![]()
![]()
![]()
THE ENVIRONMENT
The cool Benguela current flows north along the southern African west coast between latitudes 35°S and 15°S (Newman, 1977). To the east of Cape Point (Fig. 1) temperatures increase under the influence of the Agulhas Current (Shannon, 1966), whereas north of Walvis Bay the warm Angola current may intrude from the northwest (Stander, 1964; O'Toole, 1980; Badenhorst and Boyd, 1980). Total production of the Benguela upwelling system has been calculated as 375 × 10 ̃6 tons C/y (Cushing, 1969). Production up to 1.---5 g C/m ̃2/d has been measured in upwelling plumes (Nielsen and Jensen, 1957).
Along the South West African coast, mid-winter (July) marks the onset of upwelling. Upwelling attains a maximum in spring, is of little consequence in summer except towards the south, and is often absent during autumn (Stander, 1964). Intensity of upwelling is particularly high between the Orange River and Luderitz and lessens substantially north of Walvis Bay (Stander, 1964). Between latitudes 22°S and 19°S there are frequently onshore movements during spring and summer of a surface water mass whose temperature and salinity characteristics suggest that it derives from mixing of the cold Benguela Current from the south with the warmer Angola Current from the north (O' Toole, 1980).
Off the Republic of South Africa's Western Cape upwelling is initiated in spring, is strong during summer, is moderate or absent in autumn and is at a minimum in winter (Shannon, 1966). The reason for the discrepancy in upwelling seasons off South West Africa and the Western Cape is that in the north southerly to southeasterly winds are strongest from August through October, whereas in the south winds blowing parallel to the coast are maximal in summer (Stander, 1964; Shannon, 1966). The Western Cape system is rapidly pulsed and can be divided into four quasi-independent upwelling regions: the Namaqua area near Hondeklip Bay where extensive phytoplankton blooms occur; the Cape Columbine region from where upwelling plumes may extend northwards for more than 200 km under the influence of the Benguela Current; the Cape Peninsula system where up to three discrete plumes may develop simultaneously; and the region between Cape Agulhas and Cape Point (Shannon and Anderson, 1982). The upwelling plumes are separated from oceanic waters by sharp gradients of temperature and salinity. Strong equatorward jet currents are often associated with these frontal systems (Bang and Andrews, 1974). East of Cape Agulhas limited upwelling takes place off prominent capes, especially during summer and autumn (Schumann et al., 1982).
| Year | Catch (t) | Effort (std | South African c.p.u.e. |
|---|---|---|---|
| trawler days) | (t/std trawler day) | ||
| 1964 | 162 300 | 11 116 | 14,60 |
| 1965 | 203 300 | 18 755 | 10,84 |
| 1966 | 195 000 | 18 344 | 10,63 |
| 1967 | 176 700 | 17 652 | 10,01 |
| 1968 | 143 600 | 14 346 | 10,01 |
| 1969 | 165 100 | 19 153 | 8,62 |
| 1970 | 142 500 | 19 710 | 7,23 |
| 1971 | 202 000 | 28 491 | 7,09 |
| 1972 | 243 900 | 49 776 | 4,90 |
| 1973 | 157 800 | 31 751 | 4,97 |
| 1974 | 123 000 | 26 452 | 4,65 |
| 1975 | 89 600 | 19 227 | 4,66 |
| 1976 | 143 400 | 26 804 | 5,35 |
| 1977 | 97 500 | 20 145 | 4,84 |
| 1978 | 101 700 | 17 237 | 5,90 |
| 1979 | 90 400 | 14 747 | 6,13 |
| 1980 | 101 500 | 18 522 | 5,48 |
| 1981 | 99 600 | 17 148 | 5,81 |
| Year | Trachurus | Merluccius | Merluccius |
|---|---|---|---|
| ICSEAF Divisions 1.3 & 1.4 | ICSEAF Division 1.5 | ||
| Source | Babayanet al (1983) | Terre(1983) | Anon (1983) |
| 1965 | 93,5 | 99,7 | |
| 1966 | 99,6 | 212,4 | 122,2 |
| 1967 | 72,4 | 195,0 | 199,4 |
| 1968 | 69,2 | 382,7 | 247,7 |
| 1969 | 46,5 | 320,5 | 206,2 |
| 1970 | 50,4 | 402,5 | 224,7 |
| 1971 | 214,5 | 365,6 | 229,7 |
| 1972 | 72,2 | 606,1 | 214,0 |
| 1973 | 271,4 | 377,6 | 290,3 |
| 1974 | 193,1 | 318,8 | 195,7 |
| 1975 | 242,2 | 309,4 | 178,7 |
| 1976 | 456,2 | 369,8 | 209,7 |
| 1977 | 352,1 | 277,5 | 157,7 |
| 1978 | 516,2 | 258,1 | 124,2 |
| 1979 | 401,6 | 172,3 | 130,0 |
| 1980 | 544,1 | 90,5 | 70,1 |
| 1981 | 590,5 | 100,0* | 116,4 |
DISTRIBUTIONAL ECOLOGY OF THE FISH POPULATIONS
Seasonal patterns of distribution for contributors to the Western Cape purseseine fishery have been summarized by Crawford (1980a). Adult anchovies and pilchards aged between two and four tend to spawn in warm water found east of Cape Point (Figs. 2 and 3). The cool upwelled water of the west coast is avoided. Lower lethal temperature limits for eggs of pilchards and anchovies are 13.0°C and 14.0°C respectively, their rates of development increasing with temperature (King, 1977a; King et al., 1978).
Five-and six-age group pilchards used to spawn off St. Helena Bay in warm water outside the oceanic front, but heavy exploitation of juveniles means that few fish currently attain these ages (Crawford et al., 1978).
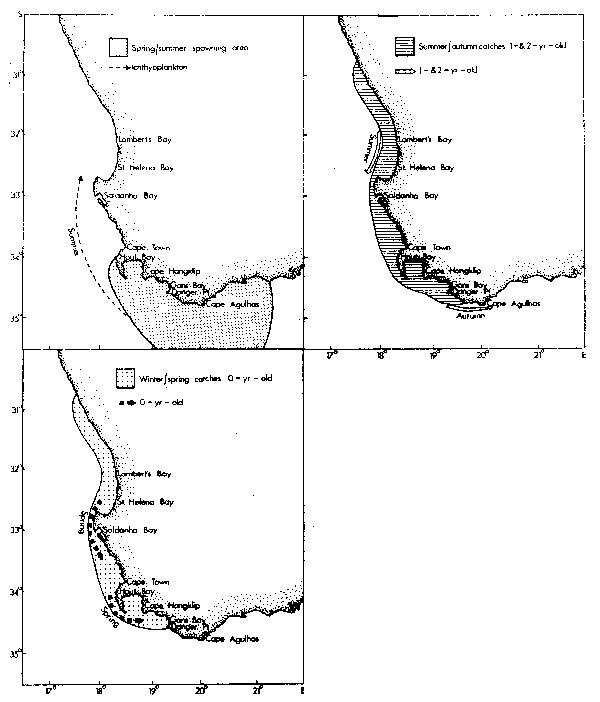
Fig. 2. The main spawning area and catch locations for different age components of the Western Cape Engraulis capensis population (modified from Crawford, 1980a).
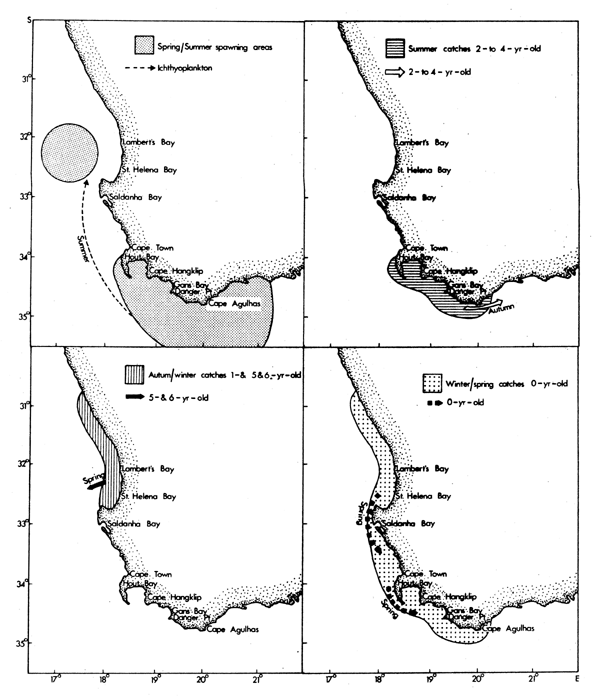
Fig. 3. The main spawning areas and catch locations for different age components of the Western Cape Sardinops ocellata population (modified from Crawford, 1980a)
Horsemackerel and roundherring also spawn along the west coast outside the oceanic front, although spawning grounds of the latter include areas that are subject to invasion by cold upwelling plumes (Figs. 4 and 5). Eggs of these species again develop more rapidly in warmer water, but can tolerate temperatures at least as low as 12.6°C (horse-mackerel) and 11.0°C (roundherring) according to King et al. (1977) and O'Toole and King (1974). The spawning season for anchovies, pilchards and horsemackerel is spring and summer, whereas that for roundherring is from late winter through early summer (Davies et al., 1981).
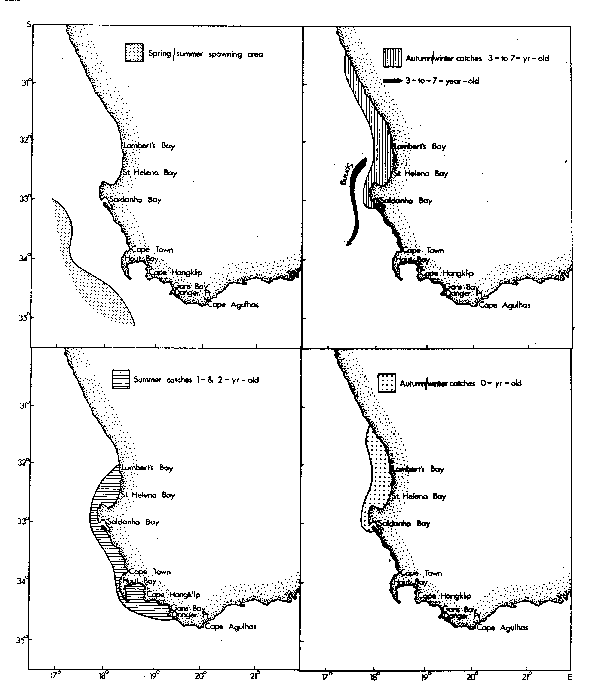
Fig. 4. The main spawning area and catch locations for different age components of the Western Cape Trachurus trachurus population (modified from Crawford, 1980a).
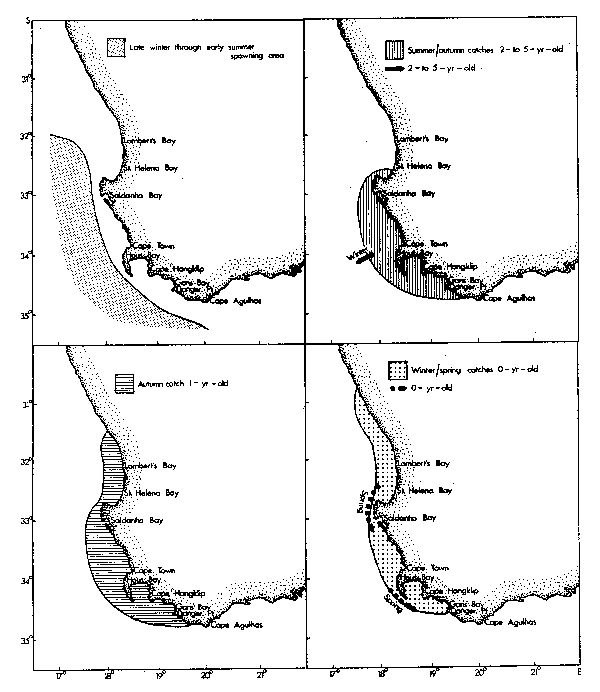
Fig. 5. The main spawning area and catch locations for different age components of the Western Cape Etrumeus teres population (modified from Crawford, 1980a)
Shelton and Hutchings (1982) have demonstrated the importance of the Good Hope Jet (Bang and Andrews, 1974) in the northward transportation of anchovy eggs and larvae. Ichthyoplankton spawned east of Cape Point may drift towards the jet in surface currents set in motion by the prevailing southeasterly winds of spring and summer (Duncan and Nell, 1969).
Recruitment of nought-year-olds of anchovies, pilchards, horsemackerel and round-herring takes place along the west coast from mid-autumn onwards. At this age they frequently occur together in mixed shoals and initially are most abundant to the north of St. Helena Bay. In winter, maximum standing stocks of plankton occur in the north of Cape Columbine (de Decker, 1973; Crawford et al., 1980; Shannon and Anderson, 1982). At the onset of spring there is a definite tendency for the young fish to move south into newly initiated regions of upwelling, and maturing anchovies and pilchards continue to the spawning grounds east of Cape Point. At the close of the spawning season they frequently migrate eastwards well past Cape Agulhas.
