![]()
![]()
![]()
1. COMPARATIVE REGIONAL STOCK ASSESSMENT ANALYSIS OF THE SHRIMP RESOURCES FROM NORTHERN BRAZIL TO VENEZUELA by N.M. Ehrhardt
2. REGIONAL ASSESSMENT OF RED SNAPPER, LUTJANUS PURPUREUS by A. Charuau, K. Cochrane, D. Die, A. Lárez, L.A. Marcano, T. Phillips, S. Soomai, R. Souza, S. Wiggins and M. IJspol
3. REGIONAL ASSESSMENT OF THE BRAZIL-GUIANAS GROUNDFISH FISHERIES by A. Booth, A. Charuau, K. Cochrane, D. Die, A. Hackett, A. Lárez, D. Maison, L.A. Marcano, T. Phillips, S. Soomai, R. Souza, S. Wiggins and M. IJspol
4. REVIEW OF ENVIRONMENTAL CONSIDERATIONS IN MANAGEMENT OF THE BRAZIL-GUIANAS SHRIMP AND GROUNDFISH FISHERIES by P. Charlier
1.1. Comparative analysis of southern brown shrimp, P. subtilis
1.2. Comparative analysis of pink spotted shrimp, P. brasiliensis, abundance trends
1.3. Fishing mortality rate analyses
1.4. Conclusions
The shrimp resources in the northern Brazil-Venezuela region support one of the most important shrimp fisheries in the world. These resources include four of the larger penaeids (southern brown shrimp Penaeus subtilis1, pink spotted shrimp P. brasiliensis, southern pink shrimp P. notialis and southern white shrimp P. schmitti) and the smaller seabob shrimp (Xiphopenaeus kroyeri). The general distribution and abundance of the species differ markedly amongst countries in the region. The brown shrimp commercial category consists of the brown shrimp P. subtilis, the pink shrimp P. notialis and the pink spotted shrimp P. brasiliensis. The species composition in this “brown shrimp mix” varies among countries and fleets as a consequence of regional differences in abundance and of fleets targeting different species according to market preferences. There are differences among commercial category landings as well, due to differences in growth and spatial size availability of the species to the national fleets. Generally, the brown shrimp, P. subtilis, is more abundant in the eastern (Brazil through Suriname) than in the western (Guyana through Venezuela) regions of the shelf, while the pink spotted shrimp, P. brasiliensis, is far more important in Guyana and Suriname than in the remaining countries. The species is not caught in the Brazilian fishery and usually very large individuals are caught off the Venezuelan coast, but the species is secondary to P. subtilis in the inshore areas of the Gulf of Paria. Information on species composition in the landings in the brown shrimp category in the different countries has been conspicuously lacking. Consequently, most of previous stock assessments were performed on data where the species were combined and, fundamentally, based on global equilibrium production models using compounded catch and effort information.
A significant fishery for sea-bob (X. kroyeri) exists for some years in Suriname but there is an absence of information concerning the abundance, spatial distribution and population dynamics aspects of this species.
White shrimp (P. schmitti) assessments have only been carried out for the artisanal fishery that operates in the Orinoco-influenced region of the Gulf of Paria, but no other assessments are available regionally for the species. During the period 1996-2000, five shrimp workshops were organised by FAO and CFRAMP to assess the abundance and status of exploitation of the shrimp stocks in the Brazil-Venezuela region. These assessments were done separately by species and sexes due to the usual sexual dimorphism found in the species. Calibrated length- and/or age-based sequential population analysis techniques were used to assess abundance and fishing mortality of the stocks. A significant effort was invested in integrating databases required by the methods used. In the analyses presented in this report, only the complete regional assessments for P. subtilis in Guyana, Suriname, French Guiana and Brazil, and for P. brasiliensis in Guyana and Suriname are compared.
Although assessments for P. notialis were completed in Guyana, similar assessments are still not available for the other countries. No assessments are available for the sea-bob fishery and an assessment for the white shrimp, P. schmitti, was completed for the Gulf of Paria only. Hence, the following comparative regional analyses are for P. subtilis and P. brasiliensis only.
1.1.1. Abundance trends
1.1.2. Environmental factors and abundance
The absolute abundance in number of shrimp by sexes was estimated by calibrated length cohort analysis in the cases of Brazil, Guyana and Suriname, while an age-based calibrated sequential population analysis was applied in the case of French Guiana. Figure 1.1 shows the sexes combined relative abundance of the brown shrimp in the different countries. The importance of the species in the Brazilian fishery is evident in the figure as well as the seasonal characteristic of the species abundance. The abundance of P. subtilis in French Guiana is significant relative to the abundance estimated in Guyana and Suriname. It shows a smoother seasonal abundance pattern as a consequence of the age slicing procedure required to generate the catch-at-age matrix needed for the age-based stock assessments used in French Guiana. It is conspicuous in the Brazil stock abundance trend (Figure 1.1) that the species undergoes significant periods of low and high abundance. For example, lower abundance dominated the period 1982-1986 (12-60 in graph), while higher abundance levels are observed during 1987-1994 (72-156), followed by lower abundance in 1995-1997 (168-192) and significantly lower in 1998 (204) and 1999 (216). This pattern of abundance when looked at over a two-decade scale shows a significant increasing trend in abundance from 1982 (12) through 1993 (144) and then a decreasing trend observed from 1993 to 1999 (216).
Figure 1.1 Total monthly abundance of P. subtilis from Brazil to Guyana
In Figure 1.2, the relative abundance of P. subtilis in Brazil and French Guiana is portrayed in standardised units (observation minus the mean and then divided by the standard deviation) or anomalies. Specific details in the French Guiana abundance trends that were not clearly observable because of scale factors in the relative drawings in Figure 1.1 show that significant changes in abundance were conspicuous in the recent history of the stock in that country. In effect, three peaks in abundance are observed in the 1991, 1995, and 1998 seasons. A conspicuous decreasing trend in abundance is also depicted during the period 1995-1999, which is coincident with a similar trend observed in the Brazilian stock.
Figure 1.2 Average monthly abundance anomalies of P. subtilis in Brazil and French Guiana
Brown shrimp stock abundance in French Guiana is compared with the abundance of the species in Suriname in Figure 1 3. The seasonal abundance trends are slightly off phase - an indication of possible recruitment time differences - however, the relative (anomaly) levels of abundance appear very approximate indicating the possibility of similarities in the relative population processes occurring in that region and stocks. It is observed that the Suriname stock follows a very similar and more conspicuously decreasing trend in abundance than the one observed in French Guiana during the period 1995-1999.
Figure 1.3 Average monthly abundance anomalies of P. subtilis in French Guiana and Suriname
The relative seasonal abundance and intensity of brown shrimp in Suriname and Guyana appear coincident in some years (Figure 1.4) but more generally, they appear to have some differences in recruitment times as well as intensity. A low level of abundance is also present in the last year (1997) for which assessments were performed in Guyana, which may also be indicative of a potential decreasing trend in abundance of P. subtilis in Guyana. General opinions from the fishery support this hypothesis that the abundance available to the fleets from Guyana has been lower in recent years.
Figure 1.4 Average monthly abundance anomalies of P. subtilis in Guyana and Suriname
Figure 1.5 Average monthly abundance anomalies of P. subtilis in Guyana and Brazil
In Figure 1.5, an attempt is made to compare the relative abundance of the most geographically separated of the P. subtilis stocks analysed (Brazil and Guyana). Two conditions characterise the trends: 1) a conspicuous difference in phase in the seasonal abundance of the species in the two countries is apparent and 2) a relative abundance pattern that is similar in the general anomaly trend. These two conditions are striking because they should be indicative of fundamental differences in the seasonality of the recruitment schedule, but still follow the same population processes, which may be dynamically similar but with different relative intensities.
In general, it is possible to conclude that there is a common decreasing trend in the abundance of P. subtilis in the last five to six years of the analysis. That decreasing trend appears to be part of a longer-range trend that peaked in the mid-1990s and that has reached low levels similar to those observed in the early to mid-1980s. The factors affecting such longer-term variability have not been analysed, but they might be related to environmental changes that may be impacting recruitment in various ways. For this reason, an effort was made here to integrate regional environmental data that could serve the purpose of explaining the abundance dynamics observed in the previous figures.
One general characteristic of the Brazil-Venezuela shrimp habitat is the presence of a large influx of fresh water from extraordinarily large river basins. The influence of the Amazon on the easternmost boundary of the resources distribution to the Orinoco in the westernmost boundary is further impacted by such large river systems as the Essiquibo and Demerara in Guyana, the Corantyne (border), Nickerie, Coppename and Suriname rivers in Suriname and the Marowijne/Maroni (border), Mahury, Approuague and Oyapock (border) rivers in French Guiana.
In Figure 1.6, the outflows of the Essequibo and Demerara rivers in Guyana are shown for the period 1986-1995. Some similarities in the seasonal outflow are evident between the two river systems and in some years the maximum outflow may differ between the two rivers. In the short database presented in Figure 1.6, it is observed that some conspicuous changes in the freshwater outflow have occurred; hence the shrimp habitat has been exposed to these changes.
Figure 1.6 Flows of the rivers Essequibo and Demerara in Guyana
Figure 1.7 shows the anomalies of rainfall, through the Essequibo runoff, and P. subtilis recruitment, which is lagging by 3 months. There is a general seasonal effect of the environment on the recruitment pattern, but the intensity of the recruitment is not clearly related to the intensity of the environmental variables plotted. One plausible cause for the latter condition may be found in a parent stock abundance effect on recruitment, hence a generational effect on the recruitment trend observed in Figure 1.7.
Figure 1.8 Rainfall from 1986 to end 1996 in Belém, Brazil and Georgetown, Guyana
In Figure 1.8, the average monthly rainfall trends in Georgetown, Guyana and in Belém, Brazil are shown. In general, there is a significant discrepancy in the seasonal rainfall pattern in the two regions. Main peaks during the rainy seasons do not overlap, but they appear to be out of phase by 3 to 5 months on average over the time period examined.
Because of the potential effect of differences in the rainfall regimes between the eastern and western boundaries of the P. subtilis distributional range, the monthly averages of the rainfall over the period 1986 through 1997 were estimated and the resulting values presented in Figure 1.9.
Two major findings are portrayed in the figure: a) the hydrology regimes in Guyana and Brazil are very different regarding the timing and b) the hydrology regimes are very different in character.
In Guyana rainfall intensity shows two seasonal peaks: the first in December to January and then a more conspicuous peak from May to July, with dry seasons in February-March and September-October.
In Brazil, the hydrological cycle appears to have a single, but more protracted rainy season between January and April, followed by a similarly protracted dry season between June and November.
The effect of these cycles on the biology of P. subtilis is observed in Figure 1.10, where the seasonal biological condition factors (the seasonal change of weight per unit of length) of females and males of the species follow a negatively correlated trend with rainfall, excepting the period November-January. Similar conditions factors for the Brazil fishery were not available at the time this report was prepared and it will be important to construct similar logic to explain the biological impact of the hydrological cycles impacting shrimp in that fishery.
The previous findings prompted some further analysis of the average seasonal P. subtilis stock abundance in Brazil and Guyana. In effect, the anomalies of the 20-year (1981-2000) monthly average of the species abundance in the two countries shows a dramatic seasonal negative correlation (Figure 1.11) that explains the previously detected general difference in phase in the historic abundance patterns presented in Figure 1.5.
Figure 1.11 Seasonal abundance anomalies over 20 years of P. subtilis in Brazil and Guyana
In Figures 1.12 and 1.13, the anomalies of the abundance and rainfall with no time lags are presented for Guyana and Brazil, respectively. The relative trends are indicative of the existence of fundamentally different population dynamic mechanisms acting on the Guyana and Brazil brown shrimp stocks. Therefore, the dynamics of the brown shrimp stocks in the two geographically most separated fisheries for the species cannot be simply integrated. Therefore, common conclusions cannot be drawn for management purposes.
The findings are also indicative of the need to include environmental analyses in stock assessment work of this species. Environmental information from French Guiana and Suriname was not available at the time of this report. Hence, similar analyses should be attempted in the future to elucidate any potential population dynamic difference that may exist on the P. subtilis stocks in those two countries.
The analyses presented in this section correspond to abundance trends for the species P. brasiliensis in the Guyana and Suriname fisheries. Estimates of abundance for P. brasiliensis in the other countries of the region have not been attempted as yet.
The absolute abundance expressed in number of individuals of P. brasiliensis in the Guyana and Suriname stocks is presented in Figure 1.14. The analyses of the monthly abundance trends in this figure indicate that the seasonal trends are much more conspicuous in Suriname than in Guyana. In effect, there is very little concordance between the seasonal patterns observed in the two stocks given that in Suriname the peaks of seasonal abundance are much greater than those observed in Guyana. It is also apparent that the abundance of the stocks is different, being significantly higher in Suriname than in Guyana. A striking similarity, however, is the generally decreasing trend in the abundance of the two stocks, indicating that the stocks in these two countries may be subjected to similar population dynamics processes that are impacting recruitment.
Figure 1.14 Average abundance of P. brasiliensis in Suriname and Guyana
Figure 1.15 Anomalies in average abundance of P. brasiliensis in Guyana and rainfall in Georgetown
Figure 1.16 Cumulative rainfall in Georgetown, Guyana
The potential environmental effect on P. brasiliensis abundance is shown in Figure 1.15. In general, there appears to be a concordance of the seasonal rainfall cycles and the seasonal abundance of the stock in Guyana. The correspondence of the seasonal cycles is more conspicuous in the latter part than in the earlier part of the series. In general, the monthly rainfall pattern in Georgetown, Guyana has not followed a trend, however, the cumulative annual rainfall shows some significant changes with a conspicuous decreasing trend from 1965 through 1995 (Figure 1.16). The consequences of such a trend cannot be assessed at this time, but it may have a considerable effect on the abundance of P. brasiliensis in Suriname and Guyana.
P. subtilis
Monthly fishing mortality rate trends for males and females P. subtilis in Guyana, Suriname and Brazil are presented in Figures 1.17 to 1.19. A common feature in all the figures is the increasing trend in fishing mortality with the only exception being males of P. subtilis in Brazil. Fishing mortality rates are slightly higher in Guyana than in Brazil, while the fishery in Suriname shows the lowest fishing mortality trend among the three fisheries. It is important to note that in the above trends, the increases in seasonal fishing mortality have been more conspicuous in the later years of the series (see Figures 1.17-1.19).
Figure 1.17 Monthly fishing mortalities of male and female P. subtilis in Guyana
However, most of the monthly fishing mortality rates are lower than the monthly natural mortality rate of 0.18-0.20, assumed for the species in all stock assessments. It is important to note, however, that the levels of fishing mortality observed in the figures are rapidly approaching those of the natural mortality rate, therefore, this may be an indication that all stocks of P. subtilis in the region may be undergoing exploitation rates at levels close to those indicating full utilization.
Figure 1.18 Monthly fishing mortalities of male and female P. subtilis in Suriname
Figure 1.19 Monthly fishing mortalities of male and female P. subtilis in Brazil
P. brasiliensis
Similarly, fishing mortality rates for P. brasiliensis are presented in Figures 1.20 and 1.21 for Guyana and Suriname, respectively. The general increasing trends in the fishing mortality rates, F, are significant and there is a marked difference in the exploitation of the species in the two countries in that F is much higher in Guyana than in Suriname. The fishing mortality rates in Guyana have reached the levels of natural mortality in several occasions indicating the possibility that the fishery in Guyana may have reached levels of full utilization.
It is important to note that the trends in fishing mortality observed in all the species and fisheries are not high enough as to have created the very conspicuous decline in abundance that is reported for all the cases above. The implication of these trends may then be that environmental factors may be a more significant factor than fisheries for determining recruitment in these species.
Figure 1.20 Monthly fishing mortalities of male and female P. brasiliensis in Guyana
Figure 1.21 Monthly fishing mortalities, P. brasiliensis in Suriname
The comparative regional analyses presented here provide for the first time a vision of the historic status of abundance and exploitation of the two most important shrimp species in the northern Brazil-Venezuela region. There are stock assessments still missing in the brown shrimp category, for example P. notialis has been assessed only in Guyana and assessments for P. subtilis in Trinidad and Tobago and Venezuela have not yet been completed. It will be important to complete those assessments in order to have a full set of comparative analyses for the fishery that operates on brown shrimp.
In general, all the species are subjected to increasing trends in fishing mortality. However, the observed decreasing trends in abundance are more conspicuous than those that the exploitation rates would have generated on the stocks if all other sources of mortality had kept constant. This analysis has shown that the shrimp species in the region are subjected to great changes in the environment as demonstrated by the seasonal river run-off and rainfall trends. Preliminary analyses (not reported yet) on the impact of environmental changes on recruitment of P. subtilis in the region show that once the recruitment variability that is explained by the abundance of parent stock is removed, there appears to be a good correlation with seasonally-delayed environmental effects acting on the recruits. The expansion of the environmental analysis on recruitment should be considered as an important scientific task for the immediate future as those analyses will help elucidate characteristics of the temporal and spatial shrimp biomass dynamics and availability to the fleets.
The findings in these analyses should also prompt the need to consider the effects of environmental changes on the benchmarks (reference points) that are usually adopted to manage fisheries of species with short life spans such as the tropical shrimps. Among these are maximum sustainable yield, F0.1 and % spawning potential ratios used to control fishing mortality or any other mechanism that may be used to optimize biomass utilization in annual species (e.g., minimum size and closed seasons).
2.1. Introduction
2.2. Fishery management
2.3. Recommendations
2.4. References
The red snapper fishery is one of the most important fisheries in the region between eastern Venezuela and northern Brazil. Venezuelan fishermen from Isla Margarita, who still represent the main fishing group active in this fishery, started it in 1940. For these fishermen, the fishery is an extension of their Caribbean fisheries for red snapper, but the fleet that operates east of the Orinoco has for many years consisted of vessels larger than those operating in the vicinity of Margarita Island and Trinidad. Most of the catch comes from handlines, although longlines are being used more and more in this fishery. In Trinidad in the 1980s, the fisheries expanded to the east coast, where a fleet using fish pots and lines operated, whereas in 1998 the area of major industrial activity was the west coast.
Since the 1980s, in order to gain access to fishing grounds within the EEZs of Guyana, Suriname and French Guiana, the Venezuelan fleet has been required to land part of its catch in the host country. Although the bulk of landings in French Guiana and Suriname is made by Venezuelan vessels, some small catches of red snapper are made in Suriname by other fleets, including the national shrimp trawl, fish trawl and artisanal gillnet fleets. In Guyana, a local fleet of between 4 and 15 vessel operated over many years. This fleet has now grown to 39 vessels, including 20 Venezuela-owned vessels that are leased by a Guyana fishing company.
West of the Amazon delta, in Brazil, the fishery developed from the migration of vessels from Ceará and Piaui, because of the depletion of stocks of red snapper in those states. In Brazil, the fishery was traditionally dominated by handlines, but lately longlines and pots, as well as some lobster gillnets make significant catches.
2.2.1. Biological Information
2.2.2. Fishing Operations
2.2.3. Stock Assessments
The different countries of the region have very different priorities with regard to their red snapper stocks. For Venezuela and Brazil, red snapper is a traditional resource that has been harvested by their fleets for a long time. However, the Brazilian catches come from stocks under their own jurisdiction, whereas for Venezuela a large part of the catch comes from waters of foreign countries. In Venezuela, the main management issue is to maintain access to foreign fishing grounds and there have been few attempts at monitoring the harvest or establishing a management system that will ensure sustainable use of this resource. In Brazil, the main management issue has been the sharing of the resource between different groups of fishermen. A secondary issue in Brazil has been the appropriateness of the management measures for red snapper (such as the minimum size) that were established for the stocks in the waters to the southeast of the Amazon River. For Suriname and French Guiana, red snapper is a resource harvested mainly by a foreign fleet and therefore the greatest management priority is to recover as much of the benefits from the harvest as possible by requiring landings in their own countries, by charging license fees or by trying to establish national fleets.
It is clear, however, that all countries of the region should have a common interest in ensuring that snapper stocks are harvested sustainably and in a way that obtains the maximum benefits from the resource. In order to be able to achieve a sustainable red snapper fishery, it is essential that snapper stocks are assessed throughout their range and that the operations of their fleets are described in detail for the whole region.
