![]()
![]()
![]()
by
J.E. Carscadden
Fisheries Research Branch
Department of Fisheries and Oceans
P.O. Box 5667
St. John's, Newfoundland AlC 5Xl
Canada
Resumen
El Capelin (Mallotus villosus) es una especie pequeña que forma cardúmenes y es abundante en el Atlántico Noreste en el área de Newfoundland. Es una especie importante como especie forrajera. Hay cinco stocks de capelin en el área, aun cuando algunos de ellos pueden en realidad ser stocks complejos. A excepción de un área principal de desove ubicada mar afuera, todo el capelin desova en la playa o muy cercana a élla. La población madura se compone principalmente de individuos de tres a cuatro años de edad y la mayor parte mueren después del desove.
Durante décadas el capelin ha sido explotado por una pequeña pesquería dómestica (25 000 ton.), pero durante los años 1970s esta especie fue fuertemente explotada por una pesquería internacional mar afuera. Las capturas totales de capelin llegaron a su máximo a mediados de los años 1970s con 360 000 toneladas y después de eso han declinado. En estos momentos no hay ninguna pesquería mar afuera pero la pesquería costera ha aumentado en los últimos añops debido a la demanda de huevas de capelin en el mercado Japonés.
Las evaluaciones del capelin se han hecho usando modelos de producción, técnicas acústicas y modelos para seguir la secuencia de la abundancia de capelin. Solo el último método ha proporcionado una perspectiva histórica de la dinámica de población de capelin. Estos modelos indican que la abundancia llegó a su máximo a mediados de los años 1970s, mayormente debido a la fuerte clase anual de 1973. Las clases anuales de 1974 y 1978 fueron débiles y la población declinó. Sin embargo la clase anual de 1979 parece haber sido fuerte dando como resultando que la biomasa desovante de capelin en el año 1982 fuera grande.
Las capturas fueron relativamente altas durante los añnos en que se produjeron pobres clases anuales, y esto tiende a implicar a la pesquería en la disminución de la población de capelin. Sin embargo, a excepción de uno de los s stocks desovantes, no se ha mostrado que la pesquería haya tenido ningún efecto mortífero en la dinámica del capelin. Las variables ambientales durante la primera etapa de la vida de capelin afectan la sobrevivencia, en el crecimiento y posiblemente el reclutamiento. La mortalidad de huevos en las arenas de la playa es afectada por la temperatura de incubación, la que en difinitiva es afectada por un complejo de factores incluyendo la densnidad de huevos, la temperatura del mar, temperatura máxima y mínima del aire, horas de luz, lluvia y amplitud de mareas. Estos factores varían solo ligeramente en la mayoría de las principales áreas de desove al este de Newfoundland y al sur del Labrador, y tales cambios sincronizados pueden contribuir a la fluctuación anual de las fuerzas de las clases anuales de capelin. El oleaje que es inducido por el viento que se dirige hacia la playa, es importante para facilitar la liberación de las larvas de la arena de la playa. Estos vientos hacia la playa también inducen el remplazamiento de las masas de agua de tal forma que las larvas son liberadas en aguas más cálidas, con una superficie rica en posibles presas, la cual ha remplazado aguas frías, que son ricas en predatores. Solamente se dispone de datos de dos años, 1978 y 1979, pero en esos años la densidad de huevos en la arena era aproximadamente igual. La frecuencia de vientos hacia la playa durante el período de eclosión fue 50% mayor en 1979, y la densidad de larvas en el ambiente cercano a la orilla fue 60% mayor. La biomasa desovante de capelin fue aproximadamente igual en 1978 y 1979 pero la clase anual de 1979 aparentemente es mucho mayor que la de clase anual de 1978. Análisis preliminares sugieren que hay una relación positiva entre el reclutamiento y la temperatura de la superficie del mar (0–20 m) durante las etapas tempranas de la existencia pelagíca.
De las observaciones cualitativas parece que hay dos especies cuya abundancia ha aumentado durante los períodos de baja abundancia de capelin, estos son la amodita (Ammodytes dubius) en el sur, y el bacalao del ártico (Boreogadus saida) en norte. Los estudios realizados sobre los efectos de las fluctuaciones de capelin y dos predatores principales, el bacalao (Gadus morhua) y el salmón (Salmo salar) no han llegado a establecer ninguna relación firme entre estas especies. Sin embargo, los autores de estos estudios fueron críticos de su trabajo mayormente debido a que los datos disponibles no fueron colectados para probar las relaciones que se habían postuladdo, o porque no dispuso de datos adecuados.
INTRODUCTION
It is probable that capelin, Mallotus villosus, is one of the most important fish species in the northwest Atlantic because of its position as a forage species (Bailey et al., 1974; Winters and Carscadden, 1978) and its commercial importance. This position as an important forage and commercial species has created controversy over the years; for instance, as early as 1835 fisherman complained that the “hauling of capelin for manure had proved very prejudicial to the success of the cod fishery” (Journal of the Newfoundland House of Assembly 1835). In the last decade, capelin has generated a great deal of interest both from the general public and the scientific community. This interest arose largely because of a large commercial capelin fishery that developed in the early 1970's and the potential impact of this fishery on the capelin stocks themselves and on predator stocks, many of which are commercially important.
It is the purpose of this paper to review some aspects of capelin biology, the history of the commercial fishery and its management, the population dynamics of capelin, the factors affecting the abundance of capelin, and the possible consequences of fluctuations of capelin on competitors and predators in the Newfoundland area of the northwest Atlantic.
Life History
Capelin (or caplin) are small, silvery fish closely related to smelt. They are a cold water, pelagic, schooling species inhabiting Arctic and sub-Arctic seas in the Atlantic and Pacific oceans. In the eastern Atlantic, capelin occur from western Norway to northern Russia and are widely distributed throughout the Barents Sea. They are also abundant around Iceland and Greenland. On the east coast of North America, capelin occur from Hudson Bay to Nova Scotia, but are most abundant around Newfoundland and Labrador. In the Pacific, capelin occur from Alaska to Juan de Fuca Strait and from the Sea of Chukotsk to Japan and Korea (Jangaard, 1974).
The relative distribution of capelin in the Newfoundland coastal zone varies seasonally but undoubtedly peaks in June and July, when beach spawning occurs. At other times of the year, capelin can be found in large concentrations in the offshore waters. While the distribution of young capelin is not well defined, the distribution and migration patterns of adults are better known and spawning stocks of capelin have been identified. There are five major stocks in the Newfoundland area; Labrador-northeast Newfoundland stock, NAFO* Div. 2J3K, (A); northern Grand Bank-Avalon stock, NAFO Div. 3L, (B); South Grand Bank (Southeast Shoal) stock, NAFO Div. 3No, (C); St. Pierre-Green Bank Stock, NAFO Div. 3Ps, (D); Gulf of St. Lawrence stock, (E) (Fig. 1). Some of these major stocks (eg. Labrador-northeast Newfoundland and Gulf of St. Lawrence) may be composed of a number of sub-stocks (Winters, 1974a; Sharp et al., 1978). These stocks have been identified through the knowledge of seasonal distribution patterns and growth differences (Campbell and Winters, 1973; Winters, 1974b), neristic (Sharp et al., 1978; Carscadden and Misra, 1980) and morphometric (Sharp et al., 1978) characters and the fishing patterns of the commercial capelin fleet. Besides these offshore stocks of capelin that migrate to the inshore areas or Southeast Shoal to spawn, there is also evidence to suggest that there are local stocks, probably relatively low in abundance. Like most species of fish, there is some degree of stock overlap during some part of the life cycle. Since most of the commercial fishery occurred on only three of the stocks (Div. 2J3K, Div. 3L, and Div. 3NO stocks), most of the subsequent discussion will be restricted to capelin occurring in these areas.
* Northwest Atlantic Fisheries Organization (formely ICNAF, the International Commission for Northwest Atlantic Fisheries
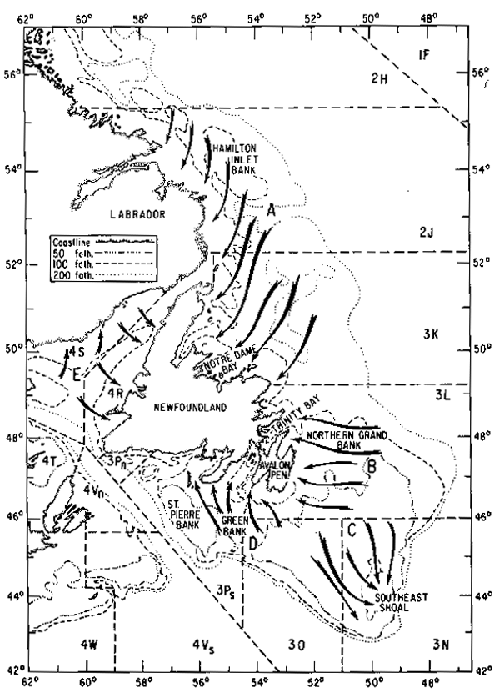
Fig. 1. Map showing major stocks and migration routes of mature capelin.
Alpha-numeric designation (e.g. 2H, 2J, 3K, etc.) are NAFO
Divisions.
A. Labrador - Northeast Newfoundland (NAFO Div. 2J3K) stock
B. Northern Grand Bank - Avalon (NAFO Div.3L) stock
C. South Grand Bank (Southeast Shoal - NAFO Div. 3NO) stock
D. St Pierre - Green Bank (NAFO Div. 3Ps) stock
E. Gulf of St. Lawrence stock
The Notre Dame Bay area is known to be an over-wintering area for part of the Labrador-northeast Newfoundland stock. It seems probable that capelin also over-winter in parts of NAFO Subarea 2; maturing capelin have been taken in research catches late in the year in this area and there is no known northward migration in the spring from Div. 3K to the inshore spawning grounds in Subarea 2. During June and July, mature capelin move inshore to spawn on beaches in northeastern Newfoundland and Labrador. After spawning, most fishes die. The few spawning survivors move offshore in the late summer and join immature fish to form feeding schools. Feeding is heavy from August to November and schools are found in offshore waters from Labrador to northeast Newfoundland.
The northern Grand Bank-Avalon and Southeast Shoal stocks appear to mix and overwinter on the northern part of the Grand Bank (Campbell and Winters, 1973; Carscadden and Misra, 1980). During late winter and early spring the fish become active, form feeding schools, and begin a migration to the spawning grounds. The northern Grand Bank-Avalon stock moves inshore to spawn on Newfoundland beaches while the Southeast Shoal stock moves south over the grand Bank to spawn on sandy substrate on the Southeast Shoal. Spawning in both areas occurs during June and July and post-spawning mortality is high at this time. Spawning survivors are believed to move to the northern Grand Bank area for feeding and over-wintering.
There are differences in the timing of spawning in different areas. Spawning on the west and south coasts of Newfoundland usually begins in late May. Spawning time usually becomes later further to the north such that spawning in Labrador may not occur until August in some years (Templeman, 1948). Spawning on the Southeast Shoal, the only known offshore spawning area for capelin, occurs in June and July.