Five-and six-age group pilchard, adult horsemackerel and adult roundherring have been most available to the purseseine fleets from late summer through winter, moving offshore in other seasons to spawn.
Chubmackerel spawn off Cape Columbine (Fig. 6) in winter and early spring (Baird, 1975; 1977) when onshore currents, set in motion by northwesterly winds, bring relatively warm (14-15°C) water nearer to the coast (Shannon, 1976). Nought-year-olds are fully recruited at the beginning of each year, and occur inshore along the west coast in much the same region as adult roundherring with which they may form mixed shoals (Geldenhuys, 1978). They then assume a progressively more offshore existence, the adults generally only being encountered by purseseiners on their inshore spawning migration off Cape Columbine (Baird, 1978a). By this stage myctophids dominate their diet (Baird, 1978b) and it is noteworthy that Cape Columbine is also the region where most purseseine catches of Lampanyctodes are recorded (Crawford, 1980b).
Off South West Africa anchovies spawn inshore between the Cunene River and Mercury Island from late spring to early autumn (King, 1977b; le Clus and Kruger, 1982; le Clus and Melo, 1982). Serial spawning is likely and large adults are more fecund than smaller individuals (le Clus, 1979a). Eggs are carried north by the prevailing currents and most larvae occur in the warmer water north of Walvis Bay (O'Toole, 1977; Badenhorst and Boyd, 1980). Large shoals of juvenile anchovies recruit to the fishery in the vicinity of Walvis Bay, usually from May, whereas adults are caught inshore north of Cape Cross from autumn to early winter (le Clus and Melo, 1982; Thomas et al., 1982).
There are two main centres for the spawning of pilchard eggs: the Walvis Bay vicinity where production takes place from mid-spring to early summer, in seasurface temperatures ranging from 12.0°C to 16.5°C, and between latitudes 19°S and 21°S where eggs are spawned offshore in midsummer and early autumn in water temperatures of 16.5°C to 22.8°C (Stander, 1963; Matthews, 1964; King, 1977; le Clus and Thomas, 1982). Average oocyte mass of spring and early summer spawners is significantly greater than that of pilchards producing eggs in late summer and autumn, perhaps as an adaptation to the lower temperatures encountered (le Clus, 1979b). Dispersal of ichthyoplankton is in a northern direction (O'Toole, 1977) towards a nursery ground for juvenile pilchards near the border of Angola and Namibia/S.W. Africa(Schulein, 1973; Cram, 1977). Nought-year-olds recruit to the fishery from May between Palgrave Point and Walvis Bay, whereas older age groups are caught from autumn (le Clus and Thomas, 1982). The largest size-classes were traditionally caught in the Walvis Bay vicinity (Schulein, 1971; Cram, 1977).
Larvae of Sufflogobius were the most important element in ichthyoplankton sampled off South West Africa during the summers of 1972 to 1974 (O'Toole, 1978b). Spawning occurred over a wide area in inshore waters between July and February but was most intensive south of Walvis Bay from spring to early summer (O' Toole, 1978b). Acoustic records indicated that the species was most abundant between Cape Cross and Luderitz (Cruickshank et al., 1980).
Predators of schooling pelagic fishes include Cape fur seals Arctocephalus pusillus (Rand, 1959a), three species of guano producing seabirds, namely Cape gannets Sula capensis, jackass penguins Spheniscus demersus and Cape cormorants Phalacrocorax capensis (Davies, 1955; 1956; Rand, 1959b; 1960a; 1960b; Matthews, 1961; Batchelor, 1982; Crawford and Shelton, 1982), various tuna Thunnus species (de Jager et al., 1963; Nepgen, 1970), chubmackerel (Baird, 1978b), yellowtail Seriola lalandi (Crawford, 1980a) and snoek Thyrsites atun (Nepgen, 1979a). Snoek is a migratory fish species being caught off South West Africa from November to January, along the Cape west coast from April to July, and east of Cape Point from June to October (de Jager, 1955; Nepgen, 1979b).
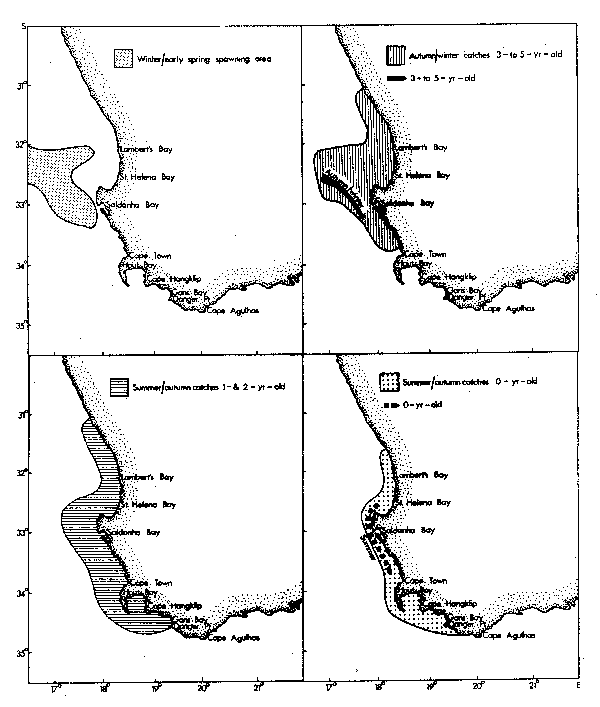
Fig. 6. The main spawing area and catch locations for different age components of the Western Cape Scomber japonius population (modified from Crawford, 1980a)
The spawning season for hakes off the Western Cape is late winter to spring (Botha, 1971), whereas newly hatched larvae occur off South West Africa from September to April, particularly in the vicinity of Walvis Bay (O'Toole, 1978a).
TRENDS IN THE COMMERCIAL FISHERIES
The Western Cape purseseine fishery commenced in 1943, but no accurate records of performance are available prior to 1950. Initially the fishery was largely preoccupied with exploitation of pilchards and horsemackerel (du Plessis 1959), and these two stocks continued to dominate landings until well into the 1960's (Table 1). Annual catches of pilchards in excess of 400,000 tons were recorded in the early 1960's, as a result of exceptionally powerful year-classes formed during the late 1950's (Newman and Crawford 1980). Subsequently recruitment returned to more normal levels, it was not possible to maintain the high yields, and managers of processing plants sought alternative resources to bolster diminishing returns. A small-meshed (13mm) net was introduced in 1964 and exploratory fishing for anchovies was initiated. Since 1966, anchovies have consistently been the major contributor to pelagic catches off the Western Cane, although substantial landings of chubmackerel were also recorded in the mid-1960's.
| Year | Combined species catch | Effort | Catch per unit effort |
|---|---|---|---|
| ('000 t) | (std. boat days) | (t/std. boat days) | |
| 1960 | 412,0 | 3,498 | 117,5 |
| 1961 | 490,9 | 3,717 | 132,5 |
| 1962 | 497,3 | 3,936 | 126,6 |
| 1963 | 427,0 | 4,482 | 95,7 |
| 1964 | 425,6 | 5,466 | 77,9 |
| 1965 | 480,1 | 9,580 | 50,1 |
| 1966 | 357,1 | 8,099 | 44,1 |
| 1967 | 509,3 | 12,059 | 42,2 |
| 1968 | 368,6 | 11,110 | 33,2 |
| 1969 | 352,0 | 10,349 | 34,0 |
| 1970 | 358,9 | 11,052 | 32,5 |
| 1971 | 324,9 | 10,126 | 32,1 |
| 1972 | 433,6 | 13,080 | 33,2 |
| 1973 | 451,4 | 12,249 | 36,9 |
| 1974 | 400,5 | 8,720 | 45,9 |
| 1975 | 407,4 | 10,329 | 39,4 |
| 1976 | 407,5 | 8,587 | 47,4 |
| 1977 | 357,2 | 11,168 | 32,0 |
| 1978 | 380,4 | 13,039 | 29,2 |
| 1979 | 380,9 | 10,983 | 34,7 |
| 1980 | 380,5 | 10,065 | 37,8 |
| 1981 | 379,2 | 10,821 | 35,0 |
The combined catch rose to almost 500,000 tons in the early 1960's, but has more recently been limited by a quota system to about 400,000 tons (Crawford, 1981a). Effort escalated considerably after the decline of the pilchard stock through an expansion in total fleet hold capacity, a move towards larger vessels and the introduction of fishing aids (Newman et al., 1979; Crawford, 1981a).
Exploitation of South West African pelagic stocks began in 1947 (Cram, 1977), and until 1968 catches were almost entirely of pilchards (Table 2). More than 1,3 million tons were caught in 1968 but landings then declined rapidly. The 1980 catch amounted to only 11,000 tons. Small-meshed (13 mm) nets were introduced but only became widely used from 1968-1969 (Cram, 1977; Schulein et al., 1978). Since then landings of anchovies have generally exceeded 100,000 tons, a peak catch of 355,000 tons being recorded in 1978. Catches of other species have been insignificant except for 140,000 tons of horsemackerel landed in 1971.
A combined-species catch of over 1,5 million tons was recorded in 1968. Total fish hold capacity of the fleet increased from c. 3,000 tons during 1957-1960 to over 14,000 tons in 1975 (Newman, 1977). However, effort measured in terms of fuel consumption peaked at around 23 million litres in 1969 before decreasing to less than 7 million litres in 1980 as a result of quota restrictions (F.H. Schulein, Sea Fisheries Research Institute, Cape Town).
Since the turn of the century hakes have formed the basis of a substantial demersal trawl fishery off southern Africa, and account for about 70 per cent of the trawl catch (Botha 1970). Catches of hake by the South African fishery (based mainly at Cape Town and in recent years at Saldanha) reached 1,000 metric tons in 1919, increased steadily to 100,000 tons in 1959 (Chalmers 1976) and attained a record of 153,000 tons in 1979 (Botha 1980). After 1962, the Cape hakes also became a sought-after target of distant-water trawlers from several foreign countries. In 1973, 14 different countries achieved a catch of just over one million tons with more than 300 large vessles, thereby establishing the Cape hake fishery as the largest of its kind in the world (Botha, 1980). The local fishery has traditionally confined its fishing activities mainly to the fishing grounds off the Western Cape and to a lesser extent to those off the southern coast of. South Africa. Foreign catches of hake have tended to concentrate in areas further north off the coast of South West Africa. After the introduction of a 200 nautical mile exclusive fisheries zone off the Republic of South Africa on 1 November 1977, the hake fishery off South Africa reverted to an almost exclusively local one, as it was before 1962 (Botha, 1980).
VARIABILITY IN THE ENVIRONMENT
Hutchings and Nelson (1983) developed a timeseries of upwelling indices for the Western Cape for the spring-summer spawning season of pilchards and anchovies. There was considerable variability between seasons, with no clear trend (Fig. 7). Upwelling was poor during the 1965/1966 and 1968/1969 seasons and good during 1974/1975 and 1975/1976. The indices are based on wind records and commences in 1960. In earlier years seasurface temperature information is available for inshore waters from routine research cruises in the St. Helena Bay region (Buys, 1959). A cool anomaly, probably indicative of powerful upwelling, occurred between 1953 and 1957. Warmer temperatures were experienced during the 1952/1953 and 1957/1958 seasons (Fig. 7).
A cool water anomaly for the Walvis Bay region off South West Africa during the mid-1950's lagged behind that of the St. Helena Bay area by about two years (Fig. 7; Stander, 1963). Boyd (1979) investigated variability of seasurface temperature off South West Africa during the period 1969 to 1978 and found it to be low between 1972 and 1975 and during the 1977/1978 spawning season for anchovies.
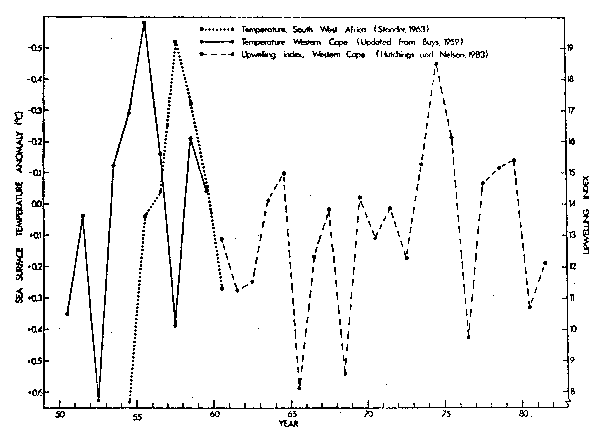
Fig. 7. Seasurface temperature anomalies and upwelling indies for the southern Benguela system, 1950-1982
VARIABILITY OF STOCKS DURING THE PERIOD OF EXPLOITATION
Recruitment
The most extensive timeseries of recruitment (defined in this paper as numbers of nought-year-old fish in the population) available for fisheries of the southeast Atlantic are those derived from Virtual Population Analysis (VPA) (e.g. Centurier-Harris, 1977; Newman and Crawford, 1980; Armstrong et al., 1983; Babayan et al., 1983; ie Clus and Melo, 1983; Payne and Leslie, 1983; Thomas, 1983). In all VPA's conducted the natural mortality rate (M) has been assumed constant throughout the period of investigation, and in many instances age compositions of catches in earlier years have been calculated from age at length information pertaining to a subsequent period (Crawford et al., 1978). Additionally the relatively shortlived nature of species exploited by the pelagic fisheries has often meant that cohorts have not contributed in a significant manner to the catches for more than a few years, thereby increasing the likelihood that trends derived from VPA may reflect availability of cohorts to the fishing fleets rather than absolute variations in year-class strength. The purseseine fisheries of the Southern Benguela system catch a mixture of species with preferential fishing for certain species (Crawford 1981a; 1981b) so that VPA indices may also be influenced by availability of other species. For these reasons VPA estimates of recruitment have been compared wherever possible with independent indices to test their validity. On account of assumptions that have to be made regarding terminal mortalities, VPA estimates for the most recent years cannot be considered reliable.
