![]()
![]()
![]()
Compilado por
Suzana Anita Saccardo
Instituto de Pesquisa e Desenvolvimento Pesqueiro
Superintendencia do Desenvolvimento da Pesca
PDP/SUDEPE Av. Indianápolis, 1123
São Paulo (SP)
BRASIL
Summary
The results obtained from the studies of the biology and stock assessment of Sardinella brasiliensis that inhabit the littoral zone of southeast Brasil (22°-28°S) are presented, as provided by the Permanent Group for the Studies of Sardines.
Related to reproduction, the analysis of frequencies of maturity stages and gonadal development indices show that peak spawning occurs in summer, particularly in the months of January and February. The average size at onset of sexual maturity was estimated to be 16.7 cm and the average fecundity was 23,000 eggs per adult female.
Age determination for the study of growth, obtained by reading otolith rings, showed that the catch comprises principally of individuals with two or three classes of rings, in which the most frequent group has four rings and a corresponding size of about 18 cm. The value of the growth constant "k" is also discussed. Total landings of S. brasiliensis have diminished drastically in recent years, passing through a maximum of 228,000 tons in 1973 to 90,000 tons in 1982. The MSY calculated by the application of the general production model was 170,000 tons.
Natural mortality was estimated at 0.74 using the method of Pauly (1980) and then this was applied to preliminary cohort analyses. The results of these analyses are also discussed.
Evaluations of total biomass derived from hydroacoustic studies made during the period 1974-1980 yielded values ranging from 142,000 to 414,000 tons.
The available information for stock diagnoses do not appear to explain conclusively the reasons that have provoked the decline in catches. Various hypotheses about this topic are discussed.
INTRODUCTION
Entre las especies pelágicas de importancia económica del Brasil, la sardina es aquella que está sujeta al mayor esfuerzo de pesca por parte de la flota comercial. La sardina se captura en gran cantidad en el área comprendida entre los 22°S y 28°S (Cabo de São Tomé a Cabo Santa Marta Grande), a una profundiad máxima de operación de aproximadamente 70m. (Fig. 1).
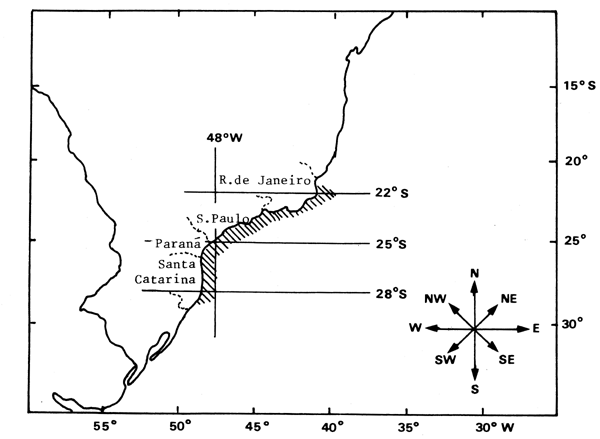
Fig.1 - Area de pesca de 1a sardina (Sardinella brasiliensis)
Existe un gran volumen de datos y un gran número de trabajos sobre esta especie, iniciados en 1950. En vista de la importancia de este recurso la SUDEPE (Superintendencia del Desarrollo de la Pesca), promueve, desde 1974, reuniones anuales del Grupo Permanente de Estudios sobre la Sardina - GPE (*)-, con la finalidad de actualizar las informaciones disponibles sobre la situación del recurso y recomendar medidas de administración.
En 1979, durante la reunión del Grupo Permenente, se constató que los estudios eran dispersos y existían diferencias en la metodología de los muestreos y en los criterios de obtención de datos por lo cual no era posible tener un cuadro sinóptico satisfactorios de los conocimientos de la especie. Por este motivo se creó un Programa de Estudios Biólogicos sobre Sardina (PIEBS) que está siendo llevado a cabo por investigadores del GPE y que tiene por objectivo el seguimiento continue de la población de sardina, los cambios en su estructura y las variaciones de los parámetros biológicos. Este programa de estudios analiza las informaciones de caracter biológico y las relaciona con los datos de captura y esfuerzo de pesca con la finalidad de evaluar la situación del recurso y contribuir a una adecuada administración pesquera.
Durante el 2o Encuentro del Grupo de Trabajo y Entrenamiento sobre Evaluación de Stocks (GTT) de 1981 promovido por la FAO, se revisaron los trabajos existentes sobre la especie y se analizaron los más recientes datos disponibles, llegándose a hacer una estimacion del nivel de explotación de la población y el estado de la pesca (PDP/SUDEPE, 1981a).
La presente contribución pretende documentar los últimos resultados sobre biología y evaluación de stock de sardina según fueron presentados en la última reunión del GPE y en el GTT.
* El Grupo Permanente de Estudios sobre la Sardina (GPE) está constituido por científicos de la SUDEPE (Ministerio de Agricultura), Instituto de Pesca (Secretaría de Agricultura del Estado de San Pablo) e Instituto Oceanográfico (Universidad de San Pablo).
BIOLOGIA
Estructura de la especie
Existen evidencias de que la especie no se comporta homogéneamente a lo largo de su área de explotación en la costa brasilera.
Estudios basados en el análisis de la distribución de frecuencia de tallas (Richardson et al., 1959; Richardson y Sadowski, 1960) y del período y lugar del desove (Rossi-Wongtschowski, 1977; Projecto Megalosar, 1980) sugieren la existencia de dos grupos diferenciados: uno entre 22° - 25° S y otro entre 26° - 28°S con épocas de desove diferentes. Los jóvenes de cada uno de estos grupos penetrarían en la region esturino-lagunar de Cananéia (25° 01'S) en épocas distintas: el grupo septentrional en abril y el meridional en octubre saliendo en mayo (Richardson y Sadowski, 1960). Algunos estudios bioquímicos y morfológicos contribuyen a apoyar esta observación.