The general environmental conditions present during spawning have been characterized. Capelin spawning inshore in Newfoundland and Labrador prefer water temperatures of 5.5°C- 8.5°C and pebbles ranging in size from 0.5–2.5 cm in diameter (Templeman, 1948). Spawning normally occurs at night or on dull cloudly days, although there are exceptions. On the Southeast Shoal, spawning occurs in a water depth of approximately 50 m where bottom temperatures are 2–4°C (Pitt, 1958a).
Inshore spawning normally occurs for 4–6 weeks; most spawning occurs on the beaches but if water temperatures on the beach become too high, the fish will spawn in deeper water adjacent to the beaches. The extent of deepwater (non-beach) spawning is probably very variable from year to year and other factors besides water temperature on the beach may exert an influence.
Capelin mature at three or four years of age with faster growing fish maturing earlier (Winters, 1974b). In the spawning populations, three- and four-year-olds usually predominate, although other ages, including two, five, six, and seven do occur in varying proportions. As already noted, spawning mortality is high, usually being greater than 80% (Carscadden and Miller, 1980).
There is no doubt that capelin are important as a forage fish in the Newfoundland area. Perhaps the most extensively studied capelin predator is cod and there are a number of studies (see Akenhead et al., 1982; Lilly et al., 1981 for reviews) that substantiate the observation that cod consume capelin. The broad seasonal patterns of cod predation on capelin in Div. 2J3KL and Div. 3NO have been described but in spite of the numerous feeding studies, “there is no estimate, based on adequate seasonal and spatial sampling of the contribution of capelin to the total food consumption of a single cod stock in a given year” (Lilly et al., 1981). However, several studies have estimated that for the Newfoundland area in general, capelin comprise about 30% of the cod's diet (Akenhead et al., 1982).
Although cod is the capelin in predator that usually evokes the most concern, other marine species feed heavily on capelin as well. Capelin accounts for 45–100% of the food of adult salmon, depending on size, in Newfoundland coastal waters (Lear, 1972). Greenland halibut in the length range of 20–80 cm feed heavily on capelin (Lear, 1969) and American plaice (20–49 cm) in Div. 3L depend on capelin for 14–38% of their food (Pitt, 1973). Haddock, yellowtail flounder, winter flounder, skates, sea ravens, and a variety of other demersal species consume large quantities of capelin eggs and capelin during the spawning season (Pitt, 1958b; Templeman, 1968).
Marine mammals also feed heavily on capelin. Capelin are an important food for whales, including fin whales (Mitchell, 1975), minke whales, sei whales (Sergeant, 1963) and humpback whales (Lien, 1980), in the Newfoundland area. Sergeant (1973) has estimated that at least 25% of the diet of harp seals is capelin and other less abundant species of seals such as hooded, grey and harbour seals, also consume significant quantities of capelin.
Seabirds are also capelin predators; murres (Tuck, 1960), greater shearwaters. puffins, gannets, kittiwakes (Winters and Carscadden, 1978), and gulls (Lewis, 1937) all feed on capelin.
THE FISHERY AND ITS MANAGEMENT
It is only in the last decade that man has become a significant predator of capelin although capelin have been taken during the spawning season for decades. It has been estimated that prior to the 1950's 20,000–25,000 tons of capelin were taken annually in Newfoundland for bait and fertilizer and to a lesser extent for food for dog teams (Templeman, 1968; Barnes, 1974). However, with the decline in the use of dog teams and fertilizer for gardens and less demand for capelin as bait, inshore landings declined considerably. During the 1970's, interest in capelin as a commercial species increased with effort being concentrated in the offshore area in the early and mid 1970's and in the inshore area in the late 1970's.
In 1972, the first substantial offshore catches of about 70,000 tons of capelin were reported. These catches increased rapidly, peaking in 1976 at about 370,000 tons and declining since then. The catches were taken at different times of the year in different areas and details of the catches by areas (Div. 3L, Div. 3NO, and Div. 2J3K) are given in Fig. 2,3 and 4.
The annual offshore capelin fishery normally began in March or April on fish, most of which were maturing and feeding, on the northern Grand Banks (Div. 3L). Thus, fish from two stocks, the northern Grand Bank-Avalon stock and the Southeast Shoal stock, were fished by a fleet dominated by USSR midwater trawlers. The fishing fleet followed the capelin as they moved to their spawning grounds; however, once the inshore spawning component entered Canadian territorial waters, the fleet followed the Southeast Shoal (Div. 3NO) capelin to their spawning grounds where the fishery on spawning capelin continued into July. Most of the vessels operating on the Southeast Shoal were USSR midwater trawlers and Norwegian purse-seiners.
The fishery in Div. 3L developed very rapidly with the peak catch reported in 1974, followed by a gradual decline from 1975–80 (Fig. 2.). It should be noted that in 1975, Canadian ports were closed to the USSR fishing fleet because Canadian authorities accused the Soviets of taking approximately double their quota in Div. 3L. If true, the catch in 1975 (Fig. 2) should be approximately 60,000 tons. In addition, the “inshore” catch in 1974 is shown at approximately 8,000 tons, higher than the preceding and following years. This may be an anomaly caused by the grouping of the data; in the present grouping, Canadian catches were assumed to have been taken inshore since Canada has no fleet of large vessels fishing capelin. However in 1974, some catches were reported from large (500–999 GRT) purse-seiners chartered by Canadian companies. Thus, the catches may have been taken offshore and the inshore catch estimates for 1974 may be over- estimated and the offshore catch estimates under-estimated. It should also be noted that each year some unknown fraction of the Div. 3L offshore catch may have been fish from the Southeast Shoal stock. In general, the trend of catches for Div. 3L since 1974 has been a decrease in the offshore portion (offshore fishery closed in 1979) and an increase in the inshore portion. The inshore catch consists of mature capelin and has been taken entirely by Newfoundland fishermen who are attempting to satisfy the Japanese market for roe capelin.
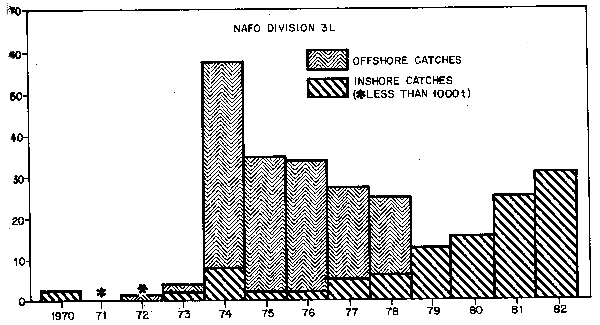
Fig. 2. Nominal catches of capelin in NAFO Div. 3L, 1970–82. 1982 catch data are preliminary.
| Fig. 3. Nominal catches of capelin in NAFO Div. 3No, 1970–82. | 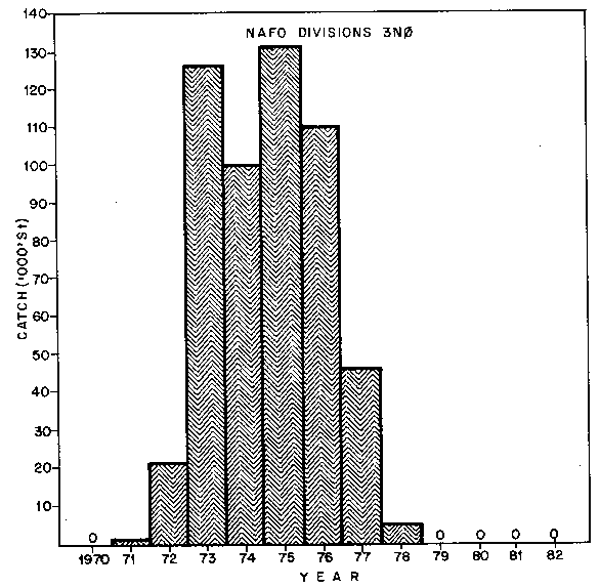 |
The catches (Fig. 3) of the Southeast Shoal stock (Div. 3NO) rose rapidly from about 21,000 tons in 1972 to a level of about 100,000 to 132,000 tons for four years between 1973–1976 and dropped quickly to 47,000 tons in 1977 and 5,000 tons in 1978. The fishery was closed in 1979 and has remained closed since then. It was the conclusion of scientists that “the intense commercial fishery on the spawning grounds in Div. 3N may have substantially reduced the spawning stock size in recent years, and the possibility of recruitment overfishing should be taken into account” (Anon., 1979).
Each year when the fishery on the Southeast Shoal stopped with the cessation of capelin spawning, the attention of the fleet shifted to Div. 2J and Div. 3K in late August or early September. This fishery, which caught feeding capel in most of which would mature and spawn the following year, was prosecruted mainly by USSR midwater trawlers and continued until November or December.
The first large catch in the Div. 2J3K capelin fishery was reported in 1972 at 46,000 tons. Catches peaked in 1976 at about 216,000 tons and declined after that to about 5,000 tons in 1980. During the period 1970–1980, inshore catches remained at a low level below 2,500 tons (Fig. 4).
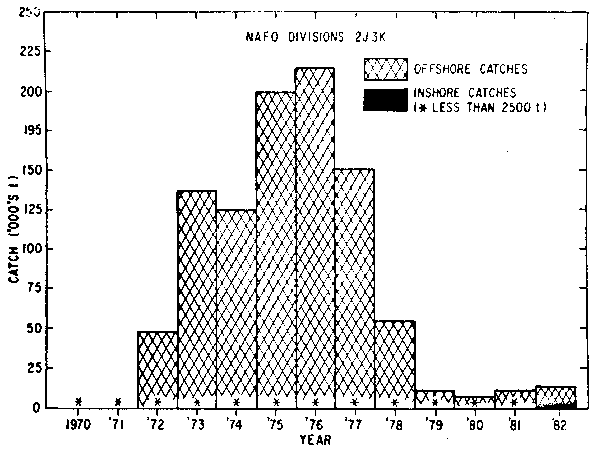
Fig. 4. Nominal catches of capelin in NAFO Div. 2J3K, 1970–1982. 1982 catch data are preliminary.
The management of capelin came under the jurisdiction of ICNAF, and later NAFO, until Canada extended its fisheries jurisdiction to 200 miles (January 1, 1977). At this time, the Div. 2J3K stock came solely under Canadian management because it occurred completely within Canadian jurisdiction although Canada has continued to request that biological advice be provided through NAFO. The Div. 3NO stock overlaps the Canadian zone and biological advice and management recommendations have continued under the auspices of NAFO. Biological advice for the Div. 3L stock continues to be provided through NAFO although all of the fishery occurs within the coastal zone. The biological advice provided for the management of capelin has, with few exceptions, been adopted by the fisheries managers in NAFO and in Canada and as a result, the biological advice and some of the rationale for that advice is provided below.
