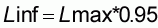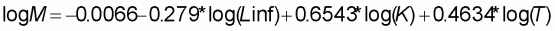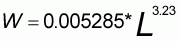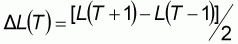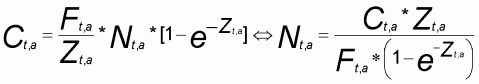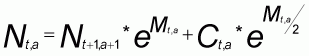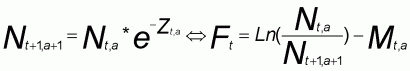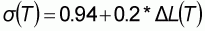![]()
![]()
![]()
Martial Laurans, Didier Gascuel et Mariama Barry
ENSAR, Laboratoire Halieutique, 65 rue d Saint Brieuc, 35042 Rennes, Cedex,
France
Email: [email protected]
Email: [email protected]
Introduction
Le thiof (Epinephelus aeneus) est l'espèce la plus importante en terme de captures parmi les sept espèces de mérou (Brusle, 1985) présente sur les côtes sénégalaises. Elle est principalement ciblée par la pêche artisanale; les techniques de la ligne et du filet maillant sont les plus employées. La pêche industrielle le capture au chalut dans des proportions beaucoup plus faibles. Poisson de base dans le plat national sénégalais (le thiboudien), aujourd'hui la majorité des captures sont exportées vers l'Europe, principalement en France. Le poids économique de cette espèce est élevé, néanmoins, le suivi de son exploitation n'est pas réalisé. Dans un travail complet sur l'espèce (Cury et Worms,1982), une première évaluation par l'approche structurale concluait que le stock au nord du Cap-Vert était sous-exploité. Depuis, aucun travail similaire n'a été mené. Un plus grand nombre de travaux a été réalisé sur sa biologie, en premier lieu sur les migrations entre la Mauritanie et le Sénégal (Champagnat et Domain 1978; Domain, 1980; Cury et Worms, 1982; Cury et Roy, 1988). L'étude de la croissance et de la reproduction ont surtout été réalisées en Méditerranée (Brusle et Brusle, 1975; Ezzat et al., 1981; Bou-Ain et al., 1983a; Bou-Ain et al., 1983b). En effet, le thiof est très présent sur les côtes du Maghreb (notamment en Tunisie) jusqu'en Israël. Le régime alimentaire est essentiellement basé sur des poissons et des crustacées (Cadenat, 1954; Bou-Ain et al., 1983a) et de céphalopodes (Caverivière, com pers).
Comme la plupart des espèces de la famille des serranidés, le thiof est une espèce hermaphrodite protérogyne. La période d'inversion sexuelle est assez bien connue (Brusle, 1975; Bou-Ain et al., 1983a). Elle est variable, mais peut-être définie comme se réalisant autour de la taille de 75 cm. Les jeunes individus (moins de 30 cm) se trouvent surtout en zone côtière, notamment en zone estuarienne (estuaire du Sine Saloum et du fleuve Sénégal au Sénégal). Ensuite, ils se trouvent à des profondeurs plus importantes (entre 30 et 100 mètres, Caverivière, 1994) à la fois dans des zones rocheuses (moins qu'Epinephelus guaza) et surtout dans des milieux avec de fortes proportions de zones sableuses (Champagnat, 1978; Brusle, 1985; Bou-Ain et al., 1983a).
Dans le présent papier, la courbe de croissance du thiof est revue, la limite du stock sur lequel nous travaillons est discutée, ensuite les résultats de l'évaluation de stock par l'approche structurale sont présentés.
