelos de interacción biológica pueden ser totalmente empíricos (por ejemplo, basados en expresiones polinómicas) o basarse en modelos pesqueros más familiares, como el modelo de producción de Schaefer (Lotka-Volterra) o el más detallado de Beverton y Holt. En esta última categoría figuran los modelos de análisis de cohortes múltiespecíficos que se utilizan para analizar las capturas mixtas en el Mar del Norte. No parece que el uso de modelos de producción permita obtener mucha información biológica, aunque sus conceptos y supuestos pueden servir de base para algunos razonamientos bien fundados.
En general, los modelos multiespecíficos tienden a ser sencillos y poco realistas o, en cambio, difíciles de manejar y poco atractivos para quien esté iniciándose en el arte de preparar modelos. Una excepción interesante es quizás el enfoque de Larkin y Gazey (1982) de aplicar al desarrollo de las pesquerías del Golfo de Tailandia un modelo sencillo de la biomasa, que puede procesarse en una pequeña computadora personal y exige relativamente poca experiencia en materia de programación. El modelo representa un sistema cerrado, pero debería ser fácil de adaptar a un sistema abierto, como una zona de afloramientos costeros, añadiendo factores de migración.
Podría ser interesante formular el sistema peruano, tal como lo ha concebido Walsh(1981), utilizando el enfoque de Larkin y Gazey. Walsh ha añadido a los conocimientos comunes sobre la historia de la anchoveta peruana la observación de que tras el colapso de la población de anchoveta se produjo un aumento de la sedimentación de materia orgánica, elemento que puede ofrecer la información necesaria para comenzar a entender las interacciones dentro de un sistema tan variable como es ése.
Un sistema como el que se encuentra frente al Perú representa un reto para los especialistas en modelos del ecosistema. Cuanto mayores sean las perturbaciones que han de describirse, mejor ha de ser el modelo. Preparar un modelo de un sistema en estado estable no constituye un problema, pero probablemente ese modelo se revelará poco satisfactorio cuando se produzca una perturbación y no es fácil que incluya los mecanismos necesarios para responder en forma realista a una perturbación que no se haya producido antes.
Como en la mayoría de los campos de las ciencias naturales, las ciencias pesqueras se ven más limitadas por la creatividad e ingenio del hombre que por la complejidad del equipo necesario. En la mayoría de los casos, para una buena investigación pesquera, bastan pocos elementos de equipo (aunque para determinadas investigaciones puede ser necesario equipo complejo). El uso de computadoras está resultando cada vez más esencial para procesar y analizar grandes colecciones de datos y preparar modelos complicados. Pero raras veces es necesaria una computadora grande para ello; pequeñas computadoras, poco costosas, con medios adecuados para el almacenamiento de datos, son a menudo suficientes y no requieren una costosa manutención.
A menudo resulta desaconsejable, desde el punto de vista económico, adquirir un buque de investigación grande. A veces se requieren barcos grandes porque las operaciones requieren una amplia autonomía o han de hacerse con mal tiempo, pero en otros casos la adquisición de buques grandes se debe solamente a razones de prestigio. En cualquier caso, es preciso sopesar las razones en pro de la adquisición de un buque grande (a menudo a un costo nominal) contra los costos reales que impondrá su operación. La mayoría de las veces, los costos operativos son el factor decisivo para el éxito de un programa de investigación en el mar. Al mismo tiempo, hay que tener cuidado en que el equipo del barco (máquinas, hidráulica, electrónica, etc.) sea tal que resulte fácil encontrar repuestos y proceder al servicio en el país en que vaya a utilizarse (puede ser distinto del país de construcción); de no hacerse así, las reparaciones pueden resultar muy costosas y llevar muchos meses, con las consiguientes pérdidas de fondos y tiempo.
A menudo se desperdicia mucho esfuerzo y dinero porque los artes adquiridos para el buque de investigación no son adecuados, especialmente en el caso de las redes de cerco y arrastre. Hay que tener en cuenta el tipo de peces y la cantidad necesaria así como la capacidad del barco y su tripulación. Si no es fácil disponer del asesoramiento de expertos, es prudente dirigirse a un consultor.