En estudios bioquímicos se constató que los patrones electroforéticos de las proteínas totales del cristalino de sardina pueden ser usadas como marcas naturales para el estudio de la estructura de la especie (Vazzoler y Ngan, 1976).
Por otra parte, los datos de proporciones corporales y carácteres merísticos de 3351 ejemplares capturados entre 22° - 28°S en el período de 1970 a 1974 (Rossi-Wongtschowski, 1978) analizados aisladamente no permiten caracterizar los individuos provenientes de cada grupo debido a la gran variabilidad individual causada por la enorme plasticidad de la especie, asociada a los movimientos de los individuos en el sentido N-S e viceversa, resultando en un mosaico. Sin embargo, el análisis de las variaciones del número de branquispinas del primer arco branquial izquierdo permitieron sustentar algunas hipótesis sobre la existencia de estos grupos.
En relación a la morfología de los otolítos se constataron diferencias estadísticamente significativas en las relaciones entre la longitud del rostro y antirostro; y en los pesos de los otolítos con la talla de los ejemplares (Rossi-Wongtschowski et al., 1982). El análisis de las características de los otolítos (tipo de núcleo y forma) de los ejemplares capturados en diferentes áreas y épocas del año sugiere la existencia de mezcla de los grupos entre sí.
Están aún en fase de estudio los análisis del tipo bioquímico y las proporciones corporales como medio para caracterizar morfológicamente los posibles grupos (Projecto Megalosar, 1980, 1981).
Estructura de la población:
Proporción Sexual: generalmente existe una mayor frecuencia de hembras que de machos durante el transcurso del año, con excepción de la época del desove (verano), cuando las frecuencias entre ambos sexos tienden a igualarse (Matsuura, 1977; PDP/SUDEPE, 1982 b).
Composición por tamaños: Se observa heterogeneidad en la composición de los individuos por clase de talla a to largo del área de distribución (Projecto Megalosar, 1980); la tabla l muestra la amplitud de talla y las clases más frecuentes en la pesquería comercial.
| región | amplitud (mm) | clase más frecuente (mm) |
|---|---|---|
| 22o - 23oS (Cabo Frío) | 150-260 | 240;250 |
| 23o - 24oS (Angra dos Reis; Santos) | 130-240 | 200 |
| 25o - 26oS (Cananéia; Paranaguá) | 170-210 | 190 |
| 26o - 28oS (Itajaí; Florianópolis) | 120-230 | 180 |
Considerando también los datos obtenidos de la pesca artesanal, la cual incluye ejemplares con tallas inferiores a 170 mm, se verifica que el reclutamiento a la pesca ocurre principalmente en los meses de agosto y septiembre.
La mayor frecuencia de individuos pequeños se observa en la región estuarino-lagunar entre los 25o y 26oS.
Composición por clases de edad: la pesca incide principalmente sobre dos o tres clases de edad, encontrandose una composición diferencial a lo largo del área: en la región entre 22o-23oS predominan dos individuos más viejos y en la región 25o-26os los más jóvenes (Projecto Megalosar, 1981).
Se observe que la especie comienza a ser capturada cuando posee 3 anillos en sus otolitos, permaneciendo disponible para la pesca durante un periodo correspondiente a la formación de 3 o 4 anillos más (PDP/SUDEPE, 1982 b).
Reproducción
Madurez gonadal: Estudios histológicos de los ovarios permitieron establecer las caracteristícas del desarrolo gonadal y una escala de madurez adecuada (ISAAC-NAHUM et al., 1983).
Considerando las características macro y microscópicas de las gónadas de hembras adultas se verifica que la mayor parte de los individuos permanece en fase de reposo gonadal de abril a septiembre, madurando en seguida rápidamente; el estadio de pre-desove es alcanzado en noviembre por los individuos de Rio de Janeiro y posteriortnente por aquellos de San Pablo y Santa Catarina, lo cual puede sugerir que el desove se inicia más tempranamente en la region Norte. (Fig.2).
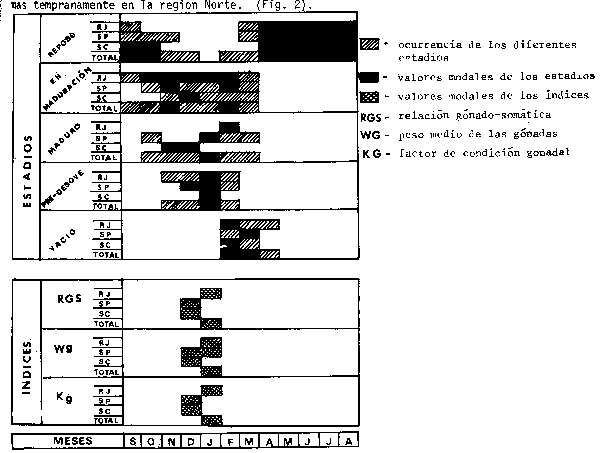
Fig.2.- Cuadro comparativo señalando los meses en que aparecen cada uno de los estadios de madurez y también sus mayores frecuencias así como los valores modales de los índices de madurez para los Estados de Río de Janeiro, San Pablo y Santa Catarina y para el área global (período Septiembre 1981 - Agosto 1982).
En el análysis de la relación gónado-somótica, peso y factor de condición gonadales se notó que estos valores aumentan con el tamaño del pez, o sea, individuos mayores poseen gónadas mayores y por tanto índices más altos que los ejemplares menores; sin embargo, parece que no hay diferencias acentuadas en relación con la época de desove entre las diferentes clases de talla, significando que todos los peces desovan aproximadamente en el mismo período (PDP/SUDEPE, 1982 b). Los valores modals de estos indicardores de desarrollo gonadal ocurren entre los meses de diciembre y enero (Fig.2).
Fecundidad.- La fecundidad real media, obtenida para 16 hembras según el método de Ciechomski et al., (1973), y considerando apenas los ovocitos que pertenecen a la última moda de la distribución de frecuencia de sus diámetros, fue estimada en 23 000 ovocitos por hembra (PDP/SUDEPE, 1982 b).