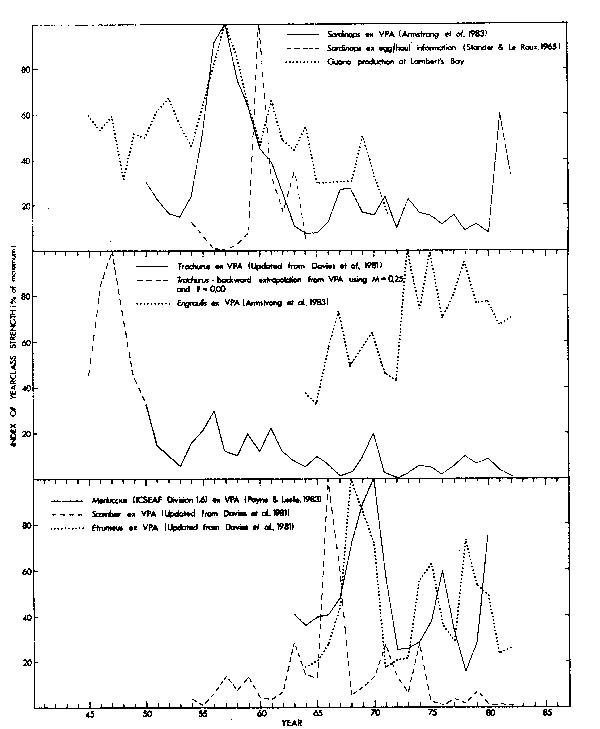
Fig. 8. Indices of recruitment for neritic stocks off the Republic of South Africa's Western Cape, 1945-1982
Western Cape Sardinops population:
VPA indices suggest that exceptionally good recruitment of pilchards occurred in the mid-1950's, especially 1956 and 1957 (Fig. 8), and as a result spawning biomass peaked during 1960/1961 (Newman and Crawford, 1980; Armstrong et al., 1983). Numbers of pilchard eggs recorded per haul of surface plankton nets also peaked marked1y in 1960 (Fig. 8; Stander and le Roux, 1968) and if used as measures of spawning biomass (e.g. Smith, 1972) again suggest a major upward perturbation of recruitment in the mid-1950's. Further evidence for this phenomenon is provided by a large increase in the amount of guano harvested at Lambert's Bay from 1956 to 1958 (Fig. 8). Lambert's Bay is located at the northern extremity of the fishing grounds and in the region where the nought-year-old pilchards recruit to the fishery (Crawford and Shelton, 1978; Crawford, 1980a).
VPA indices further suggest a second, but minor, upswing in recruitment after 1965. From 1969 through 1980 cohort strength of pilchards fluctuated around a level approximately 15 per cent of the 1957 maximum.
Western Cape Engraulis population:
VPA indices of year-class strength for the Western Cape anchovy population are only available from 1964. Recruitment was low in 1964 and 1965, fluctuated around an intermediate level from 1966 to 1972, reached a peak in 1973 and remained high until 1980 (Fig.8). The powerful year-classes from 1973 onwards resulted in a 1979 stock size that was double that of the mid-1960's (Fig. 9). Independent indices of anchovy recruitment are not available, but off the Western Cape handline catches of an important predator, Thyrsites, closely followed VPA estimates of total anchovy biomass from 1969 to 1978 (r = 0,708; p<0,02; n = 10). This relationship broke down in subsequent years (Fig. 9). The combined annual catch of Thyrsites in the Southern Benguela system rose from 8,000 tons in 1973 to 20,000 tons in 1977. It is unlikely to have been greater in previous years, but in 1978 more than 81,000 tons were landed, and this high exploitation possibly influenced stock size. Handline catches of Thyrsites off the Western Cape were exceptionally good in 1976 (Fig. 9) and may have resulted from an unusually high availability of Thyrsites to fishermen in that year. Purseseine catches of pilchards, another major forage (Nepgen, 1979a), were greater in 1976 than in any year since 1965 and 2,5 times the mean for the period 1969-1982 (Table 1). The increased VPA estimates of anchovy recruitment and biomass and the good handline catches of Thyrsites in the late 1970's may all have resulted from anchovies being more readily available to both fishermen and predators during this period. However, consistency of the trends makes an absolute increase in the biomass of anchovies after 1972 the more likely explanation. The large increase in recruitment in 1973 and the plentiful cohorts that followed could then be regarded as the most significant events in the dynamics of the anchovy stock since exploitation was initiated.
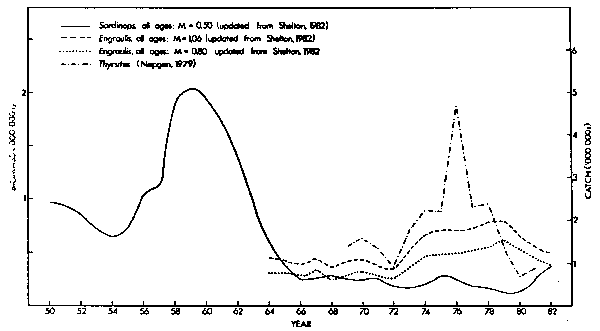
Fig. 9. Trends in biomass of Sardinops and of Engraulis and in handline catches of Thyrsites off the Republic of South Africa's Western Cape, 1950- 1982. Natural mortality rates of 0,80 (Newman and Craeford, 1980; Crawford, 1980c) and of 1,06 (MacCall, 1974) have been used for Engraulis
Western Cape Trachurus population:
Between 1950 and 1958 annual length compositions of purseseine catches of horsemackerel off the Western Cape were characterized by the progression of a distinct mode (Fig. 10). Geldenhuys (1973) identified this as consisting mainly of the 1946, 1947 and 1948 year-classes, so that the high catches of the early 1950's appear to have resulted from powerful recruitment during the late 1940's (Fig. 8). Only 65 per cent of horsemackerel otoliths have discernible age rings (Gendenhuys, 1973) making VPA of doubtful reliability (Newman and Crawford, 1980). VPA applied to the best available age information suggests that recruitment since 1950 has been considerably lower than during the late 1940's. A number of minor peaks, separated by intervals of up to five years, occurred between 1956 and 1970. Since 1970 recruitment has never exceeded 10 per cent of the 1947 maximum (Fig. 8).
Western Cape Scomber population:
VPA suggests that powerful recruitment of chubmackerel occurred during 1966 and 1967 (Fig. 8). The highest chubmackerel catches on record were landed between 1967 and 1969 (Table 1), when much of the harvest was netted close inshore (Crawford, 1981d). It is possible that environmental factors may have resulted in increased availability of chub-mackerel to the purseseine fleet during this period. However, catches of adult chubmackerel pursed offshore peaked during 1969 and 1970 at about 60,000 tons before declining to negligible quantities by 1975 (Baird, 1978a). The development of the 1966 and 1967 cohorts can be traced on distributional charts of catches and their length compositions, and from the charts it is evident that the 1966 and 1967 year-classes provided the bulk of the offshore catches of adults until 1971 (Crawford, 1981d). With their passing from the fishery the chubmackerel catch declined.
Minor increases in year-class strength of chubmackerel have been estimated by VPA for the late 1950's, 1963, 1971 and 1974, but even the best of these was less than 30 per cent of the 1966 peak (Fig. 8). Since 1974 recruitment has been consistently poor.
Western Cape Etrumeus population:
As with the anchovy population, VPA indices of roundherring recruitment are only available from 1964, when commercial fishing with the small meshed net was first permitted. Round herrings have relatively large scales, which they shed readily when disturbed. This frequently leads to clogged meshes and a net that is both difficult to purse and that will later require considerable time and effort to clean. Fishermen, therefore, generally avoid roundherrings when they can be recognized and when other fish are plentiful (Crawford, 1981b), complicating interpretation of VPA. Recruitment appears to have been at its highest from 1968 to 1970 with peaks also during 1974/75 and 1976/77. Catch per unit effort indices for the mixed-species purseseine fishery of the Western Cape were low from 1968 to 1972 (Table 5, Crawford 1981a), that is after the decline of the pilchard stock but before the increase of the anchovy population. A scarcity of fish during this period may have resulted in a higher proportion of effort being directed towards shoals of roundherrings. However, the 1969 and 1970 year-classes persisted into the catches of 1972 and 1973 respectively (Geldenhuys, 1978).
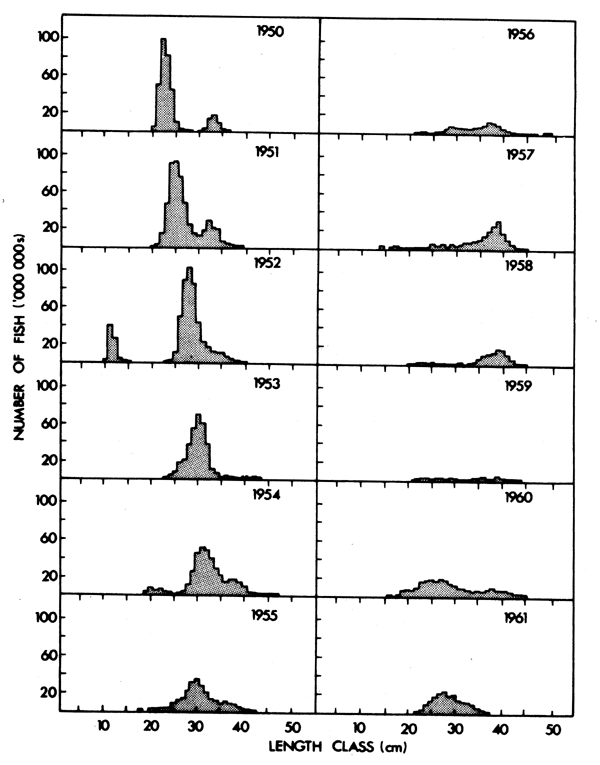
Fig. 10. Caudal length distribution of purseseine catches of Trachurus off Republic of South Africa's Western Cape, 1950-1961
Merluccius spp. in ICSEAF Division 1.6:
VPA estimates for Merluccius spp. in ICSEAF Division 1.6 (Payne and Leslie, 1983) indicate a large increase in year-class strength between 1968 and 1971 and a smaller peak in 1976. The cohorts of other years were moderate. (The 1980 increase cannot be considered reliable at this stage.) Selection factors for hake are 10, 80 and 100 per cent for fish aged one, two and three or older respectively (Payne and Leslie, 1983). The good year-class of 1976 appears to have improved catch rates in the late 1970's (Table 3). However, although the apparently powerful cohorts of the late 1960's and early 1970's were detected by VPA's at an early stage (Newman et al., 1976), they had no noticeable impact on indices of catch per unit effort in the early 1970's. From 1971 to 1973 there were large increases in both catch and effort (Table 3) as foreign participation in the fishery peaked.
South West African Sardinops population:
Age information for catches of pilchards off South West Africa are only reliable from 1967 (Thomas, 1979; 1983). Earlier Nawratil (1960) had used scales to estimate age composition of the catches, but Newman (1970a) discussing the results of a tagging experiment concluded that Nawratil's (1960) age compositions were invalid. Off South West Africa and Angola catches at the northern extremity of the pilchard fishing grounds are often of the smaller size-classes (Monteiro, 1962; Schulein, 1971). Angolan catches (from Schulein 1971) were exceptionally high during 1956 and 1957 (Fig. 11), suggesting powerful recruitment in these years. Numbers of pilchard eggs recorded per haul of surface plankton nets in the vicinity of Walvis Bay peaked markedly three years later. The steep decline in this parameter after 1960 has been attributed to an oceanographic anomaly resulting in conditions unsuitable for sexual development of pilchards (Stander and de Decker, 1969), but decreasing biomass of spawners following a return to more normal levels of recruitment may equally well have been the cause.
VPA (Thomas, 1983) indicates that recruitment increased from 1969 to a maximum in 1972 and then declined sharply (Fig. 11). The year-classes of the early 1970's gave rise to improvements in overall stock size and biomass of spawners, trends that are borne out by catch rates, aerial estimates of stock size and egg surveys (Fig. 12) as discussed fully by Butterworth (1980).
