![]()
![]()
![]()
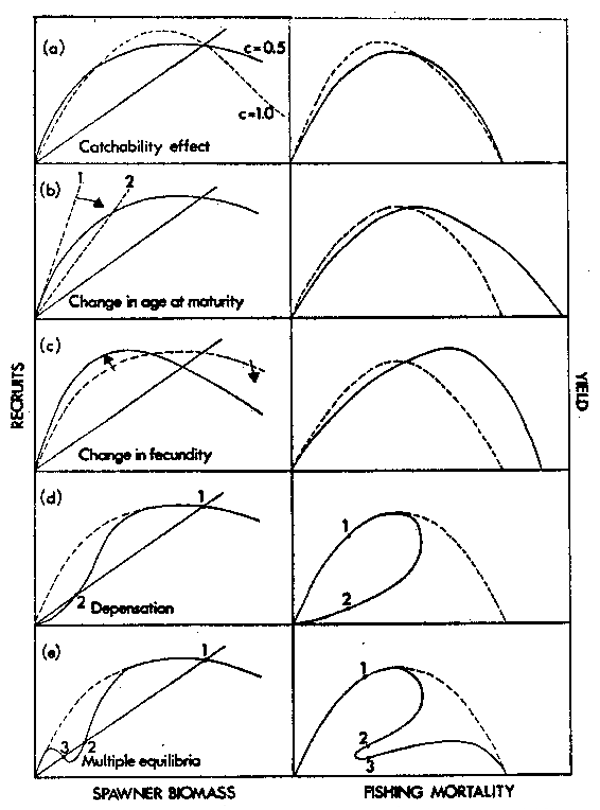
Figure 11. A summary of the effect of density-dependent population responses on the shape of both the stock-recruit and yield curves. Replacement at a relatively low fishing mortality is indicated by the solid diagonal line on the stock-recruit curves.
(a) A comparison between stocks exhibiting the stock-recruit relationship R = Ne-bN where c = 1 (a normal Ricker curve) and c = 0.5 (spawner biomass increasing faster than spawner density).
(b) The effect of reducing age at maturity at low stock sizes. The steeply sloping dashed line (1) represents replacement if no change in maturity has occurred at an equivalent F value.
(c) The effect of a compensatory increase in fecundity as spawner stock is reduced (solid curve).
(d) The effect of depensation in the stock-recruit relationship. A stable equilibrium occurs at 1, but 2 represents an unstable equilibrium.
(e) A stock-recruit curve for a population escaping depensation at a low spawner biomass, for example due to a Type III functional predator response on pre-recruits. Points 1 and 3 are stable equilibria, but point 2 is unstable.
STOCK-RECRUIT PLOTS FOR THE SOUTH AFRICAN ANCHOVY AND PILCHARD
The 18 stock-recruit data points for the South African anchovy fall on the lefthand limb of a shallowly domed recruitment curve with an arbitrary c value of 0.5 (Figure 12). The first two data points, 1964 and 1965, are low and may indicate an underestimate of recruitment since the fishery only partially exploited the stock in the first two years. The points are all well above the replacement line in the absence of fishing (F = 0) but replacement lines in keeping with fishing mortality values before and after the good recruitment of 1973 pass through the respective groups of points. In 1979, 1980 and 1981 recruitment fell below the replacement line and the population appears to be headed back down the lefthand limb of the recruitment curve. Since the topography of the lefthand limb was defined as the population grew and has been shown to be fairly gently sloping, management should be able to prevent collapse provided there is no gross environmental anomaly.
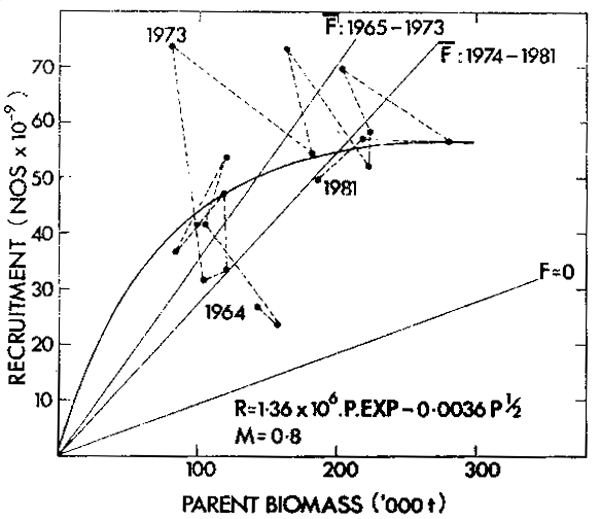
Figure 12. A generalized recruitment curve for anchovy at m = 0,8 where parent biomass is one-year-old fish and older and recruitment is calculated at 6 months of age. Replacement at F = 0 and at average F values during the periods 1965–1973 and 1974–1981 is shown.
Data points on the pilchard stock-recruit plot (Figure 13) have been differentiated into two groups. The open circles correspond to years before 1965 including the period of high biomass and subsequent decline. The closed circles are for data between 1966 and 1980, a period of relative stability, despite the fact that effort is directed towards this species for canning. The parent biomass axis incorporates the large changes in maturity that have been observed in the population. In the absence of this compensation and at present levels of fishing mortality, the replacement line would have been almost vertical (Figure 13).
The resilience displayed by the pilchard to heavy fishing pressure, having fallen below replacement recruitment on a number of occasions in recent years, cannot easily be explained by a conventional recruitment curve. The resilience may be evidence of a stable equilibrium at a low population size, as described by Peterman et al. (1979) and illustrated in Figure lle. Even if fishing on the pilchard were to cease entirely, recovery would not be automatic if the lower descending righthand limb were still cut by the F = 0 replacement line. This may explain the lag in the recovery of the Japanese sardine Sardinops melanostica and the failure of the Californian sardine Sardinops caerulea to recover despite the reduction of fishing pressure.
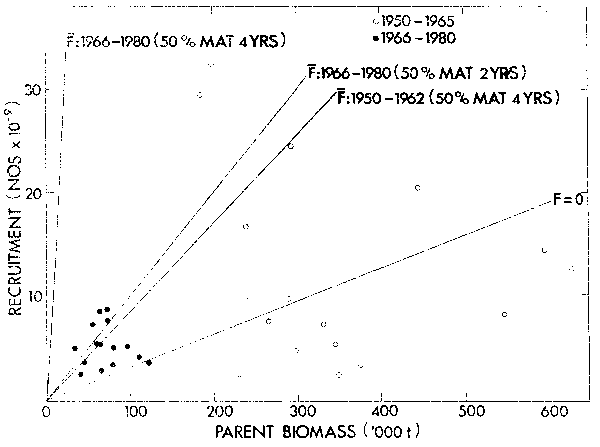
Figure 13. Scatter plot of pilchard recruitment at six months of age against parent biomass (incorporates the density-dependent change in age maturity). Open circles are 1950 – 1965 stock-recruit values and closed circles 1966 – 1980 values. The replacement line at F = 0 applies to the 1950 –1965 values only; replacement lines at average F for 1950 –1962 and 1966 – 1980 take into account the change in age at maturity between these periods, but replacement at average F during 1966 – 1980 if age at maturity had not changed is also shown.
STOCK-RECRUIT CURVE AND MANAGEMENT
A curve fitted to a set of stock-recruit points should not be extrapolated beyond those points without caution. Although a Ricker or Beverton and Holt curve can often be fitted to the data, in a stock where fishing mortality is increasing it is the area of extrapolation at lower spawner biomass which could be most significant in determining the pattern of future yields. We maintain that by examining the density-dependent processes that are displayed at intermediate stock sizes a preview of the topography of the recruitment curve at lower stock sizes can be obtained and used in a cautious approach to management. Once a suitably shaped recruitment curve has been decided upon, the incorporation of a stochastic element with a similar variability to that observed in the stock-recruit data points can be useful in determining the potential variability of yields at different levels of fishing mortality.
As an example of this approach Armstrong et al. (1983) used the observed variance in the South African anchovy recruitment as stochastic input into a regenerating yield model. A 100-year run of this model at F = 0,5 is shown in Figure 14. By running the model 50 times over a 20-year time horizon, a long-term mean yield, C.O.V. of the yield and probability of the yield falling below 100 000 tons was determined for a series of F values (Figure 15).