The red snapper (Lutjanus purpureus) is only one of many species harvested by these fishing fleets, but it is the main target species for most of them. For the Venezuelan demersal fleet, red snapper constituted 85% of the landings during 1997-1999, whereas for the Brazil fleet it constituted 86% of the handline catch (1974-1976) and 83% of the trap catch (1998-1999). In Guyana, approximately 93% of the catch is red snapper. Other important species are groupers (Epinephelus spp.) and vermilion snapper (Rhomboplites aurorubens). The biological information on these other species is limited and there have been no assessments of the status of their stocks.
Knowledge on growth and reproduction are available for red snappers from French Guiana (Charuau and Die 2000), Brazil (see e.g. Filho et al. 2000), for the southern Caribbean resource fished by Venezuela (Caraballo, 1989; Gonzalez and Eslave, 1999; Lárez, 1994; Lugo, 1986), the Atlantic resource fished by Venezuela (Gonzalez et al. 1998), the eastern part of Venezuela (Gonzalez et al. 1998) and for the north and northeast coasts of Trinidad and west coast of Tobago (Manickchard-Heileman and Phillips,1993).
In the period between 1997 and 1999, 240 Venezuelan vessels (greater than 14m length) were actively fishing red snapper outside the Venezuelan EEZ. Of these, 41 were licensed to operate in French Guiana and 20 in Guyana (Table 2.1). The number of Venezuelan vessels licensed to fish in Suriname from 1997 to 2000 has varied between a minimum of 82 (2000) and a maximum of 134 (1999).
Table 2.1 Number of vessels targeting red snapper (L. purpureus)
|
Country |
Total Number of Vessels |
Venezuelan Vessels |
|
Brazil |
86 |
0 |
|
French Guiana |
46* |
41 |
|
Suriname |
82-134# |
81-134 |
|
Guyana |
39 |
20(operated by a Guyana company) |
|
Trinidad |
(36)** |
+ 36 vessels < 14 m length reported to be fishing
since 1997 |
|
Venezuela |
240*** |
na |
* The 5 non-Venezuelan vessels were from BarbadosThe Venezuelan fleet uses handlines and longlines to harvest snappers. Typically handlines have from 5 to 7 hooks and each handline boat uses 10 handlines at any one time. Longline vessels carry 1 or 2 longlines, each with 400 hooks. A single handline has the same catching power as a longline with 150 hooks. Over the period 1997-1999, the demersal fleet used only handlines. These continue to fish throughout the region with the greatest portion of the catch estimated to come from the waters of Suriname and French Guiana and lesser quantities from Guyana and Venezuela (Table 2.2). Landings also take place throughout the region with the largest part of the catch landed in French Guiana, Suriname and Venezuela. Landings outside the region continue, for example, 280 tonnes of red snapper were sampled from the landings in Martinique by Venezuelan demersal vessels during 1997. Catches of other species, apart from some catches of vermilion snapper, are all landed in Venezuela.# In 2000, 1 of the 82 vessels was Trinidad owned
** Vessels fishing in the EEZs of other countries only. There is also an internal fleet, which also targets red snapper at various times.
*** This number includes vessels operating in waters of other countries.
In Guyana, Venezuelan and Guyanese vessels using handlines and traps land most red snappers, but small amounts are landed by shrimp trawlers and artisanal gill-netters. Some data on snapper landings has been obtained for the last three years. A limited amount of length frequency data has also been collected over that same period. From 1995 to 1999, most red snappers landed by handlines were between 30 and 45 cm standard length (SL). Most fish caught by traps were between 20 and 35 cm SL.
In Suriname, some Venezuelan vessels land part of their catch locally. In 1997, the total landed catch was 1500 t (Table 2.2). According to data from Venezuela, catch per unit effort in Suriname is similar to that of Guyana and lower than that from French Guiana.
In French Guiana, vessel licenses are transferable so that the number of individual vessels from Venezuela harvesting red snappers in that country during a year can be greater than 41. However, the average number of vessels operating at any point in time in the waters of French Guiana has always been smaller than 41, but it has been rising, with 20 vessels at the beginning of the 1990s and 25 vessels at the end of the 1990s. In the same period, the number of fishing days per year has risen from 4500 to about 8000 and there has been a corresponding increase in landings, from 1200 to 1800 tonnes. Catch per unit effort remained relatively constant at around 25 kg per hour fished until 1999 when it decreased to 20 kg per hour fished. The average size of the fish in the landings, however, has decreased dramatically in the last 10 years, from 45 cm in the early 1990s to 35 cm at the end of that decade.
Table 2.2 Landings of red snapper recorded by country in which landing occurred
|
Country |
Landings |
Year of landings |
|
Brazil |
3 000 |
1997 |
|
French Guiana |
1 653* |
1999 |
|
Suriname |
1 500 |
1997 |
|
Guyana |
700 (estimated) |
1999 |
|
Trinidad |
3 |
|
|
Venezuela |
570 (estimated) |
1999 |
* Nominal catch, taking into account the maximum permitted 25% returning to Margarita, Venezuela at the end of the trip.In Brazil, the fishery is conducted with a mixture of gears (handline; longline; trap and gillnet) and red snapper is also taken as a by-catch in the lobster fishery. Most of the catch comes from longline and trap. The Brazilian fleet uses from 20 to 45 hooks for each handline, compared to the 5 hooks used by the Venezuelan fleet. Junior and de Brito (1999) have estimated conversion factors to combine fishing effort of traps and handlines for the industrial and semi-industrial fleets. During 1997 this fleet landed around 3000 t of red snapper, of which 350 t were caught with traps and the rest with handlines. For 1997, catch per trap and day was 9 kg for the industrial fleet and 33 kg for the semi-industrial fleet. For the same period the catch per hook per day was 2.5 and 3.2 kg, respectively.3 Data were not available at the Workshop. Landings and trip information can be made available for the artisanal fleet, but there is no formal data collection system for the industrial fleets, with specific reference to those vessels operating pots on the east coast. The landings of the artisanal fleets are recorded in larger categories, such as “snapper”. This data needs to be disaggregated in order to pull out data on L. purpureus. Data are available from 1989 to present.
The results of analyses undertaken on L. purpureus relevant to management are summarised in Table 2.3. The only thorough stock assessment to date conducted on red snappers was performed by Charuau and Die (2000) with data exclusively from French Guiana. Length frequency samples and landings from Cayenne were used to develop a catch-at-size matrix for the period 1989 to 1999. These data were converted to catch-at-age by using the growth curve of Perodou (1994). A cohort analysis was then conducted to estimate fishing mortality and population abundance by age. According to this assessment, fishing mortality has significantly increased in this fishery, while recruitment has increased at the same time. Catch in weight per unit effort has not shown any clear trend over this same period. Analyses conducted by Charuau and Die (2000) showed a strong retrospective trend in fishing mortality and recruitment suggesting that estimates might be biased. The bias may be the result of changes in selectivity or targeting. The management implications of the assessments are that effort should be reduced by between 25 and 50% from current levels.
Table 2.3 Summary of assessment results for L. purpureus relevant to management by country. References for the information contained here are presented in the text where applicable
|
Country |
Estimated Status of Resource |
Assessment Method |
Comments |
|
Brazil |
Over-exploited and risk of recruitment over- fishing |
Catch-curve analysis and per recruit reference
points. |
72% of catch by handlines and longlines and 63% of catch by
traps is of immature fish. |
|
French Guiana |
Fully to over-exploited |
VPA and per recruit analyses |
Increasing numbers of young fish in catch |
|
Suriname |
No work done |
|
|
|
Guyana |
Preliminary data collation only |
|
|
|
Venezuela |
|
|
|
|
i) -Caribbean coast (no reference available) |
Over-exploited |
Catch-curve analysis and per recruit reference
points |
|
|
ii) -Atlantic coast |
No work done |
(Lárez, 1994). |
56% of catch from longline and handline fisheries is immature
fish |
|
Trinidad and Tobago |
Fully exploited |
Catch-curve analysis and per recruit reference
points |
|
A yield per recruit analysis undertaken by Manickchard-Heileman and Phillips (1993) suggested that red snapper off the north and north east coasts of Trinidad was fully exploited and that increased yields could be obtained by increasing the age at first capture to 3 years from the mean age at first capture at the time of the study of 1.4 years (281 mm total length). They estimated the length at first maturity to be 270 mm total length for males and 390 mm total length for females.
Preliminary results of assessments undertaken on the fishery in Brazil showed that over 60% of the catch of red snapper is of immature fish, which gives considerable cause for concern. The regulations there require that no more than 15% of the landings are less than the size at maturity, but this regulation is not enforced (Souza, 2000).
Despite the scarcity of reliable assessments, there is a substantial amount of data being collected on the red snapper fishery in the region (Table 2.4). This should be processed and used for appropriate assessments as soon as possible.
Table 2.4 Data currently available on red
snapper.
Another important source of information is the database
of the 1988 Nansen survey.
|
Country |
Landings |
Effort |
Biological and Other |
|
Brazil |
1974-1999 |
1974-1999 |
1974-1999 |
|
French Guiana |
1985-1999 |
1985-1999 |
Length (1986-1999), growth, maturity |
|
Suriname |
1998-1999 |
1998-1999 |
|
|
Guyana |
1995-present |
1995-present |
Length frequency of landings (1995-1999) some missing
months |
|
Venezuela |
|
|
|
|
i) Caribbean coast |
1981-1999 |
1981-1999 |
Length frequencies (1986) |
|
ii) Atlantic coast |
1989-1999 |
|
|
|
Trinidad |
No directed fishery Some data are available from artisanal and
semi-industrial fisheries, but not specifically for red snapper. |
No directed fishery Some data are available on artisanal and
semi-industrial fisheries, but not specifically for red snapper |
A formal data collection programme for snappers does not
exist. Length frequency data available for one year (1996/1997). |
2.3.1. Enforcement And Control
2.3.2. Management Measures
2.3.3. Monitoring
2.3.4. Research and Data Collection
In general, this is a very poorly regulated fishery with neither the coastal state nor the flag state, in the case of Venezuelan vessels fishing in the region, implementing adequate controls to ensure sustainable utilisation of the resource. In order to address this, both the coastal and port states need, in the first instance, to ensure enforcement of existing regulations aimed at conservation of the resource, such as minimum sizes and effort regulations. Inadequate enforcement capability in most if not all countries is a serious limitation.
Where assessments have been undertaken, there are strong signs of over-exploitation of the resource. The most sophisticated stock assessment analyses are available from French Guiana. It was estimated that effort in French Guiana needs to be reduced by 25-50%.
Of equal concern is that no full assessments have been undertaken in Brazil, Suriname, Guyana and Venezuela. High priority should be given to utilising whatever data are currently available in each country in order to assess the resource on a national basis, as well as to attempt a regional assessment. Until this has been done no increases in effort should be considered in any of these countries.
The regional nature of the Venezuelan dominated fishery, stretching from French Guiana to Venezuela, needs to be monitored in a controlled and reliable manner. This will require:
All of the above tasks would be facilitated and made more effective if a regional database was established and maintained.
Current research and monitoring efforts need to be strengthened across the region. This should include:
At this stage, little is known about the stock structure of resource within the region, and identification of this structure would contribute considerably to effective management. A tagging study could be an effective means of investigating this. In the interim, the precautionary approach would be to assume that each country has its own stock and to manage each of those in a sustainable manner.
Caraballo, I., 1989. Determinación de algunos parámetros poblaciones del pargo colorado, Lutjanus purpureus (Poey, 1875) (Pisces: Lutjanidae) de la región oriental de Venezuela. Tesis de Grado, Universidad de Oriente. Cumaná. 93p.
Charuau A. and D. Die, 2000. Red snapper (Lutjanus purpureus) fishery in French Guiana. FAO Fish. Rep. No. 628: 72-86.
Filho, M. Asano, R.F. Curtrim Souza and D. Dinnis Bezerra, 2000. Aspects of fishing for snapper (Lutjanus purpureus) on the north coast of Brazil. FAO Fish. Rep. No. 628: 45-53.
Gonzalez, L.W. et al., 1998. Edad, crecimiento y mortalidad de pargo en las Guayanas. Boletin de Investigaciones Marinas y Costeras 27.
Gonzalez, L.W. and N. Eslave, 1999. Edad y crecimiento del pargo colorado, Lutjanus purpureus Poey, 1875 (Teleostei: Lutjanidae) de la región oriental de Venezuela. Revista de Biologica Marina y Oceanografia 34 (1): 99-107.
Junior and de Brito, 1999. (Full reference unavailable)
Lárez, A., 1994. Dinamica de la exploitación del pargo colorado, Lutjanus purpureus (Poey, 1875) (Pisces: Lutjanidae) y del mero Epinephelus flavolimbatus (Poey, 1875) (Pisces: Serranidae) en el oriente de Venezuela y areas adyacentes. Tesis Maestria, Universidad de Oriente. Cumaná.
Lugo, T.M., 1986. Aspectos de la reproducción del pargo colorado Lutjanus purpureus (Poey, 1875) (Pisces: Lutjanidae) de la región oriental de Venezuela. Tesis de Grado, Universidad de Oriente. Cumaná. 96p.
Manickchard-Heileman, S. and D. Phillips, 1993. Preliminary stock assessment for the fishpot fishery of Tobago. Technical Report. FAO/UNDP, Port of Spain, Trinidad and Tobago. 37p.
Perodou, J.B., 1994. Dynamique de la population exploitée de vivaneau rouge (Lutjanus purpureus) de Guyane. Complementarité des analyses globale et structurale. Thèse de docteur de l’Université de Lille. 261p.
Souza, R. F. Curtrim, 2000. The fishery for red snapper, Lutjanus purpureus on the north coast of Brazil. Report prepared at the Fourth Workshop on the Assessment and Management of Shrimp and Groundfish Fisheries on the Brazil-Guianas shelf, Cumána, Venezuela, 2-13 October 2000. (Not published).
3.1. Introduction
3.2. Background to the Groundfish Fisheries in the Region
3.3. Fishery Management
3.4. Resource Status
3.5. The Road Ahead
3.6. Monitoring, Enforcement and Control
3.7. Future Research and Data Collection
3.8. Proposed New Management Measures
3.9. References
SYNOPSIS
Groundfish are important socially and economically as they sustain the domestic fish market by providing an accessible and affordable protein source. Certain species are also exported and therefore earn much foreign exchange. From a social perspective, groundfish provide the basis for an artisanal fishing culture. Management issues are complicated because of the variety of gears used, the multispecies nature of the groundfish fisheries and because they are harvested by many countries. To further complicate matters, there is a paucity of data pertaining to their biology, productivity and even catch and fishing effort. Confidence in stocks assessments is therefore low. Those results that are available illustrate a bleak to mediocre picture of the state of resources. Most resources are fully to over-exploited with little to no legislation in place to improve the situation. Proactive and rationale management based on sound scientific information is therefore imperative. To maintain sustainable harvesting, research and monitoring need to be strengthened across the region. This should include on-going collection of accurate and adequate catch, effort, size frequency and age data for each country. All landings should be recorded, wherever they occur, identifying the country of origin of each and hence elucidate of stock structure and movement patterns within and, most importantly, between countries.
The groundfish resources in the Brazil-Guianas shelf region (situated between the Amazon and Orinoco Rivers) are important for commercial and social reasons. Commercially there is strong domestic market demand for affordable and accessible fish protein together with a source of valuable foreign exchange when exported. Social reasons include the reliance of many rural fishers on an artisanal fishing livelihood.
In the past, there has been considerable confusion as to the value of the groundfish fishery as a whole. Shrimp has been perceived as the most important resource in the region because of the foreign exchange that it generates. In contrast, groundfish has been given a lower status because most landings are consumed domestically with its earnings unaccounted for. Groundfish have therefore been ignored or given only limited attention. Commensurate with shrimp’s perceived economic importance is the allocation of resources for research and management. The few groundfish species that are exported in any appreciable amount are red snapper (Lutjanus purpureus), lane snapper (L. synagris) and green weakfish (Cynoscion virescens). Red snapper is arguably the most important groundfish in the region as its distribution range is throughout the region and it is primarily exported. Red snapper was discussed in Chapter 2 and will not be reviewed here in detail. Export quality fish are beginning to be recognised as important, and management issues are now being addressed. There are, however, other species with minor export potential, but considerable social importance. Rural fishers are dependent on a sustainable harvest of resources to ensure social stability as domestic sales contribute to their economic security.
The groundfish resources are harvested various countries, using a variety of fishing gears, which each harvest a variety of species. These fisheries are therefore multigear, multispecies and multinational. Fishing methods can be classified as industrial or artisanal depending on the level of mechanisation. Industrial fleets are dominated by trawlers and large pot/line vessels and are often owned internationally.
These vessels can undertake long fishing trips as they have large cold storage facilities. It is these vessels that fish for the export market. Artisanal vessels are smaller, have reduced cold storage facilities and are restricted to length of time spent at sea and area fished. The gear used is varied and can in many instances be readily changed to meet specific fishing conditions. It is these vessels that fish for the domestic market. In certain circumstances, there has also been conflict between resource users such as industrial shrimp trawlers landing species targeted by artisanal line/pot fishers.
To address the management of groundfish in the region, this review outlines species given priority together with a synopsis of data that is pertinent for management. Specific management issues have been highlighted together with those measures that are currently in place. To conclude possible future data collection and assessment initiatives have been proposed.
3.2.1. Fishing Operations
The consultative process that included representatives from all participating countries identified the most important species (Anon. 2000). Those species with specific importance to at least one country were considered key species with certain species of common interest to the region. The species reviewed are the Lutjanidae: red snapper (L. purpureus) and lane snapper (L. synagris) and the Sciaenidae: king weakfish (Macrodon ancylodon), whitemouth croaker (Micropogonias furnieri), smalleye croaker (Nebris microps), Jamaica weakfish (Cynoscion jamaicensis) and green weakfish (Cynoscion virescens) (Table 3.1).
Table 3.1 Priority groundfish resources harvested in the Brazil-Guianas shelf region. The symbol § indicates country specific priority species.
Fish are harvested as either targeted catch (10) or as by-catch (20), using either hooks (H), trawls (T), pots/traps (P), fyke nets/Chinese seines (F) or gillnets (G).
|
CountrySpecies |
Brazil |
French Guiana |
Suriname |
Guyana |
Venezuela |
Trinidad andTobago |
|
L. purpureus |
§10HP |
10H* |
10H |
10H |
§10H |
10H |
|
L. synagris |
? |
10H* 20P |
10T |
20T |
§20H |
§10HP20TG |
|
M. ancylodon |
§? |
|
|
§20TFG |
§20T |
20T |
|
M. furnieri |
|
|
|
20TFG |
§20T |
§20THG |
|
N. microps |
|
|
§20T |
20TFG |
§? |
|
|
C. acoupa |
§? |
|
|
|
|
|
|
C. jamaicensis |
|
|
|
|
|
§20THG |
|
C. virescens |
|
|
§10T |
20TFG |
§20T |
|
*Catches from Venezuelan vessels
Brazil
Fishing areas in Brazil are restricted to their territorial waters using both artisanal and industrial vessels. Longlines (pargueira) and/or traps (manzuá) are the dominant gears deployed (Table 3.1) landing a variety of fish (Filho et al. 2000). The most important groundfish landed is red snapper, which is targeted by 86 industrial vessels (Table 3.2). No data are available on numbers of small or medium-sized artisanal vessel. Catch and effort data are restricted to L. purpureus (Tables 3.2 and 3.3). In the North Region, all snapper is landed in the State of Pará at 11 different landing points (Filho et al. 2000).
French Guiana
Fishing in French Guiana occurs between the Maroni River and Oyapock Rivers by local artisanal and industrial Venezuelan vessels (Charuau and Die, 2000). Snappers are the most important fish group landed and are fished by 46 Venezuelan vessels licensed by the European Union and fish using hooks and pots (Tables 3.1 & 3.2).
Another snapper directed fishery was initiated in 1996 using with pots on muddy grounds. Species targeted include vermilion snapper (Rhomboplites aurorubens) and L. synagris. Increasing amounts of R. aurorubens (180 tonnes in 2000) are being landed and this is possibly a response to declining catches of L. purpureus (700 tonnes in 2000). Most L. synagris are donated to crew at the end of trips.
The main by-catch of the hook and line fishery comprises ten species of groupers (not landed in Cayenne), various carangids, pelagic sharks (processed on board) and, seasonally, king mackerel Scomberomorus spp.