The biological advice, provided through ICNAF and later NAFO, was initially requested because of the increasing commercial offshore fishery during the early 1970's. The first advice concerning the total allowable catch (TAC) of capelin pertained to 1974 when it was recommended by the Standing Committee on Research and Scientists (STACRES) of ICNAF that the catch should not exceed 250,000 tons. The following year (1975) it was recommended that the total TAC could be increased to 500,000 tons and maintained for three years. Furthermore, it was noted that the fishery should be restricted to mature capelin approaching and during the spawning season and countries participating in the fishery should conduct surveys of both the adult and juvenile stock to monitor the effect of the fishery. Also in 1975, a crude estimate of relative stock sizes resulted in a recommendation that 300,000 tons could be allocated to the northern area (Div. 2J3K) and 200,000 tons to the southern area (Div. 3LNOPs). Further subdivision of the northern TAC was not practical at that time due to insufficient knowledge of stock separation. However, there was evidence to suggest that the fishery in Div. 3L was operating on several spawning components which spawn later inshore in Div. 3L, on the Southeast Shoal, and possibly in Div. 3Ps. It was recommended that only 10,000 tons be reserved for a TAC for Div. 3Ps, since this stock was believed to be small, but that this amount could be taken in Div. 3NO if not taken in Div. 3Ps. It was concluded that it would be desirable to concentrate the fishery as much as possible on the mature capelin in Div. 3NO to minimize potential adverse effects on capelin and other species. This would also reduce the possibility of overfishing any one of the spawning components present in Div. 3L earlier in the season and for the inshore spawning stock, it would minimize any adverse effect on the inshore migration and feeding success of the Div. 3L cod. The final sub- division of the 200,000 TAC in the southern stock was no more than 10,000 tons allocated to Div. 3Ps and no more than 50,000 tons allocated to Div. 3L with the remainder allocated to Div. 3NO. If these maxima were not achieved in Div. 3L and 3Ps, the uncaught amounts could be taken in Div. 3NO (Anon., 1975).
The biological advice remained essentially unchanged until 1979 when evidence of poor recruitment in the capelin population resulted in a recommendation of reductions in the TAC's. STACRES advised that the 1979 TAC in Div. 3LNO should be 16,000 tons and to protect the spawning stock in Div. 3N and during its migration through Div. 30 to Div 3N, there should be no commercial fishery for capelin in Div. 3NO. This meant that 16,000 tons could be taken only in Div. 3L. It was agreed by NAFO that there would be no offshore fishery for capelin in Div. 3L and Canada claimed only 10,000 tons to be taken inshore. Therefore, the quota for capelin in Div. 3L in 1979 was essentially 10,000 tons. In the northern stock, it was recommended that the 1979 TAC be reduced from 300,000 tons to 75,000 tons.
The recruitment prognosis remained poor in 1980 and the biological advice remained unchanged for the stocks in Div. 3LNO that is, a TAC of 16,000 tons with no fishing in Div. 3NO. In the north, the advice was to close the fishery or allow a small nominal TAC. It was noted that a small fishery of 10,000–15,000 tons in the north would allow scientists to better assess the status of the stock in 1980 and to quantify the advice for the next year. An experimental fishery with a TAC of 5,000 tons was conducted in the fall of 1980 to provide data for assessment purposes.
In 1981 there was some evidence of improvement in the southern area and based on the exploitation rate that had been recommended in 1980 and 1981, a TAC of 30,000 tons to be taken in Div. 3L was recommended while the Div. 3NO fishery should remain closed. In the northern area, there was conflicting evidence in the scientific data and as a result, the advice remained unchanged from the 1980 advice; that is, that the fishery remained closed or a small nominal fishery of 10,000–15,000 tons be allowed. An experimental fishery with a TAC of 10,000 tons operated in Div. 2J3K during the fall of 1981.
No scientific advice was requested for 1982 and the 1981 TAC's remained in effect for that year.
An assessment of the stocks conducted in mid 1982 (Anon., 1982) suggested that the stocks had improved dramatically, largely as a result of the strong 1979 year-class. The TAC recommended for the Div. 3L stock was 60,000 tons and for the Div. 2J3K stock, 50,000 tons. Although some improvement was detected in the Div. 3NO stock from surveys conducted in 1981, the abundance estimates were still much lower than historical estimates and it was recommended that this fishery remain closed in 1983.
In general terms, because of the uncertainties involved in the scientific data, the known poor recruitment to the capelin stocks during the late 1970's and early 1980's and the importance of capelin as a food for other marine species, a conservative exploitation rate of 10% has been recommended since 1979 for the Div. 3L stock. The TAC's recommended for the Div. 2J3K stock have been less than 10% of the projected biomass although no specific guideline (e.g. 10%) has been adopted.
POPULATION DYNAMICS AND FACTORS AFFECTING ABUNDANCE
There had been some studies on the biology of capelin (e.g. Templeman, 1948; Pitt, 1958b; Winters, 1966, 1970a, 1970b) but detailed information on population dynamics of capelin was not available during the early phases of the large offshore fishery. There have been three techniques used since the mid–1970's to assess the status of the capelin stocks in the Newfoundland area and describe the population patterns during the last decade; a surplus production model, acoustics methodology, and sequential capelin abundance models. The use of the three assessment methods has shown a trend over time. During the early phase of the fishery, acoustic estimates and the surplus production model were the only sources of information. Both types of estimates were recognized as being crude but they were used as indications of the magnitude of the capelin resource. The surplus production model was used only in the early years of capelin assessments whereas the acoustic method has been used since 1974. As the fishery continued through the 1970's, there were enough data accumulated to develop the sequential capelin abundance models. In the most recent years up to and including 1981, these models have been used as the most reliable indications of stock status and as a starting point for projections. Ironically, these models have depended on the fishery as a data source and the results from the models were largely responsible for the advice to reduce the total allowable catch.
The surplus production model was first developed in 1973 (Campbell and Winters, 1973) and expanded in 1975 (Winters, 1975; later revised by Winters and Carscadden, 1978). Although it is extremely crude, the model was used to provide biological advice and was intended to provide a first estimate of potential long-term annual yield of capelin. It was recognized that many species of marine fish, mammals and seabirds feed extensively on capelin and that the stocks of many of these predators had declined during the 1950's and 1960's. Therefore, it was assumed that there would be capelin available to a commercial fishery in the 1970's that previously would have been necessary to sustain the predator stocks. Cod, whales, and seals were chosen for this analysis because there were estimates of their abundance and of capelin consumption (albeit crude in some cases) by these animals. This model has recently been criticized (Beddington et al., 1981) as a management tool because it is over-simplified. Although this criticism is probably valid, it should be noted that the developers of the model stressed that the estimate of surplus production be interpreted only as indicative of the order of magnitude of potential capelin yield under the assumptions used in the model, most of which could not be tested. Furthermore, the TAC that was eventually put in force was 500,000 tons, twice the level proposed by the authors based on the estimates from the surplus production model.
Acoustic estimates, using echo integration, have been produced primarily by Soviet (Serebrov et al., 1975; Bakanev et al., 1976; Seliverstov and Kovalev, 1976; Kovalev et al., 1977; Klochkov et al., 1977; Bakanev and Seliverstov, 1978; Klochkov and Seliverstov, 1978; Ermolchev et al., 1979; Seliverstov and Serebrov, 1979; Ermolchev et al., 1980; Bakanev 1980, 1981; Bakanev and Ermolchev, 1982) and Canadian (Miller et al., 1978; Miller and Carscadden, 1979a; Carscadden and Miller, 1980; Miller and Carscadden, 1980, 1981; Miller et al., 1982) scientists although one early estimate (Dragesund and Monstad, 1973) was produced from an exploratory survey by the Norwegians. The Soviets have the longest time- series of acoustic estimates of capelin, beginning in 1974 in the northern area (Div. 2J3K) and 1975 in the southern area (Div. 3LNO). The Canadian estimates are more recent with the first estimate in the northern area coming from a 1977 survey and in the southern area from a 1979 survey. In most recent years, except for 1982, the acoustic estimates are used not as absolute estimates but as trend indicators. The decision to use the Canadian acoustic estimates as relative indicators of stock status arose for three reasons. First, there were relatively few measurements of target strength available for capelin; the target strength values are critical for biomass estimation and errors in target strength produce proportional errors in the final biomass estimate. There was no information available on swimming angle of capelin in the field and it is known that target strength can change radically with swimming angle of the fish (Nakken and Olsen, 1977). Second, the personnel collecting the acoustics data and estimating biomass from these data were, at first, relatively inexperienced in this field. As a result there was a period of learning associated with the acoustics methodology which ran parallel to the accumulation of basic biological data to be used in the development of acoustic techniques (e.g. survey design) specifically for capelin. Third, there was a reluctance on the part of other scientists to accept estimates derived from acoustics, a relatively new technology to most, as valid indications of stock status. Although the problems associated with target strength and swimming angle have not been fully resolved, considerable experience has been gained by the Canadian investigators and at the same time other scientists have become more familiar with the advantages and disadvantages of acoustic techniques. In 1982, acoustic estimates were used as absolute estimates (still recognizing the potentially large sources of error associated with target strength) and as a basis for projections (Anon., 1982).
The sequential capelin abundance models (Carscadden et al., 1978; Carscadden and Miller, 1979; Miller and Carscadden, 1979b; Carscadden and Miller, 1980; 1981; Carscadden et al., 1981) which have provided a historical perspective of the dynamics of capelin in the Newfoundland area were developed because the high natural mortality of capelin prevented the use of standard sequential population analysis such as virtual population analysis (Gulland, 1965) or cohort analysis (Pope, 1972). The sequential capelin abundance models (SCAM) are similar to VPA and cohort analysis both in their design and mechanisms. They attempt to calculate for any year-class its age-specific size and fishing mortality rate based on such input parameters as catch-at-age and natural mortality rate. Like cohort analysis, some method of estimating fishing mortality in the most recent years is necessary and in all cases to date, commercial catch rates have been used to calibrate the models and so provide an estimate of terminal fishing mortality. The models differ from cohort and VPA in that some temporal partitioning occurs and the different mortality rates (i.e. high spawning mortality rate at the time of the year and natural mortality rate at other times) are applied to the population estimates at the appropriate time of the year within the model. Since recruitment to the mature portion of the population is essentially equivalent to maturation rate and the fishery operates primarily on mature fish it was necessary to incorporate annual age-specific maturation rates into the models as well. The sensitivity of the models is presently being quantified, however, it is known that they are sensitive to some of the input parameters, e.g. proportions mature- at-age and undoubtedly, like cohort analysis, they are less reliable in most recent years. However, for this discussion, it is assumed that they provide a reasonably accurate representation of capelin population dynamics over the last decade.