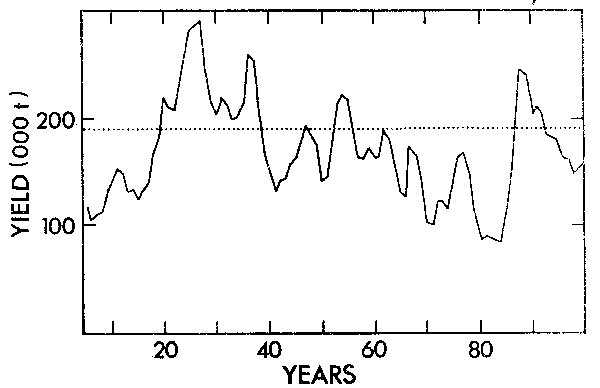
Figure 14. An example 100-year run of a stochastic anchovy yield model at F = 0,5. The stochastic input was derived from the observed deviation of Ln R/N values about a fitted Ln R/N versus regression line and hence expresses a normal distribution of Ln a values. The effect of serial correlation in density- independent egg and larval mortality and /or fecundity due to environmental cycles was examined by applying a serial correlation coefficient of 0,5 to the Ln a values (from Armstrong et al., 1983).
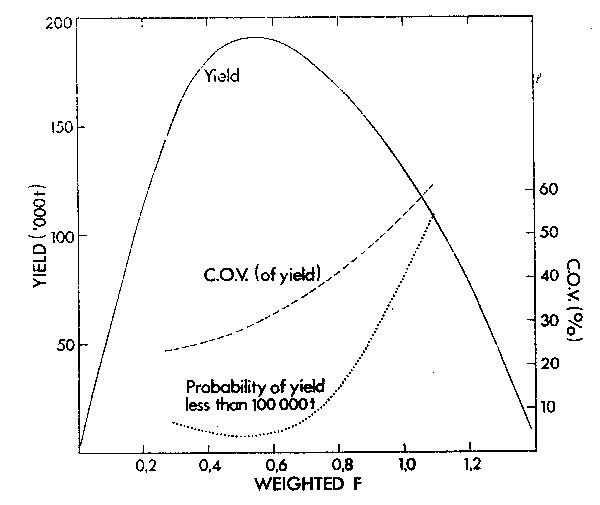
Figure 15. The long-term mean yield curve derived from 50 runs of the stochastic anchovy yield model over a 20-year time horizon. Also shown are the coefficient of variation of the yield and the probability of yield falling below 100 000 tons within 20 years.
CONCLUSIONS
The shape of the recruitment curve has a considerable influence on the average yield that can be taken, particularly at high levels of exploitation.
By examining the density-dependent population responses displayed at intermediate stock sizes, a preview of the shape of the recruitment curve at lower stock sizes can be obtained and used in a cautious approach to management.
In the South African pilchard compensatory changes in age at maturity and in gonad condition factor are important density regulatory responses. In addition, the inverse relationship between catchability and population size may be evidence that spawner density changes at a slower rate than spawner biomass (MacCall, 1980) and hence a conventional Ricker curve of recruits against spawner biomass may overestimate the severity of density-dependent mortality. The influence on the recruitment curve of a contraction in spawning range to the south after the decline of the pilchard is equivocal.
The incorporation of a compensatory change in fecundity into the generalized Ricker equation R = aNe-bNcrequires modification of the “a” term (corresponding to environment and fecundity) into a part (g) that is density-dependent (g=f(N)) and a part (a) that is density-independent. The rewritten equation takes the form R = agNe-bNc.
The combined effects of an inverse relationship between catchability and population size, a compensatory decrease in the age at maturity and a compensatory increase in fecundity will allow yield to be maintained up to a high level of fishing mortality but will precipitate a rapid collapse at a point only slightly above MSY. When depensation is superimposed on these effects the collapse is more rapid and recovery will be slower.
Reduced mortality of juveniles at a low population size due to, for example, a Type III predatory response, may give rise to a second stable equilibrium. The relative stability displayed by the South African pilchard population at a low biomass in recent years may be evidence to support the existence of a second equilibrium position.
REFERENCES
Armstrong, M.J., P.A. Shelton, R.M. Prosch and W.S. Grant. 1983. Stock assessment and population dynamics of anchovy and pilchard in ICSEAF Division 1.6 in 1982. Colln.scient.Pap.int.Commn.SE.Atl.Fish. 10:7–25
Bagenal T.B. 1973. Fish fecundity and its relation with stock and recruitment. Rapp.P.- v.Réun.Cons.int.Explor.Mer. 164:186–198.
Clark, C.W. 1974. Possible effects of schooling on the dynamics of exploited fish populations. J.Cons.int.Explor.Mer. 36:7–14.
Crawford, R.J.M. 1981. Distribution availability and movements of anchovy Engraulis capensis off South Africa. 1964–1976. Fish.Bull.S.Afr. 14:51–94.
Crawford R-J-M- and P.A. Shelton. 1981. Population trends for some Southern African seabirds related to food availability. In Proceedings of the symposium on Birds of the Sea and Shore. Cape Town, 1979. (J. Cooper, ed.). Cape Town: Southern African Seabird Group.
Crawford, R.J.M., P-A- Shelton, and D. Hutchings, 1980. Implications of availability, distribution, and movements of pilchard (Sardinops ocellata) and anchovy (Engraulis capensis) for assessment and management of the South African purse-seine fishery. Rapp.P.- v.Réun.Cons.int.Explor.Mer. 177:355–373
Csirke, J. 1980. Recruitment in the Peruvian anchovy and its dependence on the adult population. Rapp.P.-v.Réun.Cons.int.Explor.Mer. 177:307–313.
Gulland, J.A. 1977. The stability of fish stocks. J.Cons.int.Explor.Mer. 37(3):199–204.
Hewitt, R. 1981. The value of pattern in the distribution of young fish. Rapp.P.-v.Réuhn.Cons.int. Réun.Cons.int.Explor.Mer. 178:229–236.
Holling, C.S. 1973. Resilience and stability of ecological systems. Ann.Rev.Ecol.Syst. 4:1–23.
Jakobsson, J. 1980. Exploitation of the Icelandic spring-and summer-spawning herring in relation to fisheries management. 1947–1977. Rapp.P.-v.Réun.Cons.int.Explor.Mer. 1977:23–42.
Larkin, P.A., R.F. Raleigh and N.J. Wilimovsky. 1964. Some alternative premises for constructing theoretical reproduction curves. J.Fish.Res.Bd Canada. 21(3)477–484.
MacCall, A.D. 1980. The consequences of cannibalism in the stock-recruitment relationship of planktivorous pelagic fishes such as Engraulis. pp. 201–219. In IOC Workshop Report No. 28 (G.D.Sharp, ed.). IOC/UNESCO, Paris.
Newman, G.G. and R.J.M. Crawford. 1980. Population biology and management of mixed-species pelagic stocks off South Africa. Rapp.P.-v.Réun.Cons.int.Explor.Mer. 177:279–291.
Paloheimo, J.E. and A.M. Dickie. 1964. Abundance and fishing success. Rapp.P.- v.Rén.Cons.int.Explor.Mer. 155:152–163.
Paulik, G.J. 1973. Studies of the possible form of the stock-recruitment curve. Rapp.P.- v.Réun.Cons.int.Explor.Mer. 164:302–315.
Peterman, R.M., W.C. Clark and C.S. Holling. 1979. The dynamics of resilience: shifting stability domains in fish and insect systems. pp. 321–341. In Population Dynamics (Proceedings of the 20th Symposium of the British Ecological Society London 5–7 April 1978). (R.M. Anderson, B.D. Turner and L.R. Taylor, eds.). Blackwell, Oxford.
Shelton, P.A. and L. Hutchings. 1982. Transport of anchovy, Engraulis capensis Gilchrist, eggs and early larvae by a frontal jet current. J.Cons.int.Explor.Mer. 40:185–198.
Smith, P.E. and R. Lasker. 1978. Position of larval fish in an ecosystem. Rapp.P.- v.Réun.Cons.int.Explor.Mer. 173:77–84.