Table 3.2 Estimates of effort used to harvest priority groundfish resources in the Brazil-Guianas shelf region
|
|
Industrial |
Artisanal |
|||||||
|
Number of vessels |
Percentage breakdown |
Number of vessels |
Percentage breakdown |
||||||
|
Trawl |
Pots, traps & hooks |
Trawl |
Fyke |
Gillnet |
Hook and line and longline |
Pots, traps |
|||
|
Brazil |
86* |
|
100 |
? |
|
|
|
|
|
|
French Guiana |
46* |
|
100 |
0 |
|
|
|
|
|
|
Suriname |
7 |
100 |
|
? |
|
|
|
|
|
|
Guyana |
54 |
100 |
|
1331 |
? |
27 |
60 |
cadell=8 |
39* |
|
Venezuela - Paria |
88 |
? |
? |
324 |
100 |
|
73 |
9 |
4 |
|
Trinidad & Tobago |
58 |
52 |
48 |
586 |
15 |
|
50 |
25 |
10 |
*Red snapper directed effortSuriname
Industrial and artisanal vessels operate out of Suriname. Fishing grounds are in territorial waters with industrial vessels using trawling methods and artisanal gears, including drifting gillnets and Chinese seines.
Three industrial fleets fish(ed) Suriname waters; two Korean groundfish-directed midwater trawlers that operated between 1998 and 1999, a Dutch groundfish-directed bottom trawl fleet, consisting of five vessels and a domestic Suriname fleet. The Dutch fleet accounted for approximately 75% of the landings in 1998 despite fishing with smaller vessels and fewer hauls per day than the Korean vessels (Charlier et al. 2000).
Cynoscion virescens is the most important commercial species and is landed by a number of gears, the most important being trawls, drifting gillnets and Chinese seines. Lane snapper, L. synagris, has only recently gained importance in the landings in Suriname forming part of the by-catch of the shrimp trawlers and Korean and Dutch vessels (Charlier et al., 2000).
Table 3.3 Landings (tonnes) of priority groundfish resources harvested in the Brazil-Guianas shelf region using various gears. Fish are harvested using hooks (H), trawling (T), pots (P), fyke nets/Chinese seines (F) or gillnets (G)
|
Country |
Gear |
Year |
Species |
|||||
|
L. synagris |
M. ancylodon |
M. furnieri |
N. microps |
C. virescens |
C. jamaicensis |
|||
|
Brazil |
ALL |
|
No data |
- |
- |
- |
- |
- |
|
French Guiana |
H |
1997 |
10** |
- |
- |
- |
- |
- |
|
H |
1998 |
2** |
|
|
|
|
|
|
|
Suriname |
TGF |
1991 |
|
- |
- |
341 |
66.7 |
49.2 |
|
T |
1998 |
No data |
- |
- |
No data |
3151 |
- |
|
|
Guyana
|
T |
1998 |
No data |
7669 |
|
No data |
- |
- |
|
F |
1998 |
No data |
4381 |
|
No data |
- |
- |
|
|
G |
1998 |
No data |
6516 |
|
No data |
- |
- |
|
|
H |
1998 |
No data |
- |
|
No data |
- |
- |
|
|
Venezuela |
T |
1998 |
|
|
786 |
- |
- |
- |
|
T |
1999 |
50-150 |
6021 |
|
|
|
|
|
|
Trinidad & Tobago |
ALL |
1997 |
456 |
- |
608 |
- |
- |
71 |
*Average of 1987-1996Guyana
**Catches estimated from scientific sampling.
Guyana’s industrial fleet directs its fishing effort at shrimp. Groundfish are landed either as shrimp by-catch or is targeted by artisanal fishers. Fishing occurs throughout its territorial waters with landings centralised in the case of industrial vessels whilst being diffuse in the artisanal fishery.
The offshore industrial fishery consists of 125 shrimp trawlers, of which 48% are foreign owned. Foreign trawlers mainly exploit penaeid shrimp with finfish and small amounts of squid and lobster as by-catch. The locally owned trawlers mainly exploit seabob and various groundfish species such as M. ancylodon, M. furnieri, N. microps, catfishes (Arius spp.) and various Cynoscion species. Small quantities of penaeid shrimp are landed as by-catch (Hackett et al., 2000).
The artisanal fleet is diverse and is comprised of vessels using Chinese seines and nylon gillnets. The artisanal fleet is large (1331 vessels) with the Chinese seine fishery accounting for 27% (359 vessels) and nylon gillnetters for 18% (244 vessels) (Artisanal Frame Survey, 1994). The Chinese seine catch consists primarily of shrimp, M. ancylodon and N. microps. In 1997, preliminary estimates of Chinese seine catch indicated 36.8% M. ancylodon and 12.5% N. microps. A large amount of juvenile fish is caught and either discarded dead or used to produce “fish meal” (Chakalall and Dragovich, 1980). The nylon gill net vessels land M. ancylodon, N. microps, C. virescens, and acoupa weakfish (C. acoupa).
Trinidad and Tobago
Groundfish is an important resource in Trinidad and Tobago and is landed by artisanal and industrial vessels. The industrial fleet consists of shrimp directed trawlers that land groundfish as a by-catch and groundfish directed hook/pot vessels. Smaller artisanal vessels deploy various hook-based gears, pots and gillnets (Soomai et al. 2000).
The snapper fishery is one of the country’s most commercially valuable fisheries. Landings of snappers for 1997 were estimated at 318.4 tonnes with a value of TT$ 5.8. In 1996, the estimated value of exports (mainly to North America) of snappers was TT$ 1.62 million. The species of main importance in terms of landings and value are L. synagris, L. purpureus and R. aurorubens. Other species of lesser importance are grey snapper (L. griseus), dog snapper (L. jocu) and silk snapper (L. vivanus) (Soomai, 2000).
The Trinidad snapper fishery is focused mainly in the Columbus Channel and off the southeast coast. Snappers are also landed as by-catch from the demersal shrimp trawl fishery in the Gulf of Paria and south coast of Trinidad and from the pelagic gillnets set in shallow water off the south coast. Tobago also has an artisanal fish pot fishery operating off its continental shelf and shelf break north west of the island (Manickchand-Heileman and Phillips 1993).
Groundfish are predominantly landed as by-catch from the shrimp trawl fleet and by the gillnet and line fisheries, which also target the king mackerels (Scomberomorus spp) and other coastal pelagic species. The main species of commercial importance in the groundfish fishery are the sciaenids M. furnieri and C. jamaicensis, which are mainly landed by vessels operating in the Gulf of Paria and the Columbus Channel. Apart from trawling, the main gears catching these species are bottom-set gillnets, handlines and bottom-set longlines.
The Fisheries Division Vessel Census conducted in January 1998, identified 743 vessels that operate gears that capture groundfish (Chan A Shing, 1999). Of this figure, 402 vessels operate on the west coast of Trinidad. In 1994, M. furnieri accounted for 65% (627 t) of the total landings and 63% (TT$2.6M) of the total value of the groundfish landed by the artisanal fishery, apart from trawling. C. jamaicensis accounted for 9% by weight (87 t) and 15% by value (TT$0.6M) (Soomai et al., 2000).
Venezuela
The Venezuelan fishing fleet has maintained a historic presence fishing in all countries in the region. Red snapper is the most important species landed with approximately 240 boats (all larger than 14 m long) landing about 4400 t per year (Charuau et al. 2001).
During 1998, the by-catch accounted for 93% of the total catch in the nets. Of this, 33% was sold in the local market and the other 60% was dumped at sea. The main fish species landed include M. furnieri, Cynoscion spp., M. ancylodon, L. synagris, coco sea catfish (Bagre bagre), Arius spp., madamango sea catfish (Cathorops spp.), Atlantic moonfish (Selene setapinis) and various shark species. These species represent 80% of the total by-catch landings (Alió et al., 2000).
3.3.1. Management Concerns
3.3.2. Current management measures
3.3.3. Data Available for Management
The countries in the region have different management priorities for their groundfish stock(s). Despite specific differences, there is common ground; that of ensuring that resources are harvested at sustainable levels and that maximum benefits are obtained socially and economically. Specific management priorities and concerns outlined in Chakalall et al. (2000) are summarised below.
Brazil
Brazil is concerned with the prevention of overfishing, increasing production, increasing employment associated with fishing, long-term conservation of the resource and improved utilisation of by-catch.
French Guiana
French Guiana has few concerns. A decrease in the average size of some fish species has been noted and this has raised sustainability issues.
Guyana
Guyana has prioritised the need to reduce harvesting of juvenile groundfish by the Chinese seines, a reduction conflict between fishers operating the different gear types (trawlers fishing nearer inshore resulting in destruction of the gillnets and the Chinese seine gears). Concern has also been raised regarding the declining catches of M. ancylodon in the various fisheries (especially from the trawl fleet) and an increase in juveniles of this species in the Chinese seine net fishery. The management objectives for the inshore artisanal fishery is to increase the landings to a sustainable level which would enable the fishery to contribute to improved nutrition for the population, export earnings, increase employment and aggregate output thereby stimulating growth of the national economy (Shepherd et al., 1999).
Suriname
In Suriname management goals for small soft-bottom demersal fish (e.g. Macrodon ancylodon, Cynoscion virescens, Nebris microps) is to ensure the constant supply to the domestic market and to maintain reasonable prices. The main goal for the small sandy-bottom demersal fish (L. synagris) is the generation of foreign currency, since the exploitation of this resource is primarily export-oriented.
Trinidad and Tobago
Trinidad and Tobago is concerned about overfishing, understanding the socio-economic importance of groundfish to artisanal fishers in rural coastal communities and multigear-multispecies interactions between groundfish and shrimp and trawl by-catch.
Venezuela
Venezuela’s management issues are centred around sustainability issues, the interaction between their domestic and Trinidad and Tobago fleets, regional sharing of fish resources and the reduction of conflict between the industrial and artisanal sectors.
Brazil
Article 187 of the Federal Constitution of Brazil provides for the definition of an agriculture policy and explicitly includes agro-industrial activities, agriculture and livestock, fisheries and forestry. Article 225 of the Constitution identifies a number of principles that concerns the environment. Included among these principles, are the protection of fauna and the ecological management of species and ecosystems. Both these principles implicitly include fisheries. The responsibility for the application of these principles lies with Government. Groundfish regulations are restricted to red snapper where traps are required to have 8 cm square mesh (Chakalall et al. 2000).
French Guiana
Prior to the demarcation off an EEZ in 1977 access to the fishery was unlimited. The common fisheries policy of the European Economic Community (EEC), now European Union (EU), which came into effect in January 1983, covers French Guiana as an overseas department of France. The policy calls for common rules for fishing in the maritime waters, and co-ordination of structural policies of Member States to promote harmonious and balanced development of the fishing industry (Council Regulation (EEC) No. 101/76). Management is now through licensing through the EU. The licenses are nominative and free and decided by the EU.
Groundfish legislation is restricted to the snapper group. Effort has been capped at 46 vessels (41 Venezuelan and 5 are unused from Barbados), skippers have to land and sell 75% of their catches to two processors in Cayenne and that only handlines and traps may be used (Chakalall et al. 2000).
Guyana
Fisheries in Guyana are regulated by the Fisheries Act of 1957, the Fisheries Regulations of 1959, the Fisheries (Pin Seine) Regulations of 1962, the Fisheries (Aquatic Wildlife Control) Regulations of 1966 and the Maritime Boundaries Act of 1977. A new draft act has been tabled for parliamentary discussion.
Groundfish legislation is restricted to the licensing of all vessels together with the use of 45 mm cod-end mesh, while Turtle Excluder Devices (TEDs) are required on seabob trawlers (Chakalall et al. 2000).
Suriname
Fisheries in Suriname are regulated by the Decree on Marine Fishery, Decree C-14, in force since 1st January 1981. This legislation has been revised and a new fisheries law was drafted in 1992, but has not yet been promulgated. The new draft law stipulates the elaboration of annual management plans for the fishery types, in which all regulatory measures will be established. This approach should allow fisheries managers to adapt to the changing conditions of exploitation.
Regulations currently in force pertain to registration, annual fees, reporting, closed areas and destination of catch. All vessels require licensing, the snapper fishery is restricted to 100 hook and line vessels whilst the industrial trawling fleets may only operate at depths greater than 25 fathoms (45 m).
Trinidad and Tobago
The existing legislation, the Fisheries Act of 1916, was found inadequate as a legal basis upon which a modern fisheries management system can be structured. A draft Fisheries Management Act and Policy Directions for Marine Fisheries in Trinidad and Tobago in the 1990s, providing the framework for the management of both local and foreign fishing activities in the waters under the jurisdiction of Trinidad and Tobago, were produced in June 1995. The draft Fisheries Management Act, which should facilitate fisheries management, data collection, licensing, registration and enforcement, was circulated in 1997 to fishers and stakeholders in the fisheries sector for comments. It has not yet been promulgated.
In 1988, there was a Cabinet decision to place a ceiling on the total numbers of artisanal, semi-industrial and industrial trawlers. This has not been enforced. As proposed in the Management Plan for the Trawl fishery, owners of trawlers should be required to hold entitlements to the fishery and that these entitlements should be transferable provided the replacement vessel does not have a greater horsepower or fishing power, and provided that replacement of the vessel is in keeping with the level of fishing effort approved in the Plan.
Current legislation governing domestic fishing in Trinidad and Tobago is the Fisheries Act (Chapter 67:51). Under this Act, no snapper under eight inches (20 cm) in length should be landed or sold. The Act applies to rivers, inland waters and the 12-mile territorial sea, however, it does not apply to the EEZ (Chakalall et al. 2000).
Venezuela
The Government has forbidden the construction of new conventional trawling units since 1989 to try to limit the size of the fleet. Until 1993, the Government had promoted the movement of industrial units to Guiria, in order to increase the fishing effort in the Atlantic zone, and decrease it in other more traditional fishing areas. Since then, the situation has been reversed and the number of trawl units in the Atlantic zone has progressively decreased.
Venezuela’s trawl fisheries are regulated by the joint resolutions of the Ministry of Agriculture (MAC/DGSPA/No. 46) and Ministry of the Environment (MARNR/DAA/No. 103) from 30th January 1980. The fishing areas for the trawling fleet and the ones reserved to the artisanal fishers are specified, both in the coastal zone and in the island territories. A second resolution (MAC/DGSPA/No. 391) from 13th December 1990 (Annex III) regulates the activity of the trawling fleet in the Gulf of Venezuela, establishing a closed season from 15th December to 8th January, and from 15th August to 10th September. Now all these resolutions are under study, in order to establish up-to-date norms for this fishery.
The industrial fleet has two closed seasons of one month - 15 January to 15 December and 15 May to 15 June. Vessels may only fish for 16 000 fishing days per year with a maximum of 235 days at sea per boat. All vessels must be licensed with trawl nets fitted with TEDs. In the artisanal trawl sector, vessels may only fish in depths greater than 10 m and use a cod-end mesh of 3.5 cm (Chakalall et al. 2000).
Lutjanus synagris
Growth, mortality, sex ratio and reproductive parameters have only been calculated from Trinidad and Tobago (Dass 1983, Manickchand-Dass 1987). Length-weight parameters and length frequencies are available for Suriname together with size composition and the landings in 1998 (Charlier et al., 2000).
Catch and effort data available for the assessment of L. synagris are limited. Pertinent data from Trinidad and Tobago include catch and standardised effort data for all major fisheries from both the west and south coasts between 1989 and 1997, together with length frequency data from fish harvested using pots and by banking from both coasts between 1996 and 1997 (Soomai 2000). In French Guiana, catch and effort data between 1986 and 1998 have been estimated from scientific sampling but have, unfortunately, not been disaggregated from the Venezuelan data. Catch and effort data between 1994 and 1999 are available from Venezuela (Tables 3.2, 3.3 and 3.4).
Macrodon ancylodon
Limited data are available on the reproductive biology, nursery areas and feeding biology of M. ancylodon (Bianchi 1992). Catch data is available from Guyana from the trawl, Chinese seine, gill net and hook fisheries for 1994 and between 1996 and 1998 and from Venezuela between 1994 and 1999 (Table 3.3 & 3.4) (Hackett et al., 2000).
Table 3.4 Data currently available for priority
groundfish species.
Another important source of information are the results
of the 1988 surveys with the R/V Dr. Fridtjof Nansen (see IMR,
1988)
|
Country |
Data available |
Species |
|
|
|
|
|
|
L. synagris |
M. ancylodon |
M. furnieri |
N. microps |
C. virescens |
C. jamaicensis |
||
|
Brazil
|
Landings (years) |
none |
- |
- |
- |
- |
- |
|
Effort (years) |
none |
- |
- |
- |
- |
- |
|
|
Biological and other |
none |
- |
- |
- |
- |
- |
|
|
French Guiana
|
Landings (years) |
1986-1998 |
- |
- |
- |
- |
- |
|
Effort (years) |
1986-1998 (not separated from Venezuela) |
- |
- |
- |
- |
- |
|
|
Biological and other |
none |
- |
- |
- |
- |
- |
|
|
Suriname
|
Landings (years) |
none |
- |
- |
1991-1992 |
1998-1999 |
- |
|
Effort (years) |
none |
- |
- |
1991-1992 |
1998-1999 |
- |
|
|
Biological and other |
Length frequencies |
- |
- |
Length frequencies |
Length frequencies, growth, mortality |
- |
|
|
Guyana
|
Landings (years) |
- |
19941996-1998 |
- |
none |
- |
- |
|
Effort (years) |
- |
19941996-1998 |
- |
none |
- |
- |
|
|
Biological and other |
- |
Length based population parameters, mortality |
- |
Length based population parameters, mortality |
- |
- |
|
|
Venezuela
|
Landings (years) |
1994-1999 |
1994-1999 |
- |
- |
1994-1999 |
- |
|
Effort (years) |
1994-1999 |
1994-1999 |
- |
- |
1994-1999 |
- |
|
|
Biological and other |
No data |
No data |
- |
- |
No data |
- |
|
|
Trinidad & Tobago
|
Landings (years) |
1989-1997 |
- |
1989-1997 |
- |
- |
1989-1997 |
|
Effort (years) |
1989-1997 |
- |
1989-1997 |
- |
- |
1989-1997 |
|
|
Biological and other |
Length frequencies, growth, reproduction, mortality |
- |
Length frequencies, growth, mortality |
- |
- |
Length frequencies growth, mortality |
Micropogonias furnieri
Otolith-based growth parameters have been calculated by Manickchand-Heileman and Kenny (1990) in Trinidad and Tobago. Catch, effort and length frequency data are available from Trinidad and Tobago between 1989 and 1997 (Soomai et al., 2000). Catch data from 1998 are available from Venezuela (Tables 3.2, 3.3 and 3.4) (Alió et al., 2000).
Nebris microps
Biological information is restricted to Guyana, which only has anecdotal information related to distribution and nursery areas. The growth parameters have been calculated using length-based methods, but are considered unrealistic (Hackett et al., 2000). Catch or effort data are restricted to 1991 and 1992 for Suriname data (Tables 3.3 and 3.4).
Cynoscion jamaicensis
Biological data such as length-weight, growth and mortality parameters have been estimated from length frequency data from Trinidad and Tobago. Catch and effort are restricted to Trinidad and Tobago for the period 1989 to 1997 (Table 3.3) (Soomai et al., 2000).
Cynoscion virescens
Biological parameters, such as length-weight, growth and mortality parameters, have been estimated from length frequency data in Suriname (Charlier et al. 2000). Catch (but not effort data) is available from Suriname between 1998 and 1999. Catch and effort data between 1994 and 1999 has also been collected by Venezuela (Table 3.3).
Lutjanus synagris
Per recruit analysis conducted on the Suriname stock of L. synagris revealed that the stock is under- to fully-exploited from both yield- and spawner biomass -per-recruit perspectives (Table 3.5; Charlier et al. 2000).
Per-recruit analysis by Maingot and Manickchand-Heileman (1987) showed that L. synagris in Trinidad and Tobago was under-exploited. Analysis of recent catch trends (Booth et al. 2000) showed that on the west coast catches have decreased slightly (1.16% year-1) as opposed to the rapid increases in south coast fishery over the same time period (35% year-1). Effort trends for both coasts were similar with a rapid decrease of more than 60% after 1993. The reasons for this decline are not known. Concomitant with the decrease in fishing effort has been an increase in catch-per-unit-effort (CPUE). Trends in CPUE are often used as a proxy for resource abundance and suggest that there is an apparent recovery of the stock at a rate of 23% year-1 on the west coast and a 34% year-1 on the south coast. The true magnitude of this recovery is however unknown and is probably less than that required for an economically viable fishery as the fishers that left the pot fishery have not returned (Table 3.5).