The sequential capelin abundance models have been useful for two stocks - Div. 2J3K (SCAM 2J3K) and Div. 3L (SCAM 3L) for 1972–1980. The results of SCAM 2J3K (Carscadden and Miller, 1981) are shown in Table 1 and Fig. 5. The results of this model were not accepted by assessment scientists (Anon., 1982) for biomass estimates in 1981 because the fishing mortality rates estimated in the most recent years were very low (F = 0.01). Instead acoustic estimates for 1981 were used (Table 1, Fig. 5) and therefore, some caution should be exercised in comparing the 1981 estimates with estimates from previous years because of the different methods of assessment. The results of the model indicate that total biomass (Jan. 1) varied between 870,000 tons and 3,697,000 tons between 1972 and 1980 with the biomass peaking in 1975 and 1976. During the same time period, the total biomass at the approximate start of fishing (Sept. 1) varied between 428,000 tons in 1980 and 4,025,000 tons in 1975. For this stock, exploitable capelin biomass on Sept. 1 was calculated by multiplying estimated population numbers-at-age by a calculated partial recruitment factor for each age. This partial recruitment factor was estimated by expressing fishing mortality rate at age each year as a proportion of the highest fishing mortality rate in that year (Carscadden and Miller, 1981). No estimate of exploitable biomass was available for 1981, but between the years 1972 and 1980, the peak biomasses occurred in 1975 and 1976 and declined after that. The results from the acoustic survey suggest that the total biomass (Sept. 1) in 1981 increased by a factor of approximately 4 times from the low abundance in 1978 and 1979. The large biomasses in the mid 1970's were the result of the strong 1973 year-class. The next strongest year-class was the 1969 year-class which resulted in biomasses at intermediate levels in the early 1970's. The 1974–1978 year-classes were very small and the biomass declined during the late 1970's.
| Age/Year | 1972 | 1973 | 1974 | 1975 | 1976 | 1977 | 1978 | 1979 | 1980 | 1981 |
|---|---|---|---|---|---|---|---|---|---|---|
| TOTAL POPULATION AT START OF YEAR | ||||||||||
| 2 | 52.2 | 53.0 | 42.1 | 224.4 | 22.2 | 7.7 | 6.4 | 18.0 | 62.5 | |
| 3 | 60.5 | 38.4 | 38.0 | 30.2 | 161.0 | 15.7 | 5.6 | 4.5 | 13.0 | |
| 4 | 24.5 | 37.6 | 24.5 | 21.8 | 16.3 | 81.8 | 8.7 | 3.1 | 2.9 | |
| 5 | 2.4 | 7.8 | 15.2 | 10.3 | 4.5 | 3.6 | 26.9 | 2.8 | .5 | |
| 6 | .4 | .5 | 1.6 | 3.8 | 1.4 | .5 | 1.0 | 5.2 | .5 | |
| Biomass | 1,987,000 | 2,220,000 | 2,274,000 | 3,697,000 | 3,642,000 | 2,435,000 | 1,238,000 | 501,000 | 870,000 | |
| (tons) | ||||||||||
| TOTAL POPULATION (1 Sept.) | ||||||||||
| 2 | 42.7 | 43.4 | 34.4 | 183.7 | 18.2 | 6.3 | 5.3 | 14.7 | 51.2 | 71.0 |
| 3 | 42.9 | 28.6 | 26.7 | 20.6 | 98.1 | 11.2 | 4.3 | 3.3 | 9.2 | 10.0 |
| 4 | 9.1 | 19.0 | 12.3 | 5.8 | 4.6 | 33.1 | 3.7 | .6 | 1.1 | 2.0 |
| 5 | .6 | 2.1 | 4.9 | 1.7 | .6 | 1.5 | 6.3 | .5 | .1 | 1.0 |
| 6 | .1 | .1 | .4 | .3 | .1 | .2 | .1 | .5 | .1 | 1.0 |
| Total | ||||||||||
| biomass | 1,987,000 | 2,251,000 | 1,848,000 | 4,025,000 | 2,843,000 | 1,474,000 | 434,000 | 428,000 | 1,177,000 | 1,794,000 |
| (tons) | ||||||||||
| Exploit. | ||||||||||
| biomass | 485,000 | 1,048,000 | 984,000 | 1,516,000 | 1,613,000 | 644,000 | 245,000 | 334,000 | 169,000 | |
| (tons) | ||||||||||
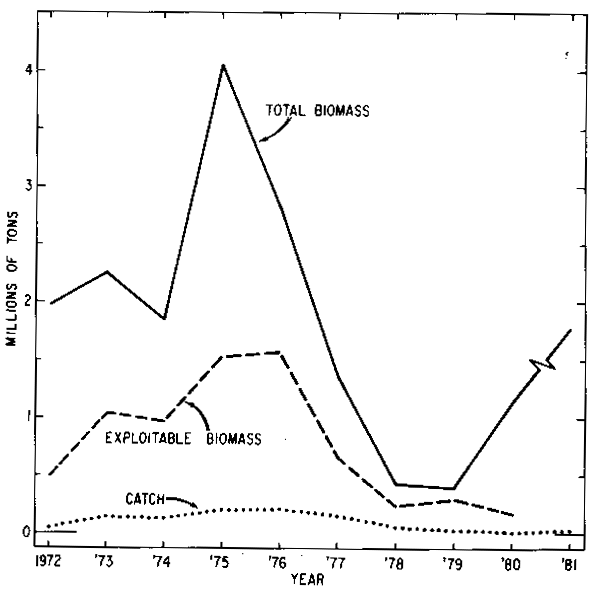
Fig. 5. Estimates of total biomass (1 Jan.) and exploitable biomass (1 sept.), 1972–80 from SCAM 2J3K acoustic estimate (fall 1981) and catch for Div. 2J3K capel in stock.
The 1979 year-class appears stronger than the 1974-1978 year-classes; this year-class has appeared in abundance as immature fish during research vessel surveys. Although the 1982 data have not been analyzed completely, observations from our surveys, from other scientists and from fishermen around the Island indicate that the spawning biomass of capelin significantly increased in 1982 and this was due largely to the 1979 year-class. An examination of the catches (Fig. 5) over the 1972–1981 period suggests that catches were very low in relation to the biomass available.
The results of the analysis using SCAM 3L are given in Table 2 and Fig. 6. It should be emphasized that these estimates are considered less reliable than the results for the Div. 2J3K stock using SCAM 2J3K. Estimates from the Div. 3L model could be calculated only up to 1977 because of a lack of reliable independent estimates of abundance in most recent years. Estimates of abundance for 1978–1981 are based on predicted values of year-class strength at age 3 derived from a relationship between year-class strengths in the Div. 2J3K and Div. 3L stocks (Carscadden and Miller, 1979; Carscadden et al., 1981) which suggests that parallelism in year-class strength of capelin occurs in all areas of the northwest Atlantic. Estimates of abundance for 1981 acoustic surveys (Miller et al., 1982) suggested a total biomass of 1,764,000 tons (ages 1–6) and 1,103,000 tons of age 3 fish and older in Div. 3L, or approximately 3.5 times the projected estimate from Carscadden et al., 1981). The results of a Soviet acoustic survey (Bakanev and Ermolchev, 1982) estimated approximately 421,000 tons of immature capelin in Div. 3L and northern Div. 3N. Unfortunately this estimate is not strictly comparable to the Canadian estimate because very few older mature capelin were observed. The Canadian survey was conducted closer to the coast and undoubtedly detected significant numbers of age 3 fish and older (compare the estimated numbers of 1973 year-class in the two surveys) that were migrating inshore to spawn.
The capelin stock in Div. 3L was highest in abundance in the mid 1970's and intermediate in abundance in the early 1970's, largely due to the strongest year-classes, the 1973 and 1969 year-classes. According to the estimates of biomass for the late 1970's shown in Table 2 (assuming the parallelism of year-class strengths), the biomass dropped during this period. Although the acoustic estimates from the late 1970's were never used as absolute for assessment purposes, the trends were taken as indicative of stock status and support the results of SCAM 3L. The 1979 year-class was also relatively strong in this area resulting in a higher spawning biomass in 1982. As in the Div. 2J3K stock, the catches in Div. 3L have been relatively low in relation to the estimates of capelin biomass for this stock.
Thus, for both the northern (Div. 2J3K) and southern (Div. 3LNO) areas, the large commercial capelin fishery that operated during the 1970's grew very rapidly, peaked in the mid 1970's and declined rapidly in the late 1970's; this fishery pattern coincided with the pattern of capelin biomass. The catch levels declined as the population biomass declined because of poor year-classes between 1974 and 1978. The fact that the peak catches of the fishery occurred during the years in which small year-classes were produced tends to implicate the fishery in the decline. In fact, the fishery has been cited as a probable contributing factor only in the decline of the Div. 3NO stock (Anon., 1979). The analyses that have been conducted to date for the Div. 3L and Div. 2J3K stocks suggested the catch was not large in relation to the biomass available and therefore, was unlikely to have had deleterious effects on the capelin stocks. Furthermore, the 1979 year-class, which appears to be large, resulted from the spawning in a year in which both the capelin biomass and the fishery were small.
Although the factors affecting the variation in year-class strength of capelin are not fully understood, there is evidence available to suggest that environmental factors operating on the early stages of the life history are affecting survival and therefore recruitment. Frank and Leggett (1981a, b; 1982a, b) have been studying factors affecting capelin egg and larval mortality on a capelin spawning beach in Newfoundland since 1978. The time between egg deposition on the beach and hatching in the gravel was influenced by average incubation temperature; this incubation temperature varied between years and between cohorts in a single year and between low, mid, and high tide locations on the beach. Water temperature, maximum and minimum air temperatures and hours of sunlight all determined the average incubation temperatures. The interacting effects of egg density, water temperatures, maximum and minimum air temperature, hours of sunlight, rainfall and tidal amplitude explained over 70% of the temporal and spatial variance in egg mortality. The climatic and hydrographic variables that influence egg mortality vary only slightly over most of the major spawning sites in eastern Newfoundland and southern Labrador areas and the authors suggest that synchronous changes in these variables may contribute to the annual fluctuations in capelin year-class strength (Frank and Leggett, 1981b).