South West African Engraulis population:
VPA estimates of recruitment of anchovies off South West Africa (le Clus and Melo, 1983) fluctuated around a relatively stable level from 1972 to 1981 (Fig. 11). Ageing in earlier years is considered unreliable. The 1982 value is low but only preliminary. The strongest year-class occurred in 1978 and there were also peaks in 1973, 1975 and 1980. Apparently no increase in recruitment has been maintained over two successive years. Independent estimates of anchovy egg production from the 1978/79 to 1980/81 spawning seasons (ie Clus and Melo, 1982) match trends in recruitment of one year earlier (Fig. 11). Anchovies mature from one year of age (Crawford, 1981e; Armstrong et al., 1983).
South West African Trachurus population:
Off South West Africa VPA indices of recruitment of horsemackerel (Babayan et al., 1983) increased markedly from 1967 through 1978, with minor fluctuations (Fig. 11). A large catch of juvenile fish was recorded by the South West African purseseine fleet in 1971 (Table 2; Newman and Centurier-Harris, 1977) but foreign trawlers using midwater nets have been responsible for the bulk of the catches in other years.
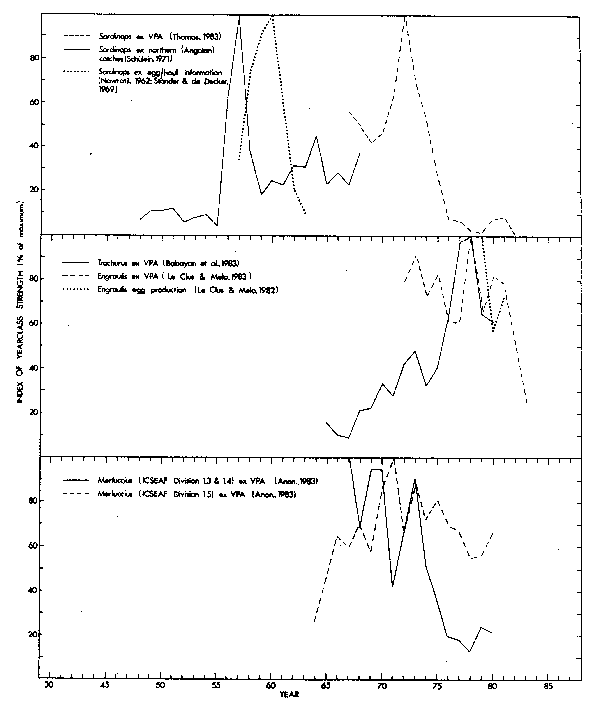
Fig. 11. Indices of recruitment for neritic stocks off South West Africa 1948-1982.
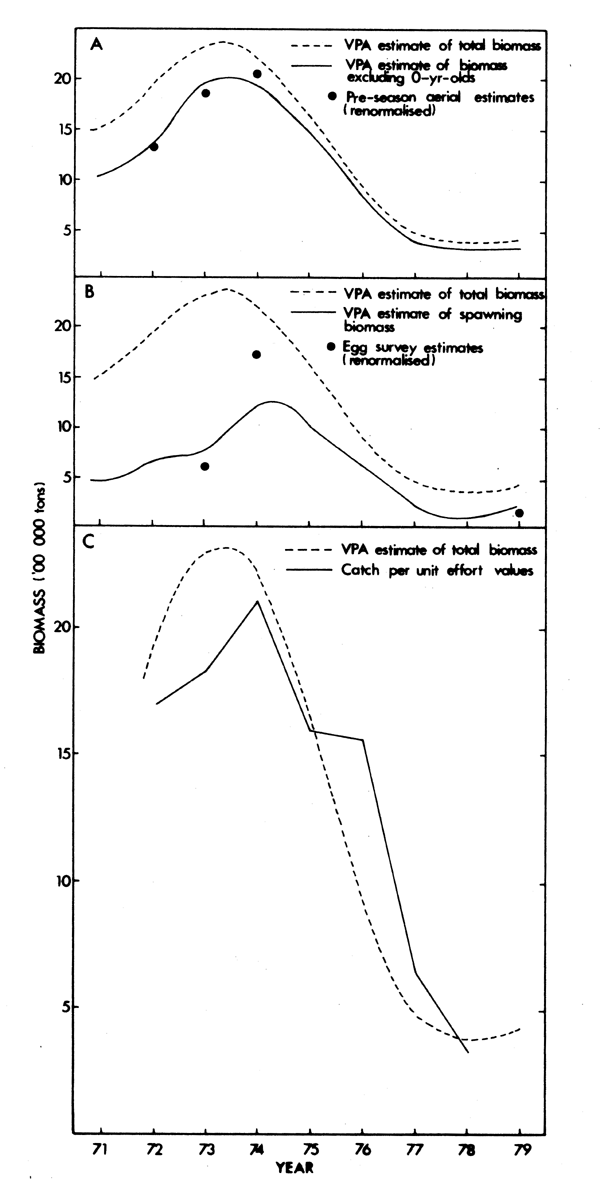
Fig. 12. Trends in Virtual Population estimates of biomass of Sardinops compared with (A) aerial estimates of stock abundance, (B) estimates of abundance from egg surveys and (C) catch rates, 1971-1979 (modified from Butterworth, 1980)
Merluccius spp. in ICSEAF Divisions 1.3, 1.4 and 1.5:
The fisheries for hake in ICSEAF Divisions 1.3, 1.4 and 1.5 are multinational. Assessments have previously been based on the assumption that one population was responsible for catches from Divisions 1.3 and 1.4 and a second for those from Division 1.5 (e.g. Newman et al., 1976a; 1976b). Preliminary results from a recent study of stock identity based on meristic characters confirm the existence of two populations within these Divisions, but indicate that the boundary between the two occurs at about latitude 23°S near the middle of Division of 1.4 (Lleonart et al., 1983). Therefore not too much significance can be attached to VPA indices of recruitment for the two formerly recognized populations (Fig. 11). Year-class strength for hakes in Divisions 1.3 and 1.4 apparently declined, with fluctuations, by approximately 80 per cent since 1967, whereas the general recruitment trend for hakes in Division 1.5 was an increase from 1964 through 1971 and subsequently a moderate decline.
Biomass
VPA estimates of biomass for some of the more important fish stocks in the south-eastern Atlantic are presented in Tables 6 and 7. During the 1950's and early 1960's Sardinops was the major resource off the Western Cape and South West Africa, but in both regions biomasses subsequently declined to less than 10 per cent of the maximum levels. After fluctuating between 600,000 and one million tonnes from 1950 to 1955, the Western Cape population reached a size of about two million in 1959, but fell to below 200,000 tonnes in the early 1970's (Newman and Crawford, 1980; Davies et al., 1981; Shelton, 1982). The South West African population was estimated from tagging at 5,67 million tonnes for the period 1957-1960 and 6,80 million tonnes between 1963 and 1966 (Newman, 1970a). The VPA estimate for 1967 was 6 million tonnes but this had decreased to 1,4 million tonnes by 1976 (Thomas, 1983). The decline was then arrested by favourable recruitment, leading Cram (1977) to speculate that stock had recovered. In retrospect it is evident that the "recovery" was shortlived, the biomass climbing to slightly over 2 million tonnes in 1973 and 1974 but then plummeting to less than 50,000 tonnes by 1979 (Thomas, 1983).
| Year | Sardinops | Trachurus | Engraulis | Merluccius | Merluccius |
|---|---|---|---|---|---|
| ICSEAF | ICSEAF | ||||
| Divisions | Divisions | ||||
| 1.3 & 1.4 | 1.5 | ||||
| Ages | All | 1 + older | All | 2 - 7 | 2 - 9 |
| Source | Thomas | Modified | Le Clus | Anon | Anon |
| (1983) | from Babayan | & Melo | (1983) | (1983) | |
| et al., (1983) | (1983) | ||||
| 1957-1960 | 5,670* | ||||
| 1963 | 6,804* | ||||
| 1964 | 6,804* | ||||
| 1965 | 6,804* | 729 | |||
| 1966 | 6,804* | 1,092 | 706 | ||
| 1967 | 5,955 | 982 | 726 | ||
| 1968 | 4,161 | 947 | 1,063 | 735 | |
| 1969 | 2,402 | 1,047 | 1,130 | 643 | |
| 1970 | 1,449 | 1,177 | 1,230 | 606 | |
| 1971 | 1,376 | 1,439 | 1,281 | 626 | |
| 1972 | 1,733 | 1,477 | 646 | 1,451 | 689 |
| 1973 | 2,114 | 1,832 | 748 | 1,033 | 730 |
| 1974 | 2,125 | 2,056 | 572 | 878 | 624 |
| 1975 | 1,582 | 2,203 | 551 | 1,025 | 625 |
| 1976 | 836 | 2,461 | 504 | 1,061 | 824 |
| 1977 | 315 | 2,583 | 554 | 775 | 841 |
| 1978 | 100 | 3,085 | 739 | 557 | 481 |
| 1979 | 48 | 3,551 | 481 | 392 | 462 |
| 1980 | 51 | 3,799 | 463 | 294 | 460 |
| 1981 | 125 | 3,821 | 519 | 333 | 432 |
| 1982 | 91 | 289 |
* From tagging (Newman, 1970a).
| Year | Sardinops | Trachurus | Engraulis | Scomber | Etrumeus | Merluccius |
|---|---|---|---|---|---|---|
| 1950 | 960 | 620 | ||||
| 1951 | 950 | 690 | ||||
| 1952 | 861 | 670 | ||||
| 1953 | 713 | 610 | ||||
| 1954 | 641 | 552 | 38 | |||
| 1955 | 740 | 440 | 51 | |||
| 1956 | 1 040 | 362 | 48 | |||
| 1957 | 1 130 | 330 | 37 | |||
| 1958 | 1 900 | 252 | 56 | |||
| 1959 | 2 040 | 220 | 67 | |||
| 1960 | 1 940 | 230 | 69 | |||
| 1961 | 1 700 | 171 | 70 | |||
| 1962 | 1 400 | 151 | 42 | |||
| 1963 | 980 | 92 | 45 | |||
| 1964 | 592 | 94 | 310 | 80 | 22 | 565 |
| 1965 | 382 | 86 | 310 | 69 | 34 | 618 |
| 1966 | 243 | 59 | 290 | 144 | 48 | 639 |
| 1967 | 260 | 48 | 343 | 250 | 58 | 577 |
| 1968 | 290 | 45 | 250 | 201 | 52 | 548 |
| 1969 | 241 | 46 | 290 | 160 | 47 | 563 |
| 1970 | 233 | 5 | 323 | 110 | 62 | 563 |
| 1971 | 252 | 3 | 283 | 90 | 57 | 632 |
| 1972 | 182 | 4 | 260 | 102 | 50 | 616 |
| 1973 | 180 | 3 | 370 | 96 | 46 | 468 |
| 1974 | 202 | 1 | 460 | 80 | 40 | 324 |
| 1975 | 280 | 1 | 483 | 102 | 80 | 231 |
| 1976 | 250 | 2 | 500 | 87 | 100 | 218 |
| 1977 | 184 | 4 | 520 | 111 | 120 | 160 |
| 1978 | 171 | 3 | 544 | 6 | 97 | 199 |
| 1979 | 130 | 5 | 610 | 10 | 48 | 180 |
| 1980 | 130 | 11 | 520 | 14 | 63 | 158 |
| 1981 | 270 | 21 | 440 | 25 | 80 | 131 |
| 1982 | 370 | 3 | 380 | 33 | 82 |
Biomass indices for the horsemackerel population of the Western Cape decreased markedly after the early 1950's (Table 7). The indices are based on purseseine catches only, although the species is also caught demersally (de Villiers, 1977). The population appears to have the centre of its distribution offshore (Fig. 4; Crawford, 1980c) and catchability to purseseiners increases with increasing population size (in prep.), suggesting that a large stock expands its geographic distribution and in so doing becomes more readily available to the purseseine fleet.
Biomass of the chubmackerel population of the Western Cape increased from a level of about 50,000 tonnes during the 1950's to 250,000 tonnes in 1967, largely as a result of powerful 1966 and 1967 year-classes (Newman and Crawford, 1980). The stock then decreased with depressed biomasses being calculated for the late 1970's (Table 7). Biomass levels of hakes in each of the ICSEAF Divisions considered have also deteriorated in recent years (Tables 6 and 7), whereas that of the anchovy population off South West Africa remained fairly stable during the 1970's fluctuating between 450,000 and 750,000 tonnes (Table 6).
Increased population sizes have recently been observed for three stocks in the south-east Atlantic, those of anchovy and roundherring off the Western Cape and horsemackerel off South West Africa (Tables 6 and 7). In their respective regions the anchovy and horse-mackerel stocks are currently the most important of those for which biomass estimates are available. Interestingly both increased in a sigmoid fashion, following large decreases in the local pilchard populations, and they attained their maxima 10 to 13 years after the collapses of the pilchard stocks were complete (Fig. 9 and 13).