Ulltang, O. 1980. Factors affecting the reaction of pelagic fish stocks to exploitation and requiring a new approach to assessment and management. Rapp.P.-v.Réun.Cons.inst.Explor.Mer. 177:489– 504.
by
Bernard Einar Skud
U.S. National Oceanic and Atmospheric Administration
National Marine Fisheries Service
Northeast Fisheries Center
Narragansett Laboratory
South Ferry Road
Narrangansett, Rhode Island 02882
USA
Resumen
Los biólogos pesqueros han notado que las correlaciones entre factores físi- cos del ambiente y la abundancia de peces fallan con frecuencia. Las explicacio- nes para estas fallas han generalmente asumido que los datos para la comparación fueron inadecuados o que las relaciones fueron fortuitas o enganosas. El examen de las medidas de la abundancia y los factores ambientales de la caballa del Atlántico (Scomber scombrus) y la sardina de California (Sardinops sagax caerulea) indican que la falla de estas correlaciones occuren cuando las especies cambian del compoente dominante al componente subordinado de la biomasa total de pelágicos. Cuando especies que compiten, tales como el arenque del Atlántico (Clupea harengus) y la anchoveta de California (Engraulis mordax), tomaron la dominancia, su abundan- cia fue correlacionada positivamente con los mismos factores ambientales. La rela- ción para especies subordinadas fue negativa. Un cambio similar en la dominancia y una relación inversa con los factores ambientales fue reportada para gup (Poecilia reticulata) y para el Xiphoronorus maculatus en estudio experimentales. Aun cuando los cambios en dominancia no podrían explicir todas las fallas de las correlaciones, los estudios de la relación entre el ambiente y la abundancia deben considerar la densidad de las especies que compiten entre ellas y en particular la posición de las especies en la jerarquía de la dominancia.
INTRODUCTION
“All correlations fail eventually.” This facetious adage has been credited to Lionel A. Walford, but has been echoed by others who have examined the durability of correlations between physical factors of the environment and the abundance of fishes. Gulland (1978) remarked how few of these correlations had stood the test of time. Ware and Henriksen (1978) commented that“ …Every scientist knows that the path of fisheries research is littered with the bones of similar correlations that have been eventually proven spurious….” Akenhead (1981) said that, without a mechanistic model, such correlations can be expected to fail as conditions extended to the correlation change. Cushing (1978) summarized the situation as follows: “Although it is clear that climate, in a general sense, affects fish stocks, the nature and mechanisms of the relation are not well understood. Attempts at direct empirical correlations between environmental factors and year-class strength have seldom been successful because of the wide choice of factors and the limited number of observations … It seems that we need a deeper understanding of the various biological and physical events in the ocean and their links to the factors directly controlling year-class strength.”
I will discuss examples of these correlations that have failed. My intent is to show that these “failures” should not be summarily dismissed on the assumption that the data were faulty, that the criteria for comparison were ill-founded, or the relation spurious, rather that, with the usual contraints of accepting or rejecting an hypothesis, the results should stimulate the questions: How? and Why?
ATLANTIC MACKEREL AND TEMPERATURE
Taylor et al. (1957) correlated mackerel, Scomber scombrus, landings with air temperature data and reported a positive and highly significant relation from 1820 to 1890, but thereafter the relation was not significant. McHugh (1976) considered this 70-year correlation of particular note, because annual mean temperatures showed no particular trend prior to 1900, but did show a marked increase thereafter. Taylor et al. (1957) concluded that there was nothing in the temperature record to account for the precipitous decline of mackerel in the 1880's and the continued low abundance in the 1900's.
Bell and Pruter (1958) reexamined the mackerel data and compared temperature records from the open ocean (Riehl, 1956) with fluctuations in the landings from 1880 to 1930. Their results (r = -0.15) confirmed the insignificant relation for this period, but the authors questioned the methodology used by Taylor et al. (1957). Bell and Pruter stated:
“ … The authoritativestudy of Sette and Needler (1934) judiciously leaves the question of specific cause or causes of catch fluctuations in Atlantic mackerel unanswered. If a rela- tionship between temperature and mackerel is to be demonstrated, it seems that it should have been more noticeable during the recent period when temperatures rose to a new higher level than during the earlier years (1820–1890) when, although fluctuating, the long-term trend of temperature was relatively level.”
Following the lead of Lett and Kohler (1976), Winters (1976) and Ware and Henriksen (1978), who reported a competitive interaction between mackerel and herring, Clupea harengus, in the Gulf of St. Lawrence during the 1960's, Skud (1982) examined the relation of the two species prior to 1960 in the Gulf of Maine and in the Canadian maritime provinces. His results supported the findings of a competitive interaction in the Gulf of St. Lawrence. He found that landings of the two species were negatively correlated and observed that the decline of mackerel in the 1880's was coincident with an increase in the landings of herring. Based on the conclusion that the species were competitive, Skud (1982) reexamined the relations of landings with temperature. He confirmed the positive relation before 1900 (Taylor et al., 1957) and found a significant, negative correlation for mackerel from 1906 to 1960. During the same period, the relation between herring landings and temperature showed a significant and positive correlation. After 1960, when the abundance of herring declined and the abundance of mackerel increased to its highest level since the decline of the 1830's, the relation between abundance and temperature was negative for herring and positive for mackerel, however, the periods of dominance for each species was too short to test the significance of the correlations.
From this analysis and the study of other species, Skud (1982) concluded that in competing species, the dominant responded positively to changes in the environment that improved survival, and that the subordinate species responded negatively to the same factors. If dominance changed, the response of the two species also changed. He concluded that the dominant species control the abundance of the subordinate and therefore negated or masked the “normal” response of the subordinate to environmental change. He explained the failure of the mackerel-temperature relation reported by Taylor et al. (1957) as a change from the dominant to a subordinate pelagic species.
PACIFIC SARDINE AND SALINITY
A change comparable to that of the herring-mackerel complex also occured with the California sardine, Sardinops sagax caerulea, and anchovy, Engraulis mordax. As the abundance of larvae and stock size of sardines declined after 1940, the abundance of anchovy larvae and adults increased (Smith 1972). The deposition of fish scales in marine sediments showed similar fluctuations since 1940 and provided estimates of abundance for both species since 1810 (Soutar and Isaacs 1969). “Most if not all investigators have found that the hypothesis that the Pacific sardine and northern anchovy are direct competitors unavoidable. This hypothesis is not supported by the less-than-significant
positive correlation between the scale deposition of the two species in the Santa Barbara sediment…” (Soutar and Isaacs 1974).
Radovich and MacCall (1979) summarized the status of the research on the sardine- anchovy complex as follows:
“The hypothesis of interspecific competition from the Pacific sardine … has been evoked (Murphy 1966), seemingly verified (Silliman 1969), and set back (Soutar and Isaacs 1974). Smith (1972) showed that anchovy population to have been rela- tively large (between 2 and 3 million tons) in 1940 and 1941. Such a population size at a time when sardines were also abundant is utterly inconsistent with the competition hypothesis.”
Soutar and Isaacs' (1974) data showed that a third species, Pacific hake (Merluccius productus), dominated the pelagic biomass during most of the period of record, but its fluctuations in abundance were not considered in the interpretation of the correlation between sardine and anchovy. For example, the abundance of these two species increased from 1905 to 1925 (a strong positive correlation), when the abundance of hake declined 80%. This change suggests that the two species interact with hake and the long-term, positive correlation between sardine and anchovy may have occured because of the decline in the pelagic biomass. That is, total biomass may have a greater effect on the abundance of sardines and anchovy than the competition between them.
The size of the anchovy population in 1940 and 1941 was large relative to the years after 1925, but Soutar and Isaacs' (1974) data show that it was small relative to the period from 1810 to 1925. The sardine population in 1940 and 1941 also was small relative to earlier years. Comparable levels of abundance, when both populations and the total pelagic biomass were low, is not inconsistent with the hypothesis of competition as claimed by Radovich and MacCall (1979). Levels of abundance of two competing species will be comparable, at some intermediate level, whenever there is a change in dominance. The period most inconsistent with the hypothesis occured at the turn of the century when the combined abundance of anchovy and hake was approximately half of its maximum, and the high abundance of anchovy in the 1880's occured when both sardine and hake populations were low. For these reasons, and because all three species apparently interact, I do not agree that the hypothesis of competition can be dismissed.
Bogdanov (1968) also reviewed the sardine-anchovy complex. He made particular reference to the positive correlation (+0.845) between salinity and the recruitment of sardines from 1934 to 1941 (Walford 1946), and the negative correlation (-0.639) for the same factors from 1942 to 1955 (Marr 1960). The change was assumed to be evidence of a fortitous relation except by Bogdanov (1968), who considered this change to be of paramount importance and extensively reviewed the relations of salinity, upwelling and temperature. Although he did not resolve the question, his astuteness merits recognition, because the change in the correlation was associated with the change in dominance. Recruit- ment of sardines declined steadily after 1940 and anchovies attained dominance of the pelagic biomass during the latter period.