Los científicos pesqueros tienen la responsabilidad profesional y ética de adquirir, mejorar y evaluar sus “herramientas” (que incluyen técnicas de análisis, modelos, conceptos unificadores, etc.). De esas tres actividades, la más difícil y necesaria es la evaluación. Un breve estudio de los diversos sesgos que se encuentran en la información pesquera y los análisis de las pesquerías Ileva a una conclusión muy importante e inquietante: la preponderancia de errores en las ciencias pesqueras conduce a la sobreexplotación de los recursos pesqueros.
Los sesgos indicados no parecen proceder de la base teórica de la ciencia pesquera. Se deben, más bien, a los métodos de aplicación y medición, en los que influyen enormemente las circunstancias. Los ejemplos siguientes podrán ser útiles para poner en claro la naturaleza del problema.
Si el método utilizado para estimar la abundancia no es sensible a los cambios en la abundancia real, el análisis de la pesquería puede subestimar el grado real de impacto que tiene en un recurso. En consecuencia, los modelos correspondientes de rendimiento tenderán a sobreestimar la intensidad óptima de explotación.
Captura por unidad de esfuerzo. Este índice tan popular no es a menudo sensible a los cambios en la abundancia real, especialmente en las pesquerías pelágicas de superficie (por ejemplo clupeidos), en las que tanto la distribución de la pesca como la de los peces son no aleatorias. Cuando los recursos disminuyen, las dimensiones de los cardúmenes pueden permanecer invariadas y el área de distribución de los recursos puede contraerse, mientras la abundancia en el centro permanece relativamente constante. Los pescadores responden a una disminución de la abundancia adaptando sus artes y eliminando los barcos menos eficientes, con objeto de mantener elevada la captura por unidad de esfuerzo nominal. En las pesquerías multiespecíficas, el esfuerzo nominal dirigido a una especie determinada se define a menudo como el esfuerzo que ha resultado en la captura de esa especie, lo que hace que la CPUE resulte insensible a los cambios en la abundancia real.
Pesquerías en desarrollo. A menudo, cuando se inicia una pesquería, es inevitable un período de aprendizaje. La pesquería inicial, con una abundancia elevada y una explotación ineficiente, se convertirá con el tiempo en una pesquería donde la abundancia será menor pero la eficiencia habrá aumentado. Durante esa transición, la captura por unidad de esfuerzo nominal no disminuirá y ello dará la impresión de que el recurso tiene gran elasticidad o capacidad de recuperación. Además, a medida que se extiende el radio geográfico de una pesquería en desarrollo, puede suceder que se vayan explotando progresivamente segmentos vírgenes de la población durante un período prolongado.
Reconocimientos de ictioplancton. Los cambios compensatorios en la fecundidad, la edad de madurez y la mortalidad de huevos o larvas harán que en estos reconocimientos se subestimen los cambios en la abundancia de la población.
Reconocimientos acústicos. Cuando la densidad es elevada puede resultar imposible detectar cuantitativamente los peces que se hallan en el centro de los cardúmenes, mientras con densidades menores la detección es mejor.
Capturas. La falta de comunicación de las capturas puede conducir inadvertidamente a una explotación excesiva. La aplicación de cuotas de captura para proteger los recursos puede dar lugar a que aumenten los casos de capturas no declaradas, no comunicación de datos o identificación equivocada de especies.
Mejora progresiva. La mejora progresiva de los trabajos de monitoreo puede dar una falsa impresión de estabilidad, mientras de hecho las capturas están disminuyendo.
Regresión funcional. El uso de la regresión estándar “Y sobre X”, en vez de una regresión funcional (que permite reconocer los errores de observación en las variables independientes) se traducirá en una subestimación del valor absoluto de la pendiente de la regresión. En los modelos de producción, ello conduce a menudo a una sobreestimación del rendimiento máximo sostenible y del esfuerzo óptimo. En las relaciones población desovante-reclutamiento esto puede dar la impresión errónea de que el reclutamiento es independiente del tamaño de la población. No se dispone aún de regresiones funcionales para aplicación no lineal y, por tanto, en la estimación de modelos de regresión no lineal ese sesgo es actualmente inevitable.