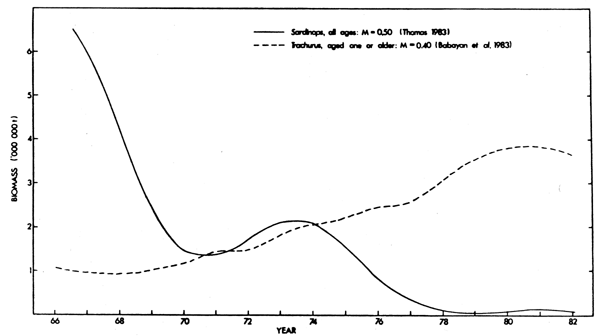
Fig. 13. Trends in biomass of Sardinops and Trachurus off South West Africa, 1966-1982
The Verhulst-Pearl logistic growth curve was applied to both the Western Cape
Engraulis and South West African Trachurus populations in a manner similar to the catch-corrected
model described by MacCall (1980). Constants for the instantaneous natural rate
of decrease in biomass (M-G) were derived using mass at age information and instantaneous
natural mortality rates of 0,8 and 0,4 for anchovies and horsemackerel respectively
(Crawford, 1980c; Babayan et al., 1983). For anchovies M-G was calculated for ages one
to three, as contrasted with ages two to three compared by MacCall (1980), on account of
the large proportion of young anchovies in commercial catches off the Western Cape
(Crawford et al., 1978). Ages two to were considered for horsemackerel since all
contribute to the midwater trawl catch (Babayan et al., 1983). Each year's VPA estimate
of biomass (Bt) was reduced by the natural rate of decrease and incremented by the next
year's recruitment (Rt + 1) to provide an estimate of the biomass (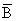 t + 1) that would
have occurred at the beginning of the next year under conditions of no fishing. Thus
t + 1) that would
have occurred at the beginning of the next year under conditions of no fishing. Thus
t + 1 = Bt + Rt + 1.
Assuming a constant , which was estimated at slightly above the maximum biomass observed each pair of Bt and t+1 was used to derive the value r in the logistic equation
Bt = B(1+e ̃ A-rt)
A mean r was then calculated and used as an initial estimate in the iterative model described by MacCall (1980) to obtain final estimates of B, A and r.
The difference between t+1 and Bt+1, that is the difference between the projected
biomass under conditions of no fishing and the VPA estimate of biomass based on actual
exploitation, was taken to be the "equivalent end-of-year catch" (cf. MacCall, 1980).
Biomass was then incremented assuming logistic growth and decreased after each year by
the equivalent end-of-year catch. Timeseries of biomasses computed in this way are
compared with estimates calculated by VPA in Fig. 14. Good agreement suggests that the
Verhulst-Pearl logistic equation describes in a reasonable manner growth of the Western
Cape anchovy and South West African horsemackerel populations in recent years.
Range
During the early years of commercial exploitation of pelagic species off the Western Cape substantial quantities of large adult pilchards were regularly caught off St. Helena Bay (du Plessis, 1959). On the basis of length information these fish are believed to have been aged five or older (Crawford et al., 1978). Following the decline of the pilchard stock in the late 1960's and the introduction of small-meshed nets in 1964 adult pilchards ceased to occur in significant numbers west of Cape Point. The bulk of the adult population, comprising fish aged two to four, currently inhabits the waters east of Cape Point (Crawford, 1981f). Results of historical egg surveys suggest that when adult pilchards were abundant off St. Helena Bay spawning also occurred east of Cape Point (Crawford, 1981f). The depletion of the pilchard stock was thus characterized by a contraction in the distribution of adults, and the contraction was towards the warmer waters of the region.
A similar phenomenon appears to have taken place off South West Africa. Prior to the collapse of the pilchard fishery shoals of large fish were encountered well to the south of Walvis Bay, and were exploited by boats operating from Luderitz (Schulein, 1971). After the stock had been substantially depleted boats from Luderitz were forced to fish north of Walvis Bay, and eventually the processing plant at Luderitz ceased operations. Over the period 1959 to 1973 the fishing range of boats operating from Walvis Bay was related to stock size, boats travelling further to the north during periods of low abundance (Cram, 1977). Quantities of pilchard eggs spawned off Walvis Bay declined substantially following the resource reduction (Matthews, 1964; King, 1977). Spawning still occurs in warmer water further north (King, 1977; le Clus and Kruger, 1982).
Fig. 14. Population growth of the Western Cape Engraulis and South West African Trachurus populations as estimated by Virtual Population Anaalysis and by fitting the logistic growth eduation, 1964-1982
VARIABILITY OF RESOURCES PRIOR TO THE INITIATION OF COMMERCIAL EXPLOITATION
Recruitment and biomass
The seabird island at Lambert's Bay is located at the centre of the recruitment grounds for pilchards, anchovies, horsemackerel and roundherring off the Western Cape (Crawford, 1980a). Since the late 1890's there have been three large oeaks in the amount of guano collected at this island, separated by intervals of approximately 30 years (Fig. 15). The final peak was significantly correlated (r = 0.783; |p<0.001; n = 20) with the eruption of pilchard recruitment during the late 1950's, leading to speculation that similar powerful cohorts may have been formed during the 1890's and 1920's (Crawford and Shelton, 1978; Newman and Crawford, 1980). The peak of the 1890's was significantly correlated (r = 0.804; p<0.02; n = 7) with catch rates of snoek, a major predator of pelagic fishes in the southern Benguela system (Crawford and Shelton, 1978), and that of the 1920's with handline catches of snoek off the western Cape {r = 0.739; p<0.01; n = 18). At many seabird islands off southern Africa, including Lambert's Bay, the guano harvest is related to numbers of breeding seabirds (Crawford and Shelton, 1978).
Prior to the initiation of commercial exploitation of pelagic fishes off southern Africa, guano harvests at seabird islands were dependent on availability of shoaling pelagic fishes (Hutchinson, 1950). Losses of guano caused by rain was also of importance (Hutchinson, 1950), but global economic factors apparently exerted little influence, in an attempt to nullify effects of rain, guano harvests at islands off the Republic of South Africa and off South West Africa were considered as the moving averages of three years (Fig. 16). Historical accumulations of guano had been cleared by the middle of the nineteenth century (Hutchinson, 1950). Trends for the two regions were similar until the mid-1940's, that is before the commencement of purseseine fishing. There were large increases in production from 1900 through the mid-1920's, followed by decreases in both regions. Yields were again high in the late 1930's and early 1940's. Trends at islands off South West Africa, in more recent years may have been influenced by the construction of guano platforms between Walvis Bay and Cape Cross from 1930 (Fig. 1), although at the same time natural breeding sites in the Walvis Bay vicinity were lost through islands in lagoons becoming joined to the mainland (Rand, 1952; Cooper et at., 1982). Guano harvests at islands off South Africa stabilized at about 2000 t in the 1950's before declining to half this level from the early 1960's. This decline corresponded to the depletion of the Western Cape pilchard stock (Crawford and Shelton, 1978). A decline of similar magnitude at islands off South West Africa during the late 1960's occurred coincidentally with the collapse of the local pilchard stock (Table 7).
Range
Neither pilchards nor anchovies were recorded as prey items of gannets breeding at Bird Island, near Port Elizabeth, in the 1930's (Rand, 1959b). Sample size is not known, but from 1978 to 1981 (n = 2031) these two species contributed 60 per cent of the diet (Batchelor, 1982). Numbers of gannets breeding at Bird Island have increased markedly in recent years, as have population sizes of penguins and Cape cormorants at islands east of Cape Point (Randall and Ross, 1979; Crawford and Shelton, 1982). It appears that abundance of pilchards and anchovies off Port Elizabeth was low at the start of the century, whereas it is now likely to be substantial.
Off South West Africa guano production at each of the major islands was high during the 1920's (Fig. 17). There were subsequent declines at each centre, but those at the two northernmost islands (Mercury and Ichaboe) were considerably less severe than at islands located further south. If the exceptional guano yields of the 1920's resulted from a high biomass of pelagic fishes, it is likely that as abundance of fish declined the stocks contracted to the north.
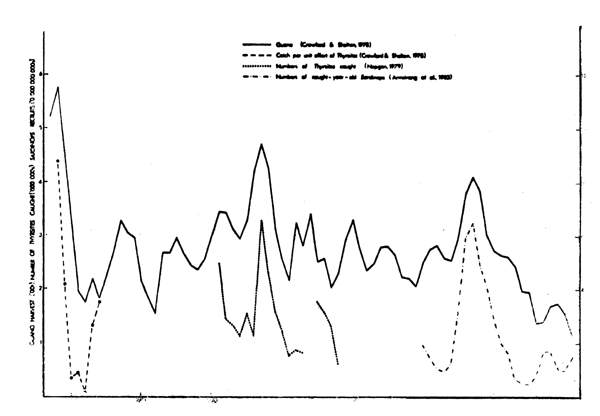
Fig. 15. Trends in the gruano harvesst at Lambert's Bay, and in catch rates of Thyrsites and in recruitment of Sardinops off the Western Cape, 1897-1972. Values shown are the moving averages of three years.
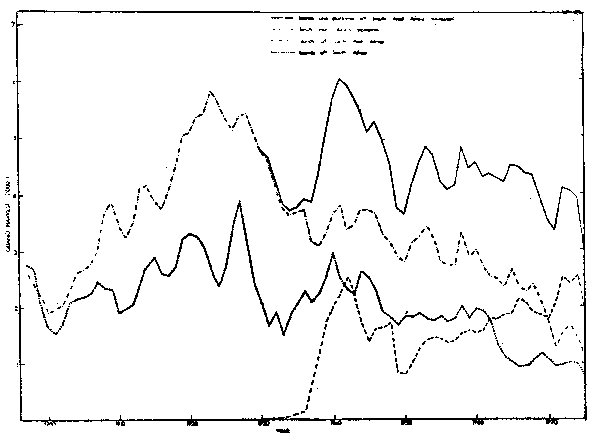
Fig. 16. Trends in the guano harvest at islands off South and South West AFrica, and at guano platforms along the South West African coast, 1987-1974. Values shown are the moving averages of three years.
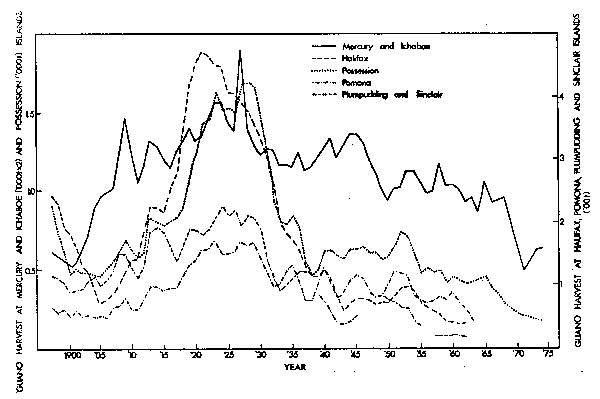
Fig. 17. Trends in the guano harvest at particular islands or groups of islands off the South West African coast, 1897-1974. Values shown are the moving averages of three years
Visual inspection of a correlation matrix of trends in guano harvests at islands off the southern African coast between 1897 and 1975 (during which period guano was collected by the South African government) suggests that the South West African islands form a distinct group (Fig. 18). Islands off South Africa are loosely correlated with one another and with those off South West Africa. The same technique has been applied by MacCall (1982) to examine subpopulation structure of anchovies off California.
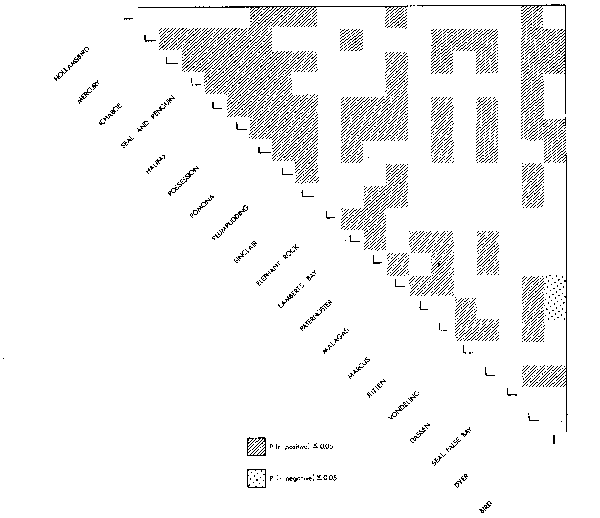
Fig. 18. Pattern of correlation between trends in guano harvests at islands off South West Africa. Islands are listed from north (Hollambird) to south (Bird) around the coast
CHARACTERISTICS OF VARIABILITY IN THE SOUTHERN BENGUELA SYSTEM
A number of generalized conclusions can be formulated from the foregoing concerning variability of neritic stocks in the southern Benguela system.