The change in the relation of the abundance of sardines to salinity was also apparent in the relation to temperature. In the earlier papers that discussed the cause for fluctuations in the abundance of sardines, I was unable to locate any specific reference to a correlation with temperature. As this is the environmental factor that is most frequently used in such studies, I assume that attempts were made, but that results showed a negative relation and, therefore were not reported. Using data from Radovich (1961), Ahlstrom (1965) and Murphy (1977), I compared the catch and catch-per-unit effort (CPUE) with sea-surface temperatures (Table 1). The responses of both species to temperature were inverse to that of salinity. These two factors are associated with upwelling and are themselves inversely related (Bogdanov, 1968). Bakun (1973) computed annual upwelling indices for the California coast (36°N, 122°W). The correlations of these indices with the abundance of sardine and anchovy larvae from 1951 to 1959 were -0.630 and +0.592, respectively. The inverse relation with the change in dominance support the thesis formulated from the herring and mackerel data (Skud 1982).
| Dominant Species | Sardine | Anchovy | ||||
|---|---|---|---|---|---|---|
| CPUE | Catch | CPUE | Catch | |||
| Adults: | ||||||
| 1932–1939 | (8) | S | -.421 | -.410 | +.353 | |
| 1940–1947 | (8) | A | +.649 | +.520 | -.655 | |
| 1941–1959 | (19) | A | +.863** | -.560* | ||
| Larvae: | ||||||
| 1951–1959 | (9) | A | -.710* | +.801** | ||
I also explained Sillimans's (1975) experimental studies on guppy(Poecilia reticulata) and swordtail (Xiphorphorus maculatus) in which he recorded the actual num,bers and biomass of each species. The relation of the two species to temperature was inversed and changed when dominance changed.
DISCUSSION
I concluded that a change in dominance explained the failure of the mackerel- temperature and sardine-salinity correlations (Skud 1982) and I think dominance is one of the unknown mechanisms which Cushing (1978) said was needed to understand the factors controlling year-class strength. As a corollary to the statement by Backiel and LeCren (1967)“…that studies of environmental influences on fish growth are of little relevance without data on population density…”, I conclude that studies of the relation of environmental factors to abundance for a given species should always consider the density of the competing species, i.e., the social hierarchy.
Not only may a change in dominance explain a change in the correlation with an environmental factor, it may also explain why the indices of abundance of the same or similar species are positively correlated in one area and not another, for example herring and mackerel in the Gulf of Maine and in the Gulf of St. Lawrence after 1960; and for cod and haddock across the North Atlantic (Carrod and Colebrook, 1978). Dominance may also explain certain observations concerning exploited stocks and the effects of fishing. Austin and Ingham (1978) echoed the conclusions of other scientists stating that”
… A heavily fished or overfished stock is more susceptible to environmental fluctuations than is a more stable or underutilized stock….” I suggest that a heavily fished stock is likely to be the dominant species and is more responsive to environmental change because it is below its equilibrium density, whereas the underutilized stock is subordinate and its fluctuations in abundance are controlled by the dominant species rather than environmental factors.
I assume that multispecie interactions, rather than two-species complexes, are the norm and that the other species, including predators, also can influence the apparent relation of environmental factors to abundance. Thus in modelling a multispecies complex, it would be necessary to incorporate the effects of a change in dominance from one species to another, along with interactions such as predation and an array of environmental factors.
Just as comparisons between species abundance and environmental factors should be made in concert with other species, so should comparisons of species abundance. Correlation matrices such as presented by Dow (1977), Sutcliffe et al. (1977), Garrod and Colebrook (1978) and Sissenwine et al. (1979) fulfill this basic need, but it is necessary to determine the position of dominance within each trophic level to interpret the meaning of each correlation and of correlations with like and different signs. Without the determination of dominance, correlations of different signs cannot be assumed to represent different responses to an environmental factor or an indication of competition, nor can correlations of a similar sign be assumed to imply similar responses. For example, differences in dominance may explain, as Garrod and Colebrook (1978) showed, why the recruitment of a species is positively correlated with the environment in one geographic area and negatively in another, or why recruitment of species with totally different early life histories had a strong affinity to a given environmental variable.
This varying response of a species probably has contributed to the polarization of the scientific community into two camps as described by Dickie (1973); one crediting major changes in yield to exploitation, the other, relating these changes to the environment. Part of the problem, as Daan (1980) pointed out is that well-documented cases of species replacement refer to heavily exploited populations, so that the effects of fishing are superimposed upon and can never be well-separated from environmental effects. Determination of dominance should help to separate these effects and properly identify species interactions and perhaps, to define the operative mechanisms that have been so elusive.
The effects of interactions discussed in this paper reinforce Burkenroad's (1951), McHugh's (1959), and others' suggestions for an ecosystems approach. And my message is simply that if a long-term significant correlation fails, it is not necessarily because the measure of abundance was inadequate or the hypothetical relation was not biologically justified, etc. The change should be a signal to reexamine the data and to examine factors that could have influenced the relation. Correlation analysis can be a useful tool, it should not be abandoned or condemned, if the results do not support the original hypothesis. The cautions that Bell and Pruter (1958), Austin and Ingham (1978), Akenhead (1981), and others have expressed concerning correlation analyses and their interpretations still pertain. However, the effects of dominance show that the loss of significance or a change in the response is not acceptable evidence that an environmental factor did not affect abundance or that the observed relation was spurious. Nor is it valid, before considering the social hierarchy, to dismiss a correlation for one species when another, in the same trophic level, has not shown a similar response or if the same species when another, in the same trophic level, has not shown a similar response or if the same species did not respond in the same way in a different area. These conclusions do not imply that all correlations that fail are the result of a change in dominance, rather, they support Bogdanov's (1968) approach. He considered the change in the relation between salinity and the abundance of the California sardine to be one of particular significance and although he did not resolve the change, he attempted to explain the phenomenon, rather than assuming that there was a problem with the data or that the relation was fortuitous.
REFERENCES
Ahlstrom, E.H. 1965. A review of the effects of the environment of the Pacific sardine. Int.Comm.Northw.Atl.Fish.Spec.Publ. 6:53–74.
Akenhead, S.A. 1981. Fisheries Oceanography and the nature of carrying capacity for larval marine fishes. Northw.Atl.Fish.Org. SCR Doc. 81/IX/142, 27 p.
Austin, H.M. and M.C. Ingham. 1978. Use of environmental data in the prediction of marine fisheries abundance. pp. 93–89. In Climate and Fisheries. Univ. R.I., Center for Ocean Management Studies.
Backiel, T. and E.D. LeCren. 1967. Some density relationships for fish population parameters. pp. 261–293. In The Biological Basis of Freshwater Fish Production (S.D.Gerking, ed.). John Wiley and Sons.
Bakun, A. 1973. Coastal upwelling indices, west coast of North America, 1946–71. U.S.Nat.Mar. Fish.Serv. Tech.Rep.SSRF-671:103 p.
Bell, F.H.and A.T. Pruter. 1958. Climatic temperature changes and commercial yields of some marine fisheries. J.Fish.Res.Bd.Canada. 15(4) : 625–683.
Bogdanov, G.A. 1968. Factors governing the reproduction of certain sardines. Prob.Ichthy.(USSR). 8(5):695–704.
Burkenroad, M.D. 1951. Some principles of marine fishery biology- Publ.Tex.Inst.Mar.Sci. 2(1):177– 212.
Cushing, D.H. 1978. Climatic variation and fisheries. In Problems and Progress in Oceanography Relevant to Fisheries (J.A. Gulland, ed.) FAO Fish.Rep. (206)(Supplement l) Appendix 2:18–19.
Daan, N. 1978. A review of replacement of depleted stocks by other species and the mechanisms underlying such replacement. J.Cons.int.Explor.Mer. Symposium on the biological basis of pelagic fish management, No. 24. Rapp.P-v.Réun.Cons.int.Explor.Mer. 177:405–423.
Dickie, L.M. 1973. Interaction between fishery management and environmental protection. J.Fish.Res.Bd.Canada. 30(12):2496–2506.
Dow, R-L- 1977. Effects of climatic cycles on the relative abundance and availability of commercial marine and estuarine species. J.Cons.int.Explor.Mer. 37(3):274–280.
Garrod, D.J. and J.M. Colebrook. 1978. Biological effects of variability in the North Atlantic Ocean. Rapp.P.-v-Réun. Cons.int.Explor.Mer.173:128–144.