Of continued concern is that a large proportion of the banking catch is immature (Booth et al. 2000).
Macrodon ancylodon
Results from the deterministic and stochastic per-recruit analyses in Guyana revealed that the stock was overfished from both growth and recruitment perspectives (Table 3.5) (Booth et al. 2000).
Table 3.5 Results of stock assessments conducted for priority groundfish species in the Brazil-Guianas region. LCA = Length-based cohort analysis, PR = per-recruit analysis and CPUE = catch-per-unit effort trend analysis
|
Country |
Assessment methods Status of resource Comments |
Species |
|||||
|
L. synagris |
M. ancylodon |
M. furnieri |
N. microps |
C. virescens |
C. jamaicensis |
||
|
Brazil
|
Methods |
Not conducted |
- |
- |
- |
- |
- |
|
Status |
Unknown |
- |
- |
- |
- |
- |
|
|
Comments |
Unknown |
- |
- |
- |
- |
- |
|
|
French Guiana
|
Methods |
Not conducted |
- |
- |
- |
- |
- |
|
Status |
Unknown |
- |
- |
- |
- |
- |
|
|
Comments |
Unknown |
- |
- |
- |
- |
- |
|
|
Suriname
|
Methods |
PR, LCA |
- |
- |
- |
PR |
- |
|
Status |
Under- to fully exploited |
- |
- |
- |
Over exploited |
- |
|
|
Comments |
Selectivity needs to be estimated |
- |
- |
- |
Reduced F in 1999 |
- |
|
|
Guyana
|
Methods |
Not conducted |
PR |
Not conducted |
Not conducted |
- |
- |
|
Status |
Unknown |
Over exploited |
Unknown |
Unknown |
- |
- |
|
|
Comments |
Unknown |
Strong evidence for growth and recruitment overfishing |
Unknown |
Key biological parameters could not be calculated |
- |
- |
|
|
Trinidad and Tobago
|
Methods |
LF, CPUE trends |
- |
PR and CPUE, bio-economic analysis |
- |
- |
PR and CPUE, bio-economic analysis |
|
Status |
Possibly over exploited but recovering |
- |
Overfished |
- |
- |
Overfished |
|
|
Comments |
Increase in CPUE |
- |
- |
- |
- |
- |
|
|
Venezuela |
None |
|
|
|
|
|
|
Earlier yield per recruit analyses of the state of the fishery in the northern part of the region (Manickchand-Heileman and Kenny 1990; Alvarez et al. 1999, Soomai et al 2000) concluded that M. furnieri was fully exploited.
Recent trends in CPUE, per-recruit analysis, a biodynamic model and bio-economic analysis all indicate that M. furnieri is now over-exploited (Table 3.5). This overfishing situation can possibly be attributed to the combined effort of six gear types operating in the Gulf of Paria (Soomai et al. 2000).
A joint analysis of the stock using Trinidad and Venezuelan data from the artisanal and industrial fleets (Alió et al. 2000) showed similar results - that of low stock levels commensurate with high levels of exploitation. The biomass dynamic model suggested an MSY of approximately 1500 t (1300 to 1600 t). Current fishing levels are considerably higher than this.
Nebris microps
No stock assessment has been conducted on this species due to unfeasible parameter values calculated from length-frequency samples (Hackett et al. 2000).
Cynoscion virescens
Various analyses undertaken on the Suriname fishery all indicate extremely high fishing mortality for Cynoscion virescens. According to the calculations, current levels of fishing effort has reduced spawning biomass per recruit (SBR) to 11-21%% of pristine levels (Charlier et al., 2000).
Cynoscion jamaicensis
Trends in Trinidad CPUE and a per-recruit analysis showed that C. jamaicensis is overfished (Soomai et al. 2000).
The end of the CFRAMP/FAO workshops saw considerable progress particularly with regard to the assessment of shrimp. The groundfish assessments were however hampered. As it is been clearly shown, little data are available for assessment despite evidence for overfishing in most stocks. Data series are short in length and most effort data are not standardised. Population parameters such as growth and mortality rates are unknown or at best calculated by inaccurate length-based methods. If sustainable management of these resources is to be realised then there must be change.
Fisheries in the region are poorly regulated with neither the coastal state nor the flag state, in the case of Venezuelan vessels fishing in the region, implementing adequate controls to ensure sustainable utilisation of the resource. In order to address this, both the coastal and port states need, in the first instance, to ensure enforcement of existing regulations aimed at conservation of the resource, such as minimum fish sizes and fishing effort regulations. Inadequate enforcement capability in most, if not all, countries is a serious limitation.
Charuau et al. (this volume) made several recommendations with regard to monitoring and enforcement in the snapper directed fishery in the region. These are pertinent to groundfish as a whole. Countries should therefore endeavour to:
All of the above tasks would be facilitated and made more effective if a regional database was established and maintained.
Evaluation of any fish resource requires basic information such as catches, fishing effort and size structure of the animals captured. Other information associated with the biology of the species, like parameters to describe growth and mortality, are also crucial. Countries in the region should therefore:
Brazil
French Guiana
Guyana
Suriname
Trinidad and Tobago
Venezuela
Alió, J.J., L. Marcano, S. Soomai, T. Phillips, D. Altuve, R. Alvarez, D. Die and K. Cochrane, 2000. Analysis of industrial trawl and artisanal fisheries of whitemouth croaker, (Micropogonias furnieri) of Venezuela and Trinidad-Tobago in the Gulf Of Paria and Orinoco River Delta. FAO Fish. Rep. No. 628: 138-148.
Alvarez, R., D.E. Altuve, J.J. Alió, L.A. Marcano, K. Cochrane and M Tandstad, 1999. Assessment of roncador (Micropogonias furnieri) within the Orinoco and Gulf of Paria region. Rome, FAO. GCP/INT/575/DEN Report on Activity No. 46 (Supplement 1): 159-167.
Anon. 2000. Report on meeting of the task group on snapper fisheries of the Brazil-Guianas Shelf. FAO Fish. Rep. No. 628: 186-188.
Bianchi, G., 1992. Demersal assemblages of tropical continental shelves. Thesis: University of Bergen, Bergen, Norway. 217p.
Booth, A., K. Cochrane, A. Hackett and D. Shepherd, 2000. Application of a multispecies-multigear per-recruit model that incorporates parameter variability to the shrimp and groundfish fisheries of Guyana. FAO Fish. Rep. No. 628: 154-172.
Booth, A. and S. Soomai, 2000. Recommendations for the management of lane snapper in Trinidad. Report prepared at the Fourth Workshop on the Assessment and Management of Shrimp and Groundfish Fisheries on the Brazil-Guianas shelf, Cumána, Venezuela, 2-13 October 2000. (Not published).
Chakalall, B., K. Cochrane, T. Phillips and J. Prado, 2000. Existing approaches to fisheries management in the Brazil-Guianas shelf. FAO Fish. Rep. No. 628: 11-31.
Chakalall, B. and A. Dragovich, 1980. The artisanal fishery of Guyana. Southeast Fisheries Center, Miami, USA. (Ms.), 27p
Chan A Shing, C., 1999. Report on the 1998 census of fishing vessels (Trinidad). Fisheries Division, Ministry of Agriculture, Land and Marine Resources, Trinidad and Tobago, Fisheries Internal Report No. 3. 42p.
Charlier, P, M. IJspol, Y. Babb-Echteld and A. Booth, 2000. Shrimp and finfish fisheries in Suriname. FAO Fish. Rep. No. 628: 87-104.
Charuau A. and D. Die, 2000. Red snapper (Lutjanus purpureus) fishery in French Guiana. FAO Fish. Rep. No. 628: 72-86.
Charuau, A., K. Cochrane, D. Die, A. Lárez, L.A. Marcano, T. Phillips, S. Soomai, R. Souza, S. Wiggins and M. IJspol, 2001. Regional assessment of red snapper, Lutjanus purpureus. FAO Fish. Rep. No. 651: 15-21
Dass, 1983. Some aspects of the biology of the lane snapper Lutjanus synagris (Linnaeus, 1758) in Trinidad. Institute of Marine Affairs: Chaguaramas, Trinidad and Tobago. Research Report No. IMA/12/83.
Filho, M. Asano., R.F. Curtrim Souza and D. Dinnis Bezerra, 2000. Aspects of fishing for snapper (Lutjanus purpureus) on the north coast of Brazil. FAO Fish. Rep. No. 628: 45-53.
Fisheries Division, Ministry of Agriculture, Land and Marine Resources, Trinidad and Tobago, 1991. Trawler Gear Survey, 1991.
Hackett, A., K. Cochrane and A. Booth, 2000. Assessment of fisheries in Guyana for bangamary (Macrodon ancylodon) and butterfish (Nebris microps). FAO Fish. Rep. No. 628: 110-123.
Maingot, J and S. Manickchand-Heileman, 1987. Yield per recruit analysis of the lane snapper Lutjanus synagris (Linnaeus, 1758) in Trinidad, West Indies. Institute of Marine Affairs, Chaguaramas: Trinidad and Tobago.
Manickchand-Dass, S., 1987. Reproduction, age and growth of the lane snapper Lutjanus synagris (Linnaeus), in Trinidad, West Indies. Bull. Mar. Sci., 40 (1): 22-28.
Manickhand-Heileman, S. and J.S. Kenny, 1990. Reproduction, age, and growth of the whitemouth croaker, Micropogonias furnieri, (Desmarest 1823) in Trinidad waters. Fish. Bull. 88: 523-529.
Manickchard-Heileman, S. and D. Phillips, 1993. Preliminary stock assessment for the fishpot fishery of Tobago. Technical Report. FAO/UNDP, Port of Spain, Trinidad and Tobago. 37p.
Shepherd, D., A. Hackett and R. Charles, 1999. Shrimp and groundfish fisheries of Guyana. National report of Guyana. FAO Fish. Rep. No. 600: 17-38.
Soomai, S. 2000. The snapper fishery of Trinidad and Tobago. Report prepared at the Fourth Workshop on the Assessment and Management of Shrimp and Groundfish Fisheries on the Brazil-Guianas shelf, Cumána, Venezuela, 2-13 October 2000. (Not published).
Soomai, S., N.M. Ehrhardt, K. Cochrane and T. Phillips, 2000. Stock assessment of two sciaenid fisheries in the west coast of Trinidad and Tobago. FAO Fish. Rep. No. 628: 124-137.
4.1. Introduction
4.2. Environmental Features and Variability of the Brazil-Guianas Marine Ecosystem
4.3. Impact of Fisheries on the Ecosystem and its Productivity
4.4. Impact of Other Human Activities on Marine Ecosystems
4.5. Relative Importance of Fisheries Impact and Other Factors on this Ecosystem and its Productivity
4.6. Conclusions and Recommendations
4.7. References
Many fisheries operating on the Brazil-Guianas continental shelf are starting to face, as in many parts of the world, declining abundance of the resources they exploit and at the same time a growing demand of persons wishing to join the fishery, thereby further increasing the fishing pressure. The landings of some fishery resources, submitted to very moderate levels of exploitation for many years, have been increasing well into the 1990s, particularly some groundfish species for which there is little domestic demand in the sparsely populated Guyanas. Other fisheries, notably the export-oriented penaeid shrimp fishery, are considered to have reached their level of maximum production decades ago and exhibit more or less “stabilized” patterns of exploitation.
Nearly all stocks, however, are currently reaching their limits in productivity, and their fisheries require careful management to prevent serious decreases in the profitability or even in production. Countries in the region are recognising this and are beginning to give greater attention to the development of strategies and work plans for sustainable exploitation.
A wide range of resource assessment techniques have been used, particularly in the context of these regional working groups, in order to define optimal levels of fishing mortality that different stocks would be able to sustain in the long term. As a result, many legal and practical proposals have been made, with the aim of managing the fisheries at these levels. It has often been noted, however, that besides the fact that parameters needed for resource assessment can never be estimated with perfect accuracy, there are also external factors, which can be globally referred to as “environmental factors”. In other words, many elements of the marine ecosystems, other than the exploited species, are important for the behaviour and productivity of the different stocks. Methodologies for the analysis of fishery resources as part of their ecosystem have been developed only recently, and they have not been applied yet in many parts of the world.
Fishing activities, in addition to the changes that they obviously bring to the structure of the exploited populations as well as of a number of non-target species, may have substantial impacts on different aspects of the environment. Environmental changes caused by fishing can, in turn, have consequences for the exploited stock habitat, and the productivity of the ecosystems. Ecosystems, on the other hand, are subject to fluctuations and natural cycles and modifications that are, in many instances, still poorly understood.
Fishing is only one of many human activities with a potential impact on marine environment. Estuarine and coastal ecosystems are particularly exposed to consequences of development within and outside their limits, along the coastline, and in the catchment areas of their rivers. It is rarely straightforward to segregate effects of fishing, other man-induced effects, and changes brought about by natural processes.
4.2.1. Marine Currents
4.2.2. Freshwater Inputs
4.2.3. Turbidity, Sediment Load
4.2.4. Nutrient Contents and Primary Production
4.2.5. Formation of Mud Banks
4.2.6. Dynamic Coastal Environment
4.2.7. Nature of the Seafloor
4.2.8. Critical Habitats for Fisheries Production
4.2.9. Natural Cycles
The Brazil-Guianas region, as it has been referred to in this series of workshops, is the marine area comprised between the Amazon River and the Gulf of Paria. It represents a coast length of nearly 2 000 km (1 600 km between Orinoco and Amazon, plus the Gulf of Paria). The continental shelf covers a remarkable area, with a width varying between 80 and 200 km, and harbours most of the fishery resources. The continental break is situated at a depth of around 100 meters. On the shore side, a variety of brackish biotopes (marshes, mangroves and lagoons) cover most of the coast, forming a transition towards freshwater and non-aquatic ecosystems landwards and playing a key role in the functioning of the coastal marine ecosystems.
The hydrodynamics of this marine region is dominated by the Guyana Current, flowing N-W along the shores, in the prolongation of the North-Brazilian Current, a division of the South Equatorial Current, generated by the Trade Winds (Cochrane, 1963). The Guyana Current is about 250 km broad and reaches maximal velocities of 1.5 to 2 m.sec-1 (NEDECO, 1968). It is not, however, “a constant flow, running parallel to the Guyana’s shore” (Rossignol, 1977, cited by Prost, 1990). Variability is generated by tidal currents and by the presence of eddies, generally cyclonic above the continental shelf, along the coastal flank of the current, and anti-cyclonic beyond the continental shelf (Prost, 1990).
Annual variations in the hydrodynamics pattern are also generated by the seasonal oscillations of the Trade Winds between more northerly latitudes in the boreal winter and more southern latitudes in boreal summer. Under their influence, the Equatorial Counter-Current (Eastward) oscillates between the latitude of the Brazil-Guianas region and a more southern position. When the Equatorial Counter-Current is situated in front of the region, it deviates part of the Guyana Current flow towards an eastward direction and thereby decreases that current’s strength. This phenomenon, known as the “annual retroflexion of the North-Brazilian Current” (Müller-Karger et al., 1988), takes place from June to December. The Guyana Current reaches its maximal strength during the first half of the year.
Temporary upwelling events have been reported along the shelf (Gibbs, 1980). Their seasonality and relationship with biogenic productivity are still poorly understood (Martec Ltd, 1995).
The region receives the discharge of two of the world’s largest rivers, Amazon and Orinoco, and of at least 10 important, if comparatively much less powerful rivers (Oyapock, Maroni, Corantyne, Essequibo, etc.). Due to the marine hydrodynamics described above, the outflow of these rivers is deviated towards the N-W and influences the coastal environment in an area situated west of each estuary. The influence of the Amazon, however, with its average 180,000 m3.sec-1 discharge (Nittrouer and De Master, 1987), is predominant throughout the region. It has been estimated that 40 to 50% of the annual Amazon run-off is transiting along the coast of the Guyana’s. Amazon waters (characterised by their geochemical parameters, and low salinity) can still be detected off Barbados (Borstad, 1982). As a result, the ecosystem in most of the coastal area of the Brazil-Guianas region presents fairly continuous features, and has been described by marine geologists as a kind of “attenuated delta of the Amazon” (Rine and Ginsburg, 1985).
The influence of the Amazon on the shelf fluctuates through the year as its run-off varies by a factor 2.5 between its maximum in May-June and its minimum in November-December (Frouin, 1997) and also because much of the Amazon water is transported offshore by the retroflexion between June and December.
A major characteristic of waters from the Amazon is, besides low salinity, a very high turbidity. The Amazon discharges per year an estimated 731 million tonnes of sediment eroded, for the largest part, from the Andes, into the sea (Gibbs, 1976). Some 499 million tonnes of this material is suspended, and 232 million tonnes is dissolved. In contact with seawater, flocculation of suspended sediments and their deposition on the sea floor take place gradually. Therefore, as river water gradually mixes with oceanic water and salinity is increasing, its sediment load decreases, from a maximum of 50-200 mg.l-1 close to the mouth, to 9 mg.l-1 in front of French Guiana, 2 mg l-1 in front of the Orinoco, 0.6-0.7 mg l-1 further north (compared with 0.2-0.3 mg l-1 for ocean water) (Frouin, 1997). The “plume” formed by turbid Amazon waters is visible until 400 to 500 km outside the estuary. This plume is 100 to 200 km broad in front of the coast of Amapá (Brazil). Beyond Cape Orange, at the boundary between Amapá and French Guiana, where the direction of the coastline turns from northly to northwesterly, the flow of turbid waters is pressed, under action of currents and winds, in a narrow band along the shores (20 to 40 km, Froidefond et al., 1988). It has been estimated that about 150.106 m3 per year of fine sediment is transported in suspension along the coasts of the Guyanas (NEDECO, 1968). The sediment load of the coastal waters varies seasonally, in accordance with Amazon influence outlined above.
Compared with oceanic waters of the North Brazil Current, the Amazon outflow has an elevated nutrient content. Primary production is limited by low penetration of light necessary for photosynthesis in turbid waters influenced by the Amazon, while it is nutrient-limited offshore. Recent measurements have shown that there is little seasonal variation of the nutrient content (silicates, 144 µmol.kg-1, phosphates, 0.7 µmol.kg-1, nitrates, 16 µmol.kg-1, etc.) in the plume of the Amazon (De Master and Pope, 1996). Primary productivity is highest in the mixing zone of the two types of water, reaching 8 gC.m-2.d-1, compared with averages of 2.18 and 0.81 gC.m-2.d-1, respectively, for turbid river water and for oceanic, clear water (Smith and De Master, 1996).
While sedimentation occurs homogeneously in front of Amapá, this process becomes discontinuous from Cape Orange northwards. In French Guiana, Suriname, Guyana and Venezuela south of the Orinoco River, the sedimentation process concentrates in regularly spaced zones, where mud banks appear. This mechanism was described by several authors, notably Augustinus (1983, 1989) for the Suriname coast, Eisma et al. (1971, 1978) for the coast of Guyana and Venezuela and Froidefond et al. (1988) for French Guiana. Mud banks extend from the shore to some 20-40 km into the sea (20 m depth), and they affect 20 to 30 km of shore. Distance between banks is of the same order. Water movement is much more important on the eastward side of the banks, exposed to dominant currents and waves, than on the westward side, sheltered by the bank itself. Part of the silt particles therefore becomes re-suspended at the eastern border, while deposition is intensive on the other border. The result is a shifting of the bank along the coast. In this fashion, mud banks migrate all the way from Cape Orange to the Orinoco Delta, with an average velocity of 1.5 km per year. In 1962, a total of 21 mud banks were counted between these 2 locations (Delft Hydraulics Laboratory, 1962).