1. VPA indices of recruitment and biomass are in many instances validated by independent estimates of year-class strength or abundance. Either VPA and the alternative estimates are both influenced in a similar manner by availability of fish stocks, or VPA adequately described population trends. VPA is least certain for non-target species, such as roundherring off the Western Cape, and during the early stages of a fishery, especially if age information is inadequate.
2. Accepting the definition of an eruptive fluctuation provided by Caughley (1970) as a steady rise in numbers over at least two generations (cohorts in our usage) followed by a marked decline, short term eruptions of recruitment occurred in most of the fish populations examined. Exceptions were the two anchovy populations and the Western Cape chubmackerel and South West African horsemackerel stocks. Time series for anchovies extend back to 1964 off the Western Cape and 1972 off South West Africa. An increase in numbers over two cohorts was observed twice in the Western Cape, but neither population can yet be considered to have suffered a marked decline on account of the uncertainty of VPA estimates for the most recent years. There was exceptional recruitment of chubmackerel off the Western Cape in 1966 and 1967 followed by a subsequent decrease, but the 1967 cohort was less powerful than its predecessor. Year-class strength of horsemackerel off South West Africa has increased considerably in recent years. The eruptions of recruitment experienced by other populations were of short duration (two to six years) and often of sufficient magnitude to sustain the fisheries for a number of succeeding years. Examples include the horsemackerel and pilchard populations of the Western Cape.
3. Short-term recruitment failures of comparable magnitude to the eruptions have not been observed, but recruitment may decline to and continue at very depressed levels. The horsemackerel population of the Western Cape is an example.
4. A number of fish populations may experience favourable recruitment simultaneously. The pilchard stocks of the Western Cape and South West Africa both experienced recruitment eruptions during the mid 1950's. Year-class strength of horsemackerel off the Western Cape increased at the same time. Anchovies and horsemackerel off South West Africa both attained peak recruitment during the late 1970's.
5. The greatest relative increases in recruitment are often experienced by the species which has the largest biomass at the time of eruption. In particular off the Western Cape pilchards had the highest biomass in 1956 and anchovies in 1973 and off South West Africa horsemackerel during 1977-1978.
6. Some large increases in recruitment cannot be linked to any obvious environmental phenomena, for example the exceptionally strong anchovy year-class in 1973 off the Western Cape. A high upwelling index in following years (Fig. 7; Hutchings and Nelson, 1983) may have influenced formation of the 1973 cohort. However, environmental factors cannot be altogether discounted as the purseseine catch of Lampanyctodes in 1973 was far larger than in any other year (Table 1).
7. Some of the large-scale increases in recruitment can be plausibly related to environmental factors. Thus the exceptionally powerful 1966 cohort of chubmackerel was formed during a season in which the upwelling index was noticeably poor. Warm oceanic water is preferred by chubmackerel for spawning (Shannon, 1976). (Parrish and MacCall (1978) found that for the period 1931-1938 increased recruitment of Pacific mackerel S. japonicus was associated with increased seasurface temperature, reduced sealevel and reduced atmospheric pressure during the spawning season.) Off South West Africa and the Western Cape strong pilchard year-classes of the mid-1950's followed cool anomalies of seasurface temperatures, expected to be indicative of good upwelling.
8. Major perturbations in the populations of pelagic fishes appear to have occurred prior to the initiation of commercial purseseining. Guano harvests at Lambert's Bay (at the centre of the recruitment grounds for pilchards, anchovies, horsemackerel and roundherring off the Western Cape) have exhibited three major peaks since the late 1890's. Each peak has been significantly correlated with an index of recruitment of a pelagic fish population or an index of abundance of a predator (Thyrsites) of pelagic fishes. Less pronounced perturbations occurred more frequently, with a mean interval of about six years.
9. Predators and commercial fisheries may compete for surplus production arising from eruptions of recruitment. The guano harvest at islands off South Africa stabilized during the early years (1943-1960) of purseseine exploitation of pelagic fishes.
10. Large fluctuations in the size of a major fish population are reflected in the combined species biomass of a region. The guano harvest at islands off South Africa declined markedly following the collapse of the pilchard stock, indicating a substantial reduction in availability of fish prey to seabird predators. Subsequent recoveries in seabird numbers have coincided with an increase in the biomass of anchovies (Crawford and Shelton, 1982).
11. Population growth of a species may be influenced by the overall biomass of fish in a system. A logistic form of growth from moderate biomass levels has been observed for the Western Cape anchovy and South West African horsemackerel stocks. Both followed severe depletions of the local pilchard populations.
12. The geographical distribution of species is sometimes related to population size. Severe collapses of the Western Cape and South West African pilchard stocks were both characterized by contractions in the range of the adult population. In each instance the reduced parent stock favoured warmer waters for spawning. A range contraction towards warmer water of the South West African pilchard stock (inferred from guano records) may have occurred prior to the initiation of commercial exploitation.
13. The southern Benguela upwelling region appears to comprise at least two sub-systems. Population trends, meristic and genetic studies, tagging and a correlation matrix for guano harvests at different seabird islands suggest that fish populations off the Western Cape are discrete from those off South West Africa.
14. The same driving mechanism may influence population trends in both subsystems of the southern Benguela region. Combined guano harvests for islands off the Republic of South Africa and for those off South West Africa revealed similar trends prior to the initiation of commercial purseseine fishing. Each subsystem experienced a cool water anomaly in the mid 1950's, although that off the Western Cape preceded that off South West Africa. Large increases in recruitment of pilchards occurred in both areas in the mid 1950's.
DISCUSSION
A number of factors may be expected to influence population variability, including intraspecific controls (e.g. Parrish and MacCall, 1978; Ricker, 1954; Csirke, 1980; lles, 1980; MacCall, 1980b), interspecific relationships (e.g. Silliman, 1969; Radovich, 1979), environmental conditions (e.g. Smith, 1978; Kondo, 1980; MacCall, 1980a; Shuntov and Vasil'kov, 1982; Skud, 1982) and commercial exploitation (e.g. Ricker, 1963).
A preliminary investigation of phytoplankton primary production off the Western Cape has suggested that, if the primary production ration available to shoaling pelagic fishes is 18 per cent (Smith and Eppley, 1982), food may be limiting the biomass of the mixed-species pelagic resource off the Western Cape (Shannon and Henry, 1983). Intraspecific population controls as well as interspecific competition might then be expected. The pilchard has exhibited the ability to reduce its size at sexual maturity at low biomass levels (Crawford et al., 1980). Other density-dependent responses of pelagic fishes in the southern Benguela region are discussed in detail by Shelton and Armstrong (this consultation) and so are not considered further here.
In a review of the replacement of depleted fish stocks by other species Daan (1980) concluded that this phenomenon was perhaps most clearly established for the group of species that included the sardine-anchovy complex. Off both the Western Cape and South West Africa, seasons and localities for spawning by Sardinops and Engraulis are similar. Juveniles of the two genera occur together in mixed shoals (Crawford, 1981e) and the feeding ecology of juveniles and adults is similar (King and Macleod, 1976). Collapses of pilchard stocks off the Western Cape and off South West Africa during the 1960's were followed in the 1970's by logistic-type growths of anchovy and horsemackerel populations. A logistic form of growth has also been described for the Californian anchovy E. mordax population (MacCall, 1980). This too followed the decline of a Sardinops stock, but MacCall (1980a) preferred to a hypothesis of species replacement one of a population recovery by the anchovy stock to more normal levels of abundance after a prolonged series of poor recruitment.
A second interspecific relationship is that between a predator and its prey. Hairston et al., (1960) concluded that, whereas interspecific competition must necessarily exist among populations of producers, carnivores and decomposers, herbivores are seldom limited by food, appear most often to be controlled by predators, and are therefore not likely to compete for common resources. Although more recently Caughley (1970) has shown that food may have an important influence on population trends of herbivores, the possible significance of predation for shoaling pelagic species has been highlighted by Radovich (1979). Off southern Africa clear relationships exist between indices of abundance of four dominant predators (Cape gannets, jackass penguins. Cape cormorants, snoek) and their prey. In the 1950's pilchards were the dominant forage of each predator (Davies, 1955; 1956; Rand, 1959b; 1960a; 1960b; Matthews, 1961), but following depletions of the pilchard resources the importance of anchovies in their diets increased greatly (Nepgen, 1979a; Crawford and Shelton, 1982; Furness and Cooper, 1982). For snoek at least the change of prey occurred immediately after the collapse of the Western Cape pilchard population (Nepgen, 1979b). Should predators increase following the eruption of a major prey population, return of the prey to more normal levels of abundance and a consequent diversification in the diet of predators could result in an increased natural mortality rate for all prey species until the predators themselves again come into equilibrium with their forage. In this manner replacement of one pelagic species by another might be delayed.
Environmental factors are a frequently cited cause of variability in pelagic fish populations (e.g. Csirke, 1980; Kondo, 1980; MacCall, 1980a; Saville, 1980). Boyd (1979) found a significant correlation between variability of seasurface temperatures and recruitment. Recruitment was favoured by reasonably constant temperatures and it was argued that feeding success of anchovy larvae was greater in stable conditions (Boyd, 1979). The present paper has shown that a number of eruptions of recruitment for neritic stocks in the southern Benguela system can be plausibly linked to environmental anomalies.
The similarity between biomass trends off South Africa and off South West Africa, as inferred from guano harvests, and simultaneous eruptions in recruitment, sometimes over a wide geographic region, may be evidence that environmental phenomena operating over a scale of hundreds of km are more important than localised events in determining recruitment in the southern Benguela system. At this scale the use of satellite imagery for environmental monitoring as an aid to fisheries management may be useful (e.g. Shannon and Anderson, 1982). Skud (1982) cites instances in which changes in abundance of dominant and subordinate species of fish were respectively positively and negatively correlated with environmental factors that improved survival. In Californian waters a number of species experienced poor recruitment during the late 1940's and early 1950's (MacCall, 1980a). Off southern Africa year-class strength for two species improved markedly on at least two occasions. In each instance the largest relative increase in recruitment was experienced by the stock having the largest biomass.
The Japanese sardine S. melanosticta fishery recently increased dramatically after a period of poor catches lasting about 30 years (Kondo, 1980). Biomass of the Californian anchovy was high in the early 1940's and again in the mid 1960's (MacCall, 1980a). Off the Western Cape there may have been three large eruptions of recruitment since the late 1890's at intervals of approximately 30 years. Guano indices suggest that smaller perturbations also occurred at intervals of about six years. Such a happening would have ensured the formation of at least one strong year-class for each generation of pilchards. Anchovies, with their shorter lifespan (Crawford et al., 1978), are likely to be adapted to a shorter cycle and estimates of recruitment indicate an alternation of good and not-so-good year-classes (Figs. 8 and 11).
Moderate fishing of stocks that prior to exploitation include a large number of age groups can cause a major reduction in population size (Ricker, 1963). Off southern Africa intensive fishing of the multispecies pelagic stocks with smallmeshed nets has led to a current dominance of species having a short generation time (Newman and Crawford, 1980). Species characterised by a greater longevity, such as pilchards, horsemackerel and chubmackerel, now contribute little to the landings and a high fishing mortality on the younger ages maintains the parent populations at a low level (Newman and Crawford, 1980). Reductions in the geographic range of egg production, as have been observed for both pilchard populations, could adversely affect the probability of larvae encountering favourable environmental conditions, increase chances of recruitment failure (Saville, 1980) and decrease likelihood of favourable eruptions (Newman and Crawford, 1980).
In conclusion it appears that variability of the multispecies neritic complex in the southern Benguela system is to some extent biologically controlled, especially at the biomass levels. However, environmental factors and exploitation patterns are likely to be of even greater importance, the former in influencing largescale eruptions of recruitment and the latter in determining species composition. Eruptions of recruitment are naturally infrequent, and frequency of occurrence may be further reduced by changes in age structure and geographic distribution of species brought about by fishing.
ACKNOWLEDGEMENTS
We thank K. van Niekerk for assistance with programming of the model to examine logistic growth, K.L.M. Bentz and P.B. Crawford for typing the manuscript, and A. van Dalsen for the artwork. We are grateful to H.G. v.d. Boonstra, A.J. Boyd, D.S. Butterworth, F. le Clus, A.I.L. Payne, F.H. Schulein and R.M. Thomas for useful comments on the manuscript. One of us (RJMC) is indebted to SANCOR funding that enabled participation at the Costa Rica consultation.
REFERENCES
Anon. 1983. Report of the standing committee on stock assessment (Stock). Colln.scient.Pap.int.Commn.SE.Atl.Fish. 10 (in press).
Armstrong, M.J., P-A- Shelton, R.M. Prosch and W.S. Grant. 1934. Stock assessment and population dynamics of anchovy and pilchard in ICSEAF Division 1.6 in 1982. Colln.scient.Pap.int. Commn.SE.Atl.Fish. 10 (in press).