Talla media del inicio de la primera madurez gonadal.- El valor encontrado para ambos sexos y para el area global fue 167 mm (PDP/SUDEPE, 1982 b).
Tipo de desove.- A pesar de que las curvas de distribución de diámetros de ovocitos intraovaricos se presentan bimodales, el comportamiento de estas de curvas muestran un patrón de desove total, una vez que los ovocitos representados por la última moda crecen, aumentando su diámetro, mientras que los otros, menos diferenciados, permanecen estables. Este hecho, reforzado por la presencia de ovocitos que pertenecen a la primera moda en los ovarios vacious, hace pensar que estas células estarían destinadas a ser reabsorbidas por el ovario y no a evolucionar y ser eliminadas después de la liberación del primer lote (PDP/ SUDEPE, 1982 b).
Epoca y área de desove.- Rossi-Wongtschowski (1977) y Projecto Megalosar (1980), sugieren la existencia de dos areas principales de desova: una entre 22o - 25oS, de octubre a junio (con picos en diciembre-enero y marzo-abril), y otra entre 26o - 28oS, de diciembre a enero (con pico en enero).
Sin embargo hemos encontrado grandes variaciones de año para año, probablemente debido a cambios en las condiciones ambientales. Los resultados más recientes indican que hay reproducción en toda el área de distribución y un período de desove más restricto entre noviembre y abril con picos en enero-febrero (PDP/SUDEPE, 1982 b). Según Matsuura (1977) el desove ocurre con mayor intensidad entre las isobatas de 5l a 100 m.
Crecimiento
Los estudios sobre crecimiento de S. brasiliensis en el Brasil no siempre abarcaron toda el área de mayor densidad de la distribución de la especie. Estos ban sido realizados a través del análysis de las distribuciones de frecuencia de tallas (Richardson et al., 1959; Santos y Fratzen, 1965; Matsuura, 1977; Rijavec et al., 1977), lectura de anillos de escamas (Richardson et al, 1959) y otolTtos (Vazzoler, et al., 1982a, 1982b; Rossi-Wongtschowski et al., 1982; (PDP/SUDEPE, 1982 b).
Fue constatado que los otolitos son más adecuados para la lectura que las escamas (Rossi-Wongtschowski et al., 1982). y se comprobó la periodicidad y época de formación de los anillos de crecimiento (Vazzoler et al., 1982a).
La longevidad de los individuos defiere a lo largo del área de distribución: se encontraron peces hasta 7 años en las regiones entre los 22o-25oS, hasta 6 a-os entre 26o - 28oS y en la de 25o - 26oS (región estuarino-lagunar) solamente individuos hasta 4 años (Rossi-Wongtschowski et al., 1982; Vazzoler et al., 1982a e b; PDP/SUDEPE, 1982 b).
| región | L (mm) | K | to | W (g) | Autores |
|---|---|---|---|---|---|
| 22o-23oS | 23,5 | 0,54 | -0,24 | 112,0 | Vazzoler et al., 1982 b |
| 24,3 | 0,50 | 0,00 | Santos y Fratzen, 1965 | ||
| 23o-25oS | 24,4 | 0,44 | 0,00 | Matsura, 1977 | |
| 33,5 | 0,87 | 0,00 | Rijavec et al, 1977 | ||
| 22,4 | 0,46 | -0,75 | 100,8 | Vazzoler et al., 1982 b | |
| 25o-26oS | 17,7 | 0,39 | -0,74 | 46,1 | Vazzolaret al., 1982 b |
| 26o-28oS | 22,5 | 0,37 | -1,02 | 88,9 | Vazzolar et al., 1982 b |
Comparando los resultados encontrados para el área 23o-25oS se verifican diferencias entre los parámetros indicados (Fig. 3), las cuales podrían ser debidas a las diferentes metodologías empleadas.
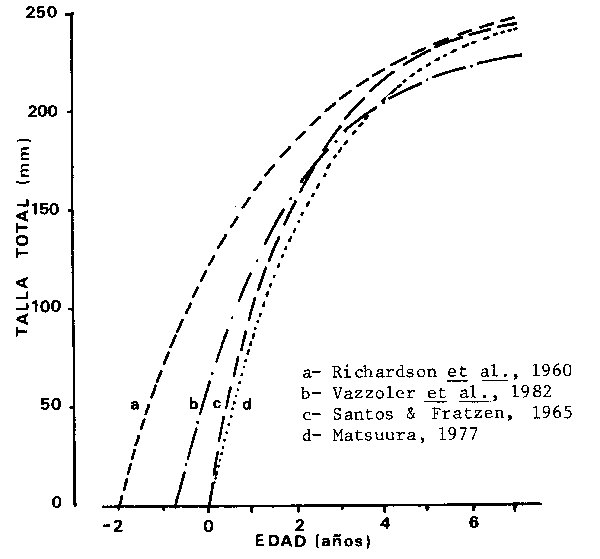
Fig. 3 - Comparación gráfica de las curvas de crecimiento de S. brasiliensis en la región entre 23o-25oS obtenidos por varios autores (adapto de Vazzoler et al., 1982 b)
En los resultados obtenidos por Vazzoler et al. (1982 b), se observaron patrones de crecimiento distintos según sea el área considerada (Fig. 4); los peces de la región estuar no-lagunar presentan crecimiento menor que los de mar abierto y entre estos últimos existe una variación clinal en el sentido Norte-Sur, siendo que los ejemplares que habitan la región más al North presentan crecimiento más intens tanto en talla cino en peso.