| Age/Year | 1972 | 1973 | 1974 | 1975 | 1976 | 1977 | 1978 | 1979 | 1980 | 1981 |
|---|---|---|---|---|---|---|---|---|---|---|
| TOTAL POPULATION AT START OF YEAR (SCAM 3L) | ||||||||||
| 3 | 12.7 | 9.0 | 8.2 | 5.1 | 24.5 | 9.9 | 5.1 | 5.0 | 6.0 | 9.9 |
| 4 | 3.9 | 6.1 | 4.7 | 3.0 | 1.8 | 13.1 | 5.3 | 2.4 | 2.4 | 2.8 |
| 5 | .3 | .8 | 3.0 | .5 | .3 | .4 | 4.7 | 1.1 | .5 | .5 |
| Biomass (tons) | 386,000 | 311,000 | 368,000 | 216,000 | 529,000 | 483,000 | 358,000 | 167,000 | 150,000 | 302,000 |
| TOTAL POPULATION (ACOUSTIC), June 1981 | ||||||||||
| Age | CANADIAN | USSR | ||||||||
| 1 | 43.0 | 42.9 | ||||||||
| 2 | 33.0 | 42.1 | ||||||||
| 3 | 45.0 | 3.1 | ||||||||
| 4 | 10.0 | 3 | ||||||||
| 5 | 3.0 | 2 | ||||||||
| 6 | 1.0 | - | ||||||||
| Total Biomass (tons) | 1,764,000 | 421,000 | ||||||||
| 3+ biomass (tons | 1,103,000 | |||||||||
| Fig 6. Estimates of total biomass (1 Jan.) 1972–81 from SCAM 3L acounstic estimate (June 1981) and catch for Div. 3L capelin stock. | 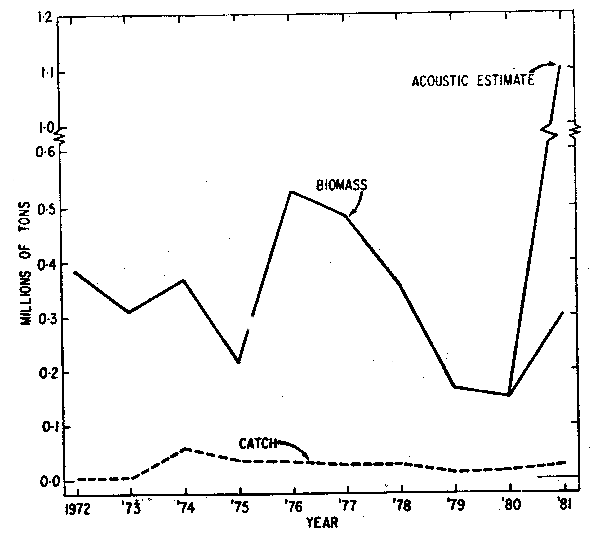 |
Capelin eggs hatch in the gravel and the larvae remain there until washed into the near-shore environment by wave action (Frank and Leggett, 1981a). Larval growth during the residence in the gravel was regulated by the interacting effects of water temperature and hours of sunlight. The swimming performance of the larvae on their release into the water was directly related to their condition which in turn was negatively related to the length of time spent in the beach gravel and to the temperature conditions during that time (Frank and Leggett, 1982a).
The emergence of larval capelin from beach gravel was correlated with onshore wind- induced wave action (Frank and Leggett, 1981a). There was a positive correlation between density of larvae in the beach gravel and the length of time separating the occurrence of onshore winds. The onshore winds that were instrumental in releasing the larvae from the gravel also induced rapid water mass exchange in the inshore waters and as a result, cold, high-saline, predator-laden waters which were present during periods of offshore winds were replaced with warmer, less-saline waters, which contained more potential prey (Frank and Leggett, 1982b). Thus, the net effects of the onshore winds were to facilitate the release of larvae from the gravel and their entry into the nearshore waters; to induce water mass changes in the nearshore waters such that the resulting waters were warmer and less-saline; to cause a 3–20 fold reduction in the potential predator complex; to increase the potential food resource for larval capelin by 2 to 3 times. Furthermore, Frank and Leggett (1982b) reported that the release of large numbers of capelin coupled with the reduction of predators during onshore winds resulted in predator satiation. As a result, the capelin larvae initiate their drift in an environment that is predictable with respect to wind conditions and consistent over the species range in Newfoundland (Frank and Leggett, 1982b). However, the timing and the frequency of these onshore winds is unpredictable and of low frequency since the summer winds in Newfoundland are generally offshore (Frank and Leggett, 1982b).
Data available for two years, 1978 and 1979 (Frank and Leggett, 1981a) indicate that egg densities in the gravel were similar. However, the frequency of onshore winds during the hatching period was 50% greater and larval densities in the nearshore environment were 60% greater in 1979. This increase in larval density also agreed with the observation that the proportion of larvae exiting the beach in good physical condition in 1979 was higher than 1978. These findings are interesting when compared to the estimates of biomass of adult capelin (Fig. 5 and 6) for 1978 and 1979 and year-class strengths resulting from these spawnings. Adult biomass in both 1978 and 1979 was low but there was an increase in released larvae in good physical condition in 1979, because of a greater frequency of onshore winds, and the 1979 year-class is apparently larger than the 1978 year-class. The spawning biomass of capelin in 1982 was much improved, largely because of the contribution of the 1979 year-class.
Once released from the beach gravel into the pelagic environment, the larval capelin
are transported quickly from the immediate beach area and into the onshore waters (W.C.
Leggett, pers. comm.). During the next few months, they are restricted to surface waters
(Jacquaz et al., 1977; Fortier and Leggett, 1983) where growth and survival are probably
influenced by prevailing hydrographic conditions. We have assumed that hydrographic
conditions during this larval phase in the surface layers influence recruitment and have
recently begun investigating the relationship between recruitment and two hydrographic
variables, temperature and salinity. Preliminary results from these investigations are
reported here. The temperature data are from Station 27 (47°32'50“N. 52°35'10”W),
located off St. John's, Newfoundland and estimates of year-class strength of capelin as
two-year-olds from northern (Div. 2J3K) stock (Table 1) were used as estimates of
recruitment. Although the temperature data are not from exactly the same area, the
Labrador current is the dominant hydrographic feature in the Div. 2J3K area and the
Station 27 area; hence, the temperatures at Station 27 should reflect temperature trends
in the northern area. Temperatures used were the sums of the monthly means for July–
December from surface to a depth of 20 m (S. Akenhead, pers. comm.). The abundance of
the 1969 year-class as two-year-olds was estimated from the abundance of three-year-olds
using the same assumptions as in the model. The recruitment data were transformed to
natural logarithms because Hennemuth et al. (1980) reported that recruitment in most of
the fish stocks they investigated exhibited a lognormal pattern. There was a significant
relationship (Fig. 7) between recruitment (R) and water temperature (TEMPSUM) (![]() = 11.74
+ 0.27 TEMPSUM, r = 0.71). Although the time series is relatively short, the relationship
was tested using detrended data (Box and Jenkins, 1976). The data were detrended by taking
differences:
= 11.74
+ 0.27 TEMPSUM, r = 0.71). Although the time series is relatively short, the relationship
was tested using detrended data (Box and Jenkins, 1976). The data were detrended by taking
differences:
TEMPSUMo = TEMPSUMt - TEMPSUMt-1
where Rt is recruitment in year t and Tt is temperature summed in year t. The relationship
was still positive (![]() nRo = -0.03 + 0.21 TEMPSUMo, r = 0.66) but was not quite significant
at the 5% level (Fig. 7).
nRo = -0.03 + 0.21 TEMPSUMo, r = 0.66) but was not quite significant
at the 5% level (Fig. 7).
The association between surface salinity and recruitment was also tested using salinity values from Station 27 over the same time period. There was no significant relationship between salinity and recruitment.
It is possible that predation has some influence on year-class variation in capelin and Dawe et al. (1981) investigated the effect of inshore predation by squid (Illex illecebrosus) on the abundance of capelin using correlation analysis. Squid are summer migrants into the Newfoundland area. Their populations fluctuate widely from year-to- year and while in the inshore area, they feed heavily and grow rapidly. In the analysis, the authors found a negative correlation between the abundance of squid and year-class strength (ages 0+1) of Div. 2J3K capelin. However, they noted that there was a number of problems with their analysis, usually related to the unavailability of proper data to evaluate their hypothesis. For instance, there are no quantitative studies on squid feeding and although squid appear to have the potential to have a significant impact on prey species, the biomass of the prey species cannot be calculated because it is not known if squid prey on larvae, 0-group, 1-group or older fish. The authors were forced to use estimates of capelin year-class size at ages 2,3, or 4 as indicators of abundance of age groups 0 and 1 since neither estimates of abundance of the younger age groups nor estimates of natural mortality were available. The analysis may have been confounded by temporal trends since the time series for capelin was only 8 years. In this respect it is interesting to note that in 1979, squid abundance was very high (Dawe et al., 1981) while the 1979 year-class of capelin appears to be good. Thus, this correlation may disappear with the addition of the 1979 point. Furthermore, recent squid feeding data suggests that capelin are not important components of the diet of squid during the inshore phase of the life cycles of both species (E.G. Dawe, pers. comm.).
AFFECTS OF CAPELIN FLUCTUATIONS ON COMPETITOR AND PREDATOR STOCKS
Although the trends in abundance of capelin during the last decade are known, there is very little information on the effects of the fluctuations on possible competitor and replacement species. In the southern part of the capelin range, the only resident species that might be considered to occupy a similar niche is sand lance (Ammodytes dubius) while in the north this position would be taken by arctic cod (Boreogadus saida). While there are no detailed quantitative data available for these species, it appears that the populations of sand lance (G.H. Winters, pers. comm.) and arctic cod (H. Lear, pers. comm.) did increase during the late 1970's when capelin abundance had declined. It is interesting to note that the increase in arctic cod, whose normal distribution is much further north than that of capelin, and the decrease in capelin abundance occurred during a time when colder water temperatures predominated (Fig. 7).
Vesin et al. (1981) noted that the abundance of squid (Illex illecebrosus) was high when capelin abundance was low and they estimated that with the decline of capelin in the Newfoundland area in the late 1970's, substantial quantities of zooplankton would be released to other predators. This led them to speculate that this release of zooplankton may be important in regulating changes in year-class of other species, notably arctic cod and squid. Except for the estimates of surplus zooplankton, no quantitative analyses were presented to support the hypothesis.
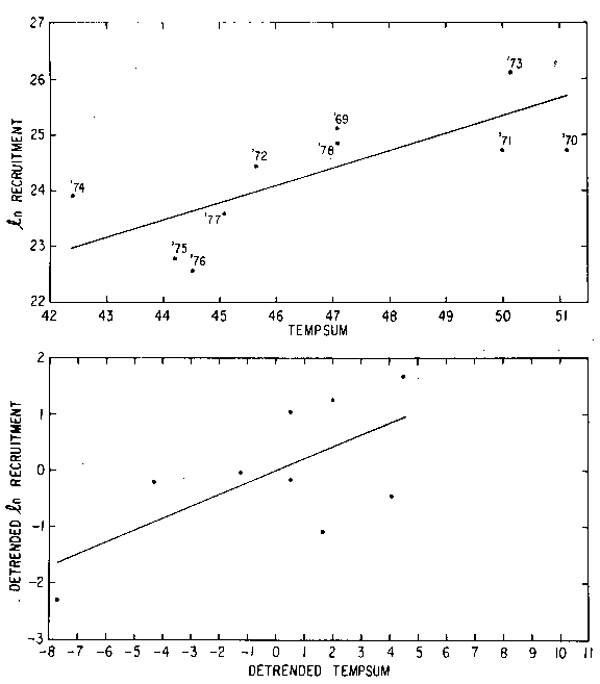
Fig. 7. Relationship between recruitment and water temperature.