Gulland, J.A. 1978. Problems and progress in oceanography relevant to fisheries. FAO Fish.Rep. (206)(Supplement 1) : 9–21.
Lett, P.F. and A.C. Kohler. 1976. Recruitment: A problem of multispecies interaction and environmental perturbations with special reference to Gulf of St. Lawrence Atlantic herring (Clupea harenqus harenqus). J.Fish.Res.Bd.Canada. 33(6):1353–1371.
Marr. J. 1960. The causes of major variations in the catch of the Pacific Sardine Proc. World Sci. Meeting. Biol.Sardines. FAO III:667–791.
McHugh, J.L. 1959. Can we manage our Atlantic coastal fishery resources? Trans.Am.Fish.Soc. 88(2):105–110.
McHugh, J.L. 1976. Effects of climatic change on fisheries. Sea Grant Reprint Series. Mar.Sci.Res.Con., St. Univ. N.Y. at Stony Brook Contrib. No. 177:8 p.
Murphy, G.I. 1966. Population biology of the Pacific sardine (Sardinops caerulea). Proc.Calif.Acad. Sci., Ser.4. 34(1):84 p.
Murphy, G.I. 1977. Clupeoids. pp. 283–308. In Fish Population Dynamics (J.A. Gulland, ed.) J.Wiley, N.Y.
Radovich, J. 1961. Relationships of some marine organisms of the Northeast Pacific to water temperature. Cal.Fish. Game, Fish. Bull.112, 62 p.
Radovich, J. and A.D. MacCall. 1979. A management model for the central stock of the northern anchovy, Engraulis mordax. Calif. Coop. Oceanic Fish. Invest. Rep. XX : 83–88.
Riehl, H. 1956. Sea surface temperatures of the North Atlantic, 1887–1936. Proj. NR 082– 120, Office of Naval Research. Univ. of Chicago, Dept. of Meterology. 129 p.
Sette, O.E. and A.W.H. Needler. 1934. Statistics of the mackerel fishery off the east coast of North America, 1804–1930. U.S.Bur.Fish., Investig.Rep.No.19.48 p.
Silliman, R.P. 1969. Population models and test populations as research tools. Bio. Science. 19 : 524– 528.
Silliman, R.P. 1975. Experimental exploitation of competing fish populations. Fish.Bull. U.S. 73(4) : 872–888.
Sissenwine, M.P., B.E. Brown, J-E- Palmer and R.J. Essig. 1979. An empirical examination on population interactions for the fishery resources off the Northeastern USA. Workshop on “Multi-species fisheries management advice.” St. John's, Newfoundland. 27 pp. (Mimeo).
Skud, B.E. 1982. Dominance in Fishes. The relation between environment and abundance. Science. 216:144–149.
Smith, P.E. 1972. The increase in spawning biomass of northern anchovy, Enqraulis mordax. Fish.Bull.U.S. 70:849–874.
Soutar, A. and J.D. Isaacs. 1969. History of fish populations inferred from fish scales in anaerobic sediments off California. Calif.Coop.Oceanic Fish.Invest. Rep. 13:63–70.
Soutar, A. 1974. Abundance of pelagic fish during the 19th and 20th centuries as recorded in anaerobic sediment off the Californias. Fish.Bull.U.S. 72(2):257–273.
Sutcliffe, W.H., Jr., K. Drinkwater and B.S. Muir. 1977. Correlations of fish catch and environmental factors in the Gulf of Maine. J.Fish.Res.Bd.Canada. 34(1):19–30.
Taylor, C.C., H.B. Bigelow and H.W. Graham. 1957. Climatic trends and the distribution of marine animals in New England. U.S.Fish.Wildl.Serv.Fish.Bull. 57(115):289–345.
Walford, L. 1946. Correlation between fluctuations in abundance of the Pacific sardine and salinity of sea water. J.Mar.Res. VI(1):48–53.
Ware, D.M. and B.L. Henriksen. 1978. On the dynamics and structure of the Southern Gulf of St. Lawrence herring stocks. Can.Fish. and Mar.Ser. Tech.Rep.No. 800:83 p.
by
B.J. Rothschild
and
D.G. Heimbuch
University of Maryland
Chesapeake Biological Laboratory
Box 38, Solomons
Maryland 20688
USA
Resumen
La extensión de la jurisdicción de los países costeros ha hecho que se ponga un mayor énfasis en la ordenación de pesquerías. A pesar de este énfa- sis todavía se han explorado muy poco los modelos que toman en consideración todo el proceso de ordenación en términos de la incertidumbre de la relación stock-reclutamiento, la información requerida para la ordenación, y la estre- cha interrelación entre la información para la ordenación y el funcionamiento de la ordenación misma. En este trabajo se desarrolla un modelo basado en estos elementos.
El trabajo describe los elementos fundamentales en la toma de decisiones (un evento casual, una acción de manejo, el saldo de una cuenta, etc) y mues- tra cómo estos pueden ser conectados para constituir una estrategia de manejo. El trabajo muestra como la estrategia de manejo y el desempeño del manejo pue- den ser afectados a través de la obtención de información adicional; se pro- porciona un esquema para evaluar los beneficios de información adicional (por ejemplo, proveniente de reconocimientos de campo y el uso de correlaciones ambientales) relacionadas a los costos de la información adicional.
Se pueden seleccionar estrategias de manejo alternativas por medio de varios criterios. Aquéllos considerados en este trabajo son el máximo y mínimo, valor monetario esperado y utilidad.
El trabajo muestra la construcción y evaluación de diez diferentes estre- gias; estas se diferencian en si son normativas o adaptables, en si toman o no en cuenta colapsos catastróficos de los stocks, y si toman en cuenta mues- treos intensivos o limitados o programas de investigación. De momento, el principal valor práctico del modelo es en (1) demuestra la interrelación ínti- ma de la performance de la ordenación y la información (2) proporciona las bases para el diseño de un sistema de manejo e información de la pesquería sobre una base cuantitativa y (3) proporciona los medios por los cuales la adquisición de información y toma de decisiones de la ordenación de pesquerías pueden ser armonizadas.
INTRODUCTION
The advent of extended jurisdiction has generated considerable concern for the management of fisheries and considerable rhetoric on the nature of fisheries management and its objectives. Most of the concern, however, has not focused on management per se (see, FAO 1980). Rather, it has focused on various important, but relatively specialized topics such as optimum yield versus maximum sustained yield, data requirements for management, multiple-species management, preventing stock collapses, satisfying economic and social development requirements of small-scale fisheries, and development of various economic approaches to management, such as fishing-effort limitation. Unfortunately, the studies and activities relating to these topics have not been undertaken in concert; rather each has been emphasized at various times and this has detracted from developing an approach which considers management as an entity in its own right.
Because of this, the important technical topics enumerated above have tended to become side issues and management remains a political process that operates with only a very limited knowledge base. As a result, the long chain of management decisions tends to be disjointed, oriented toward no particular purpose, and inefficient with respect to yielding benefits to society.
Obviously, recovery from this state of affairs will require conceptual foundations for linking together all management decisions. The greatest utility of this conceptual foundation will be to provide a basis for interrelating information needs, the multiple decisions that need to be made in an uncertain environment, and the alternative outcomes of the decision process. A conceptual foundation such as this is of critical importance because it will define fishery management where it has not been heretofore defined. The definition will enable all concerned to have an improved perception on “just what fishery management is”, but more importantly it will provide a quantitative framework for allocating budgetary funds to various parts of the decision making activity.
Development of the management process needs to be undertaken in the context of a management information system. The information system may be considered as a series of activities. The activities are 1) obtaining bioeconomic data on the operation of the fishery, 2) synthesizing these data into reports or other forms of information, 3) constructing “regulations” based upon the information, 4) executing or implementing the regulations, 5) engaging in enforcement activity and 6) evaluating the entire process (viz, activities 1–5). Figure 1 is a diagram showing a hypothetical time- sequence of events that occur in each activity. It is the task of the manager to identify the various events, to decide upon the human and budgetary resources that will be committed to each, and to schedule each event so that the entire set of activities works in the most cost-effective fashion.
In observing many fishery-management operations, we can note that these management activities are often not well defined and the various events in management occur as ad hoc reactions. The reactive mode of management and its lack of definition makes it difficult to isolate problems which are recurrent and therefore it is hard to achieve solutions. Because of this management is expensive and does not appear to work well.