Variabilidad aleatoria. La variabilidad aleatoria y la correlación serial del reclutamiento son factores que hacen que los redimientos obtenibles sean inferiores al rendimiento máximo sostenible determinístico. A pesar de ello, es este último rendimiento el que se utiliza a menudo como criterio de una pesquería plenamente desarrollada.
Rendimiento potencial. La conocida estimación del rendimiento potencial propuesta por Alverson-Pereyra-Gulland (Y = 0,5 MB∞) tiende a sobreestimar los rendimientos sostenibles que pueden conseguirse. Con frecuencia, las pesquerías se desarrollan inicialmente con un abundancia anormalmente alta de recursos, lo que da una sobreestimación de la abundancia virgen B∞ . La fórmula entraña además que el reclutamiento al nivel de rendimiento máximo sostenible es igual al reclutamiento virgen, cosa que raramente sucede. En caso de recursos muy fluctuantes, el redimiento potencial puede no resultar apropiado para los niveles más bajos de los ciclos de abundancia. Lamentablemente, se suele seguir usando la estimación del rendimiento potencial mucho tiempo después de disponer de información mejorada sobre la pesquería.
Formulación de modelos. La mayor parte de los modelos pesqueros se formulan en términos deterministas y tienden a un equilibrio, suavizando las transiciones entre los distintos estados. Si lo que existe en realidad es un desequilibrio sostenido, con catástrofes o estados estables múltiples, esos modelos sobreestimarán la estabilidad de los recursos. Puede ser atractivo suponer una productividad multiespecífica constante, pero la experiencia muestra que la productividad total puede ser baja por varios años durante las transiciones (si es que este concepto tiene validez alguna).
Lo que los científicos y administradores pesqueros desean, al igual que los pescadores, es que los recursos sean saludables y productivos. Por ello, puede haber una tendencia inconsciente a sobreestimar la productividad o la abundancia (cosa que se ha observado también en otras ciencias: véase Gould, 1978). Así pues, se suele tolerar, por ejemplo, la presunta presencia de un sesgo hacia la sobreestimación de la abundancia real, mientras ante un presunto sesgo hacia la subestimación se suele adoptar una actitud más crítica.
Esta peligrosa y extendida tendencia al sesgo que se observa en la ciencia pesquera puede evitarse en gran medida, pero el remedio no es fácil:
| - | Los científicos y administradores han de estar conscientes de la tendencia inadvertida a la sobreexplotación y de las varias maneras en que ello puede suceder. |
| - | Los métodos y análisis han de someterse periódicamente a revisión crítica, para detectar ese tipo de error. |
| - | El uso de métodos adecuados es importante. Ello entraña a menudo análisis más largos y detallados. Los métodos abreviados son a menudo peligrosos si los resultados se consideran definitivos. |
| - | Es preciso preparar métodos y modelos más sólidos. Al mismo tiempo, hay que estudiar y entender mejor los sesgos inherentes en los métodos existentes. |
En la ciencia pesquera, los criterios de trabajo han de ser: conocimiento, responsabilidad, oportunidad, aplicabilidad y perspectiva (tener en cuenta el contexto).
Los subsistemas físico, biológico y humano de las pesquerías están sometidos a variabilidad en todas las escalas de tiempo y sus interacciones son importantes para la ordenación.
Los cambios en la abundancia pueden deberse a dinámica intraespecífica, competencia, predación, pesca o causas ambientales. Los datos sobre la variabilidad de la pesquería y los recursos permiten elucidar esas causas.
Los distintos síntomas de cambios adversos en la situación de los recursos pueden agruparse en tres categorías principales: disminución de la abundancia, disminución del reclutamiento, e intensidad de pesca relativamente alta. Esos síntomas han de considerarse en el contexto de la pesquería, para poder establecer niveles de referencia.
Se han examinado varios aspectos del monitoreo de operaciones pesqueras y del muestreo biológico. Es necesario llevar un diario o preparar una relación histórica, además de registrar datos como los de captura y esfuerzo. En el muestreo ha de tenerse en cuenta la presencia de múltiples especies en las capturas.