Any given point along the coast is normally the site of mud bank formation for a number of years, and then of erosion until the coming of the next bank. As the intertidal part of the bank is exposed to air at low tide, it hardens to a certain point and may become colonised by pioneer mangrove trees (Avicennia germinans), resulting in a seaward progression of the shore, and up to several kilometres can be gained from the sea during the few years of presence of the mud bank. This will not prevent erosion from taking place, however, once the mud bank phase has been succeeded by an “inter-bank” phase, as trees will be uprooted by the waves and the coastline will recede. In this fashion, the migration of coastal mud banks defines and modulates, for a large part, the environment at both sides of the shore. Froidefond et al. (1988) used aerial photographs to calculate the land gains and losses made around each bank over the years and concluded that they roughly compensated each other in front of French Guiana. Therefore the total mud bank area also remains constant. The velocity of the migration of the banks, however, is thought to vary by periods. On the very long term, there has been a net land gain in Suriname, where the coastline has progressed northwards, in the last 6 000 years, with an average speed of 2 m per year in eastern Suriname and 10 m per year in western Suriname (Augustinus, 1983).
It is important to note that the substrate on which the mud banks carry out their shifting movement is itself made of old consolidated mud deposits, of the same origin, that has accumulated during millions of years since the tertiary era. This type of substrate is found to the depth of 20 m and is replaced, further on the shelf floor, by a patchwork of sand and clay mixtures, then by sand of diverse characteristics. Another important feature, present along most of the continental shelf of the region, is a former (drowned) coral reef, located in the depth zone of 45-80 m, at the level of a former coastline. With the exception of French Guiana, detailed maps of the nature of the sea bottom are not available in the region. It has been observed (Vendeville, 1995, Charlier, 1995) that the type of substrate may play a more important role than the depth gradient in the distribution of fish assemblages.
The Brazil - Guianas continental shelf shelters relatively productive fishing grounds, compared with surrounding marine ecosystems. Fundamental biological processes on the Brazil-Guianas shelf have not yet been really studied. There are several research programmes under way, starting to systematically record basic ecological parameters. In French Guiana, the “Programme National Environnement Côtier” pursues a multidisciplinary approach of the coastal environment. Regional co-operation has been implemented through the “Ecolab” programme. For the time being, mechanisms governing the different levels of production are not quantified. Important factors for this productivity are, however, known to be:
It is difficult to point to elements, within this large marine ecosystem, that would have a markedly greater influence on the productivity, and particularly, on fisheries productivity. Traditionally, attempts have been made to find particular biotopes where key processes (spawning places, nurseries, food sources) in the biological cycle of important species are thought to concentrate. The identification of such places would open possibilities for taking particular protection and/or management measures.
Mangrove forests are sites of high primary (vegetal) production. Particularly on these coasts, the mangrove forests are said to exhibit exceptional growth rates. This high productivity is due to the important nutrient supply from inshore, but also to the dynamic characteristics of the coastal landscape. Large parts of the mangrove forest are regularly washed away, before reaching their “climax” stage at which productivity decreases, while new areas are colonized, on top of mud banks, by fast growing pioneer trees. The transfer of this vegetal biomass to other production levels starts with its degradation, by bacteria, fungi, etc. into elements that can be mobilized by a quite diverse detritivorous community. Detritivorous species, among which are many small crustaceans but also juveniles of decapods and of fish species, are the key players in this trophic chain.
These processes, however, do not necessarily take place in the mangrove forest itself. Organic matter is exchanged with the adjacent coastal waters, where the next trophic levels are much more abundant. Therefore, the many fish species, often cited in published reports to illustrate the importance of the mangrove forest for the marine resources, are really associated with the productivity of the mangrove rather than spending a important part of their life cycle in the mangrove forest.
It is probably in the shallow fringe of coastal waters, on and around mud banks, that a major part of trophic transfers involving exploitable species takes place. Isolated experiments in French Guiana and Suriname have detected important concentrations of fish eggs, larvae and juveniles (Vendeville, 1995). It is particularly complicated to operate sampling devices on the very soft bottom of this zone, and this has hampered attempts to investigate its role in a quantitative way. The shallow fringe apparently serves as nursery place for many marine fish species. It may also be a key habitat for shrimp species. Penaeus subtilis nurseries, for instance, that have been found to date in coastal marshes like Mana (Rossignol, 1972), cannot account by themselves for the entire recruitment into the adult shrimp fishery.
The ecology of the mud banks is only now being investigated, particularly in French Guiana (see above). Diatoms seem to account for a high primary productivity (Daniel Guiral, personal communication). On the other hand, densities of Tanaid crustaceans of up to 13,000 individuals per m2 have been observed and are attracting large numbers of shore birds (Teunissen, 2000). According to Frouin (1997), polychaet annelids are abundant on the soft mud substrate.
The role of estuaries (other than the Amazon’s), as privileged zone for mixing of fresh and seawater, also remains to be explored. Personal observations in Suriname (large numbers of juveniles caught by Chinese seines in certain months) suggest that they may play a particular role in the recruitment of some fish species.
There is a zone considered by fishermen as very important for brown shrimp (Penaeus subtilis) recruitment in Brazil (José A.N. Aragão, personal communication). Situated between 00°20’N to 01°10’N and 47°00’W to 47°55’W, it is called “lixeira” by fishermen (“rubbish zone”), because of the concentration of very small shrimp, associated with an important zone of seaweed production. Owing to its geographical position, this area must be a mixing zone of Amazon river water and oceanic water and is therefore certainly the site of high primary production. Trawling in that area has been prohibited year round by the Brazilian authorities.
Water salinity and temperature, currents, rainfall and river flow are expected to shape, to some extent, natural productivity and fisheries. Since Lowe (McConnell) showed in 1962 that most fish species had long, not well defined breeding seasons, however, very few relationships between any of these factors and seasonal features of fish or shrimp life cycles have been demonstrated. In the case of brown shrimp, Penaeus subtilis, rainfall maxima and recruitment peaks were found to coincide in several countries (Garcia et al., 1984, Charlier et al., 1995, Maharaj et al., 1998, Ehrhardt et al., 1998). Attempts to link inter-annual variations of shrimp recruitment (or other vital parameters) with environmental factors have not been successful yet, however.
Longer-term cycles have been invoked to explain variations, although their influence has never been demonstrated, and the mechanisms of their possible impact remain completely unexplored. Solar activity cycles have been mentioned as possibly connected to rainfall and brown shrimp recruitment (Garcia et al., 1984). El Niño Southern Oscillations (ENSO) are reflected, in this part of the world, by drought episodes during El Niño events. Mol et al. (2000) investigated the link between breeding success of fresh-water armoured catfish (Hoplosternum littorale) in Suriname and the occurrence of El Niño. Fluctuations in oceanic circulation are also likely to exert an influence on production processes in the coastal ecosystem. Finally, global warming could also be expected to impact this ecosystem, as it does many others.
4.3.1. Direct Effects
4.3.2. Impact on the Structure of Fish Communities
4.3.3. Indirect Effects
4.3.4. Trophic Effects
4.3.5. Modifications of the Physical Environment
A series of studies were undertaken under co-ordination of the International Council for the Exploration of the Sea (ICES). Based on these studies, an ICES Working Group concluded in 1988 that fishing possibly had effects on the benthic communities, but that these effects could not be quantified, as they combined with other causes like pollution and eutrophication. More recent investigations have taken a broader scope, and considered the entirety of the ecosystems. In March 1999, ICES has convened a symposium on ecosystem effects of fishing. Much of the information available worldwide on this broad subject has been reviewed on this occasion. Since ecosystem effects of fishing have been little studied in the Brazil - Guianas region, several cases are taken, in the discussion below, from contributions to that symposium.
Among the many ways in which fishing interferes with the ecosystem, the most obvious are its effects on exploited marine organisms populations, inflicting fishing mortality, decreasing biomass, modifying age structure. It is the object of stock assessment methodologies, in the most classical sense, to investigate, which levels of these direct effects are compatible with a sustainable exploitation.
Practically all fishing gear also catches non-target species that cannot, in general, be returned alive to the sea. This unintended fishing mortality has reduced drastically several fish populations around the world, particularly demersal species in areas where there is an intensive shrimp trawl fishery. Fish populations can even be reduced outside the fishing grounds. Chong et al. (1998), cited by Blaber et al. (2000), mention fin-fish populations in 20-60 m depth being reduced by 60%, in Malaysia, as a result of shrimp trawling in a shallower area, where large amounts of young fish were destroyed. This shrimp by-catch issue is well known in the Brazil-Guianas region, where studies have, in several countries, attempted to quantify the impact on several commercial species. In Venezuela, for example, the by-catch taken by the shrimp trawlers was estimated at 96,000 tonnes annually, representing 22% of the national fish and shellfish production (Alió and Marcano, 2000). By-catch/shrimp ratios are typically between 5 and 15 to 1 in the region (Villegas and Dragovich, 1984, Marcano et al., 1995). Analysis of the species and sizes composition of the by-catch reveals that many commercial species are included, that only a small part is utilised, and that undersized (young) individuals generally predominate. It is also felt that the species composition has changed over the years and that several species have practically disappeared from the by-catch, indicating a dramatic shrinking of their populations, notably in the case of sharks (José Alió, personal communication). Effects of fishing on Chondrichthyans has been the subject of considerable attention because species of this group are generally characterised by biological parameters that make them particularly vulnerable to overfishing (slow growth, low reproductive potential, position at or near the top of food webs) and also because a large part of their catch is taken as by-catch, which means an absence of management. A review provided by Stevens et al. (2000) discusses the position of sharks and rays in ecosystems worldwide and shows, for example, that there is a danger of local, if not global extinction for some species.
Finally, two cases of direct impacts of fishing on marine organisms populations can be quickly mentioned, for the sake of comprehensiveness of this outline, but will not be further developed upon, as they do not work back on the productivity of the system (from a fisheries point of view). There is the problem of ghost fishing (fish being killed by abandoned or lost fishing gear), which is included, for example, in an overview proposed by Goñi (1998). There is also the case of accidental capture of marine mammals and turtles. Abundant literature is available on this subject, and in the best cases, the mortality inflicted by fishing gears has been estimated and compared with other mortality causes. In Australia, for example, it was concluded that the northern shrimp fishery did not represent a major factor, compared to the other threats posed to turtle populations in that region (Poiner and Harris, 1996, cited by Blaber et al., 2000). Nevertheless, measures to reduce this mortality caused by fishing activities were introduced.
Because of the selectivity of fishing gears, and of differences between species in terms of vulnerability and resilience to fishing pressure, fishing generally induces, in the long term, changes in the species and size composition of fish communities. Many cases have been described in various regions of the world. In the North Sea for instance, trawl survey data collected in 1906-1909 and in 1990-1995 have been compared (Rijnsdorp et al., 1996). A decrease in species diversity was observed, as well as a shift in the size distributions towards smaller sizes. Although differences in survey gear used in each period made comparisons difficult, the data indicated a decrease in general abundance.
There are also well-documented examples from lower latitudes. In the Gulf of Thailand, the expansion of the trawl fishery from 1961 caused a severe decrease in abundance of large, long-lived fish and of several families of smaller fish (Pauly, 1988) and an increase in shrimp and squid. In another tropical environment (Southeast Gulf of Carpentaria, Australia), Harris and Poiner (1991) compared results of scientific surveys carried out before the start of the prawn fishery (1964) with those obtained, with similar technology, 20 years later (1985-1986). They examined the changes that had occurred in the abundance of 82 taxa (fin-fish species or small groups of species). They found that 52 of the 82 taxa had not changed significantly in abundance, 18 had decreased and 12 had increased. Taxa showing an increase were generally bentho-pelagic, while benthic taxa, more vulnerable to the prevalent fishing gear (prawn trawl), had more often shown a decrease. Discussing possible causes of observed changes, they suggested that fishing effort and discarding by-catch could be invoked as main cause for 18 taxa only, and that changes for other taxa were related to factors like environmental modifications (themselves possibly caused by fishing).
The relatively low level of impact of fishing on the fish community structure in this case, compared with what happened in the Gulf of Thailand, was interpreted as a result of lower level of fishing intensity in that region. Scientists are attempting to relate structural changes observed in fish communities to the level of fishing exploitation. Several methodologies have been elaborated to quantify these structural changes, based on diverse indexes reflecting overall size composition, diversity (in species and sizes), and dominance (whether many species are more or less equal in importance or there are dominant components). One of the final objectives would be to be able to assess the degree of exploitation by measuring diversity, dominance and size spectra through relatively simple indexes, particularly in data-limited situations. These indexes could also be useful to compare the behaviour of fish communities located in different types of environment, when exposed to various levels of fishing. The effectiveness of such indexes at providing diagnostics is discussed by Bianchi et al. (2000). While there is enough evidence that intensive fishing alters size spectra, as average length of fish caught decreases, the rate of change varies between ecosystems. In tropical environments, it appears, for example, that increases in fishing effort are much less sharply reflected than in high-latitude environments and that an increase in diversity is often observed in response to heavy exploitation. In any case, this approach provides a unique tool for the analysis of changes, within a given ecosystem, over time. Data collected by surveys and other observation programmes at different stages of fisheries development therefore have an irreplaceable value.
In the Brazil-Guianas region, Bianchi (1992) provides a description of fish assemblages, but an analysis of the type described above has not been undertaken yet here. Basic data collected in different periods are available, however, in several countries. They have been generated by a number of surveys, starting in the 1950s, conducted with research vessels, generally based on sampling by bottom trawls, and more recently on board of commercial vessels chartered for investigation purposes. How appropriate and comparable data from different surveys are remains to be investigated, but the bottom trawl, as a sampling device, has in any case undergone little modification in those decades. It is believed that comparisons of the results obtained by the earliest and by recent surveys would provide useful insights of the changes in abundance of main species, and help understand the impact of fishing on this ecosystem. In several countries, moreover, observer programmes are carried out, generally on trawlers. Even when such programmes do not necessarily cover extended periods, they provide valuable material for comparisons. In Venezuela, for example, there is an on-going observers programme on shrimp trawlers (Marcano et al., 1995). In Brazil, surveys have been carried out by federal agencies and recently with the research vessel “Paulo Moreira” (José A.N. Aragão, personal communication). In Suriname, an observer programme has been run, on board several types of trawlers, from 1994 to 1999. Partial results were discussed in local reports (Charlier, 1996). There is currently no observers programme in French Guiana, but several short-term surveys have been conducted in the nineties (Moguedet et al, 1994, Vendeville, 1995) and a series of 3 cruises is planned for 2001 (“GREEN” Program: “Guyane REsource ENvironment).
Indirect effects of fishing on the ecosystem include trophic effects, acting on the different components of the food webs, their interactions and their dynamics, and effects produced by modifications brought by fishing to the physical environment.
As all organisms present in a marine ecosystem feed on each other, variations in some species’ biomass can impact other species because they are prey, predators, or competitors for common prey. A well-known interpretation of the effects of exploitation is the famous “fishing down of the food web” theory (Pauly et al., 1998), whereby the species located in the superior part of the food web (generally large size, slow-growing) are the first to suffer biomass reduction as fishing pressure raises. As the higher trophic levels become depleted, the catches are increasingly made up of (generally smaller and fast-growing) species belonging to lower trophic levels.
Fisheries on demersal resources of Georges Bank may provide an illustration for another facet of trophic effects. There, increasing exploitation in the 1960s more than halved total fish biomass. However, the decrease in abundance of target species (gadoids and flounders) coincided with an increase of other species (dogfish and skates), occupying a similar niche (Fogarty and Murawski, 1998). According to Garrison and Link (2000), the global trophic structure of that fish community (the proportional importance, in biomass, of each trophic level) has remained static.
Trophic effects result from numerous and intricate interactions, and there is considerable speculation about which types of mechanisms are prevalent. How much is the abundance of prey species constrained by predator populations? To which extent does the production at the lower trophic levels vary independently and govern the biomass of species at higher trophic level? Answers to these questions probably vary from ecosystem to ecosystem, but they are critical for understanding changes brought about by fishing and by other causes. The so-called mass-balance trophic models (for example, Christensen and Pauly, 1992) provide a useful analytic tool. These models try to describe and quantify, as accurately as the data allow, trophic relationships between all species (or groups of species) participating in a system. Such steady state models have also been extended to form dynamic ecosystem models, as in the case of ECOSIM (Walters, Christensen and Pauly, 1997). Used in this way, they can also be used to investigate likely effects of modifications (brought about by fishing strategies, regulations, etc.), to compare different states, in real life, of ecosystems, to explain changes that have occurred, and relate them to fishing. This approach is closely related to the one discussed above, dealing with fish community structures, as feeding interactions represent a major structuring factor in a marine ecosystem. The first approach intends to capture global trends, while the second one proceeds the other way round, focusing first on the mechanisms within the system.
A rich literature exists on the application of mass-balanced models to investigate various ecosystems. Also, for the Brazil-Guianas region, they represent yet another path for understanding the developments in fisheries and their environmental impacts.
Fishing gears can alter, in a more or less persistent way, the habitats of fish populations. We do not know, generally, what the different species exactly require in order to complete successfully the different steps of their life cycle. But we do know that habitats have to fulfil different functions as shelter (hiding from predators), foraging area for food, breeding area, nursery area, and the capacity to fulfil one or more of these functions can be impaired by damage provoked by fishing gear action. The damage can be on the seabed, the benthos, or on the water quality.
Seabed
Bottom trawls are, of course, the type of gear impacting the seabed most directly. Their otter boards imprint tracks on the seafloor, ploughing grooves that can vary from a few cm up to 0.3 m deep (Jones, 1992). The area between these grooves is affected by the footrope, and eventually by bobbins and chains disposed along the footrope. Trawling can also be responsible for some re-arrangement of sediment cover. Fine particles, in particular can be re-suspended, be transported by currents over some distance to areas with another type of substrate, and interfere there with the settlement of benthic organisms. Several authors have shown that a modest layer of silt, for example, can inhibit the settlement of bivalves.
The higher the total gear weight, the more visible and durable the tracks on the seafloor. The effects vary also in accordance with the type of substrate, as gear will plough deeper in soft bottom than in hard bottom. They depend as well on the water movements close to the bottom, being stronger and more durable in waters that are not affected by strong currents and tides. Krost et al. (1990, cited by Jones, 1992) found that tracks remained for 5 years, in sandy mud in the Baltic Sea. Finally, the proportion of the area that is actually swept plays a role in the extent of eventual damage. Calculations have been made in some heavily exploited fishing grounds. In the Kiel Bay (Baltic Sea), Krost et al. (see above) found that 19% of their study area bore trawl tracks. In the Netherlands, Rijnsdorp et al. (1991, cited by Jones, 1992) calculated that every square meter of the seabed is trawled, on average, 7 times a year.
What consequences perturbations of the seabed can bring for the fish populations is difficult to establish. Mobile fishing gear tends to smooth out structures on the bottom and reduces habitat complexity (Dorsey and Pederson, 1998), thus the number of niches available for marine life. Langton (1998) discusses the case of larval cod, evolving from eggs and larvae stages distributed in the water column, that “settle onto the entire surface of Georges Bank but, because of predation, [they] are very quickly restricted to an area of gravel pavement on the northern edge”.
In the Brazil-Guianas region, the extension of the zones that are actually used by the shrimp fishery (and other types of trawlers) has not been estimated. In Suriname, it is considered that the shrimp fleet, in the course of the first years of operation, has deliberately cleared the area useful for shrimp trawling of most obstacles small enough to be removed.
Benthos
Communities of benthic organisms represent a valuable part of the marine ecosystems, and they play an important in the life cycle of exploited species and of important organisms in the food chain. A relationship between damage to the benthos and changes in some fish species populations has been observed well before the mechanisms responsible for this relationship could be identified. On the shelf off northwestern Australia, a trawl fishery has been targeting snappers (Lutjanus genus) and emperors (Lethrinus genus). Abundance of these species declined in 1962-1983 (Sainsbury, 1988, cited by Bianchi et al., 2000), and increased again after the shelf was closed to trawling. Among several possible explanations, these variations were shown to be most likely related to alterations of the habitat structure (Sainsbury, 1997, cited by Bianchi, 2000), caused by the destruction of large epibenthic organisms (sponges, alcyonarians, gorgonians) by the trawl gear (and their recovery after closure). The biomass of species Nemipterus spp. and Saurida spp., occurring mostly over open sand, had increased in the trawling episode. In Tasman Bay, New Zealand, Saxton (1980) noted a decline in juvenile fish with the removal of bryozoan beds (cited from Jones, 1992).
The damage inflicted to benthos varies in accordance with the type of gear, the type of substrate and the type of benthic community. Several general observations emerge from much experimentation reported in the literature (including Creutzberg et al. 1987; Stevens 1990; Jones 1992; Pranovi et al. 2000; Bergman and Hup 1992).