Thus, we see the potential of considerable improvement in the management process through designing systems that explicitly involve the six activities outlined above and the events that are components of each activity. These events and activities would naturally need to be responsive to and interacting with the great chance uncertainties that are associated with any fisheries management process. These include, inter alia, the year-to-year fluctuations in fish stocks, the collapses and explosions, and whether or not a stock that is heavily fished has a propensity to increase in abundance with a reduction in fishing.
While the establishment of the six activities is a relatively easy task, the identification of the events, the analysis of the costs-and-benefits of each, the proper time-placement of each event in its activity, and the coordination among activites is a much more complex task. Fortunately, however, there are a variety of operations research techniques that can facilitate the design of a management information system.
Fig. 1. Six management activities, obtaining data, developing information, constructing regulations, implementing regulations, enforcement and evaluation. The X's refer to hypothetical events in the time stream. Important tasks of management are to identify these events, the magnitude of human and budgetary resources associated with each and their placement in time.

These include program evaluation and review techniques (PERT) and mathematical programming. But there is one technique that seems particularly suitable in the sense that it has a natural relationship to the process of making a sequential decision in time in an uncertain environment. This technique is decision theory.
In this paper we lay a decision-theoretic foundation for the design of a fishery management system. We begin by discussing the fundamental structure of the decision problem taking into account a) chance events, b) decision-making strategies, and c) consequences or payoffs. We then show how these may be employed in various management philosophies (i.e. perfect-knowledge management, maxi-min management, expected-monetary-value management, and expected-utility management. Next, we show how various sampling schemes can improve the management process and give examples which show how comparisons might be made among different management situations such as, prescriptive or adaptive strategies, the timing involved in estimating recruitment, taking into account or ignoring the possibility of catastrophic stock collapses, various sampling distributions and costs for field programs.
FUNDAMENTAL STRUCTURE OF DECISION PROBLEM
As implied earlier the activity of management can be characterized as a series of decisions; decision theory provides a context for analyzing these decisions.
The typical decision problem in ts simplest form requires the specification of three components. These are 1) the chance events about which a decision is made, 2) a list of decisions or actions that will be undertaken before observing the chance event and 3) the consequences given the particular chance event and the action that is taken.
A) Chance Events A chance event is an event that occurs as the result of a random or chance process. Examples of chance events are the occurrence of a head in a flip of the coin or drawing an ace from a deck of cards. The most evident chance event in fish-population dynamics is the magnitude of year-class strength. With respect to year-class strength, the chance process may be thought of as being motivated by Nature. In effect Nature selects the size of the year-class, O, according to some probability distribution. If the decision problem involves a single year, then the outcome of the chance process might be described in terms of a good year class O=G, a poor year class O=P, and a catastrophically poor year class O=C. If, however, the decision problem considers a sequence of years, say N years, then the outcome would be of the form
O = (O1 = i, O2 = i, …, On =i)
where i = G, P or C (Note that O is sometimes called Nature's strategy). The linkages among the O1's are obviously an important part of the problem. The O1's may be independent of one another or they may be dependent. The simplest form of dependence would occur if there were a stock-recruitment relationship. Such a relationship would imply that the recruitment in any year would depend upon recruitment in previous years and the amount of fishing in those years.
B) The Decision Makers Strategies The decision maker attempts to guess O in a single year problem or O in a multiple year problem. He then undertakes some action or set of actions. This action may be denoted by A for a single year or
A = (A1, A2, …, AN)
for the multiple N-year problem. A is frequently called a strategy.
Strategies may involve sampling. In the context of decision analysis, “sampling” refers to undertaking a survey, or a field program, or some other method of acquiring additional information regarding the system with the intent of reducing uncertainty and improving the decision process. It is obvious that if sampling is considered, then its cost and the value of its benefits will be an important part of the analyses.
Strategies may be prescriptive or adaptive. A prescriptive strategy is one in which each year's actions are determined in the first year and an adaptive strategy is one in which each year's actions are determined sequentially each year based upon observations of the stock in that year or in previous years.
For the purpose of this exposition the actions that comprise strategies involve setting the magnitude of fishing mortality in each year. Thus A=H would refer to high fishing mortality, A=L would refer to low fishing mortality and A=Z would refer to zero fishing mortality.
C) Consequences or Payoffs Generally speaking a O and A (where the number of elements in O and A may each number from 1 to N ) will determine a particular payoff. Payoffs are usually measured in monetary units. Intuitively, it is easy to think that sustained heavy fishing on small year classes would result in low payoffs. An example of an integration of these three components in a decision tree is shown in Figure 2.
Figure 2 shows, for example, that if the year class is good (O=G) and the manager decides to fish heavily, then the payoff will be 25 monetary units. (The payoffs were chosen for demonstration purposes, but are intuitively of the right magnitude).
An important simplifying assumption should be mentioned at this point. That is, if in reality fishery problems were independent single-year problems, then it would be most profitable to catch as much fish as possible. This is of course not really the case since there would be no individuals left to spawn - so the example here is really intended to reflect what would happen with very short lived fish and the payoff includes the maintenance of an appropriate size spawning stock.
THE MANAGEMENT PROCESS
The preceding discussion developed the notion that the decision making process - that is, the process of management - involves listing Nature's strategies, the decision makers strategy, and the payoffs. But how does one select the right action or strategy? In this section we discuss the question of selecting the right action or strategy and how the strategy might be improved by increasing information on nature's strategy through sampling. First the problem is considered as a single-year problem to demonstrate the methodology and then it is considered as a multiple year problem.
Perfect knowledge management
If in fact O were known - that is, if it were known in advance whether a year class was G, P, or C, then the correct course of action would be obvious (Figure 2). That is
a) If O=G, choose H and gain 25 units.
b) If O=P, choose L and gain 5 units.
c) If O=C, choose Z and gain -5 units.
Each action would thus, given that O is known, maximize the return. But, as it is in the real world of fisheries O is never known with certitude. Thus, procedures need to be applied which take into account the uncertainty that is associated with O.
Maxi-min management (information not available)
If O is not known, then one approach to selecting A is to pick that A that results in the greatest minimum value for all possible O. Choosing this action protects the manager against the worst possible outcome.
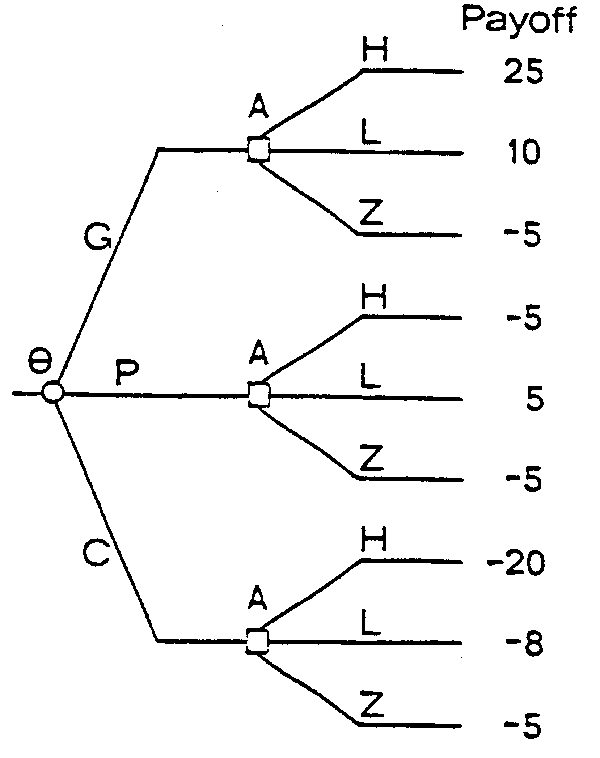
Fig. 2. Example of decision tree showing integration of the chance event, O, the action or strategy, A, and the payoff in numerical monetary units.
For example, referring to Figure 1, we see that for A=H, heavy fishing, the minimum value is -20; for A=L, the minimum value is -8; and for A=Z, the minimum value is -5. Thus under maxi-min management the action, Z, would be taken and, taking the action Z ensures that whatever O is, the maximum loss will be -5.