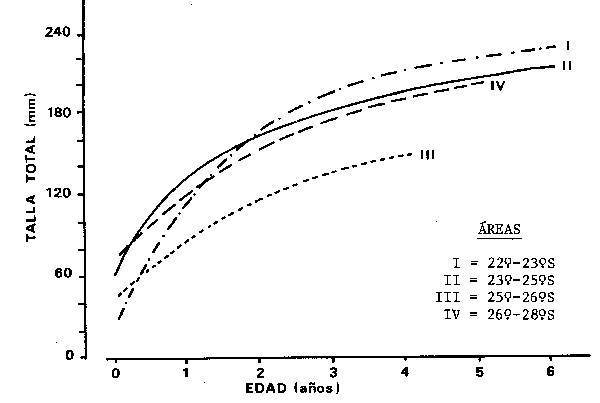
Fig. 4 - Comparación gráfica de las curves de crecimiento en talla de S. brasiliensis para las cuatro regiones estudidas por Vazzolar et al. (1982 b)
EVALUACION DE STOCK
Caracterización de la flota y de la pesquería
La pesca de sardina, inicialmente artesanal, comenzó a desarrollarse en términos industriales en la década del 1940 en los Estados de Río de Janeiro y San Pablo con la mecanización de la flota, y más recientemente en la década del 1960 en el Estado de Santa Catarina(PDP/SUDEPE, 1981a).
Existen actualmente un total de 378 embarcaciones conocidas con el nombre de"traineiras": 88 registradas en el Estado de Santa Catarina, 65 en San Pablo y 225 en Río de Janeiro. La tabla 3 muestra los promedios de las principales características físicas de estas unidades.
| características físicas del barco | SC | SP | RJ |
|---|---|---|---|
| Eslora(m) | 20,8 | 19,6 | 14,1 |
| Capacidad de la bodega (m3) | 51,8 | 36,6 | 26,1 |
| Arqueo bruto (ton.) | 66,5 | 51,7 | 24,6 |
| Edad (años) | 10 | 19 | 19 |
| Número de barcos | 88 | 65 | 225 |
Podemos constatar que los barcos de Santa Catarina (SC) son mayores y más modernos que los de San Pabo (SP) y Río de Janeiro (RJ) (PDP/SUDEPE, 1982a), 1o que probabtemente determina diferencias en el poder de pesca entre estas flotas.
La pesca es realizada durante las noches sin luna; los cardúmenes son localizados por métodos visuales por el proero (vigía) y son posteriormente cercados con una red de cerco que mide cerca de 500-675 m de largo y 40-76 m de altura (Wiefels y Jabtonski, 1979] A continuación, 1os peces son cargados a cubierta con pequeos sacos. Toda la operación dura aproximadamente 3 horas.
Desembarques de sardina
La disponibilidad de sardina para la pesca difiere de año para año y particutarmente de mes a mes, a pesar de no obedecer a un patrón definido de comportamiento (PDP/SUDEPE, 1981a).
Considerando los datos disponibles desde 1964, los desembarques de sardina presentan valores ascendentes hasta 1973, cuando la producción alcanzó su máximo de 228 037 ton, a partir de entonces se observa una tendencia de disminución hasta 1976 (105 053 ton) mantem'endose estable en los cuatro anos siguientes alrededor de las 145 000 ton. En los últimos dos aos las capturas disminuyeron llegando hasta el valor mínimo de 90 000 ton. en 1982 (Fig. 5).
Esfuerzo y abundancia
Analizando la tendencia del esfuerzo aplicado a la especie se nota que a pesar del aumento de la intensidad de pesca, en el período 1964-1973, el índice de abundancia (captura/lance) se mantuvo estable; en los últimos años, después de una brusca caída del esfuerzo (entre 73 y 78), este se mantuvo en el mismo nivel, mientras que la abundancia que aumenta inicialmente, comienza a disminuir a partir de 1978 (Fig. 5).
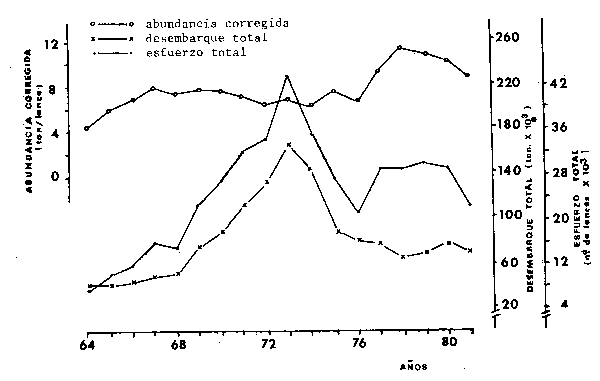
Fig. 5 - Desembarque total (ton,), esfuerzo total estimado (no de lances) y índice de abundancia de San Pablo (corregido), para S. brasiliensis en el período 1964-1981.
La interpretación de estos datos parece incierta, debiéndose resaltar que la medida de esfuerzo empleada puede no ser adecuada para estimar la densidad promedio real de la población. Esto por la modalidad de la pesquería, en que sólo se realiza el lance cuando el cardumen ha sido avistado, por tanto los datos de captura/lance miden más las variaciones en el tamaño del cardumen que la abundancia del recurso (ACMRR, 1976).
Por otro lado, los datos de "numero de viajes" tampoco serían satisfactories ya que debido a los altos costos operacionales de la flota, especialmente en los últimos años, se nota una tendencia a solo realizar viajes cuando hay certeza de la disponibilidad de cardúmenes en determinada área.
Por otro lado, los datos sobre "horas de búsqueda" no parecen confiables, siendo necesario en el presente encontrar una forma de integrar las diversas medidas de esfuerzo para poder determinar con exactitud las oscilaciones de la abundancia.
Consideraciones sobre la crisis de la producción en el. período 1981-1982
Existe una gran preocupación por parte de los administradores y empresarios de la pesca de sardina con respecto a la disminución brusca de las capturas en los últimos dos años.
Para tratar de explicar esta disminución se formularon las siguientes hipótesis:
1o) disminución del poder de pesca o del esfuerzo debido a la salida de algunas embarcciones grandes que fueron transferidas a la pesquería de atunes, o que se retiraron debido a la inviabilidad económica de la pesca.
2o) no disponibilidad del recurso en las áreas de pesca accesibles a la flota comercial.