Top: ![]() n recruitment vs. TEMPSUM
n recruitment vs. TEMPSUM
Bottom: detrended ![]() n recruitment vs. detrended TEMPSUM
n recruitment vs. detrended TEMPSUM
(see text for details)
A great deal of concern has been expressed about the effects of fluctuations in capelin abundance on predator stocks and this has prompted scientific studies using existing data on two of these relationships - capelin and cod (Gadus morhua), and capelin and salmon (Salmo salar). The effects of fluctuations of capelin biomass on cod growth and on the inshore cod fishery were investigated by Akenhead et al. (1982). There was no statistically significant effect of capelin biomass on cod growth detected. However, after making corrections for the biomass of cod, the authors did detect significant relationships between cod trap catches versus capelin biomass as well as between total inshore gear catches versus capelin biomass and water temperature. However, there were a number of uncertainties in the data and the authors suggested that the statistically significant relationships not be accepted as unequivocal proof that such relationships do exist in nature but that they be used to determine the direction of future research.
Recently, the failure of the 1977 smolt-class of salmon prompted Carter (1979, 1980) to state that the decline of capelin due to overfishing caused the failure of this smolt- class. It appeared that an unusually high mortality of this smolt-class had occurred during its first winter at sea (Reddin and Carscadden, 1981). There were virtually no feeding data available to indicate whether capelin were important in the diet of salmon during their first sea-winter but the size of young capelin in relation to smolts suggested that they would be suitable prey. Assuming that capelin were eaten by salmon, Reddin and Carscadden (1981) showed, using correlation analyses, that in some cases there were statistically significant relationships between salmon catches and abundance of young capelin. However, the authors noted that these relationships were significant only because of the 1977 smolt-class. Indeed, when the analyses were repeated (Carscadden and Reddin, 1982) using data updated by one year, none of the relationships was statistically significant.
The analyses of the cod-capelin and salmon-capelin relationships failed to produce unequivocal proof of the dependence of the predator on capelin. In both analyses, the data available were not collected to test the postulated relationships, thereby making the results suspect, or in some instances, the proper data were not available. For instance, cod trap catches were assumed to be an indicator of inshore cod abundance which further assumes that fishing effort has not changed over the time period in question. A better indicator would have been catch per effort to account for annual changes in effort unfortunately effort data were not available. In both studies, it was assumed that the predators were dependent on capelin and a change in the abundance of capelin would result in the dynamics of the predator. However, the diet of both cod (Akenhead et al., 1982) and adult salmon (Reddin and Carscadden, 1982) is spatially and temporally variable and in the absence of capelin, both species would feed on other prey if available. In this case, an absence of capelin probably would not result in starvation of the predator nor an immediate drop in abundance. Instead, a reliance on other prey or less prey might result in slower growth and/or reduced fecundity, if in fact there was a detrimental effect at all. Such effects would be subtle and long-term making them difficult to detect.
ACKNOWLEDGEMENTS
D.S. Miller has contributed significantly in the development of and computer programming for the acoustic techniques and sequential capelin assessment models. G.H. Winters has provided valuable advice and helped in the development of the sequential capelin abundance models. Scott Akenhead provided the monthly Station 27 data, 0–20 m. None of the information presented here would have been possible without the capable assistance of a number of technicians in the Pelagic Section, Fisheries Research Branch, St. John's.
REFERENCES
Akenhead, S.A., J.E. Carscadden, H.Lear, G.R. Lilly and R. Wells. 1982. Cod-capelin interactions off northeast Newfoundland and Labrador. p. 141–148. In Multispecies approaches to fisheries management advice. (M.C. Mercer, ed.). Can.Spec.Publ.Fish.Aquat.Sci. 59:169 p.
Anon. 1975. Int.Comm.Northwest Atl.Fish.Redbook. STACRES Spec.Meet. Jan. 1975.
Anon. 1979. Int.Comm.Northwest Atl.Fish.Redbook. STACRES Spec.Meet. Feb. 1979.
Anon. 1982. Rep.of Scientific Council. NAFO SCS Doc. 82/18, Ser.No. N577, 77p.
Bailey, R.J.F., K.W.Able and W.C. Leggett. 1974. Seasonal and vertical distribution and growth of juvenile and adult capelin (Mallotus villosus) in the St. Lawrence estuary and western Gulf of St. Lawrence. J.Fish.Res.Board Can. 34:2030–2040.
Bakanev, V.S. 1980. Results of USSR investigations on capelin in Divisions 2J and 3K, Autumn 1979. NAFO SCS Doc. 80/43. Ser.No. NO75. 6p.
Bakanev, V.S. 1981. Results of Soviet investigations on capelin in Northwest Atlantic in 1980. NAFO SCR Doc. 81/11/10. Ser.No. N274, 10p.
Bakanev, V.S. and V.A. Ermolchev. 1982. Hydroacoustic assessment of capelin abundance in NAFO Divisions 3LNO in May–June 1981. NAFO SCR Doc. 82/61, Ser.No. N554, 10p.
Bakanev, V.S. and S.A. Seliverstov. 1978. Capelin investigations in the waters off southern Labrador (Division 2J) and on the north Newfoundland Bank (Division 3K) in the autumn of 1977. ICNAF Res.Doc. 78/30, Ser.No. 5191, 11p.
Bakanev, V.S., S.A. Seliverstov and L.I. Serebrov. 1976. Preliminary instrument estimate of abundance and biomass of capelin off south Labrador and the north Newfoundland Bank (Divisions 2J and 3K). ICNAF Res.Doc. 76/VI/54, Ser.No. 3840, 16 p.
Barnes, M. 1974. Preface to Final Report of 1973 Newfoundland Inshore Capelin Development Program by Andrew Duthie, Industrial Development Branch, Newfoundland Region, St. John's Newfoundland.
Beddington, J., D. Botkin and S.A. Levin, 1981. Mathematical models and resource management. p. 1–5. In Renewable Resource Management, Proceedings of a Workshop on Control Theory Applied to Renewable Resource Management and Ecology. Lecture Notes in Biomathematics No. 40, Springer-Verlag. N.Y.
Box, G.E.P. and G.M. Jenkins. 1976. Time series analysis, forecasting and control. Holden-Day, Toronto.
Campbell, J.S. and G.H. Winters. 1973. Some biological characteristics of capelin, Mallotus villosus, in the Newfoundland area. ICNAF Res.Doc. 73/90, Ser. No. 3048, 8p.
Carscadden, J.E. and D.S. Miller. 1979. Biological aspects of capelin and a sequential capelin abundance model for the Division 3LNO stock. ICNAF Res.Doc. 79/11/33, Ser.No. 5359, 20 p.
Carscadden, J.E. and D.S. Miller. 1980. Analytical and acoustic assessments of the capelin stock in Subarea 2 and Div. 3K, 1979. NAFO SCR Doc. 80/13, Ser.No. NO45, 19 p.
Carscadden, J.E. and D.S. Miller. 1981. Analytical assessment of the capelin stock in Subarea 2 + Div. 3K using SCAM. NAFO SCR Doc. 81/4, Ser. No. N268, 8 p.
Carscadden, J.E., D.S. Miller, and G.H. Winters. 1978. Biological aspects of the Div. 3NO capelin stock in 1977 and a capelin sequential abundance model. ICNAF Res.Doc. 78/VI/52, Ser.No. 5217. 20 p.
Carscadden, J.E. and R.K. Misra. 1980. Multivariate analysis of meristic characters of capelin (Mallotus villosus) in the northwest Atlantic. Can.J.Fish.Aquat.Sci. 37:725–729.
Carscadden, J.E. and D.G. Reddin. 1982. Salmon-capelin interactions: here today, gone tomorrow. CAFSAC Res.Doc. 82/23. 10 p.
Carscadden, J.E., G.H. Winters and D.S. Miller. 1981. Assessment of the Division 3L capelin stock NAFO SCR Soc. 81/VI/25, Ser.No. N304, 14 p.
Carter, W.M. 1979. Where have all the salmon gone? Int.Atl.Salmon Found.Newsletter 9(4).
Carter, W.M. 1980. Capelin and Atlantic Salmon. Int.Atl.Salmon Found. Newsletter 10(1).
Dawe, E.G., G.R. Lilley and J.A. Moores. 1981. An examination of the influence of squid (Illex illecrbrosus) on recruitment in several finfish stocks off eastern Newfoundland and Labrador. NAFO SCR Soc. 81/VI/25, Ser.No. N304, 14 p.
Dragesund, O. and T. Monstad. 1973. Observations on capelin (Mallotus villosus) in Newfoundland waters. ICNAF Res.Doc. 73/33. Ser.No. 2967, 14 p.
Ermolchev, V.A., S.M. Kovalev and A.S. Seliverstov. 1979. Methods and results of echometric surveys on the assessment of the Grand Newfoundland Bank capelin abundance in spring-summer 1979. ICNAF Res.Doc. 79/31, Ser. No. 5357, 14 p.
Ermolchev, V.A. S.M. Kovalev, V.S. Mamylov and A.S. Seliverstov. 1980. Assessment of capelin stocks in Divisions 3LNO in May-June 1979. ICNAF Res.Doc. 80/42, Ser.No. N0-74, 9 p.
Fortier, L. and W.C. Leggett. 1983. Vertical migrations and transport of larval fish in a partially mixed estuary. In review Can.J.Fish.Aquat.Sci.
Frank, K.T. and W.C. Leggett. 1981a. Wind regulation of emergence times and early larval survival in capelin (Mallotus villosus). Can.J.Fish.Aquat.Sci. 38:215–223.
Frank, K.T. and W.C. Leggett. 1981b. Prediction of egg development and mortality rates in capelin (Mallotus villosus) from meteorological, hydrographic, and biological factors. Can.J.Fish.Aquat.Sic. 38:1327–1338.
Frank, K.T. and W.C. Leggett. 1982a. Environmental regulation of growth rate, efficiency, and swimming performance in larval capelin (Mallotus villosus) and its application to the match/mismatch hypothesis. Can.J.Fish.Aquat.Sci. 39:691–699.
Frank, K.T. and W.C. Leggett. 1982b. Coastal water mass replacement: its effect on zooplankton dynamics and the predator-prey complex associated with larval capelin (Mallotus villosus). Can.J.Fish.Aquat.Sci. 39:991–1003.
Gulland, J.A. 1965. Estimation of mortality rates. Annex to Arctic Fisheries Working Group Report. Cons.Perm.int.Explor.Mer, 1965. CM, Doc. No. 3.
Jacquaz, B., K.W. Able and W.C. Leggett. 1977. Seasonal distribution, abundance, and growth of larval capelin (Mallotus villosus) in the St. Lawrence estuary and northwestern Gulf of St. Lawrence. J.Fish.Res.Board Can. 34:2015–2029.
Jangaard, P.M. 1974. The Capelin (Mallotus villosus) biology, distribution, exploitation, utilization and composition. Bull.Fish.Res.Board Can. 186:70 p.
Klochkov, D.N., and A.S. Seliverstov. 1978. Assessment of abundance and biomass of the spawning stock of capelin in ICNAF Division 3N in summer of 1977. ICNAF Res.Doc. 78/29, Ser. No. 5190, 6 p.