Babayan, V., P. Kolarov, K. Prodanov, Y.A. Komarov, B. Vaske and A. Wysokinski. 1983. Stock assessment and catch projection for Cape horsemackerel in ICSEAF Divisions 1.3 and 1.4 and 1.5. Colln.scient.Pap.int.Commn. SE.Atl.Fish. 10 (in press).
Badenhorst, A. and A.J. Boyd. 1980. Distributional ecology of the larvae and juveniles of the anchovy Enqraulis capensis Gilchrist in relation to the hydrological environment off South West Africa, 1978/79. Fish.Bull.S.Afr. 13:83-106.
Baird, D. 1975. The South African mackerel: its biology and fishery, S.Afr.Shipp.News Fishg.Ind.Rev. 30:46-51.
Baird, D. 1977. Age, growth and aspects of reproduction of the mackerel. Scomber japonicus in South African waters (Pisces: Scombridae). Zoologica Africana. 12:347-362.
Baird, D. 1978a. Catch composition and population structure of the commercially exploited mackerel Scomber japonicus, 1954-1975. Fish.Bull.S.Afr. 10:50-61.
Baird, D. 1978b. Food of the mackerel Scomber japonicus from Western Cape waters. Fish.Bull.S.Afr. 10:62-68.
Bang, N.D. and W-R-H- Andrews. 1974. Direct current measurements of a shelf-edge frontal jet in the southern Benguela system. J.Mar.Res. 32:405-17.
Batchelor, A.L. 1982. The diet of Cape Gannets, Sula capensis breeding on Bird Island, Algoa Bay. M.Sc. thesis, University of Port Elizabeth. 35 pp.
Botha, L. 1970. S.A. trawlfish landings from 1955 to 1968 with special reference to the hake. S.Afr.Shipp.News Fishg.Ind.Rev. 25(1):69-77.
Botha, L. 1971. Growth and otolith morphology of the Cape hakes Merluccius capensis. Cast. and M. paradoxus. Franca. Investl.Rep.Div.Sea Fish. S. Afr. 97:1-32.
Botha, L. 1980. The biology of the Cape hakes Merluccius capensis Cast. and M. paradoxus Franca in the Cape of Good Hope area. Ph.D. thesis. University of Stellenbosch. 182 pp.
Boyd, A.J. 1979. A relationship between sea surface temperature variability and anchogy Engraulis capensis recruitment off South West Africa. 1970-1978. Fish.Bull.S.Afr. 12:80-84.
Butterworth. D.S. 1980. The value of catch-statistics-based management techniques for heavily fished pelagic stocks, with special refeence to the recent decline of the Southwest African pilchard stock. Colln.scient.Pap.int.Commn.SE. Atl.Fish. 2:69-84.
Buys, M.E.L. 1959. The South African pilchard (Sardinops ocellata) and maasbanker (Trachurus trachurus). Hydrographical environment and the commercial catches. 1950-57. Investl.Rep. Div.Fish.S.Afr. 37:1-176.
Caughley, A. 1970. Eruption of ungulate populations, with emphasis on Himalayan thar in New Zealand Ecology. 51:53-72.
Chalmers, D.S. 1976. Weight conversion factors, length/weight relationships and annual landings of some South African trawl-caught fish. Fish.Bull.S.Afr. 8:1-4.
Cooper, J. R.K., P.A. Shelton and R.J.M. Crawford. 1982. Distribution, population size and conservation of the Cape cormorant Phalacrocarax capensis. Fish.Bull.S.Afr. 16:121-143.
Cram, D.L. 1977. Research and management in southeast Atlantic pelagic fisheries. Rep.Calif. coop.oceanic Fish.Invest. 19:33-62.
Crawford, R.J.M. 1979. Implications of recruitment, distribution and availability of stocks for management of South Africa's Western Cape purse-seine fishery. Ph.D. thesis. University of Cape Town. 467 pp.
Crawford, R.J.M. 1980a. Seasonal patterns in South Africa's Western Cape purse-seine fishery. J.Fish.Biol. 16:649-664.
Crawford, R.J.M. 1980b. Occurrence and distribution of lantern-fish Lampanyctodes hectoris catches in the South African purse-seine fishery. 1968-1976. Fish.Bull.S.Afr. 13:111-136.
Crawford, R.J.M. 1980c. Some biological parameters for six contributors to South Africa's Western Cape purse seine fishery. Colln.scient.Pap.int.Commn.SE.Atl.Fish. 7:85-91.
Crawford, R.J.M. 1981a. Catch per standard boat-day and deployment of effort in the South African purse-seine fishery, 1964-1976. Investl.Rep.Sea Fish.Inst.S.Afr. 122:1-24.
Crawford, R.J.M. 1981b. Distribution, availability and movements of round herring Etrumeus teres off South Africa. 1964-1976. Fish.Bull.S.Afr. 14:141-181.
Crawford, R.J.M. 1981c. Distribution and availability of horse mackerel Trachurus trachurus off South Africa, 1964-1976. Fish.Bull.S.Afr. 15:1-42.
Crawford, R.J.M. 1981d. Distribution, availability and movements of mackerel Scomber japonicus off South Africa. 1964-1976. Fish.Bull.S.Afr. 15:123-166.
Crawford, R.J.M. 1981e. Distribution, availability and movements of anchovy Engraulis capensis off South Africa. 1964-1976. Fish.Bull.S.Afr. 14:51-94.
Crawford, R.J.M. 1981f. Distribution, availability and movements of pilchard Sardinops ocellata off South Africa, 1964-1976. Fish.Bull.S.Afr. 14:1-46.
Crawford, R.J.M. and P.A. Shelton. 1978. Pelagic fish and seabird interrelationships off the coasts of South West and South Africa. Biol.Conserv. 14:85-109.
Crawford, R.J.M. and P.A. Shelton. 1982. Population trends for some southern African seabirds related to availability. In Proceedings of the Symposium on Birds of the Sea and Shore. (J. Coopper, ed.) Cape Town. 1979. Cape Town: Southern African Seabird Group.
Crawford, R.J.M., O.M. Centurier-Harris, G.H.L. Wingate and B.D. Kriedmann. 1978. Revision of species and age composition of landings in the South African purse-seine fishery, 1950 to 1976. Fish.Bull.S.Afr. 10:69-88.
Crawford, R.J.M., P.A. Shelton and L. Hutchings. 1980. Implications of availability, distribution and movements of pilchard (Sardinops ocellata) and anchovy (Enqraulis capensis) for assessment and management of the South African purse-seine fishery. Rapp.P.-v.Réun.Cons.int.Explor.Mer. 177:355-373.
Cruickshank. R.A., J. Cooper and I. Hampton. 1980. Extension to the geographical distribution of pelagic goby Sufflogobius bibarbatus off South West Africa and some mensural and energetic information. Fish.Bull.S.Afr. 13:77-82.
Csirke, J. 1980. Recruitment in the Peruvian anchovy and its dependence on the adult population. Rapp.P.-v.Réun.Cons.int.Explor.Mer. 177:307-313.
Cushing, D.H. 1969. Upwelling and fish production. FAO Fish.Tech.Pap. 84:1-40.
Daan, N. 1980. A review of replacement of depleted stocks by other species and the mechanisms underlying such replacement. Rapp.P.-v.Réun.Cons.int.Explor.Mer. 177:405-421.
Davies, D.H. 1955. The South African pilchard (Sardinops ocellata): bird predators, 1953-1954. Investl.Rep.Div.Fish.Un.S.Afr. 18:1-32.
Davies, D.H. 1956. The South African Pilchard (Sardinops ocellata) and maasbanker (Trachurus trachurus): bird predators, 1954-1955. Investl. Rep.Div.Fish.Un.S.Afr. 23:1-40.
Davies, S.L., G.G. Newman and P.A. Shelton. 1981. A review of the South African multispecies pelagic fishery in ICSEAF Division 1.6 in 1980. Colln.scient.Pap.int.Commn.SE.Atl.Fish. 8:51- 74.
De Jager, B.v.D. 1955. The South African pilchard (Sardinops ocellata). The development of the snoek (Thyrsites atun) a fish predator of the pilchard. Investl.Rep.Div.Fish.Un.S.Afr. 19:1-11.
De Jager, B.v.D., C.S.de V. Nepgen and R.J. Van Wyk 1963. A preliminary report on South African west coast tuna. Investl.Rep.Div.Sea Fish.S.Afr. 47:1-40.
De Villiers, G. 1977. A preliminary review of trends in South African catches and abundance of horse mackerel (Trachurus trachurus capensis) Colln.scient.Pap.int.Commn.SE.Atl.Fish. 4:99-104.
Draganik, B. 1977. Horse mackerel (Trachurus trachurus, L.) fished in the Southeast Atlantic. Colln.scient.Pap.int.Commn.SE.Atl.Fish. 4:25-66.
Duncan, C.P. and J.H. Nell. 1969. Surface currents off the Cape coast. Investl.Rep.Div.Sea Fish.S.Afr. 76:1-19.
Du Plessis, C. G. 1959. The South African pilchard (Sardinops ocellata). Trends in the pilchard fishery of the Union of South Africa. 1943-58. Investl.Rep.Div.Fish.S.Afr. 38:1-28.
Furness, R.W. and J. Cooper. 1982. Interactions between breeding seabird and pelagic fish populations in the Southern Benguela Region. Mar.Ecol.Prog.Ser. 8:243-250.
Geldenhuys, N.D. 1973. Growth of the South African maasbanker Trachurus trachurus Linnaeus and age composition of the catches, 1950-1971. Investl.Rep.Sea Fish.Brch.S.Afr. 101:1-24.
Geldenhuys, N.D. 1978. Age determination of the South African round herring Etrumeus micropus and length and age composition of the commercial catches, 1965-1973. Investl.Rep.Sea Fish.Brch. S.Afr. 115:1-16.
Hairston, N.G., F.E. Smith and L.B. Slobodkin. 1960. Community structure, population control and competition. Am.Nat. 94:421-425.
Harris, T.F.W. and L.V. Shannon. 1979. Satellite-tracked drifter in the Benguela Current System. S.Afr.J.Sci. 75:316-317.
Hutchings, L. and B. Nelson. 1983. The influence of environmental factors in the Cape pelagic fishery SANCOR 5th National Oceanographic Symposium. Grahamstown, January 1983. Symp.S.Afr.Coun.scient.ind.Res. S.288:BIII.
Hutchinson, G.E. 1950. Survey of contemporary knowledge of biogeochemistry. 3. The biogeochemistry of vertebrate excretion. Bull.Am.M.Nat.Hist. 96:1-554.
lies, T.D. 1980. Environmental pressure and intra- and inter-year-class competition as determinants of year-class size. Rapp.P.-v.Réun.Cons.int.Explor.Mer. 177:315-331.
King, D.P.F. 1977a. Influence of temperature, dissolved oxygen and salinity on incubation and early larval development of the South West African pilchard Sardinops ocellata. Investl.Rep.Sea Fish.Brch.S.Afr. 114:1-35.
King, D.P.F. 1977b. Distribution and relative abundance of eggs of the South West African pilchard Sardinops ocellata and anchovy Engraulis capensis, 1971/72. Fish.Bull.S.Afr. 9:23-31.
King, D.P.F. and P.R. Macleod. 1976. Comparison of the food and the filtering mechanism of pilchard Sardinops ocellata and anchovy Engraulis capensis off South West Africa, 1971-1972. Investl.Rep.Sea Fish.Brch.S.Afr. 111:1-29.
King, D.P.F., M.J. O'Toole and A.A. Robertson. 1977. Early development of the South African maasbanker Trachurus trachurus at controlled temperatures. Fish.Bull.S.Afr. 9:16-22.
King, D.P.F., A.A. Robertson and P.A. Shelton. 1978. Laboratory observations on the early development of the anchovy Engraulis capensis from the Cape Peninsula. Fish. Bull. S.Afr. 10:37-45.
Kondo, K. 1980. The recovery of the Japanese sardine - the biological basis of stock-size fluctuations. Rapp.P.-v.Réun.Cons.int.Explor.Mer. 177:332-354.
Le Clus, F. 1979a. Fecundity and maturity of anchovy Engraulis capensis off South West Africa. Fish.Bull.S.Afr. 11:26-38.
Le Clus, F. 1979b. Dry mass of yolked oocytes of the South West African pilchard Sardinops ocellata in relation to maturity stages and spawning cycles, 1972-1974. Investl.Rep.Sea Fish.Brch.S.Afr. 119:1-29.
Le Clus, F. and I. Kruger. 1982. Time and space distribution of temperature, salinity, plankton and fish eggs off South West Africa in 1980/81 - A preliminary data report. Colln.scient.Pap. int.Commn.SE.Atl.Fish. 9:121-146
Le Clus, F. and Y.C. Melo. 1982. Assessment of the the 1981 anchovy population in ICSEAF Divisions 1.3, 1.4 and 1.5. Colln.scient.Pap.int.Commn.SE.Atl.Fish. 9:147-161.
Le Clus, F. 1983. Virtual Population Analysis of the anchovy stock off South West Africa. 1972-1982. Colln.scient.Pap.int.Commn.SE.Atl.Fish. 10 (in press).