3o) disminución de la biomasa de sardina por fallas de reclutamiento u otras causas similares.
En relacón con la primera hipótesis cabe mencionar que se esán fomentando mejoras en los equipamientos de los barcos, principalmente para la adquisición de instrumentos de búsqueda y localización de cardúmenes, de manera de aumentar la productividad de cada viaje. Por otro lado se trató de estimar la capacidad real de pesca de la flota operativa a la fecha, con la finalidad de determinar el potencial de producción máxima de la misma. De esta manera, considerando el ao en que se hicieron las mayores capturas (1973) se calculó e1 índice de abundancia máxima en la forma captura/barco, y sabiendo el número de barcos que efectivamente operaron en la pesca durante el año de 1982, se estimó asií un potencial capturable de cerca de 160 000 ton/año. Este valor debe ser entendido apenas como un indicador de potencial de pesca, si la abundancia de sardina fuese igual a la ocurrida en 1973, y en la medida que la relación viaje/barco se mantenga dentro de los valores medios observados en los años anteriores.
Podemos sin embargo concluir que la flota actual a pesar de sus limitaciones técnicas, parece capacitada a capturar un total bastante pró al MSY calculado para el recurso.
Las otras hipótesis permanecen en el campo de la especulación, ya que todavía no existe un seguimiento sistemático de las fluctuaciones de la biomasa del stock desovante y del reclutamiento, ni tampoco de las variaciones de los paráiwtros ambientales. Con todo, se ha observado el estrechamiento del área de mayor densidad de la especie, y en los últimos aos la pesca en los extremes Norte y Sur ha bajado sensiblemente, hecho que podría indicar una disminución en eltamao del stock.
Modelo de producción general
Los datos estadísticos de la pesca de sardina se obtienen por muestreos ya que no es posible cubrir el total de operaciones efectuadas. Conociendo el monto total de los desembarques y la CPUE estimada para una muestra estadísticamente significativa de la flota, obtenemos el esfuerzo de pesca total estimado para cada período. '
Los datos de esfuerzo del período 1964-1973 fueran corregidos según Matsuura (1981) de acuerdo con el aumento del promedio del arqueo bruto de los barcos durante esos años (Tabla 4).
Usando el "lance" como medida de esfuerzo y la CPUE estimada sólo para la flota del Estado de San Pablo (por ser estas estadísticas más confiables), se aplica e1 modelo de Fox (1970) para el cálculo del rendimiento máximo sostenible (MSY) para e1 período 1969-1981.
Los parámetros fueron estimados a través de una regresión funcional debido a las caractenstícas de las variables usadas (Ricker, 1975) obteniéndose los siguientes valores y ecuaciones (Fig. 6):
MSY = 169 923 ton.
Fopt= 30 525 lances
ln U = 2.7174 - 3.2772 - 10-5 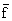
Ye = . e (2.7174-3.2773.10-5)
r = 0.76
| AÑO | DESEMBARQUE TOTAL | CAPTURA/LANCE | FACTOR DE CORRECION | CAPTURA/LANCE | ESFUERZO TOTAL | T |
|---|---|---|---|---|---|---|
| NO-SP-PR-SC (t) | SP | DE ESFUERZO | SP CORREGIDO | ESTIMADO-f | (2 AñOS) | |
| Y | J | J | (Y.) | |||
| 1964 | 38.772 | 2,7 | 0,61 | 4,4 | 0.012 | - |
| 1965 | 50.777 | 3,6 | 0,61 | 5,9 | 8.606 | - |
| 1966 | 59.553 | 4,0 | 0,63 | 6,7 | 8.009 | - |
| 1967 | 80.413 | 5,0 | 0,63 | 7,9 | 10.179 | - |
| 1968 | 76.221 | 4.8 | 0,65 | 7,4 | 10.300 | - |
| 1969 | 113.768 | 5,4 | 0,70 | 7,7 | 14.775 | - |
| 1970 | 135.400 | 5,9 | 0,78 | 7,6 | 17.816 | 16.296 |
| 1971 | 161.027 | 6,2 | 0,87 | 7,1 | 22.680 | 28.248 |
| 1972 | 170.706 | 6,0 | 0,93 | 6,4 | 26.673 | 24.677 |
| 1973 | 228.037 | 6,6 | 0,97 | 6,8 | 33.535 | 30.104 |
| 1974 | 177.089 | 6,2 | 1,00 | 6,2 | 28.563 | 31.049 |
| 1975 | 136.104 | 7,6 | 1,00 | 7,6 | 27.908 | 23.236 |
| 1976 | 105.276 | 6,5 | 1,00 | 6,5 | 16.196 | 17.052 |
| 1977 | 145.576 | 9,3 | 1,00 | 9,3 | 15.553 | 15.925 |
| 1978 | 144.685 | 11,3 | 1,00 | 11,3 | 12.804 | 14.229 |
| 1979 | 149.542 | 10,8 | 1,00 | 10,8 | 13.846 | 13.325 |
| 1980 | 146.277 | 9,2 | 1,00 | 9,2 | 15.900 | 14.873 |
| 1981 | 111.967 | 8,6 | 1,00 | 8,6 | 13.019 | 14.460 |
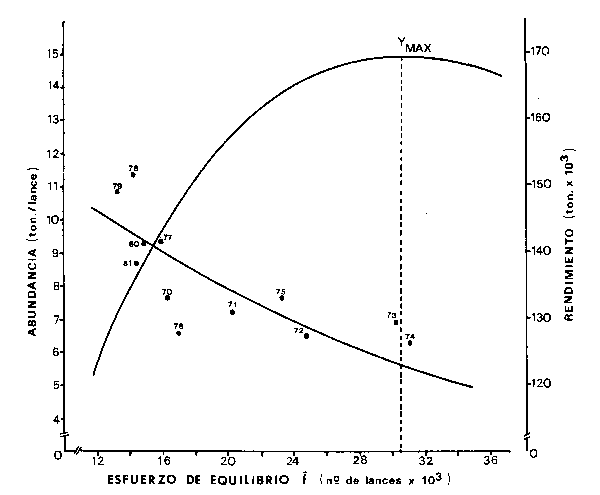
Fig.6- Relaciones entre abundancia/esfuerzo de equilibrio,y rendimiento/ esfuerzo de equilibrio, para los datos 1969-1981, ajustadas según modelo de Fox (1970)
El uso de este modelo fue ampliamente discutido tomando en cuenta las siguientes consideraciones: a) los datos de las estadísticas de pesca utilizados presentan generalmente gran margen de error; b) la unidad de esfuerzo usada puede no representar la verdadera intensidad de la pesca tanto por diferencias en el poder de pesca de la flota, que es heterogenea, como por las razones discutidas anteriormente; c) este modelo presento sabidos inconvenientes cuando aplicado a peces pelágicos en los cuales ocurren concentraciones de los cardúmenes en determinadas áreas a medida que la población se reduce, siendo que la CPUE puede no acusar a1teraciones, una vez que éstas son compensadas por cambios en el coeficiente de capturabilidad; y d) el modelo no puede prevenir eventos descontinuos como fallas en el reclutamientoo en el desove, tan comunes en las poblaciones de peces pelágicos, que frecuentemente resultan en colapsos de stocks.