Expected monetary value (EMV) Management
Intuitively we can see that if we had some idea regarding the probability of each possible outcome of O, then we ought to be able to do better than we would protecting ourselves against the absolutely worst case as when the maxi-min criterion is used. Suppose we have some means of knowing that O=C is a relatively rare event and that the possible outcomes have the following probabilities the weighted average value of each action or the expected monetary value of each action is given by
Pr (O=G) = 0.45
Pr (O=P) = 0.45
Pr (O=C) = 0.10
E (V A=H) = .45 (25) + .45 (-5) + .10(-20) = 7.0
E (V A=L) = .45 (10) + .45 (-5) + .10(-8) = 5.95
E (V A=Z) = .45 (-5) + .45 (-5) + .10(-5) = -5.0
Employing the EMV criterion in this case, one would choose the action, H, exerting a maximum amount of fishing effort because this would maximize the average return from fishing.
Expected Utility Management
The result which advises us to maximize fishing effort may seem curious, but it is not. This is because the result contains only part of the information in the decision problem. That is each action may be met with responses of different variability. One measure of this variability is variance about the expected monetary value.
Var (V|A=H) = 283.5
Var (V|A=L) = 27.2
Var (V|A=Z) = 0
One can see that if action H is chosen then the results will be met with high interannual variability. On the other hand if one were to choose action Z the results would not differ from year to year. Thus the manager is faced with the question: Should he pick action H and maximize expected return, but suffer the high probability that in some years he might have disasterous results while in other years he might have very good results or, on the other hand, should he take action L in which he would not make as much on the average, but what he would make would be relatively more certain. Unless he practiced maxi-min management he would not choose Z, because in this particular case a loss of 5.9 units could be guaranteed. The problem now becomes a little more difficult because there is no obvious solution as to whether to choose H with a high EMV and a high uncertainty in payoff or to choose L with a lower EMV which is more certain.
The tradeoff between uncertainty in payoff and expected monetary value is conventionally assessed in terms of utility. The notion of utility is generally referenced to the notion of a lottery. Each lottery is comprised of a series of tickets. Each ticket in the lottery has a certain payoff and a probability of winning associated with it. Suppose you were offered a ticket with a $10 payoff and a 20 percent chance of winning. To break even, on the average, you would expect to pay $2.00 for the ticket. If you pay more for the ticket you are risk prone and if you pay less for the ticket you are risk averse. The utility, U(V), corresponding to a value, V, is the probability of winning the lottery that leaves the decision maker (that is the buyer of the ticket) indifferent between buying the ticket and keeping the value as a certain outcome.
Utility functions can be constructed to represent almost any attitude toward risk. Three general categories of attitude toward risk are risk averse, risk neutral and risk prone. All risk averse utility functions are concave, risk neutral utility functions are straight lines and risk prone utility functions are always convex (Figure 3a). In addition to these general categories, attitudes toward risk can be classified further based on how risk aversion (or proneness) is affected by asset position. Three such classes are constant, increasing and decreasing risk aversion (proneness). All utility functions that describe constant risk aversion are of the form
u (v) = a - beV (1) (Keeney and Raiffa 1976)
For convenience, we will restrict subsequent discussions on utility to utility functions representing constant risk aversion.
The parameters, a and b, in equation (1) scale the utility function and the parameter , represents the strength of risk aversion. Larger values of represent greater aversion to risk and generate more bow-shaped utility functions (Figure 3b). Typically u(V) is scaled to the interval 0, 1 such that u(Vmin)=0 and u(Vmax)=1.
with
Vmin)=-20
Vmin)=25
and arbitrarily set to .05
u(u)=1.1 (1-e-.05(v+20)
See Figure 2
IMPROVING KNOWLEDGE ON NATURE'S STRATEGY
Under perfect-information management, 0 was known and the correct decision could always be made. The perfect-information situation does not exist, but it is useful for comparative purposes. Under maxi-min management, 0 did not have to be known, but the range of payoffs for each action was required. The fact that 0 is not known under maxi-min management results in a very conservative approach. Under EMV management, information on the probability of any particular 0 was required and the utilization of information on 0 enabled a more finely-turned criterion. The major problem with EMV is that decision makers may vary in their assessment of the worth of a particular EMV; this being particularly true when the variance of any payoff varies with the amount of the payoff. Utility theory accounts for this, but it requires the difficult analysis of attitudes toward risk.
A key question in the EMV or expected utility process involves whether or not information on 0 should be obtained and if it is obtained how precise should this information be. The degree to which 0 is understood is a central issue in improving the performance of management. Mechanisms for understanding 0 are of considerable practical importance because they may involve complicated long-term research or extensive at–sea sampling programs.
Deciding on how to investigate 0 is not generally a simple problem that a large element in improving the performance of management is to increase our knowledge of 0. There are in most instances numerous approaches to increasing knowledge about 0. While knowledge on 0 generally can be increased at some cost, not all increases in knowledge on 0 will be cost effective.
Perhaps, the simplest way of estimating 0 is to assume that causal factors have remained stable over the years and hence use past information to estimate 0 for the future. Par exemplo, suppose a fishery was observed for twenty years and there were 10 G year classes, 8 P year classes and 2 C year classes. The following estimates could be made; Pr (0=G) = 10/20, Pr (0=P) = 8/20, Pr (O=C) = 2/20. However, an assumption implicit in the above procedure is that the underlying mechanisms which cause large and small year classes is the same year–in–and–year–out. That is, in the future, Pr (0=G) is very close to 10/20; Pr (0=P) is very close to 8/20 and Pr (0=C) is very close to 2/20. To the intuition, it seems unlikely that conditions will stay the same for a number of years and hence it is desirable to collect and employ additional information.
The notion that conditions stay the same over years implies that there would be no discernable time trend in recruitment. Hence, the recruitment that would be predicted each year, is just the average recruitment and the variability associated with that prediction would be relatively large. This variability could be reduced if it were known that there was some independent variable or set of independent variables which recruitment would depend upon. This one would still need to assume that the relationship between recruitment and the independent variables would need to be the same. If as we suspect this relationship changes or the set of variables changes then the quality of the predictions could substantially deteriorate.
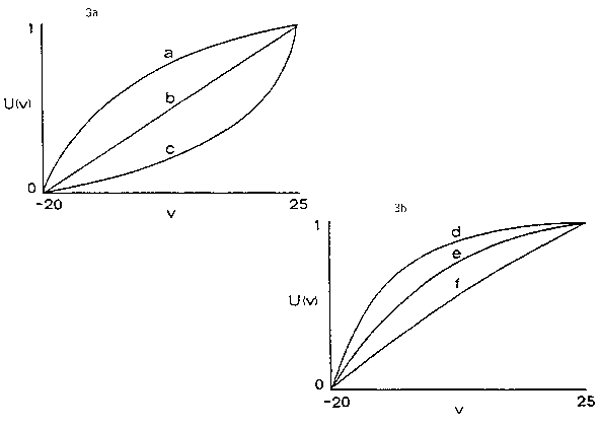
Fig. 3. Hypothetical utility functions. The ranking of actions based on expected utility, E[u(v)], depends on the shape of the utility function - both the general class of function and particular parameters affect that shape. The curves in Figure 3a are examples of risk averse (a) risk neutral (b) and risk prone (c) utility functions. The curves in Figure 3b are all constant risk averse utility functions: = 1 in d, = .01 in f and =.05 in e (the example discussed in the text).
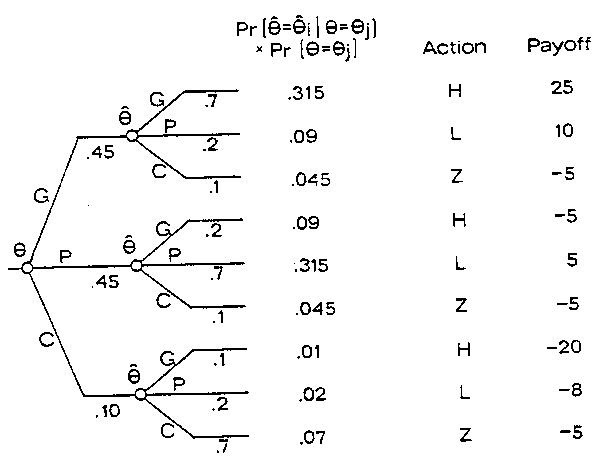
Fig. 4. A decision tree depicting the affect of sampling and associated decision rules on the probability distribution of payoffs. The decimal number below each branch is the probability of the corresponding chance event. For example, Pr(0=C)=.1 andPr (θ=P|0=C)=.2, therefore the joint probability for the history, 0=C and θ=P, is .02. The decision rules represented are if θ=G then A=H, if θ=P then A=L and if θ=C then A=Z.