Le Clus, F. and R.M. Thomas. 1982. Assessment of the the 1981 pilchard population in ICSEAF Divisons 1.3, 1.4, and 1.5 Colln.scient.Pap.int.Commn. SE. Atl.Fish. 9:163-176.
Lleonart, J., B. Mortales and A. Castellon. 1983. Estudio preliminar de la diferenciacion de stocks de Merluccius capensis en aguasde Namibia (18-285). Colln.scient.Pap.int.Commn.SE.Atl.Fish. 10 (in press).
MacCall, A.D. 1974. The mortality rate of Engraulis mordax in southern California. Rep.Calif. coop.oceanic Fish.Invest. 17:131-135.
MacCall, A.D. 1980a. Population models for the northern anchovy (Engraulis mordax). Rapp.P.-v.Réun.Cons.int.Explor.Mer. 177:292-306.
MacCall, A.D. 1980b. The consequences of cannibalism in the stock-recruitment relationship of planktivorpus pelagic fishes such as Engraulis. pp. 201-220. In Workshop on the effects of environmental variation on the survival of larval pelagic fishes. (G.D. Sharp, ed.) IOC Workshop Report No. 28. IOC/UNESCO, Paris.
MacCall, A.D. 1982. Longshore variability patterns of larval anchovy abundance.
Matthews, J.P. 1961. The pilchard off South West Africa (Sardinops ocellata) and the maasbanker (Trachurus trachurus). Bird predators, 1957-1958. Investl.Rep.mar.res.Lab. SW.Afr. 3:1-35.
Matthews, J.P. 1964. The pilchard of South West Africa (Sardinops ocellata). Sexual development, condition factor and reproduction. Investl.Rep.mar.res.Lab. SW.Afr. 10:1-96.
Monteiro, R. 1962. Sobre a ocorrencia de Sardinops ocellata (Pappe) em aguas centro-Angolanasa. Notas mimeogr. Centre Biol.Piscat. (Lisboa). 25:1-12.
Nawratil, 0. 1960. The pilchard of South West Africa (Sardinops ocellata). Age studies and age composition of Sardinops ocellata in the commercial catches, 1952-1958 and a new method for the determination of the age of Sardinops ocellata. InvestI.Rep.mar.res.Lab. SW.Afr. 2:1-43.
Nepgen, C.S. de V. 1970. Exploratory fishing for tuna off the South African west coast. Investl.Rep.Div.Sea Fish.S.Afr. 87:1-26.
Nepgen, C.S. de V. 1979a. The food of the snoek Thyrsites atun. Fish. Bull. S.Afr. 11:39-42.
Nepgen, C.S. de V. 1979b. Trends in the line fishery for snoek Thyrsites atun off the south-western Cape and in size composition, length-weight relationship and condition. Fish.Bull.S.Afr. 12:35-43.
Newman, G.G. 1970a. Stock assessment of the pilchard Sardinops ocellata at Walvis Bay, South West Africa. Investl.Rep.Div.Sea Fish.S.Afr. 85:1-13.
Newman, G.G. 1970b. Migration of the pilchard Sardinops ocellata in southern Africa. Investl.Rep. Div.Sea Fish.S.Afr. 86:1-6.
Newman, G.G. 1977. The living marine resources of the southeast Atlantic. FAO Fish.Tech.Pap. 178:1-59.
Newman, G.G. and O.M. Centurier-Harris. 1977. Stock estimates of the Cape horse mackerel (Trachurus trachurus) in the Cunene and Cape Cross Divisions. Colln.scient.Pap.int.Commn. SE.Atl.Fish. 4:67-75.
Newman, G.G. and R.J.M. Crawford. 1980. Population biology and management of mixed-species pelagic stocks off South Africa. Rapp.P.-v.Réun.Cons.int.Explor.Mer. 177:279-291.
Newman, G.G., R.J.M. Crawford and O.M. Centurier-Harris. 1976a. Stock assessment of the hake (Merluccius capensis and Merluccius paradoxus) on the Cunene and Cape Cross fishing grounds (ICSEAF Divisions 1.3 and 1.4). Colln.scient.Pap.int.Commn.SE.Atl.Fish. 3:125-132.
Newman, G.G., 1976b. Stock assessment of the hake (Merluccius capensis and Merluccius paradoxus) on the Orange River fishing grounds (ICSEAF Division 1.5) Colln.scient.Pap.int.Commn.SE.Atl. Fish. 3:133-140.
Newman, G.G., 1976c. Stock assessment of the hake (Merluccius capensis and Merluccius paradoxus) on the Cape of Good Hope fishing grounds (ICSEAF Division 1.6). Colln.scient.Pap.int.Commn. SE.Atl.Fish. 3:141-150.
Newman, G.G., 1979. Fishing effort and factors affecting vessel performance in the South African purse-seine fishery, 1964-1972. Investl.Rep.Sea Fish.Brch.S.Afr. 120:1-34.
Nielsen, G.S. and A. Jensen. 1957. Primary oceanic production: the autotrophic production of organic matter in the ocean, Galathea Rep. 1:49-135.
O'Toole, M.J. 1977. Investigations into some important fish larvae in the South East Atlantic in relation to the hydrological environment. Ph.D. thesis, University of Cape Town.
O'Toole, M.J. 1978a. Aspects of the early life history of the hake Merluccius capensis Castelnau of South West Africa. Fish.Bull.S.Afr. 10:20-36.
O'Toole, M.J. 1978b. Development, distribution and relative abundance of the larvae and early juveniles of the pelagic goby Sufflogobius bi barbatus (von Bonde) off South West Africa, 1972- 1974. Investl.Rep.Sea Fish.Brch.S.Afr. 116:1-28.
O'Toole, M.J. 1980. Seasonal distribution of temperature and salinity in the surface waters off South West Africa. 1972-1974. Investl.Rep.Sea Fish. Inst.S.Afr. 121:1-25.
O'Toole, M.J. and D.P.F. King. 1974. Early development of the round herring Etrumeus teres (de Kay) form the south east Atlantic Vie Milieu. fasc. 3(Ser.A). 24:443-452.
Payne, A.I.L. and R.W. Leslie. 1983. The state of the hake stock in ICSEAF Divisions 1.6 and 2.1/2.2 assessed from information collected up to 1981. Colln.scient.Pap.int.Commn.SE.Atl.Fish. 10 (in press).
Radovich, J. 1979. Managing pelagic schooling prey species, pp. In Predator-prey systems in fishery management. (H. Clapper, ed.) Sport Fishing Institute, Washington, D.C.
Rand, R.W. 1952. Guano enterprise in South West Africa. Ostrich. 23(3):169-185.
Rand, R.W. 1959a. The Cape fur seal (Arctocephalus pusillus): Distribution, abundance and feeding habits off the south-western coast of the Cape Province. Investl.Rep.Div.Fish.Un.S. Afr. 34:1-75.
Rand, R.W. 1959b. The biology of guano-producing seabirds: The distribution, abundance and feeding habits of the Cape gannet, Morus capensis, off the south-western coast of the Cape Province. Investl.Rep.Div.Fish.Un.S.Afr. 39:1-36.
Rand, R.W. 1960a. The biology of guano-producing seabirds: The distribution, abundance and feeding habits of the Cape penguin, Spheniscus demersus, off the south-western coast of the Cape Province. Investl.Rep.Div.Fish.Un.S.Afr. 41:1-28.
Rand, R.W. 1960b. The biology of guano-producing seabirds: 3. The distribution, abundance and feeding habits of the cormorants Phalacrocoracidae off the south-western coast of the Cape Province. Investl.Rep.Div.Fish.Un.S.Afr. 42:1-32.
Randall, R. and G.J.B. Ross. 1979. Increasing populations of Gape gannets on Bird Island, Algoa Bay, and observations on breeding success. Ostrich. 50:168-175.
Ricker, W.E. 1954. Stock and recruitment. J.Fish.Res.Bd.Canada. 11(5):559-623.
Ricker, W.E. 1963. Big effects from small causes: Two examples from fish population dynamics. J.Fish.Res.Bd.Canada. 20(2):257-264.
Saville, A. 1980. Discussions and conclusions of the symposium on the biological basis of pelagic fish stock management. Rapp.P.-v.Réun.Cons.int.Explor.Mer. 177:513-517.
Schülein, F.H. 1971. Rekruteringstudies oor die Suidwes-Afrikaanse sardyn, Sardinops ocellata, 1954-1968. M.sc. thesis, University of Stellenbosch. 62 pp.
Schülein, F.H. 1973. Recruitment studies on the South West African pilchard, 1958-1972. S.Af.natn.oceanogr.Symp.Abstr. Cape Town: 29.
Schülein, F.H., D.S. Butterworth and D.L. Cram. 1978. An assessment of the Southeast Atlantic pilchard population in ICSEAF Divisions 1.4 and 1.5, 1953-1977. Colln.scient.Pap.int.Commn. SE.Atl.Fish. 5:35-44.
Schumann, E.H., L.-A. Perrins and I.T.Hunter. 1982. Upwelling along the South Coast, of the Cape Province. S.Afr.J.Sci. 78:238-242.
Shannon, L.V. 1966. Hydrology of the south and west coasts of South Africa. Investl.Rep.Div.Sea Fish.S.Afr. 58:1-62.
Shannon, L.V. 1976. Introductory notes on the abiotic environment of the large South African mackerel. SANCOR/ECOR/LSSA/NPES 1st Interdisciplinary Conference on Marine and Fresh-water Research in Southern Africa, Port Elizabeth, July 1976. Symp.S.Afr.Coun.scient.ind.Res. S122(Fiche 8):B12-D4.
Shannon, L.V. and F.P. Anderson. 1982. Applications of satellite ocean colour imagery in the study of the Benguela Current system. S.Afr.J.Photogrammetry Remote Sensing Cartography. 13(3):153-169.
Shannon, L.V. and J.L. Henry. 1983. Phytoplankton primary production in the Benguela Current from ship and satellite measurements. SANCOR 5th National Oceanographic Symposium, Grahamstown. January 1983. Symp.S.Afr.Coun.scient.ind.Res. S.288:B18.
Shelton, P.A. 1982. The South African multispecies pelagic fishery in ICSEAF Division 1.6 in 1981. Colln.scient.Pap.int.Commn.SE.Atl.Fish. 9:327-336.
Shelton, P.A. and L. Hutchings. 1982. Transport of anchovy, Engraulis capensis Gilchrist, eggs and early larvae by a frontal jet, J.Cons.int.Explor.Mer. 40:185-198.
Silliman, R.P. 1969. Population models and test populations as research tools. Bioscience. 19:524- 528.
Skud, B.E. 1982. Dominance in fishes: The relationship between environment and abundance. Science. 216:144-149.
Smith, P.E. 1972. The increase in the spawning biomass of northern anchovy, Engraulis mordax. Fish.Bull. 70:849-874.
Smith, P.E. 1978. Biological effects of ocean variability, time and space scales of biological response. Rapp.P.-v.Réun.Cons.int.Explor.Mer. 173:117-122.
Smith, P.E. and R.W. Eppley. 1982. Primary production and the anchovy population in the Southern California Bight: comparison of time series. Limnol.Oceanogr. 27(1)1-17.
Stander, G.H. 1963. The pilchard of South West Africa (Sardinops ocellata). Temperature: its annual cycles and relation to wind and spawning. Investl.Rep.mar.Res.Lab.SW.Afr. 9:1-57.
Stander, G.H. 1964. The pilchard of South West Africa (Sardinops ocellata). The Benguela Current off South West Africa. Investl.Rep.mar.Res.Lab.SW.Afr. 12:1-43.
Stander, G.H. and A.H.B. de Decker. 1969. Some physical and biological aspects of an oceanographic anomaly off South West Africa. Investl.Rep.Div.Sea Fish.S.Afr. 81:1-46.
Stander, G.H. and P.J. le Roux. 1968. Notes on fluctuations of the commercial catch of the South African pilchard (Sardinops ocellata) 1950-1965. Investl.Rep.Div.Sea Fish.S.Afr. 65:1-14.
Thomas, R.M. 1979. Age studies on the South West African pilchard (Sardinops ocellata) in ICSEAF Divisions 1.4 and 1.5. Colln.scient.Pap.int.Commn.SE.Atl.Fish. 6:191-203.
Thomas, R.M. 1983. Catch, age, growth and stock assessment of the SWA pilchard, Sardinops ocellata, in Divisons 1.3, 1.4 and 1.5 for 1982. Colln.scient.Pap.int.Commn.SE.Atl.Fish. 10 (in press).
Thomas, R.M., J.D. Hewitson and H. Bohme. 1982. Suggested explanation for a shift in catch positions of juvenile anchovy off South West Africa in 1978. Fish.Bull.S.Afr. 16:95-97.
Troadec, J.-P., W.G. Clark and J.A. Gulland. 1980. A review of some pelagic fish stocks in other areas. Rapp.P.-v.Réun.Cons.int.Explor.Mer. 177:252-277.
![]()
![]()
![]()