Otros modelos
Mortalidad natural: la tasa de mortalidad natural fue calculada a través de la ecuación de Pauly (1980) para clupeidos, según la cual:
log10 m/0.6 = 0.0066- 0.279 log10 L + 0.6543 log10 K + 0.4634 log10 T
usando paraámetros con los siguientes valores :
L = 24.0cm K = 0.62 T = 22°C
El valor encontrado fue M=0.74
Los parámetros de crecimiento empleados en esta ocasión fueron calculates a partir de datos oarciales de frecuencia de clases de talla y por tanto el valor de M representa apenas una primera aproximavión de la mortalidad natural de la especie, la que tendrá que ser revisada cuando se obtengan parámetros más satisfactorios.
Con estos valores se aplicaron los modelos de rendimiento por recluta (Beverton y Holt, 1957; Ricker, 1975) y análisis de cohorte (Pope, 1972) con la intención de obtener una evaluación preliminar del stock por estos métodos, estimandose una mortalidad por pesca de F= 0.8 y una captura total permisible de 201 000 ton. Estos todavía se considerran como resultados preliminares dado el limitado volumen de información disponible, y los estudios respectivos deben ser continuados por el GPE.
Utilización de sardina viva como carnada en la pesca de atún
Con la implantación de la pesca de atún, las autoridades y empresarios de la pesca de sardina manifestaron cierta preocupacion en relación al impacto que podría causar la captura de ejemplares muy jôvenes de sardina para ser usados como "carnada-viva".
Conociendo el número de embarcaciones que operan en este tipo de pesca (alrededor de 60 embarcaciones) y la cantidad promedio de sardina viva usada para estos fines por cada barco se estimó en 1 000 ton. el total de sardina capturada anualmente.
Considerando que la talla promedio de las sardinas usadas como carnada es de10 cm y empleando un valor de 0.74 para la mortalidad natural se calculó cual sería el rendimiento en peso de éstas en caso de que no fuesen pescadas.
Se obtuvo un valor de 2 400 ton. de sardinas con una talla de 17 cm, las cuales dejarían de ser reclutadas al stock adulto. Convience recordar que no toda esta cantidad disponible a la pesca serí efectivamente capturada, debido a los egectos posteriores de la mortalidad. De esto se deduce que esta pesquería no implica alteraciones importantes para el stock adulto, a pesar de esto se ha recomendado el cuidadoso seguimiento y reglamentación de la mísma.
Estimación de la biomasa total
Levantamientos acústicos cuantitativos.- en el período de 1974/80 se realizaron 8 campañas a lo largo de toda el area de ocurrencia para estimar la biomasa de sardina a través de métodos hidroacústicos cuantitativos (Rijavec y Amaral, 1977), habiéndose encontrado valores que variaron entre 142 000 y 414 000 ton, con media de 250 000 ton. En esta ocasión se consideró que esta era una sub-estimación, visto que las capturas anuales de 1977-80 presentaban valores de alrededor de 140 000 ton.
Infelizmente la discontinuidad de estas campañas ha impedido el seguimiento sistemático de las fluctuaciones, lo que habría permitido interpretar mejor la situación actual y dimensionar el stock.
Huevos y larvas.- Matsuura (1977), utilizando resultados de campañas de reconocimiento de huevos y larvas realizadas en 1969/70, 1970/71 y 1971/72 estimó la magnitud del stock desovante entre 0.5 × 106 y 10 × 106ton, y para el período 1974/75 y 1975/76, en 1.7 × 106 y 3.3 × 106 ton. respectivamente. Al contrario de la evaluación por métodos hidroacústicos, estos valores representarían una sobre estimación.
Comportamiento de los cardúmenes en relación con las condiciónes oceanográficas
Las oscilaciones de la estructura oceanográfica parecen determiner la disponibilidad de sardina a la pesca, provocando dislocamiento y concentración de los cardúmenes en diferentes regiones del área de distribución (PDP/SUDEPE, 1981 b).
En 1979 se observó una masa de agua fría en la costa de Río de Janeiro que provocó la desaparición de los cardúmenes en esta área, los cuales se dislocaron en su mayoría para el litoral de San Pablo.
En 1980, fenómenos similares provocaron grandes concentraciones a poca profundidad en el litoral de Santa Catarina facilitando las operaciones de la flota con un aumento en la capturabilidad del recurso.
En 1981 nuevamente se observaron altas concentraciones en el litoral paulista.
Todo indica que la estructura oceanográfica característica de invierno, cuando no hay formación de termoclina en pequeñas profundidades, provoca la dispersión de los cardúmenes de sardina, ya que, aparentemente, la temperatura actuaría en estos casos como factor limitante. Por otro lado en el verano se observa una mayor concentración de los individuos a menores profundidades, favoreciendo así su captura, la cual está basada en la identificación visual de los cardúmenes.