Klochkov, D.N., A.S. Seliverstov and L.I. Serebrov. 1977. Estimate of capelin stocks in the south Labrador and northern Newfoundland Bank areas in the autumn of 1976 (Divisions 2J and 3K). ICNAF Res.Doc. 77/33, Ser.No. 5058, 11 p.
Kovalev, S.M., A.S. Seliverstov and M.L. Zaferman. 1977. Size of a spawning capelin stock on the Grand Newfoundland Bank (Div. 3N) in 1976 (South-Western Stock). ICNAF Res.Doc. 77/32, Ser.No. 5057, 8p.
Lear, W.H. 1969. The biology and fishery of the Greenland halibut (Reinhardtius hippoglossoides) in the Newfoundland area. M.Sc. Thesis, Memorial University of Newfoundland, 1969.
Lear, W.H. 1972. Food and feeding of Atlantic salmon in coastal areas and over oceanic depths. Int.Comm.Northwest Atl.Fish.Res.Bull. No. 9:27–29.
Lewis, H.F. 1937. Notes on the birds of the Labrador peninsula in 1934 and 1935. Can.Field Natur. 51(7):99–105.
Lien, J. 1980. Whale entrapment in inshore fishing gear in Newfoundland. Final Report to Fisheries and Oceans, Canada.
Lilly, G.R. 1981. Influence of the Labrador Current on predation by cod on capelin and sand lance off eastern Newfoundland. Northwest Atl.Fish.NAFO Sci.Coun. Studies 3:77–82.
Lilly, G.R., R. Wells and J. Carscadden. 1981. Estimates of the possible consumption of capelin by the cod stocks in Divisions 2J + 3KL and 3NO. NAFO SCR Doc. 81/8. Ser.No. N272, 9 p.
Miller, D.S. and J. Carscadden. 1979a. An acoustic estimate of capelin biomass in ICNAF Divisions 2J and 3K. October 1978. ICNAF Res.Doc. 79/34, Ser. No. 5360, 8 p.
Miller, D.S. and J. Carscadden. 1979b. Biological characteristics and biomass estimates of capelin in ICNAF Divs. 2J + 3K using a sequential capelin abundance model. ICNAF Res.Doc. 79/32, Ser.No. 5358, 15 p.
Miller, D.S. and J. Carscadden. 1980. An acoustic survey of capelin (Mallotus villosus) in Divisions 3LNO, 1979. NAFO SCR Doc. 80/4, Ser.No. NO46, 8 p.
Miller, D.S. and J. Carscadden.1981. Acoustic survey results for capelin (Mallotus villosus) in Divisions 2J3K and 3LNO, 1980. NAFO SCR Doc. 81/5, Ser.No. N269, 10 p.
Miller, D.S., J.E. Carscadden and B. Bennett. 1978. An acoustic estimate of capelin (Mallotus villosus) biomass, ICNAF Divisions 2J and 3K, October and November 1977. ICNAF Res.Doc. 78/43 (Rev.), Ser.No. 5205, 7 p.
Miller, D.S., B.S. Nakashima and J.E. Carscadden. 1982. Capelin acoustic surveys in NAFO Divisions 2J + 3K, 3LNO, and 3L, 1981–82. NAFO SCR Doc. 82/54, Ser.No. N547, 12 p.
Mitchell, E. 1975. Trophic relationships and competition for food in Northwest Atlantic whales. Proc.Can.Soc.Zool., 2–5 June 1974, p. 123–132.
Nakken, O. and K. Olsen. 1977. Target strength measurements of fish. Symposium on Hydroacoustics in fisheries research. Rapp.P.-v.Réun.Cons.int.Explor.Mer, 170:52–69.
Pitt, T.K. 1958a. Distribution, spawning and racial studies of the capelin, Mallotus villosus Mueller in the offshore Newfoundland area. J.Fish.Res.Board Can. 15:275–293.
Pitt, T.K. 1958b. Age and growth of the capelin. Mallotus villosus (Mueller) in the offshore Newfoundland area. J.Fish.Res.Board Can. 15:295–311.
Pitt, T.K. 1973. Food of American plaice (Hippoglossoides platessoides) from the Grand Bank, Newfoundland. J.Fish.Res.Board Can. 30:1261–1273.
Pope, J.G. 1972. An investigation of the accuracy of virtual population analysis using cohort analysis. ICNAF Res.Bull. 9:65–74.
Reddin, D.G. and J.E. Carscadden. 1981. Salmon-capelin interactions. CAFSAC Res.Doc. 81/2, 38 p.
Seliverstov, A.S. and S.M. Kovalev. 1976. Size of the capelin spawning stock on the Grand Newfoundland Bank. ICNAF Res.Doc. 76/51, Ser.No. 3837, 14 p.
Seliverstov, A.S. and L.I.Serebrov. 1979. Status of capelin stocks in Divisions 2J and 3K in 1978. ICNAF Res.Doc. 79/30. Ser.No. 5356, 12 p.
Serebrov, L.I., V.S. Bakanev and S.M. Kovalev. 1975. The state of the Newfoundland capelin stock. ICNAF Res.Doc. 75/7, Ser.No. 3435, 11 p.
Sergeant, D.E. 1963. Minke whales, Balaenoptera acustorostrata Lacepede, of western North Atlantic. J.Fish.Res.Board Can. 20:1489–1504.
Sergeant, D.E. 1973. Feeding, growth and productivity of Northwest Atlantic seals. J.Fish.Res. Board Can. 30:17–29.
Sharp, J.C., K.W. Able, W.C. Leggett and J.E. Carscadden. 1978. Utility of meristics and morphometric characters for identification of capelin (Mallotus villosus) stocks in Canadian Atlantic waters. J.Fish.Res.Board Can. 35:124–130.
Templeman, W. 1948. The life history of the capelin (Mallotus villosus) Mueller in Newfoundland waters. Nfld.Govt.Lab.Bull. 17:151 p.
Templeman, W. 1968. Review of some aspects of capelin biology in the Canadian area of the northwest Atlantic. Rapp.P.-v.Réun.Cons.int.Explor.Mer, 158:41–53.
Tuck, L.M. 1960. The murres: their distribution, populations and biology. A study of the genus Uria. Can.Wildl.Ser., Monogr.Ser. 1:206 p.
Vesin, J.-P., W.C. Leggett and K.W. Able. 1981. Feeding ecology of capelin (Mallotus villosus) in the estuary and western Gulf of St. Lawrence and its multispecies implications. Can.J.Fish.Aquat. Sci. 38:257–267.
Winters, G.H. 1966. Contribution to the life history of the capelin, Mallotus villosus, in Newfoundland waters. Fish.Res.Board Can. MS Rep.Ser. No. 870. 56 p.
Winters, G.H. 1970a. Record size and age of Atlantic capelin, Mallotus villosus. J.Fish.Res.Board Can. 27:393–394.
Winters, G.H. 1970b. Biological changes in coastal capelin from the overwintering to the spawning condition. J.Fish.Res.Board Can. 27:2215–2224.
Winters, G.H. 1974a. Rationale for partition of capelin quota in Subareas 2 and 3. ICNAF Res.Doc. 74/12, Ser.No. 3158, 2 p.
Winters, G.H. 1974b. Back-calculation of the growth of capelin (Mallotus villosus) in the Newfoundland area. ICNAF Res.Doc. 74/7, Ser.No. 3150, 16 p.
Winters, G.H. 1975. Review of capelin ecology and estimation of surplus yield from predator dynamics. ICNAF Res.Doc. 75/2, Ser.No. 3430, 25 p.
Winters, G.H. and J.E. Carscadden. 1978. Review of capelin ecology and estimation of surplus yield from predator dynamics. ICNAF Res.Bull. 13:21–30.
by
Armin Lindquist
Director
Fishery Resources and Environment Division
Fisheries Department
FAO, Via delle Terme di Caracalla
00100 Rome, Italy
Resumen
Por lo menos a partir del siglo XVI han habido varios “períodos de arenque” que han sido de gran importancia socioeconómica para la provincia de Bohul\?\n, en la costa de Skagerrak de Suecia. Durante estos períodos de arenque, el arenque entró en los confines del archipiélago cada invierno y fue pescado con redes de cerco de playa y redes agalleras. Capturas del orden de las 200 000 toneladas en una temporada de invierno eran usuales y al término de cada período — después que el arenque había estado entrando al archipiélago cada invierno por 20 a 50 años— la economía de la provincia declinóo a niveles de pobreza.
Hay registrosde la pesquería de espadín sóla para los últimos 100 años y durante ese tiempo se han observado cambios característicos en su distribución geográfica dentro del archipiélago, pero ninguna periodicidad.
La pesquería períodica de arenque y la pesquería de espadín estan relacionadas entre sí, sin embargo la pesquería de arenque está basada en clases anuales muy grandes de arenque del Mar del Norte, mientras que el espadín depende de condiciones hidrógraficas del Mar especiales. Estas condiciones hidrográficas son a su tiempo un prerequisito para que el arenque entre al archipiélago en grandes cantidades durante los períodos de arenque. Se hace referencia al trabajo por el cual el tipo de arenque fur identificado así como al posible mecanismo hidrográfico que afecta tanto al espadín como al arenque. Se sugiere que un debilitamiento de los vientos del oeste y un cambio en el flujo del Báltico hayan sido las claves de este fenómeno.
Se asume de que la secuencia de varios años en que clases anuales fuertes de arenque de Mar del Norte entraron al archipiélago son una expresión de la periodicidad que ocurrió en un área más amplia, pero no fue detectada debido a que no había pesquería de arenque en el mar abierto.
Para que ocurra el fenómeno conocido como “período de arenque” en el archipiélago de Bohusl\?\n hay dos prerequisitos, que ocurren independientemente se requiere una secuencia larga de clases anuales fuertes y un cambio en el sistema general de circulación océanica. Esto implica que cuando no hay cambios en la circulación océanica pero clases anuales fuertes, el arenque no entra al archipiélago. Esto significa que la periodicidad mostrada por la pesquería de Bohul\?\n no dará una imagén verdadera de la historia de las clases anuales fuertes del arenque del Mar del Norte.
INTRODUCTION
The discussion whether under natural conditions there is an equilibrium situation, permitting predictable yields, has been dominated by theoretical considerations. There are, unfortunately, not so many long—term series which monitor pelagic fish stocks. There are still less examples when human, i.e. fishery, influence can be considered negligible at least at the beginning and during the peak of the fishery. Fishermen know very well that availability of fish varies from year to year, but it has been difficult for them in discussions to distinguish between occurrences of fish in catchable quantities from stock sizes. As a matter of fact, this question is of little importance to the daily work of the fisherman. For the biologist it is of paramount importance - unfortunately the relation between fishability and stock size remains obscure and fishermen's observations can only be used with care. It therefore would be interesting to have examples with great natural fluctuations, which are well documented. One of the few cases where this has been possible is the case of the California sardine with the deposits of their scales in the sediment (Soutar and Isaacs 1974). It may be added that such work certainly should be carried out in other parts of the world and would give substantial information about natural fluctuations, such as those off Peru over the last 2 000 to 2 700 years (DeVries and Pearcy 1982).