Soft substrates offer little resistance to the action of mobile gear; therefore soft bottom communities are easily disturbed by fishing. On the other hand, they appear to recover faster than other communities from injuries. As soft bottoms are reworked by natural water movements, the communities already seem to be adapted, with a majority of short-lived, fast growing components. In general, in more variable environments, damage caused by fishing is short-term.
Heavier fishing gear is often used on sandy/hard bottom, in order to catch species that burrow or cling to the bottom (like some shrimps), than on soft bottom. The benthos may be more resilient to damage, but it is made of species with a lower turnover, that will take longer to recover.
The frequency of passage of the gear is an important factor. Permanent damage is likely to occur if the time between passages is shorter than the recovery time. Here again, there is an important difference between the two types of substrate.
Fishing alters benthic communities in the same fashion as explained above for the fish communities, the biomass of slow-growing species decreasing, to the advantage of faster-growing, often smaller, species. The impact of such changes on the associated shrimp and demersal fish communities is found to be negative in most cases, and the more so where the species have developed tight dependency bonds (in the more stable environments). Some species, however, are able to benefit from these changes and see their biomass and/or productivity increase as a consequence of fishing. Among these are scavengers, that take advantage of the organisms damaged or exposed by the passage of the gear. Another example is the North Sea sole that has benefited from the shift of the benthic community to small species appropriate for its small mouth.
How do these observations from various regions apply in the Brazil-Guianas region? Two main types of substrate can be distinguished in the fishing grounds. In the coastal area down to the depth of 20 m the substrate is soft mud, devoid of epibenthos. The in-fauna is probably already dominated by short-lived, adaptable species, and perturbations caused by passing fishing gears are most likely negligible, compared with the movements imposed by currents, tides, etc. Trawling is impossible in part of the zone, at the level of the banks, made of fluid mud. Other fishing gear types used in shallow water, notably the drifting gillnets, are quite harmless on this type of sea floor. Therefore, it does not seem that fishing could impact benthic communities of that zone enough to affect its role as nursery for many species.
Important trawling grounds located beyond this shallow fringe have firmer types of bottom, made of hardened mud and sand in varying proportions. There is no description of the benthos before trawling began, and even the present aspect of the bottom has not been studied, but observations from other areas suggest that benthos on this type of substrate can be damaged by fishing. Within the depth range considered shrimp trawling ground, distribution of fishing effort is heterogeneous. Productive areas are probably trawled several times a year. In a comparable context (north-western continental shelf of Australia), Moran and Stephenson (2000) found that a demersal otter trawl caused a reduction of 15.5% in large epibenthos density (sponges, cnidarians, bryozoans), for each trawl pass. Four trawl passes reduced the density to about a half. Recovery time was not investigated. In another area (in Florida) heavily fished by prawn trawlers, recovery time was estimated as less than a year (Tilmant, cited by Jones, 1992). The knowledge on the mechanisms of interactions between macrobenthos cover and fish populations is very limited. In some instances, management action has been taken without waiting for research results (in the Australian case above, trawling was prohibited within 50 m depth), on the assumption that habitat modification could add to the effects of direct fishing mortality and lead to reduced fish abundances.
Water Quality
Depending on the type of gear and of substrate, trawling can be responsible for a variable amount of sediment re-suspension, increasing the sediment load and the turbidity. By decreasing the penetration of light, this can interfere with the production processes in the lower part of the water column and on the bottom.
Together with inorganic sediments, organic elements could be re-suspended by fishing gear. This process could foster re-mobilisation by primary producers of nutrients trapped in the seabed. On the other hand, mixing surface and subsurface sediment layers could create anaerobic conditions close to the bottom, killing or severely impairing development of most floral and faunal forms of life. In coastal waters, small-scale vertical oxygen gradients can be critical (Jones, 1992). Caddy (2000) adds that re-suspension of anoxic soft-mud sediment results in release of toxic hydrogen sulphide and pollutants. Each of these processes could take place on the Brazil-Guianas shelf, particularly on the shallow, soft-bottom coastal fringe.
When a significant proportion of the biomass brought up by the fishing gear is discarded, the dumping of this organic material can have an impact. Although birds, sharks and other fish clear some of the discards of shrimp trawlers, most of it sinks towards the bottom where a part is scavenged by crabs and fish, and a part decomposes. In the Brazil-Guianas region, the amounts of dead marine organisms dumped by the shrimp fishery are in the order of several tens of thousand tons in each country. Such quantities are no likely to be cleared quickly by demersal scavengers. Dissolved oxygen levels close to the bottom could therefore be affected, when many boats operate in the same area, and particularly in shallow waters. In New Zealand, Rutherford (1987) calculated that oxygen concentrations could be reduced to 45-55% saturation by fishery waste.
In Suriname, small-scale fishermen have been complaining about “dead waters”, observed in shallow areas, following fishing activity by trawlers. These “dead waters” were scattered, according to these observations, with dead fish in larger amounts than could have been discarded by the trawlers. A vast area was devoid of live fish, as they apparently either had died or fled. Such mortality could be the result of local oxygen depletion, caused by the re-suspension of anoxic sediment combined with the presence of organic matter dumped from the vessels. The risk of such an effect is amplified if there is no vertical movement in the water column, as happens in certain tide and current situations. Shrimp fishermen have also reported cases, in deeper waters (20-40 m) of temporarily “dead zones”; where no live fish and no live benthic organism was to be found. These events were generally associated with observations of water “changing colour”, which evokes the possibility of planktonic blooms.
4.4.1. Agriculture
4.4.2. Industrial Development
4.4.3. Construction of Infrastructures
Caddy (2000) called impacts on marine ecosystems, generated by land-based human activities, Marine Catchment Basin (MCB) effects. These effects are particularly crucial in enclosed or semi-enclosed seas, but they are also important in the case of ecosystems that are strongly influenced by river run-off, as in the shallow areas of the Brazil-Guianas shelf. The coastal zone in the Brazil-Guianas region has not been the theatre of spectacular economic or industrial development. The largest part of this coast is even virtually untouched by human activities. Urban development is concentrated in the neighbourhood of river mouths and on riverbanks close to sea. Human impact is probably the highest at both extremities of the region: on the right bank of the Amazon-Para estuarine system, and along the Gulf of Paria, on the Trinidadian side. Due to the hydrodynamics of the region, the central, largest part of the area may remain basically unaffected. Effluents from the relatively industrialised and populated Belém region, situated on the East bank of Pará River, are barred from this region by the outflow of the Amazon and Pará rivers. Effluents from the west coast of Trinidad reach the Gulf of Paria and, given the Northwards current through the Columbus Channel, cannot impact any region to the South. The following land-based human activities could have a potential impact on the marine environment and on fisheries in the region.
Agriculture is a mainstay of the economy throughout the region. It has a direct impact on marine environment when it claims areas included in the “broader marine ecosystem” (including brackish zones connected with the sea). These areas are generally not suitable for agriculture, due to their salt content. It has been a policy, in Suriname for example, not to allow agricultural projects within a certain distance from the coastline, for technical as well as environmental reasons. Exceptions have been tolerated and there have been encroachments on the mangrove and associated brackish-water biota, for rice (as well as in French Guiana) and livestock development, but the areas concerned seem to remain modest.
Areas within the coastal brackish-water belt have been identified as suitable for aquaculture, particularly for marine shrimp. A few projects have been undertaken in Suriname, which do not cover, at this time, any significant part of the available area. Aquaculture seems to have a potential for development, however, and more extended areas could be affected in the future.
Effluents carrying chemicals used in agriculture or residues can also be expected to have impacts if they are drained in sufficient concentration to the sea. They can be pesticides, deleterious for some marine organisms, or fertilizers and alter the nutrient balance in the sea.
Eutrophication can be the result of increased nutrients input in an aquatic ecosystem. Such occurrences have been described in many parts of the world. While natural enrichment is vital to productive fisheries (Caddy, 2000) and additional nutrient inflow could, in first instance, boost primary production, the overall balance of the ecosystem is impaired by excessive levels, its structure can be affected, with shifts in species composition, reduction of the system diversity and complexity, and the appearance of new dominant species. The recent (over the last 30 years) increase, by a factor of 2.5 to 10, in the total biomass of benthic organisms on the Dogger Bank, is probably the result of organic matter input from populated countries surrounding the North Sea (Kröncke, 1990). Caddy (2000) notes that the Mediterranean Sea has become more productive, in terms of fisheries, since 1970, “especially in the northern statistical areas under the influence of the rivers Rhone, Po, Ebro”, draining from populated and agricultural areas.
Less desirable effects can take place, like the creation of hypoxic conditions, such as have been temporarily observed in the Gulf of Mexico, due to Mississippi River discharge, and in a number of study cases mentioned by Caddy (2000). Mainly in enclosed seas but also on shelves, hypoxic conditions can become chronic close to the bottom, destroying benthic life, as eutrophication further progresses, if there is not enough vertical mixing. In another example of negative and unpredictable indirect effects, a zooplankton increase in the Black Sea, caused by eutrophication, first fostered pelagic fish production (anchovy landings doubled), and then triggered an invasion by the ctenophore Mnemiopsis, which came to dominate the pelagic ecosystem and apparently caused the decrease of anchovy production to a tenth of its peak value (Volovik et al., cited by Caddy, 2000).
Such dramatic effects are unlikely to occur in the Brazil - Guianas region. Agricultural development can be described as very modest in terms of area used. On the other hand, farmed areas are concentrated in a coastal stretch of a few tens of kilometres breadth, the cultures are water-intensive (rice) as well as agrochemicals-intensive (sugar cane, bananas), and the drainage is directly to the sea, without treatment or monitoring of the effluents. Local effects could therefore be observed, particularly if input of an additional nutrient charge from agriculture would combine, for example, with re-suspension of organic matter trapped in sediment. Some local publications report on the concentrations of different chemicals per hectare per year allowed and/or actually applied. But estimations of amounts of nitrogen and phosphorus components generated by agriculture or urban concentrations, carried by rivers and reaching the coastal ecosystem could not be found. Esack (1991) and Boodoosingh (1992) studied the potential impact of agrochemicals on fisheries in Trinidad and Tobago. The application of large amounts of potentially hazardous chemicals, particularly in large-scale sugar cane estates, is reported. Except for occasional fish kills observed in inshore waters, connected with the use of insecticides, no concrete data on actual impact on marine life seems to be available.
Industrial activities are scarce and small-scale in the region. There is a potential risk of pollution, however, from gold exploitation along the upper parts of several rivers, and from oil extraction, both in the coastal plain and at sea.
Several mineral resources are being exploited in the region. The bauxite extraction plays a major role in the economy of (at least) Suriname, Guyana and Venezuelan Guyana. Significant threats posed to the environment by this industry are not documented in the literature. Coastal environmental reviews and coastal area management plans such as Teunissen, 2000, mention that effluents from bauxite mines and refineries may contain toxic substances, without further specification.
Gold is being exploited on a small-scale in all countries of the region. The main technology used to separate and amalgamate gold is the least expensive available and involves the application of about 1 g of mercury per 1 g of gold (Ramlal, 1999). This mercury is dispersed into the air and it is assumed that the largest part ends up in rivers, transforms into methyl-mercury and other chemical compounds, and concentrates along the food chain. According to the IUCN (1996, cited by Teunissen, 2000), high concentrations of mercury were detected in fish as far as 800 km downstream from gold mining areas in Brazil. Recently initiated investigations in Suriname indicate that mercury concentrations in fish may approach maximum norm (0.5 mg/kg) in piscivorous fish from the upper course of rivers, close to gold mines, and decreases towards the middle and lower course (Ramlal, 1999, Quik, 2000). If nothing was done to limit the use of mercury (that also directly threatens the life of the goldminers), mercury contamination could on longer-term also become a hazard for the coastal marine ecosystem. Larger scale gold mines, present as well in the region, use cyanide to separate gold. Errors can have disastrous consequences for aquatic life, as happened in 1995 in Guyana with the accidental release of cyanide into the Omai and Essequibo rivers.
Oil exploration started in 1980 in Suriname, with the creation of the State Oil Company. Fields have first been opened on-shore. Total reserves of the main one, Tambaredjo oil field were estimated at 165 million barrels, and oil production reached 10,500 barrels per day (from 421 wells) in 1997 (Teunissen, 2000).
Exploration for new reserves is carried out at several locations on-shore but also offshore in Suriname and in Guyana. Several boreholes have been drilled in recent years in the shallow coastal zone in Suriname waters (Teunissen, 2000), and several of them indicated the presence of oil and gas.
According to Teunissen (2000), the risk of a major oil spill at sea is present. The government has approved Resolutions to prevent pollution. Farmers and fishermen have reported minor oil spills in existing oilfields. The risks that this type accident would bring for the environment, particularly the marine ecosystem, have not been quantitatively assessed. New oilfields could be developed shortly in the mangrove belt. This would involve the drainage of the areas concerned, and could result in the destruction of parts of the mangrove.
Boodoosingh (1992) has drawn a list of human activities with potential effect on fisheries in Trinidad, particularly along the west coast, one of the more industrialised areas in the region. There are industrial concentrations at certain locations on the coast (Port of Spain, Point Lisas, etc.). Except for the possibility of environmental hazard if these sites were not properly monitored, there is no concrete indication of actual or potential effects on the marine ecosystem.
The development of infrastructures with possible impact on the marine environment is very limited, throughout the region. The construction of dams in Suriname (Brokopondo, in the 1960s) and in French Guiana (Petit Saut, in the 1990s) is worth mentioning. They have been preceded by environmental impact assessments and are not considered to have had any impact on marine fisheries.
The assessment of various types of impacts on an ecosystem requires knowledge of the fundamental mechanisms driving it. A detailed description of the ecosystem in pristine state would obviously be an ideal starting point, but this is generally not available, even in the areas that have been most studied. Fisheries exploitation has started long before the importance of ecological processes was recognised. Information collected at a later stage, after the system has already been subjected to different types of constraints, is at hand in the best cases. In order to understand the direction the system is taking under the influence of new factors, data recorded at different stages of development or of impact, need to be compared. In the case of the Brazil-Guianas marine ecosystem, the information is particularly scarce. The study of the fundamental processes (production and transfer between trophic levels) has only been started in recent years, and results are not yet available. It is therefore necessary to assume that the processes prevailing in various marine ecosystems must have similarities and to use the information from those that have been better studied in interpreting characteristics and behaviour of the Brazil-Guianas systems, while still considering the unique characteristics of these systems.
The marine ecosystem present on the continental shelf of the Brazil-Guianas region can be divided into two major components. The first one is the coastal segment under the predominant influence of the Amazon River and, secondarily, of other rivers; the second one is oceanic, located along the seaward edge of the coastal component.
The coastal component is characterised by its variability. Basic water quality parameters like salinity, turbidity exhibit ample variations, the sea bottom is covered by fine sediment alternatively re-suspended by currents and waves and re-deposited. On its landward edge, the coastline is unstable. Vegetal as well as faunal communities are necessarily adapted to these variable conditions. There is apparently no macrobenthos, and the mud banks are colonized by few species, present in large numbers. The trophic pyramid appears to be based, for the largest part, on the recycling of (part of) the abundant terrigeneous organic matter by microorganisms, fish larvae, crustaceans, polychaetes, etc. The physical perturbations caused by fishing are of a much lower scale than this perpetual movement and it is hard to see how a fishing gear could have a significant impact on this fluid structure.
This system is probably much more dependent on the fluctuations of terrigeneous inputs, nutrients and physico-chemical parameters, regulated by natural laws, however poorly known these are today, than it is on the impacts of fishing gear. This also suggests that this system could be vulnerable to pollution and chemical threats generated by inshore or onshore developments. No sure signs of pollution impacts have been recorded yet, which does not mean much more than that there is no monitoring. Mass mortalities of fish have been reported, however. Whatever the cause (oxygen depletion due to a conjunction of factors as hypothesised above, bacterial or algal infection, chemical contamination) they seem to confirm the vulnerability of the system to this type of impact. While human activities beyond the coastline and in the marine catchment area have been, for the time being, on a modest scale, this situation could change and their impact could become noticeable in the future. It is, indeed, foreseeable that aquaculture, timber exploitation, and other activities will come to a greater development and claim a larger share of the coastal area.
The oceanward component of the ecosystem, where most trawlable grounds are located, including those exploited by shrimp trawlers, is more likely to be affected by physical impact of fishing gear. Although it is also indirectly dependent on the inputs from rivers, seasonal oscillations of its parameters are of much lower amplitude. Its biota do not need to be adapted to variable conditions and are, therefore, more vulnerable to impacts by fisheries.
On the other hand, given the proximity of the very dynamic, muddy coastal strip, shifts probably occur in the distribution of superficial sediment types. Harris and Poiner (1991) attributed some of the changes observed in the status of demersal fish species in Southeast Gulf of Carpentaria to an increase in the proportion of the area covered by a muddy substrate between 1964 and 1985. Similarly, in the Brazil-Guianas region, changes in the abundance of particular fish species or assemblages could be related to modifications in sediment distribution and composition. Sediment surveys will be required, if the role of this type of causes, compared with that of fisheries, is to be investigated.
1. The effects of fisheries on the environment are considerable in the Brazil-Guianas region and can affect fisheries productivity. The intensity of these effects depends on local conditions, particularly the type of substrate and benthic cover, and the natural variability of the environment. They may be more marked, as far as the benthic environment is concerned, in the outer part of the shelf (beyond 20 m depth) than in the shallow, coastal, soft-bottom zone. But the impact on the structure of the fish communities themselves (species and size composition) is real both on the inshore and offshore components of the ecosystem.
2. The interactions between the fisheries and environment are difficult to define and quantify. Understanding these interactions requires a detailed knowledge of the mechanisms of the ecosystem, including its natural variability, which is not currently available. Moreover, it combines with impacts of other factors, both anthropogenic and natural, which has an unknown mechanisms, and it is difficult to separate all these effects.
3. There are specific habitats within an ecosystem that are of particular importance for its global functioning and/or for the life cycle of exploited species. The identification, monitoring and protection of such critical habitats are the subject of increasing attention worldwide. In this region, there are clear indications that the entire shallow, brackish-water stretch along the seashore (0-10 m depth) plays a key role in the mobilization of nutrients and energy transfer in the lower levels of trophic webs, and serves as nursery ground for many marine fish and shrimp species. Additional research is needed improve understanding and quantify this role. In the meantime, activities leading to the destruction of substantial amounts of juvenile fish and shrimp, including small mesh-size trawling, should be restricted.
4. The existence and the capacity of this near-coastal zone to fulfil its role are highly dependent on inputs from the neighbouring mangrove and associated habitats. The mangrove is very well represented in the region, where it dominates a major part of the shore. It is possible that all portions of the coast would not have the same importance; some processes being perhaps concentrated in certain areas, possibly in estuaries. Ecological research is needed to identify such “critical” zones with a view to their conservation. There seem to be other particularly valuable areas, with a high primary production, like the “lixeira”, in front of the Amazon estuary. Research on the processes taking place in these areas is required as well, for the elaboration of an appropriate management strategy.
5. The scope of investigations carried out in support of fisheries management should be broadened to take ecosystem considerations into account. In this respect, several directions for research are suggested:
6. Since all the concerns discussed above are common to all the countries of the region, and the costs involved in addressing them are considerable for any single country, regional co-operation is recommended.
ACKNOWLEDGEMENTS
I am very grateful to colleagues who have assisted me with their opinions, hints, bibliographic advices, particularly Suzuette Soomai, José Alió, José Augusto N. Aragão, Daniel Guiral and Philippe Vendeville, and indebted to FAO and Kevern Cochrane, who made this review possible, kindly revised the manuscript and suggested substantial improvements.
Alió, J.J. y L.A. Marcano, 2000. La pesca de arrastre en Venezuela y su futuro ante una sociedad con mayor conciencia ambientalista. FONAIAP DIVULGA, 63.
Augustinus, P.G., 1983. Coastal changes in Suriname since 1948. pp. 329-338 In: Bekker, D., and H. Ehrenburg (eds). Proceedings of the Furoris Congress on future of roads and rivers in Suriname and neighbouring region. University of Suriname and Delft University of Technology, 1982.
Augustinus, P.G., L. Hazelhoff and A. Kroon, 1989. The chenier coast of Suriname: modern and geological development. Marine Geology 90: 269-281.