Medidas de administración
En 1976, preocupada con la caída de la producción de sardina en ese año y sabedora del comportamiento de otros stocks de peces pelágicos intensamente explotados en otras regiones del munco, la SUDEPE decidió adoptar algunas medidas que permitiesen proteger el stock de la pesca excesiva. Citamos a seguir aquellas que a la fecha se encuentran en vigor:
- tamaño mínimo de captura, establecido en 17 cm, el cual corresponde a la talla media de inicio de la primera madurez sexual;
- período de veda, fijado año a año por 30 a 40 días, de acuerdo con la previsión de la época en que habrá mayor intensidad del desove;
- captura máxima de sardina en toda el área de pesca estipulada en 200 000 ton.
- control del esfuerzo a través de la reglamentación de la concesión de nuevas licencias para los barcos pesqueros que desean capturar sardina.
CONCLUSIONES Y PERSPECTIVAS
En vistas de las consideraciones citadas podemos concluir que:
- La estructura de la especie en el área no fue aún claramente definida, debiendo ser continuados los estudios para verificar las hipótesis propuestas sobre la existencia de más de una población.
- De la misma manera, deberán continur los estudios sobre crecimiento para esclarecer los parámetros (principalmente K y L ) así como una correcta interpretación de la lectura de la edad de los ejemplares para poder utilizar los datos en la aplicación de los modelos de evaluación.
- Los estudios sobre la reproducción ya permiten establecer las características del desove de S. brasiliensis, delimitando el tipo, época y área del desove.
- La pesca presenta algunas señales de colapso, la disponibilidad de los cardúmenes es escasa y han disminuido bruscamente las capturas en los últimos anos.
- Esta disminución puede ser debida a las oscilaciones en la estructura oceanográfica que provocan el desplazamiento de cardúmenes hacia áreas de difícil acceso para la flota comercial; la disminución del tamaño de la población desovante debido a sobrepesca; o fallas del reclutamiento.
- El análasisde los datos disponibles y la aplicación de modèlos matemáticos para diagnosticar la real situación del stock no ha permitido establecer las verdaderas causas de este comportamiento, y parece evidente que la aplicación de modelos de producción general no es adecuada. Es necesaria la sistematización de estudios de evaluación de biomasa por métodos independientes, como levantamientos acustico-cuantitativos y de huevos y larvas, o tal vez marcación.
- Se torna imprescindible ampliar las investigaciones que permitan conocer la real situación del recurso, ya que este representa un alimento de bajo costo y consumo popular a cuya captura están vinculados amplios sectores de la población de pescadores de la región Sudeste del Brasil.
RECONOCIMIENTOS
En esta contribución se han compilado resultados de trabajos llevados a cabo por los siguientes científicos del PDP/SUDEPE: J.C. Amaral (Brasilia), M. M. Bittencourt (Sao Paulo), R. de D. Cardoso (Santa Catarina), L. G. Domit (Paraná), V. J. Isaac Nahum (Sao Paulo), S. Jablonski (Río de Janeiro) y S. A. Saccardo (Sao Paulo) así como C.L.D.B. Rossi-Wongtschowski del Institute Oceanográfice de la Universidad de Sao Paulo y H. Valentini del Institute de Pesca-Secretaria de Agricultura-Sao Paulo.
REFERENCIAS
ACMRR Working Party on Fishing Effort and Monitoring of Fish Stock Abundance. 1976. Monitoring of fish stock abundance: the use of catch and effort data. FAO Fish.Tech.Pap.(155):101 p.
Beverton, R.J.H. y S.J. Holt. 1957. On the dynamics of exploited fish populations. Fish.Inves.Minist. Agric.Fish Food G.B. (2 sea Fish) (19). 533 p.
Ciechomski, J.D. y G. Weiss. 1973. Desove y fecundidad de la ancholta argentina (Engraulis anchoita). Physis, B.Aires, A. 32(84):137-153.
Fox, W.W. 1970. An exploited surplus-yield model for optimising exploited fish populations. Trans. Arner.Fish Soc. 99:80-38.
Isaac-Nahum, V.J., A.E.A. de Vazzoler y E.M. Zaneti-Prado. 1983. Estudos sobre estrutura, ciclo de vida e comportamento de Sardinella brasiliensis (Steindachner, 1879) na área entre 22°S e 28°S, Brasil. IV. Morfologia e histologia de ovários e escala de maturidade. Bolm Inst. oceanogr., S Paulo. (En prensa).
Johannesson, K.A. 1975. Relatório preliminar das observacões acústicas quantitativas sobre tamanho e distribuição dos recursos de peixes pelágicos ao largo do Brasil. PDP/SUDEPE, Doc.Téc. (10):13p., 11 figs.
Matsuura, Y. 1977. O ciclo de vida da sardinha-verdadeira (Introdução à Oceanografia Pesqueira). Publição esp.Inst.oceanogr., S Paulo. (4):146 p.
Matsuura, Y. 1981. Análise econômica da produção da sardinha na regiao Sudeste do Brasil. Bol. inst.oceanogr., S Paulo. 30(1) :57-64.
Pauly, D. 1980. A selection of simple methods for the assessment of tropical fish stocks. FAO Fish.Circ. (729):54 p.
PDP/SUDEPE. 1974. Relatório da Primeira Reunião do Grupo de Trabalho e Treinamento (G.T.T.) sobre Avaliação dos Estoques. Santos (SP), 04-29 de março. PDP/T (7):149 p.
PDP/SUDEPE. 1977. Relatório da Reunião do Grupo Permanente de Estudos sobre Sardinha. PDP/SUDEPE:29p., 9 tabs, 15 figs.
PDP/SUDEPE. 1978. Relatório da Reunião do Grupo permanente de Estudos sobre Sardinha. PDP/SUDEPE:15p., 7 tabs, 03 figs.