Un amplio estudio comparativo de la captura por unidad de esfuerzo como índice de la abundancia sería útil para evaluar el valor de ese índice y ayudar a definir cuáles son las mejores medidas disponibles del esfuerzo de pesca nominal.
Muchos procedimientos estándar (claves de edad-talla, criterios de madurez, determinación de la fecundidad) deberían revisarse a la luz de los progresos tecnológicos y científicos.
La identificación de poblaciones sigue siendo una tarea importante, pero difícil. Se siguen preparando nuevos métodos, pero muchos exigen el uso de tecnología compleja.
Se han comparado varios métodos de estimación de la abundancia, sin que ninguno parezca resultar superior. Es aconsejable determinar la abundancia utilizando más de un método.
Los pronósticos del reclutamiento se hallan aún en una fase primitiva de desarrollo. La naturaleza de los pronósticos depende de la forma en que vayan a utilizarse en la ordenación pesquera; para la mayoría de los objetivos, podrá bastar una evaluación cualitativa, determinando si el reclutamiento es “superior, aproximadamente igual o inferior a lo normal”.
Las tendencias recientes en pro de una ordenación multiespecífica y/o del ecosistema no son apoyadas por los conocimientos actuales. Es precisamente en este campo en el que se presentan los retos más importantes para la investigación y la preparación de modelos.
Raras veces los progresos de la ciencias pesqueras se ven limitados por falta de equipo o de instalaciones. Por lo general, no es aconsejable la adquisición de grandes buques de investigación, debido a los costos prohibitivos de operación.
Los científicos pesqueros han de evaluar los sesgos presentes en sus métodos y análisis. Un breve examen sugiere que la preponderancia de errores en la ciencia pesquera conduce a la sobreexplotación.
Alheit, J. et al., 1983 Batch fecundity and spawning frequency of various anchovy (genus: Engraulis) populations from upwelling areas and their use for spawning biomass estimates. FAO Fish.Rep./FAO Inf.Pesca, (291)Vol.3:977–86
Beverton, R., 1983 Science and decision-making in fisheries regulation. FAO Fish.Rep./FAO Inf.Pesca, (291)Vol.3:919–38
Booke, H.E., 1981 The conundrum of the stock concept: are nature and nurture definable in fishery science? Can.J.Fish.Aquat.Sci., 38(12):1479–80
Crowder, L. y J. Magnuson, 1983 Cost benefit analysis of temperature and food resource use: a synthesis with examples from the fishes. In Behavioral energetics, editado por W. Aspey y S. Lustick. Columbus, Ohio, Ohio State University Press, pp.189–221
Gould, S., 1978 Morton's ranking of races by cranial capacity. Science, Wash., 200:503–9
Jones, R., 1982 Ecosystems, food chains and fish yields. ICLARM Conf.Proc., 9:195–239
Kimura, D., 1977 Statistical assessment of the age-length key. J.Fish.Res.Board Can., 34(3):317–24
Larkin, P.A. y W. Gazey, 1982 Applications of ecological simulation models to management of tropical multispecies fisheries. ICLARM Conf.Proc., 9:123–40
Paloheimo, J. y L. Dickie, 1964 Abundance and fishing success. Rapp.P.-V.Réun.CIEM, 155:152–63
Parker, K.R., 1980 A direct method for estimating northern anchovy, Engraulis mordax, spawning Biomass. Fish.Bull.NOAA/NMFS, 78:541–4
Santander, H. et al., 1983 Egg mortality of the Peruvian anchovy (Engraulis ringens) caused by cannibalism and predation by sardines (Sardinops sagax). FAO Fish.Rep./FAO Inf.Pesca, (291) Vol.3:1011–26
Sharp, G.D., 1983 Tuna fisheries, elusive stock boundaries and illusory stock concepts. Collect.Vol.Sci. Pap.ICCAT/Recl.Doc.Sci.CICTA/Colecc.Doc.Cient.CICAA, (3):812–29
Stroud, R. y H. Clepper (eds), 1979 Predator prey systems in fishery management. Washington, D.C., Sport Fishing Institute, 504 p.
Walsh, J., 1981 A carbon budget for overfishing off Peru. Nature, Lond., 290:300–4
![]()
![]()
![]()