Bergman, M.J.N. and M. Hup, 1992. Direct effects of beamtrawling on macrofauna in a sandy sediment in the southern North Sea. ICES Journal of Marine Science 49: 5-12.
Bianchi, G., 1992. Demersal assemblages of the northern coast of South America (Orinoco river to Maroni river). In: Bianchi, G. Demersal assemblages of tropical continental shelves. Thesis: University of Bergen, Norway.
Bianchi, G., H. Gislason, K. Graham, L. Hill, X. Jin, K. Koranteng, S. Manickchand-Heileman, I. Payá, K. Sainsbury, F. Sanchez and K. Zwanenburg, 2000. Impact of fishing on size composition and diversity of demersal fish communities. ICES Journal of Marine Science 57: 558-571.
Blaber, S.J.M., D.P. Cyrus, J-J. Albaret, Chong Ving Ching, J.W. Day, M. Elliott, M.S. Fonseca, D.E. Hoss, J. Orensanz, I.C. Potter and W. Silvert, 2000. Effects of fishing on the structure and functioning of estuarine and nearshore ecosystems. ICES Journal of Marine Science 57: 590-602.
Boodoosingh, M., 1992. A review of possible impacts of human activities on fisheries in the cosatal waters of Trinidad, with emphasis on the Gulf of Paria. Fisheries Division, Ministry of Agriculture, Land and Marine Resources; Port of Spain (Trinidad). Fisheries Occasional Paper Series No. 2., 126p.
Borstad, G.A., 1982. The influence of the meandering Guiana Current and Amazon River discharge on surface salinity near Barbados. J. Mar. Res. 40: 421-434.
Caddy, J.F., 2000. Marine catchment basin effects versus impacts of fisheries on semi-enclosed seas. ICES Journal of Marine Science 57: 628-640.
Charlier, P., 1996. Preliminary analysis of the results of the observers program over the years 1993-1995. Fisheries Department, Suriname, manuscript.
Charlier, P. and Y. Babb-Echteld, 1995. Report of the survey “S94C” of the brown shrimp recruitment investigation program. Suriname Fisheries Report No. 8.
Charlier, P., L. Samson-Pawironadi and Y. Babb-Echteld, 1995. National report of Suriname. FAO Fish. Rep. No. 526 (Suppl.): 63-86, also Suriname Fisheries Report No. 4.
Chong, V.C., A. Sasekumar, S.M. Phang and M. Nazari Jaafar, 1998. Marine and coastal resource mapping for the Straits of Malacca (Malaysia). Final report, Malacca Straits Demonstration Project, GEF/UNDP/IMO (unpublished).
Christensen, V. and D. Pauly, 1992. ECOPATH II - a software for balancing steady-state ecosystem models and calculating network characteristics. Ecological modelling 61: 169-185.
Cochrane, J.D., 1963. Equatorial undercurrent and related currents off Brazil in March and April 1963. Science 142: 669-671.
Creutzberg, F., G.C.A. Duineveld and G.J. van Noort, 1987. The effect of different numbers of tickler chains on beam-trawl catches. J. Cons. Int. Explor. Mer 43: 159-168.
Delft Hydraulics Laboratory, 1962. Demerara coastal investigation. Report on siltation of Demarara Bar Channel and coastal erosion in British Guiana. Delft Hydraul. Lab., Vols. 1-3. 240p.
De Master, D.J., and R.H. Pope, 1996. Nutrient dynamics in Amazon shelf waters: results from AMASSEDS. Cont.Shelf Res.16 (3): 263-289.
Dorsey, E.M. and J. Pederson (eds.), 1998. Effects of fishing gear on the sea floor of New England. Published by the Conservation Law Foundation, Boston.
Ehrhardt, N., D. Shepherd and T. Phillips, 1998. Length-based assessment of the brown shrimp, Penaeus subtilis, fishery in Guyana. Prepared at the second CFRAMP/FAO/DANIDA stock assessment workshop on the shrimp and groundfish fishery on the Brazil-Guianas shelf, held at Georgetown, Guyana, 18-29 May 1998.
Eisma, D., S.J. van der Gaast, J.M. Martin and A.J. Thomas, 1978. Suspended matter and bottom deposits off the Orinoco delta: turbidity, mineralogy, and elementary composition. Netherl. J. Sea Res. 12: 224-251.
Eisma, D. and H.W. van der Marel, 1971. Marine muds along the Guyana coast and their origin from the Amazon Basin. Contrib. Mineral. Petrol. 31: 321-334.
Esack, Y., 1991. The potential impact of pesticides on the Gulf of Paria coastal fisheries. FAO/UNDP Project “Integrated Coastal Fisheries Management”.
Froidefond, J.M., M. Pujos and X. André, 1988. Migration of mud banks and changing coastline in French Guiana. Marine Geology 84: 19-30.
Frouin, P., M. Pujos and P. Watremez, 1997. Revue des connaissances sur la zone côtière de Guyane Française. Programme National d’Océanographie Côtière, Cayenne.
Garcia, S., E. Lebrun and M. Lemoine, 1984. Le recrutement de la crevette Penaeus subtilis en Guyane française. Rapp. techn. ISTPM No 9.
Garrison, L.P., and J.S. Link, 2000. Fishing effects on spatial distribution and trophic guild structure of the fish community in the Georges Bank region. ICES Journal of Marine Science 57: 723-730.
Gibbs, R.J., 1976. Amazon river sediment transport in the Atlantic Ocean. Geology 4: 45-48.
Gibbs, R.J., 1980. Wind-controlled coastal upwelling in the western equatorial Atlantic. Deep-sea Research 27: 857-866.
Goñi, R., 1998. Ecosystem effects of marine fisheries: an overview. Ocean and Coastal Management, 40 (1): 37-64.
Harris, A.N. and I.R. Poiner, 1991. Changes in species composition of demersal fish fauna of Southeast Gulf of Carpentaria, Australia, after 20 years of fishing. Marine Biology, 111: 503-519.
Jones, J.B., 1992. Environmental impact of trawling on the seabed: a review. New Zeeland Journal of Marine and Freshwater Research, Vol.26: 59-67.
Kröncke, I., 1990. Macrofauna standing stock of the Dogger Bank. A comparison: II. 1951-1952 versus 1985-1987. Are changes in the community of the northeastern part of the Dogger Bank due to environmental changes? Netherlands Journal of Sea Research 25 (1/2): 189-198.
Langton, R., 1998. Bottom habitat requirements of groundfishes. In: Dorsey, E.M. and J. Pederson (Eds.) Effects of fishing on the sea floor in New England. Boston.
Lowe-Mc Connell, R.H., 1962. The fishes of the British Guiana continental shelf, Atlantic coast of South America, with notes on their natural history. Journ. Linnean Soc. London, Zool. 44 (301): 669-697.
Maharaj, L., N. Ehrhardt, D. Altuve, R. Alvarez and S. Soomai, 1999. Working group report on the length-based assessment of the brown shrimp resources from the Orinoco Delta-Gulf of Paria region (1992-1996). Rome, FAO. GCP/INT/575/DEN Report on Activity No. 46 (Supplement 1): 51-70.
Marcano, L., J.J. Alió, D.E. Altuve and J. Celaya, 1995. Venezuelan shrimp fisheries in the Atlantic margin of Guyana. National report of Venezuela. FAO Fish. Rep. No. 526 (Suppl.): 1-29.
Martec Limited, 1995. Environmental factors affecting fisheries of the Guiana/Brazil continental shelf area. Martec report n° TR-95-21. Prepared for the CARICOM Fisheries Resource Assessment and Management Program. Martec Inc., Halifax, Nova Scotia. 35p.