PDP/SUDEPE. 1979. Relatório de Reunião do Grupo Permanente de Estudos sobre Sardinha. PDP/SUDEPE:12 p., 9 tabs, 12 figs.
PDP/SUDEPE. 1980. Relatório da Reunião do Grupo Permanente de Estudos sobre Sardinha, PDP/SUDEPE:28 p., 13 tabs, 15 figs.
PDP/SUDEPE. 1981a. Relatório da Segunda Reunião do Grupo de Trabalho e Treinamento (G.T.T.) sobre Avalição dos Estoques. Tamandaré (PE), Julho. (Versão preliminar):441 p.
PDP/SUDEPE. 1981b. Relatório da Reunião do Grupo Permanente de Estudos sobre Sardinha. PDP/SUDEPE:30 p.
PDP/SUDEPE. 1982a. Súmula das Reuniões Técnicas do Programa Integrado de Estudos Biológicos sobre a Sardinha, e do Grupo Permanente de Estudos sobre Sardinha. (Versão preliminar):12 p., 2 tabs.
PDP/SUDEPE. 1982b. Relatório do Programa Integrado de Estudos Biólogicas sobre a Sardinha. (Versão preliminar):14 p, 4 tabs, 21 figs.
Pope, J.G. 1972. An investigation on the accuracy of virtual population analysis using cohort analysis. Int.Comn.Northwest Atl.-Fish Res.Bull. (9):65-74 p.
Projecto MEGALOSAR. 1980. Sardinella brasiliensis (Steindachner, 1879): estudos sobre estrutura e comportamento através de métodos bioqulmicos e sobre ciclo de vida das populaçoes na área entre 22°S e 28°S, Brasil. Instituto Oceanográfico da Universidade de Sao Paulo. Informe cientifico presentado ao "Conselho Nacional de Desenvolvimento Clentífico e Tecnologico (CNPq)".
Projecto MEGALOSAR. 1981. Sardinella brasiliensis (Steindachner, 1879): estudos sobre estrutura e comportamento através de métodos bioquímicos e sobre ciclo de vida das populaçoes na área entre 22°S e 28°S, Brasil. Instituto Oceanográfico da Universidade de Sao Paulo. Informe cientifico presentado ao "Conselho Nacional de Desenvolvimento Clientífico e Tecnologico (CNPq)".
Richardson, I.D., G. Vazzoler, A. de Faria y M.N. Moraes. 1959. Report on sardine investigation in Brasil. Proc.World Sci.Meet.Biol.Sardine & Related Spec., FAO. 3:1051-1079.
Richardson, I.D. y V. Sadowski. 1960. Note on sampling of sardine(sardinella allecia) at Cananeia. State of São Paulo, Brazil. Bolm Inst.oceanogr., S Paulo. 6(1):87-97.
Ricker, W.E. 1975. Computation and interpretation of biological statistics of fish populations. Bull.Fish.Res.Board Can. (191):382 p.
Rijavec, L. y J.C. Amarel. 1977. Distribuição e ebundância de peixes pelágicos na costa Sul e Sudeste do Brasil. (Resultados da pesquisa com ecointegrador). PDP/T (24):55 p., 11 figs.
Rijavec, L., S. Jablonski, A.S.C.B. Simon, L.F. Rodrigues y M.R.Q. Pires. 1977. Contribuição ao conhecimento da biologia e dinâmica da sardinha (Sardinella brasiliensis) em águas brasileiras (Resultados preliminares). PDP/Série Doc.Ocasionais (24):28 p., 8 figs.
Rossi-Wongtschowski, C.L.D.B. 1977. Estudo das variações de relaçõ peso total/comprimento total em função do ciclo reprodutivo e comportamento de Sardinella brasiliensis (Steindachner, 1879) da costa do Brasil entre 23°S e 28°S. Bolm Inst.oceanogr., S Paulo. 26:131-180.
Rossi-Wongtschowski, C.L.D.B . 1978. Sardinella brasiliensis (Steindachner, 1879): estudos sobre estrutura da espécie na área entre 23°s (RJ) e 28°S (SC), Brasil. Inst.de Biociências, Tese de Doutorado: 2 vol., 61 p., 48 tabs, 86 figs.
Rossi-Wongtschowski, C.L.D.B., A.E.A. de M. Vazzoler y F.M. de S. Braga. 1982. Estudo sobre estrutura, ciclo de vida e comportamento de Sardinella brasiliensis (steindachner, 1879) na área entre 22°S e 28°S, Brasil.I.Morfologia dos otólitos. Bolm Inst.oceanogr., S Paulo. 31(2):57-76.
Santos, E.P. y F.M. Fratzen. 1965. Growth of sardines: quantitative aspects. Anais Acad.bras.Cienc. 37(supl):360-362.
Vazzoler, A.E.A. de y P.V. Ngan. 1976. Eletrophoretic patterns of eye-lens proteins of Sardinella brasiliensis (Steindachner, 1879) off Brazilian coast. Rev.Trav.Inst.Pêches marit. 40(3,4):781- 786.
Vazzoler, A.E.A., C.L.D.B. Rossi-Wongtschowski y F.M. de S. Braga. 1982a. Estudo sobre estrutura, ciclo de vida ecomportamento de Sardinella brasiliensis (Steindachner, 1879), na área entre 22°S e 28°S, Brasil. II. Idade e crescimento dos otólitos. Bolm Inst.oceanogr., S Paulo. 32(2):77-84.
Vazzoler, A.E.A., C.L.D.B. 1982b. Estudo sobre estrutura, ciclo de vida e comportamento de Sardinella brasiliensis (Steindancher, 1879), na área entre 22°S e 28°S, Brasil. III. Crescimento: aspectos quantitativos. Bolm Inst.oceanogr., S Paulo. (submetido a publicación).
Wiefels, R.C. y S. Jablonski. 1979. Relatório do Projeto Matriz da Pesca. PDP/SUDEPE:276 p.
![]()
![]()
![]()