The question of acquiring additional information and assessing its value can be determined using decision theory, the use of which is facilitated by a decision tree. Figure 4 shows a decision tree which would be characteristic of the fisheries management process. The particular tree shown in the figure gives the basic elements: Nature selects 0, a scientific program samples the system to guess 0, the manager makes a decision and then nature selects another 0.
Obviously, the manager will want to improve his ability to guess 0. Therefore, he should want his probability of detecting the true 0, i.e. pr(Ô = Ô) to be high and the probability of making a mistake, Pr (Ô = Ôi!O=Oi), ij to be low. However, the value of the estimation process on how the estimates affect the choice of an action as well as how good the estimates are.
Suppose the decision rules are if Ôt= G do At= H; if Ôt= P do At= L; and if Ôt=C do At=Z (Figure 4). The EMV for this combination of estimation and choice is 8.74. The best that could be done with the same probabilities for 0 but no sampling was an EMV = 7 for A=H. So that sampling scheme would result in an expected increase of 1.74 in the EMV over a program where sampling is not utilized. A similar analysis could be made for expected utility.
THE PROBLEM OF TIME AND MULTIPLE YEAR DECISION ANALYSIS
A time horizon is a point in time beyond which the benefits of planning do not excede the costs, usually because of uncertainty into the future. While time horizons in fisheries may not be long, they are probably longer than one year in most instances and hence it is important to consider multiple year decision analysis.
Multiple year decision analysis requires considering simultaneously the set of O's, one for each year and the set of possible actions that might arise in response to a particular set of O's.
The linkage among O's can be represented by transition probabilities Pr (OtOt-1, At-1,Ot-2,At-2,…). For the purpose of this elementary exposition we only consider probabilities of the form Pr (OtOt-1, At-1) which relate recruitment in year t with the recruitment and the action in year t-1.
In order to demonstrate the procedure we have contrived three stock recruitment hypotheses (Table 1). H1 represents no relationship between stock and recruitment and no chance of a catastrophic decline. H2 represents a positive monotonic stock and recruitment relationship with no chance of a catastrophic decline and H3 represents a monotonic stock and recruitment relationship with a small chance of a catastrophic decline.
In a multiple year setting the multiple-actions can also be linked together. Each set of multiple actions is called a strategy (for example a four year strategy would be (A1,A2,A3,A4). There are two approaches to linking the strategies together.
The first approach is prescriptive. In other words the alternative strategies are decided before the management routine is implemented. For example, within a three year time horizon it would be decided to engage in heavy fishing each year so A = (H, H, H). Table 2 gives two examples of a prescriptive strategy.
The second approach is adaptive. In this second approach the action in any year depends upon sampling the system to improve the guess of O. So an adaptive strategy requires specification of 1) the sampling procedure, 2) the method of estimation and 3) the decision rule which will give the alternative action based upon the estimate. Tables 3–10 give examples of adaptive strategies.
| Pr(θtθ|θt-l&At-1) | |||||
|---|---|---|---|---|---|
| θt | |||||
| Hypothesis | θt-1 | At-1 | G | P | C |
| 1 | G | H | .50 | .50 | .00 |
| L | .50 | .50 | .00 | ||
| Z | .50 | .50 | .00 | ||
| P | H | .50 | .50 | .00 | |
| L | .50 | .50 | .00 | ||
| Z | .50 | .50 | .00 | ||
| C | H | .00 | .00 | .00 | |
| L | .00 | .00 | .00 | ||
| Z | .00 | .00 | .00 | ||
| 2 | G | H | .60 | .40 | .00 |
| L | .70 | .30 | .00 | ||
| Z | .80 | .20 | .00 | ||
| P | H | .20 | .80 | .00 | |
| L | .30 | .70 | .00 | ||
| Z | .40 | .60 | .00 | ||
| C | H | .00 | .00 | .00 | |
| L | .00 | .00 | .00 | ||
| Z | .00 | .00 | .00 | ||
| 3 | G | H | .60 | .38 | .02 |
| L | .70 | .28 | .02 | ||
| Z | .80 | .18 | .02 | ||
| P | H | .15 | .80 | .05 | |
| L | .25 | .70 | .05 | ||
| Z | .35 | .60 | .05 | ||
| C | H | .02 | .08 | .90 | |
| L | .02 | .18 | .80 | ||
| Z | .02 | .28 | .70 | ||
| Strategy | t(year) | |
|---|---|---|
| 1 | 1 | H |
| 2 | H | |
| 3 | H | |
| 2 | 1 | L |
| 2 | L | |
| 3 | L |
3a
| Strategy | θt | θt–1 | |
|---|---|---|---|
| 3 | G | G | H |
| P | L | ||
| P | G | H | |
| P | L |
3b
| Pr(θt|θt& At | |||||
|---|---|---|---|---|---|
| θt | |||||
| Strategy | θt | G | P | C | |
| 3 | G | H | .70 | .30 | .00 |
| L | .60 | .40 | .00 | ||
| Z | .00 | .00 | .00 | ||
| P | H | .30 | .70 | .00 | |
| L | .40 | .60 | .00 | ||
| Z | .00 | .00 | .00 | ||
| C | H | .10 | .90 | .00 | |
| L | .20 | .80 | .00 | ||
| Z | .00 | .00 | .00 | ||
4a
| Strategy | θt | θt–1 | |
|---|---|---|---|
| 4 | G | G | H |
| P | L | ||
| P | G | H | |
| P | L |
4b
| Pr(θtθt& <t) | |||||
|---|---|---|---|---|---|
| θt | |||||
| Strategy | θt | G | P | C | |
| 4 | G | H | .90 | .10 | .00 |
| L | .80 | .20 | .00 | ||
| Z | .00 | .00 | .00 | ||
| P | H | .10 | .90 | .00 | |
| L | .20 | .80 | .00 | ||
| Z | .00 | .00 | .00 | ||
| C | H | .05 | .95 | .00 | |
| L | .10 | .90 | .00 | ||
| Z | .00 | .00 | .00 | ||
5a
| Strategy | θt | θt–1 | θtθt&θt–1 |
|---|---|---|---|
| 5 | G | G | H |
| P | L | ||
| C | Z | ||
| P | G | H | |
| P | L | ||
| C | Z | ||
| C | G | H | |
| P | L | ||
| C | Z |
5b
| Pr(θtθt& At | |||||
|---|---|---|---|---|---|
| Strategy | θt | G | P | C | |
| 5 | G | H | .70 | .28 | .02 |
| L | .60 | .37 | .03 | ||
| Z | .55 | .40 | .05 | ||
| P | H | .30 | .65 | .05 | |
| L | .40 | .53 | .07 | ||
| Z | .45 | .45 | .10 | ||
| C | H | .10 | .20 | .70 | |
| L | .20 | .25 | .55 | ||
| Z | .25 | .25 | .50 | ||
6a
| Strategy | θt | θt-1 | Atθt&θt-1 |
|---|---|---|---|
| 6 | G | G | H |
| P | L | ||
| C | Z | ||
| P | G | H | |
| P | L | ||
| C | Z | ||
| C | G | H | |
| P | L | ||
| C | Z |
6b
| Pr(θtθt& At) | |||||
|---|---|---|---|---|---|
| θt | |||||
| Strategy | θt | G | P | C | |
| 6 | G | H | .90 | .09 | .01 |
| L | .80 | .18 | .02 | ||
| Z | .60 | .36 | .04 | ||
| P | H | .10 | .88 | .02 | |
| L | .20 | .75 | .05 | ||
| Z | .40 | .53 | .07 | ||
| C | H | .05 | .10 | .85 | |
| L | .10 | .20 | .70 | ||
| Z | .20 | .20 | .60 | ||
7a
| Strategy | θt | θt-1 | Atθt&θt-1 | |
|---|---|---|---|---|
| 7 | G | G | H | |
| P | H | |||
| P | G | L | ||
| P | L |
7b
| Pr(θtθt& At) | |||||
|---|---|---|---|---|---|
| θt | |||||
| Strategy | θt | G | P | C | |
| 7 | G | H | .70 | .30 | .00 |
| L | .70 | .30 | .00 | ||
| Z | .00 | .00 | .00 | ||
| P | H | .30 | .70 | .00 | |
| L | .30 | .70 | .00 | ||
| Z | .00 | .00 | .00 | ||
| C | H | .10 | .90 | .00 | |
| L | .10 | .90 | .00 | ||
| Z | .00 | .00 | .00 | ||
![]()
![]()
![]()