Moguedet, P., D. Nérini and F. Guégen, 1994. Evaluation du volume et cartographie des captures accessoires de la pêcherie de crevettes pénéides en Guyane française. Rapport final contrat CEE DG XIV 92/3504. 100p.
Mol, J., D. Resida, J. Ramlal and C. Becker, 2000. Effects of El Niño-related drought on freshwater and brackish-water fishes in Suriname, South America. Environmental Biology of Fishes 59: 429-440.
Moran, M.J. and P.C. Stephenson, 2000. Effects of otter trawling on macrobenthos and management of demersal scalefish fisheries on the continental shelf of north-western Australia. ICES Journal of Marine Science 57: 517-524.
Müller-Karger, E., C.R. McClain and P.L. Richardson, 1988. The dispersion of the Amazon’s water: where does the Amazon water go? Nature, May 1988.
NEDECO, 1968. Suriname transportation study. Report on hydraulic investigation. The Hague, Netherlands. 239p.
Nittrouer, C.A., and D.J. De Master, 1987. Sedimentary processes on the Amazon continental shelf. Pergamon Press, New York.
Pauly, D., 1988. Fisheries research and the demersal fisheries of Southeast Asia. pp. 329-348. In: Gulland, J.A. (ed.) Fish Population Dynamics. London, John Wiley and Sons.
Pauly, D., V.S Christensen, J. Dalsgaard, R. Froese and F. Torres Jr., 1998. Fishing down marine food webs. Science 279: 860-863.
Pranovi, F., S. Raicevich, G. Franceschini, M.G. Farrace and O. Giovanardi, 2000. Rapido trawling in the northern Adriatic Sea: effects on benthic communities in an experimental area. ICES Journal of Marine Science 57: 510-516.
Prost, M.T., 1990. Les côtes des Guyanes. Report for the “Programme Environnement Côtier”. ORSTOM, Cayenne.
Quik, J.A.M. and P.E. Ouboter, 2000. Water quality monitoring in the Commewijne watershed, Suriname.. WWF-GFECP, Project FG-06. Centrum voor Milieu Onderzoek, Anton de Kom Universiteit van Suriname, Paramaribo. 18p.
Ramlal, J.S., 1999. The effects of small-scale gold mining in Suriname on the mercury concentration in fish. Instituut voor de Opleiding van Leraren, Cursus MOB Biologie, Paramaribo, Suriname. 38p.
Rijnsdorp, A.D., P.I. van Leeuwen, N. Daan and H.J.L. Heesen, 1996. Changes in abundance of demersal fish species in the North Sea between 1906-1909 and 1990-1995. ICES Journal of Marine Science 53: 1054-1062.
Rine, J.M. and R.N. Ginsburg, 1985. Depositional facies of the mudshorefave in Suriname, South America: a mud analogue to sandy, shallow-marine deposits. Journal of Sedimentary Petrology 55 (5): 633-652.
Rogers, S.I., M.J. Kaiser and S. Jennings, 1998. Ecosystem effects of demersal fishing: a European perspective. pp. 68-79. In: Dorsey, E.M. and J. Pedersen (eds.), Effects of fishing gear on the sea floor of New England. Conservation Law Foundation, 62 Summer Street, Boston, MA 02110, USA.
Rossignol, M., 1972. Etude d’un marais de la Guyane Française: le marais Sarcelle. Biologie, écologie des crevettes: Penaeus aztecus subtilis (formes juvéniles). ORSTOM, Cayenne. 23p.
Rutherford, J.C., 1987. A preliminary study of the dispersion of hoki wastes and potential oxygen depletion off the West Coast South Island. Fisheries Research Centre internal report No. 79.
Sainsbury, K.J., 1988. The ecological basis of multispecies fisheries, and management of a demersal fishery in tropical Australia. pp. 349-382. In: Gulland, J.A. (ed.) Fish Population Dynamics. London, John Wiley and Sons.
Sainsbury, K.J., Campbell, R.A., Lindholm, R. and W. Whitelaw, 1997. Experimental management of an Australian multispecies fishery: examining the possibility of trawl-induced habitat modification. pp 107-112. In: Pikitch. E.L., D.D. Huppert and M.P. Sissenwine (eds.) Global Trends: Fisheries Management. American Fisheries Society Symposium 20. Bethesda, MA, USA.
Saxton, F., 1980. Coral loss could deplete fish stocks. Catch 7(8): 12-13.
Smith, W.O.Jr. and D.J. De Master, 1996. Phytoplankton biomass and productivity in the Amazone river plume: correlation with seasonal river discharge. Continental Shelf Research 16 (3): 291-319.
Stevens, J.D., R. Bonfil, N.K. Dulvy and P.A. Walker, 2000. The effects of fishing on sharks, rays, and chimaeras (chondrichthyans) and the implications for marine ecosystems. ICES Journal of Marine Science 57: 479-494.
Teunissen, P.A., 2000. Coastal Management Plan for the North Saramacca Area in Suriname. Project sponsored by Ramsar Convention’s Grants Funds.
Vendeville, P., 1984. La pêcherie de crevettes tropicales de Guyane Française. Le problème des captures accessoires: estimations et implications. Thèse Institut National Polytechnique de Toulouse.
Vendeville, P., 1995. Reconnaissance des fonds de la bande littorale du plateau continental guyanais. Campagne de chalutage «Epaulard 94». Cayenne, ORSTOM.
Villegas L. and A. Dragovich. 1984. The Guianas-Brazil shrimp fishery, its problems and management aspects. pp. 60-70 In: Gulland, J. A. and B. J. Rothschild (eds.), Penaeid Shrimps - Their Biology and Management. Farnham, Great Britain, Fishing News Books.
Walters, C., V. Christensen and D. Pauly, 1997. Structuring dynamic models of exploited ecosystems from trophic mass-balance assessments. Reviews in Fish Biology and Fisheries 7: 139-172.
| 1 Scientific and common English names were
taken from the FAO Field Guide for the northern coast of South America by
Cervigón et al., 1993, also for reasons of consistency with
earlier reports. |
![]()
![]()
![]()
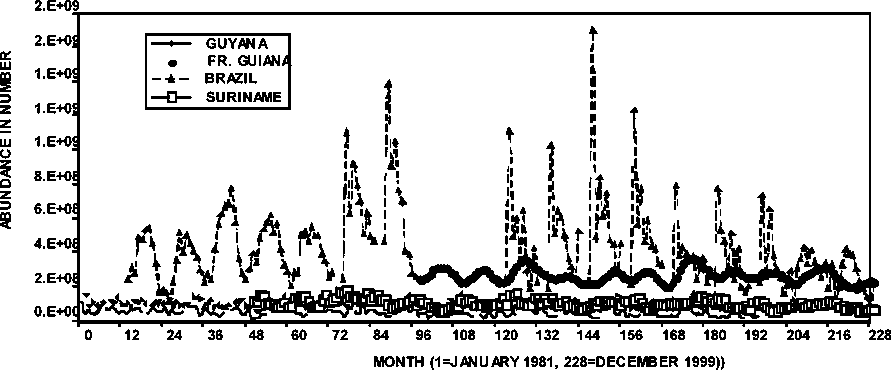{kind=link}
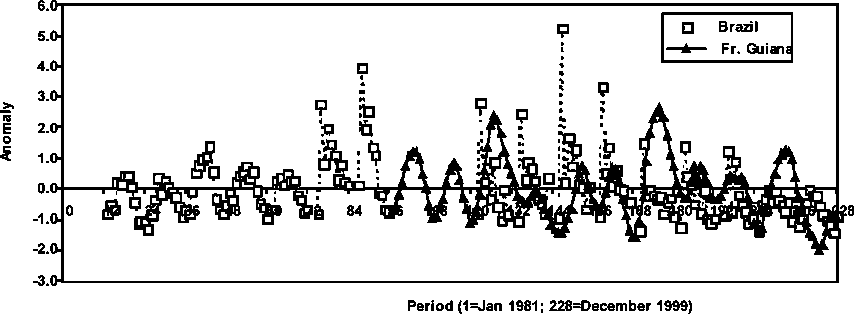{kind=link}
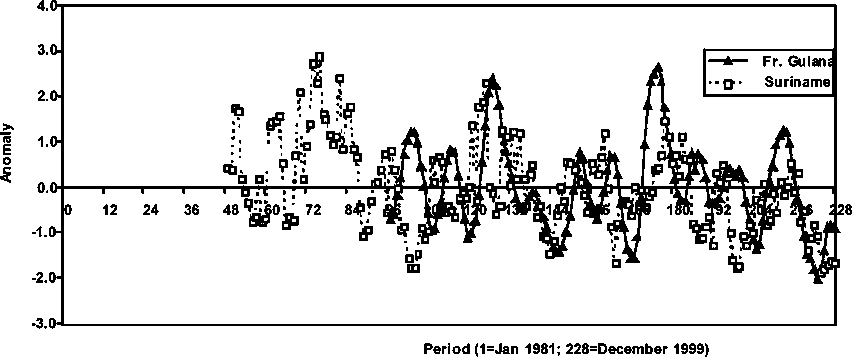{kind=link}
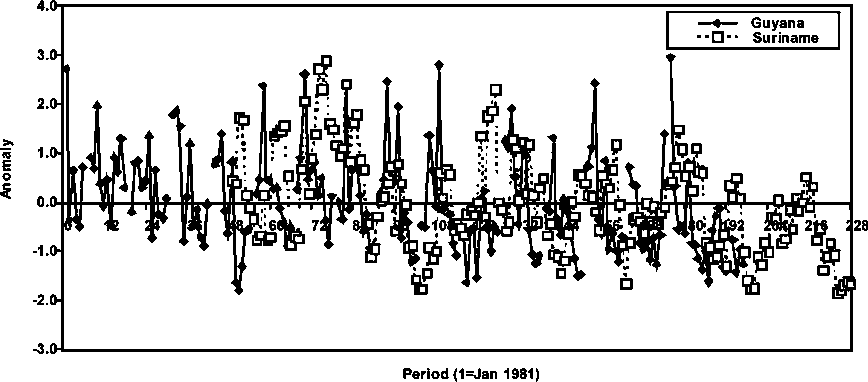{kind=link}
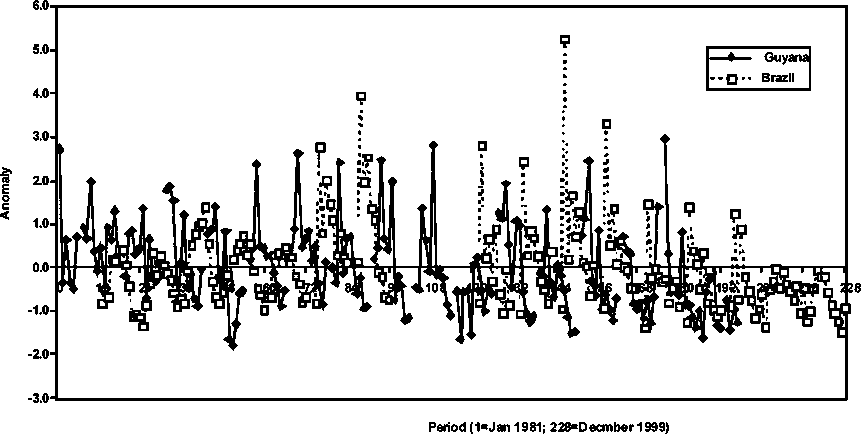{kind=link}
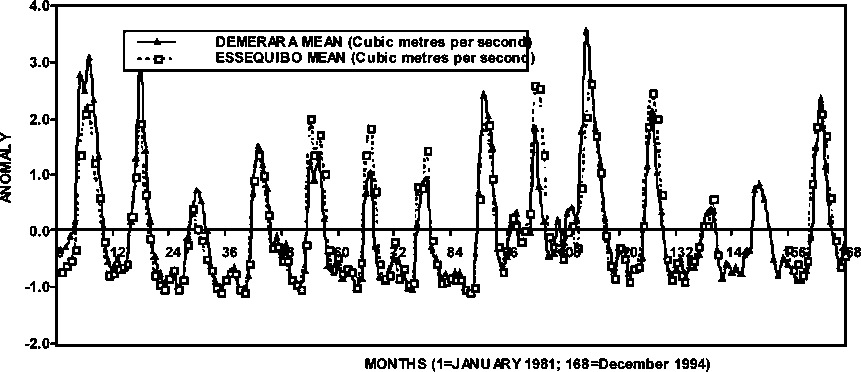{kind=link}
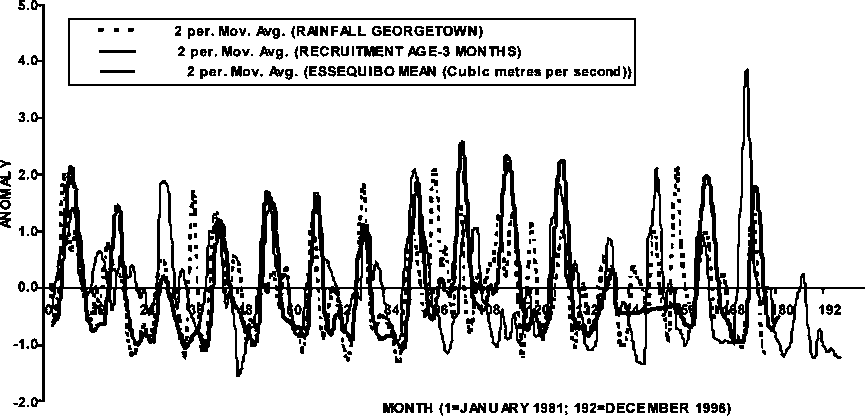{kind=link}
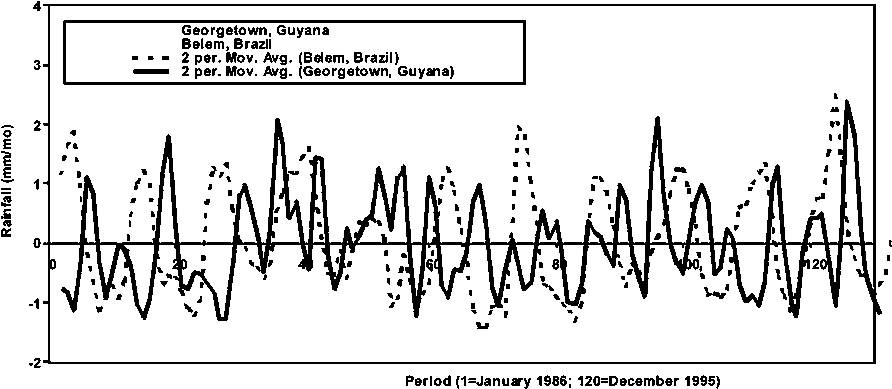{kind=link}
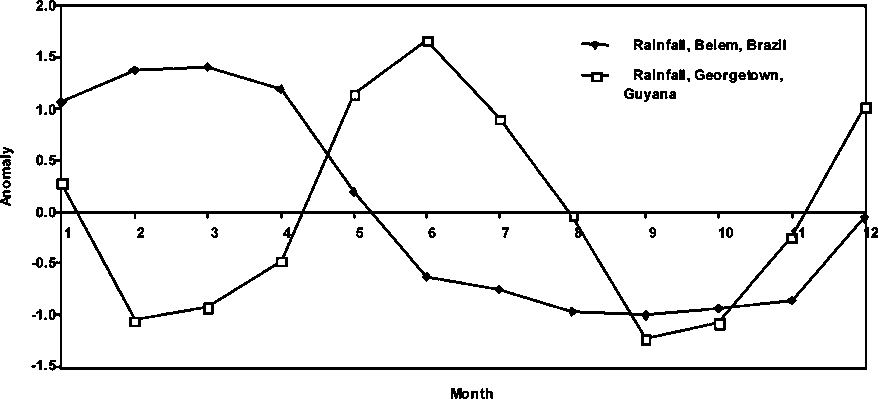{kind=link}
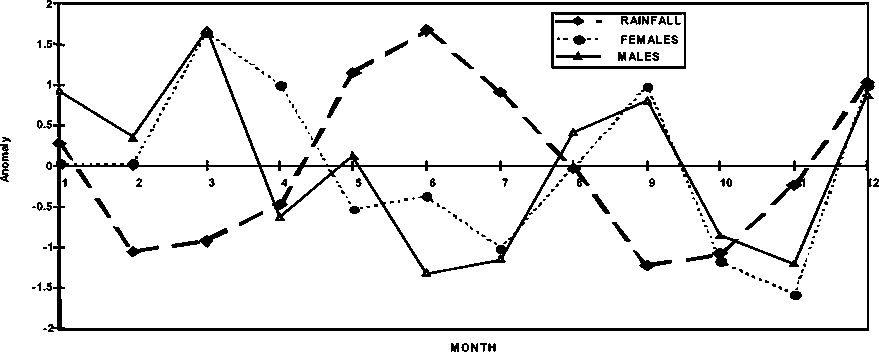{kind=link}
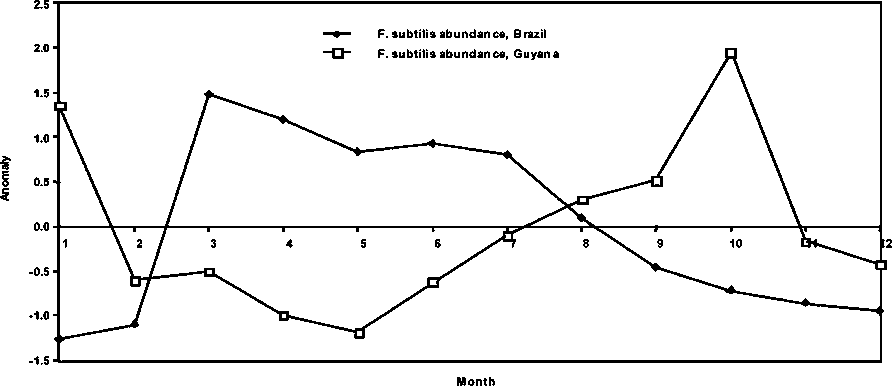{kind=link}
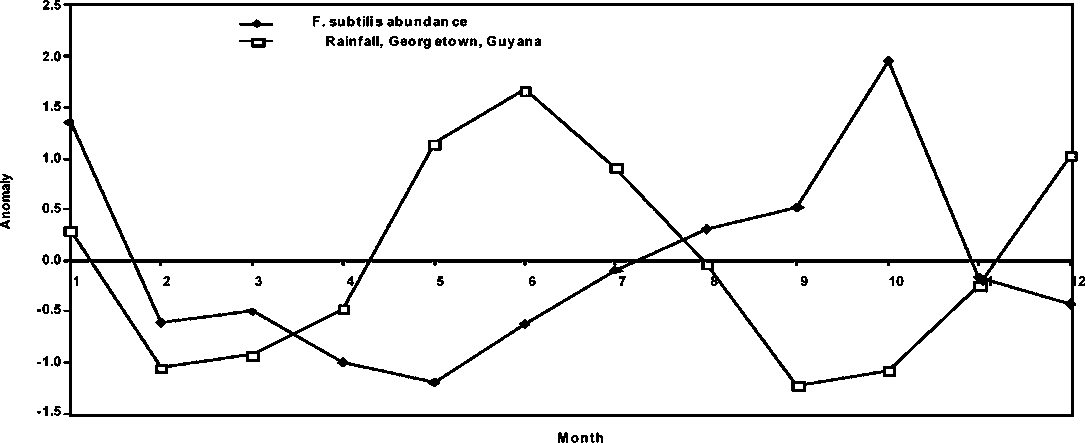{kind=link}
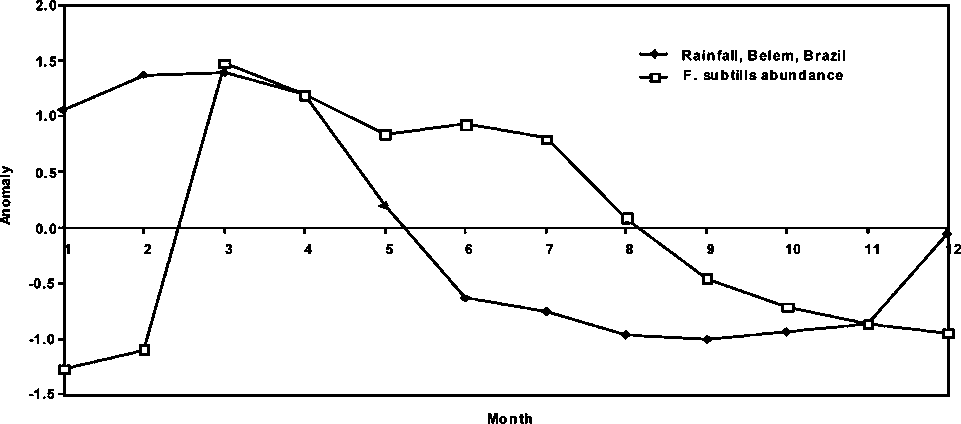{kind=link}
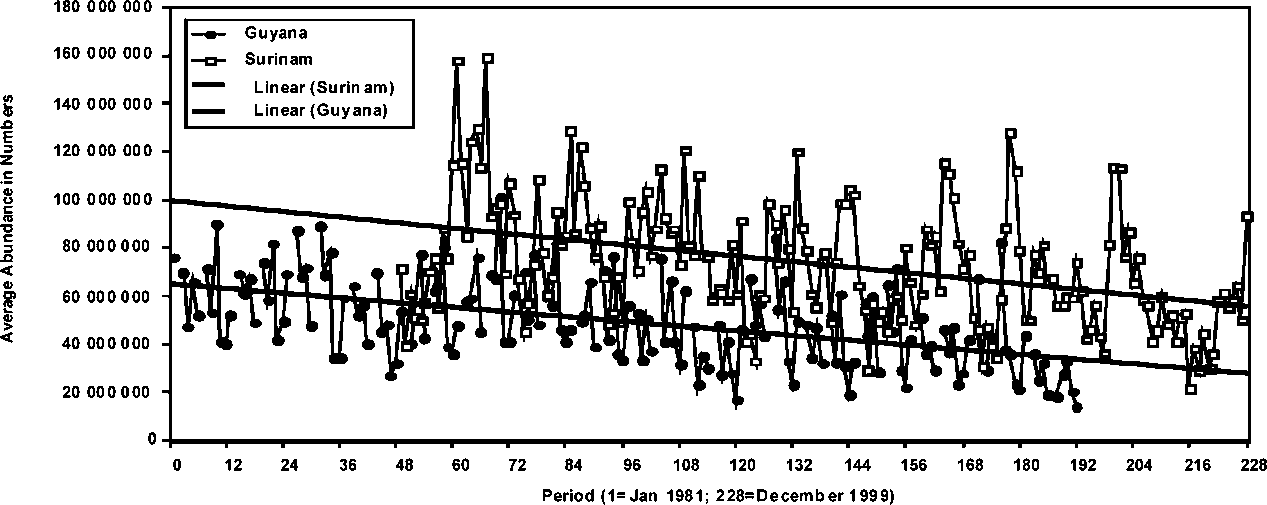{kind=link}
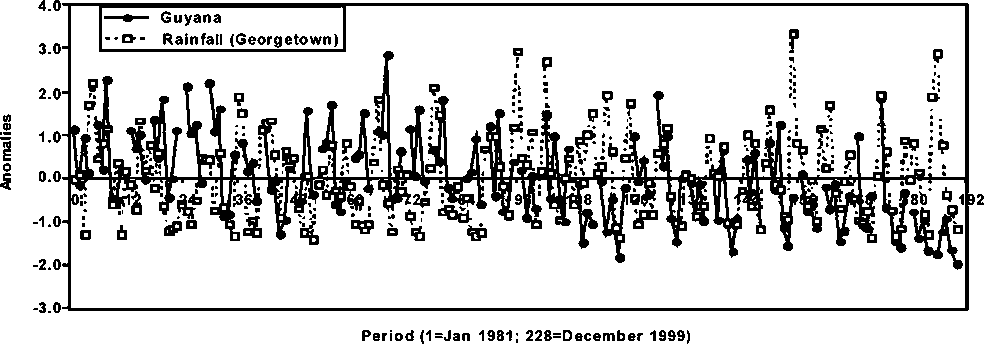{kind=link}
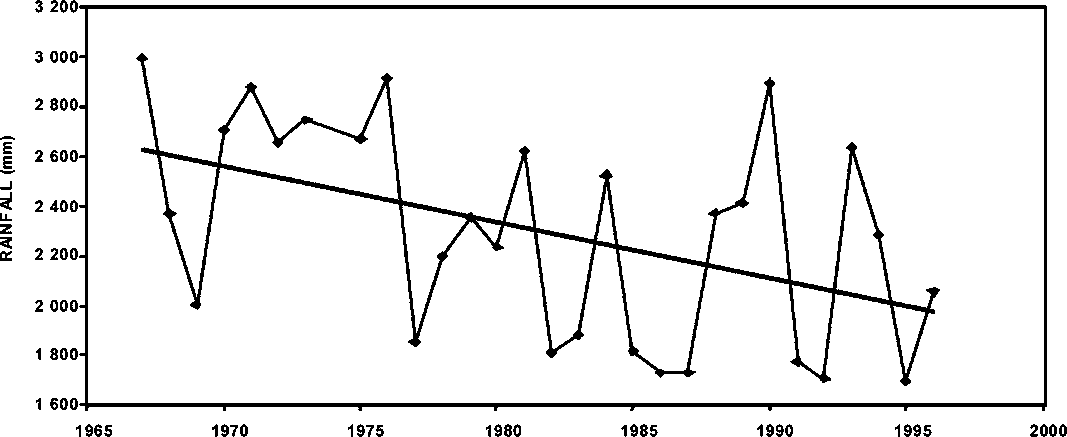{kind=link}
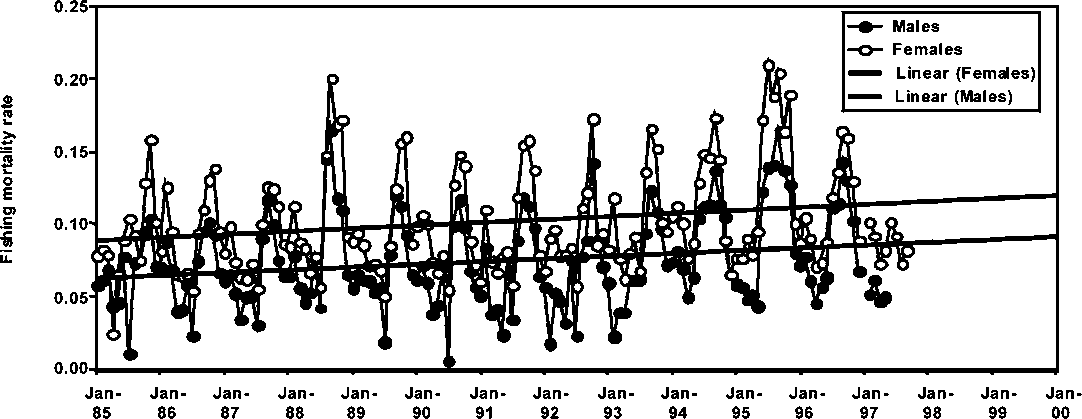{kind=link}
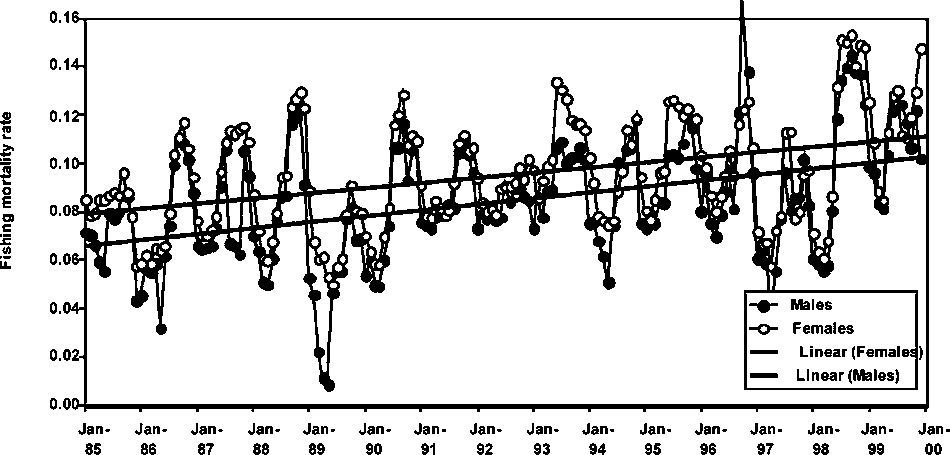{kind=link}
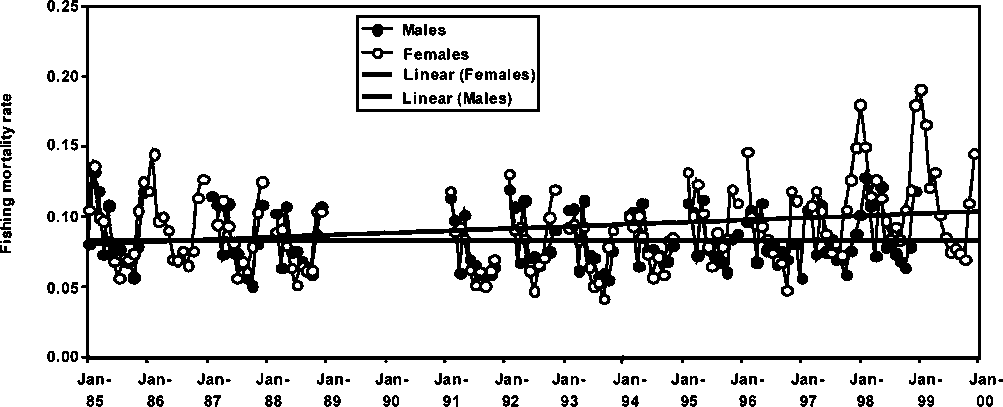{kind=link}
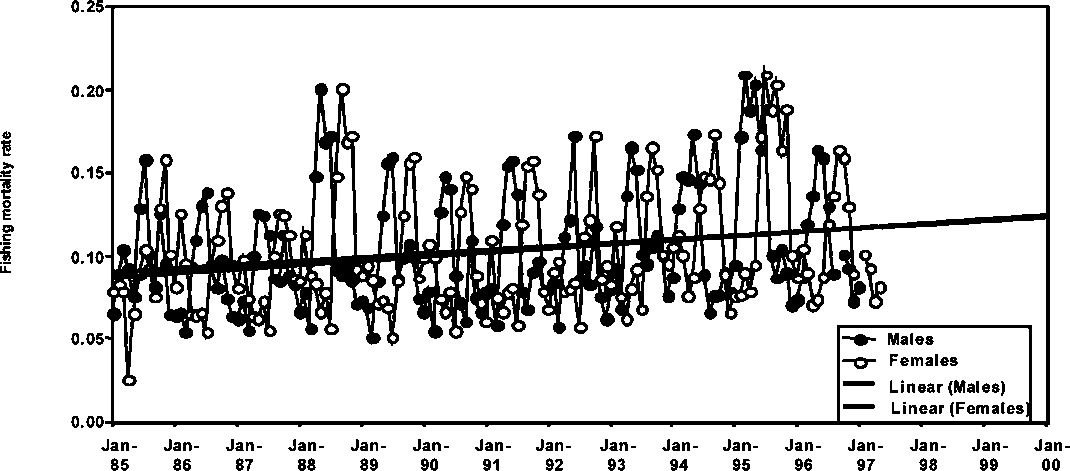{kind=link}
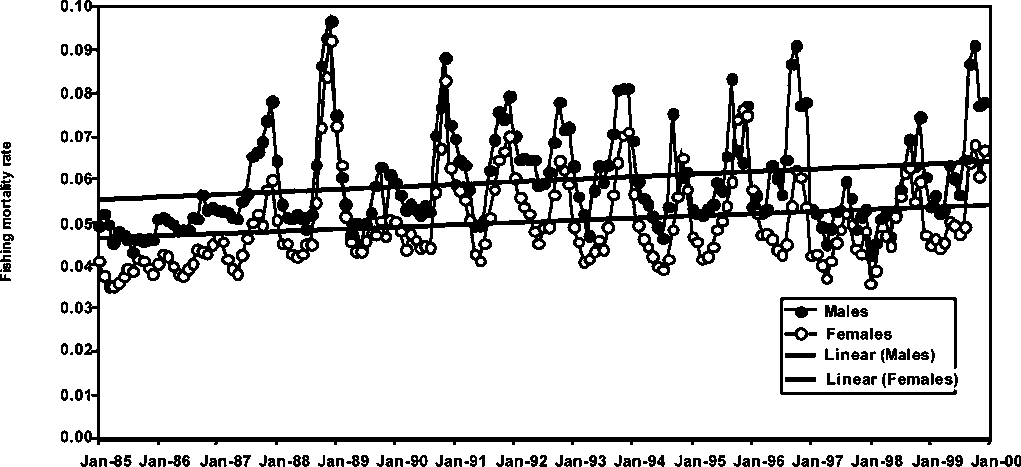{kind=link}