One example of long-term fluctuations, which have been of greatest socio-economic importance are the herring periods in the Bohuslän archipelago, on the west coast of Sweden, Fig. 1. This fishery, in which the periodic appearance of herring has been mentioned since mediaeval times, has been studied more scientifically for the first time about one hundred years ago (Ljungman 1879). Many explanations have been given for the reason why the herring occurred in great masses, some of them of a highly speculative character (as periodicity in tidal waters caused by variations in frequency of sun spots). The only detailed investigations on the herring population(s) which gave rise to the periods has been made by Höglund (1972, 1978). The reason why the herring entered the archipelago en masse during the autumn and remained there during the winter has not however been explained. Other studies in the same area have shown that another clupeid changed its distribution and that this is likely to be connected with some hydrographic change (Lindquist 1964, 1978). The purpose of this contribution is to analyse further the phenomenon of the herring periods and to show that there are at least two independent mechanisms which may cause a herring period. Detailed observations may thus give pieces of a large puzzle but not necessarily predictable fluctuations.
THE PHENOMENON
Herring is recorded to have occurred in great quantities and be fished during long periods in the Bohusl\?\n archipelago. These long periods, called herring periods, were interrupted by even longer periods when there was no herring and no fishery. Before this century there is no information whatsoever on the occurrence of herring in the open sea of the Skagerrak, as there was no herring fishery. It is well documented that the herring fishery in some of the “periods” had been of an extraordinary socio-economic importance for this province (Holmberg, 1963) which besides a moderate agriculture and some shipping had few other opportunities for earnings. The details of the herring periods have first been described by biologists and then by historians (for ref. see Höglund, 1976). According to the latter there are the following periods for which sufficient documentation is available, showing the economic problems following both the upswing when the herring appeared and the downswings when the herring ceased to enter the archipelago during the autumn and winter:
| Herring in the archipelago | 1556–89 | (1643–1698 ? ) | 1747–1808/9 | 1877–1906 | (no herring “period” since 1906) | ||||
| Herring in the open sea | …………… (no information) …………… | –1920 | 1943–54 | 1963–65 | |||||
The “period” in the seventeenth century is very questionable; Ljungman (1879) found that there was an abundant fishery at least between 1660 and 1680. For the latest period annual catches are given in Fig. 2. They are, considering the short coastal area and the gear used (see below) remarkable. During the season 1895–96 more than 200 000 t were caught. Still more remarkable are the catches in the period of the eighteenth century where catches were higher and amounted to about 270 000 t in one season, taken with beach seines only (Höglund 1976).
Fig. 1. The area investigated: (northern) Kattegat, Skagerrak.
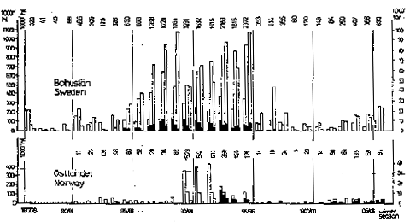
Fig. 2. Yield of herring in the Skagerrak during the winters of 1877/1878 to 1906/1907.
Earlier periods have been claimed to have taken place but a closer look shows that those periods do not stand the examination of the professional historians. For those earlier “periods” there are only scattered reports in mediaeval documents or in the sagas. The periods given above are thus those which we can accept as facts. Herring was so abundant that it was fished with gillnets and beach seines and only from the very end of the last century with purse seines.
For the time from 1903 onwards there is additional information with biological data, Hoglund and earlier authors (as Andersson, for reference see Höglund, 1979) have pointed out that the same herring as in the archipelago occurred during the winter in the open Skagerrak and they found years of abundant appearance, calling them “open sea herring periods”. As the open sea fishery started in 1903 and ceased in 1920 this is considered as being the real end of the herring period which started in 1877. There were two later periods with great herring abundance and a rich fishery in the open Skagerrak, but not in the confines of the archipelago, namely 1943–1954 and 1963–1965.
The time until 1920 with the fishery in the open sea is considered to be simply a continuation of the herring period within the archipelago — but at the end the herring for some reason did not enter the confines of the archipelago any more. The other “open sea” periods occurred independently from any invasion into the archipelago. Similar open sea periods during the winter in the Skagerrak may have occurred earlier between the recorded herring periods, without having been noticed as there was no fishery in the open sea.
Of the about 10–30 fish species of economic importance occurring in the area, only three are pelagic, namely apart from herring, sprat (Sprattus sprattus) and mackerel (Scomber scombrus). The latter is a seasonal immigrant during the warmest time of the year and is of little interest in this connection. For the former there are observations available for the fishery during the last hundred years (Lindquist 1978). Sprat occurs in shoals, either nearly devoid from herring or mixed with small herring of the same size. Before about 1970 sprat was only fished for human consumption and sales resulted in high prices. The main fishery in the Skagerrak was carried out during the winter in the confines of the archipelago and observations on quantities and localities of the fishery exist since 1860 (Lindquist 1964). From the analysis of the records it can be seen that the location of the fishery has changed several times between the northern and southern part of the coast, Fig. 3. As sprat fishermen always followed the sprat on its move one can be fairly sure that the location of the fishery reflected occurrence. As regards quantities fished and landed, there has not been any kind of “regularity” as in the herring fishery.
| Fig. 3.Location of the sprat fishery off the Swedish west coast and temperature differences between the northern and the southern part of the Swedish west coast. (a, top) Main location of the sprat fishery: N, northern part; N+S, entire coast; S, southern part. (b, bottom) Differences in the air temperature in 5-year periods between the stations Väderöbod and Vinga (see Fig. 1), from September to March. (From Lindquist, 1964. partly modified.) |  |
BIOLOGICAL BACKGROUND
Unfortunately there are rather few length measurements and age determinations of the herring which formed the bulk of the last herring period, thus not allowing for a reliable identification of all the herring. Indirect methods showed that it is very likely that it was the autumn spawning North Sea herring (NSAS) which wintered in the confines of the archipelago of Bohuslän. For the open sea period which ended in 1920 and the later open sea periods it has been shown to be NSAS. For the herring period 1747–1808/9 there is a rich material of remnants of train oil factories, as the train oil waste was buried ashore, where it still can be found in many places. Höglund investigated those semi—fossil layers consisting of vertebrae and other bones (scales disappear during the cooking process) and found that the age composition, length/ distribution, and annual ring pattern of the majority of the herring caught some 200 years ago and used for production of train oil is similar, if not identical, with the herring fished now in of the North Sea (including the Skagerrak and the Kattegat).
It can therefore safely be assumed that the herring during any of the herring periods was of the same type.
For sprat no races of the kind found in herring have been described and no meristic or morphometric groupings of sprat from the coastal fishery in the Skagerrak have been found (see further Lindquist 1968).
HYDROGRAPHICAL BACKGROUND
There are few long—term hydographical series in the Skagerrak, and those existing permit only calculations of mean temperatures. Froqner (1948) showed that there has been a substantial increase in mean temperatures in spring to autumn temperatures since the thirties (stations Torungen and Flθdevigen on the Norwegian Skagerrak coast), Fig.4. There is no tendency or any event in the temperature regime which can be connected with the onset or end of the last herring period. Lamb (1973) has shown how the intensity of westerly circulation over the British Isles changes since 1860, Fig. 5.
In order to get longer time series another approach was made, using air temperatures in the coastal area, as it has been observed that there is a firm relationship between air temperature and sea surface temperature (Frogner, 1948). Lindquist (1964) showed that even when using this method there is no direct relation between mean temperatures and the latest herring period, 1877–1906.
The picture becomes quite different, however, if for one hundred years annual temperature differences are compared between the northern and the southern Skagerrak, not absolute values of surface temperature, Figs. 3 and 6. It was found that the northern coast during the long time of observations has been either warmer or colder as compared to the southern coast, the border between “north” and “south” seems to be in the middle, where there is a boundary area of different water masses, known to fishermen.
This hydrographical background fits rather neatly with the localization of the sprat fishery, Fig. 3. As sprat is a relatively warm water fish, sprat follows or at least is found in areas which are warmest during autumn/winter. Sprat was thus found either in the northern part or in the southern part of the coast during the last one hundred years. This seems reasonable as a fish cannot compare a mean value now with that from last year or earlier. Therefore gradients only are of importance. They allow for preferences within physiological limits.
From the same hydrographic material another observation can be made: the latest herring period at the turn of the century coincides well with the time when the northern coast was warmer during winter. This also coincides with the time when the westerly winds were weaker, Fig. 5, and the colder Baltic water from the south would have gained a wider distribution.
As herring is a cold water fish a simple explanation of temperature preference as in the case of sprat cannot be given. The picture, however, may be much more complicated as the changed temperature regime may be the expression of larger hydrographic changes which are as yet unknown.
Even from this century (in the forties) there are minor trends in differences in surface temperature making it likely that there have been similar hydrographical changes as earlier.
Fig. 4. Differences between the air temperature in 5 years periods at the stations Torungen and Hallo, each month X - III.
Fig. 5. Number of days per year of general westerly circulation over the British Isles (Lamb, 1973).
HYPOTHESIS
From the studies referred to above one can draw the following conclusions:
- the fluctuations in the quantity of herring must basically have been caused by a sequence of strong year classes of mainly North Sea herring; and
- there has been a hydrographic event of a nature not known in detail occurring at the time of the last herring period.
A series of good year classes has to be assumed as the herring periods lasted for two to five decades with the herring arriving nearly every autumn/winter in the archipelago. The material from the two last herring periods shows that the majority of herring belongs to North Sea herring. When a sequence of good year classes is terminated, the stock gets more vulnerable to the fishery thus accelerating the end of the herring period. The total catches of herring in the North Sea are recorded since the beginning of this century and do not show any tendency which can be related to the Skagerrak situation (Burd 1978, Fig. 128, 132). Earlier statistics apparently do not exist.
The reason why the herring entered the confines of the archipelago en masse every autumn is unknown but is most likely connected with changes in current systems and in the outflow from the Baltic Sea.
We find thus two different mechanisms, the biological one and the hydrographical one, the latter given one of the necessary conditions for the phenomenon of a herring period in the archipelago. When there are both strong year classes and the necessary hydrographic conditions, herring appeared in the archipelago. There remains now the question of why both the sequence of good year classes and the hydrographical changes happen at the same time. This question is, however, not strictly justified, as there may have been other “periods” i.e., sequences of good year classes which were never detected because the hydrographic conditions at the most prevented or at least did not favour entering the confines of the archipelago. It is possible that both a sequence of good year classes and hydrographical changes are expressions of larger changes (westerlies) and may have the same ultimate cause, but this has to be left as an open question.
The important conclusions are (a) that the North Sea herring as such must have shown a periodicity not observed elsewhere; and (b) that temperature differences have been of greater importance for the distribution of fish and the fishery than absolute temperatures.
SUMMARY
![]()
![]()
![]()