Matériel et Méthode
- Données
Les données de capture, d'effort et de fréquence de taille proviennent du CRODT (Centre de Recherche Océanographiques de Dakar-Thiaroye). Pour la pêche artisanale, l'ensemble de ces données est issu d'une extrapolation à partir d'un plan d'échantillonnage des sorties journalières (Ferraris et al., 1994). Ce travail a commencé en 1974 dans les principaux ports de pêche de l'époque. Avec le développement de la pêche, d'autres lieux de débarquements ont pris de l'importance et ont été suivis (figure 1). A partir de 1981, toute la côte sénégalaise est suivie exhaustivement. Le littoral sénégalais est divisé en grande zone (figure 1). Aussi, les données de la pêche artisanale sont considérées comme complète à partir de cette année là. Concernant les structures de taille, les données ne sont pour le moment disponibles qu'à partir de 1989.
Pour la pêche industrielle, les données des navires étrangers sont collectées par des enquêteurs embarqués. Pour les navires sénégalais, à chaque débarquement un enquêteur est présent pour récupérer l'information (capture, temps de pêche, lieu de pêche). Aucune donnée de fréquence de taille n'est disponible pour le thiof. Les patrons pêcheurs ou les mareyeurs ne veulent pas que ce poisson soit manipulé (sortie de la glace) afin de ne pas altérer sa qualité.
Figure 1: Littoral sénégalais avec les quatre principales zones de débarquements.
- Stock
Cury et Worms (1982) ont travaillé sur le stock de thiof et ont formulé une hypothèse quant à ses limites. En utilisant les séries de données disponibles aujourd'hui, on reprend des études similaires à celles réalisées par ces auteurs. La comparaison des cpue et de leur évolution pour différentes zones est utilisée. Ces CPUE sont estimés pour le métier de la ligne moteur. En effet, ce métier cible en partie le thiof; il a la particularité d'avoir un rayon d'action limité et de faire des sorties à la journée. De ce fait, les cpue d'une zone (obtenues à partir des données des ports de cette zone) correspondent bien à des captures faites dans cette zone. Ce n'est pas le cas des pirogues glacières (sorties de plusieurs jours) qui ont la particularité de se déplacer sur de longue distance et pêcher dans plusieurs zones. Afin de montrer la saisonnalité des captures, celles-ci sont présentées par zone, année et mois. Avec les données de fréquence de taille, on analyse l'évolution du pourcentage de poissons d'une taille supérieure à 60 cm dans les captures. Le calcul est une moyenne mensuelle sur toutes les années de la série. Ce suivi est réalisé pour le métier de la ligne moteur et pour l'ensemble de la pêcherie. D'autre part, les CPUE et les captures mensuelles du métier ligne moteur à Kayar sont également calculées sur la période 1974-1994. En effet, la fosse de Kayar est souvent citée comme un obstacle à la migration de certaines espèces. Enfin, le résultat d'un marquage réalisé en 1977 est également utilisé ici.
- Courbe de croissance
A partir des échantillons de fréquence de taille de la pêche artisanale, on estime la courbe de croissance du thiof. Pour cela, les données de fréquence de taille sont groupées par trimestre de 1989 à 1996. On utilise la méthode de Shepherd (Shepherd, 1987) sous la dernière version du logiciel FISAT de la FAO. Ce logiciel permet d'obtenir un «bananoïde de confiance», décrivant les corrélations entre les paramètres Linf (longueur asymptotique) et le paramètre K (vitesse de croissance) de l'équation de Von bertalanffy qui donnent le meilleur ajustement. Afin de choisir un couple de valeur, on décide ici de fixer le paramètre Linf, comme suit (Gascuel, com pers):
|
|
(1) |
Lmax est la plus grande valeur jamais mesurée pour un individu de l'espèce. Linf est une taille moyenne asymptotique et il est logique de penser qu'elle peut-être légèrement inférieure à Lmax. Il est important de souligner que l'âge utilisé dans l'équation de la courbe de croissance est un âge conventionnel, on pose ainsi qu'au premier mois de janvier de leur vie les poissons ont 1 an.
- Mortalité naturelle (M)
Concernant la mortalité naturelle du thiof, peu de connaissances existent. Une estimation est effectuée au Sénégal (Cury et Worms, 1982) et au Congo (Fontana, 1979). A partir des paramètres de la courbe de croissance et de la connaissance de la biologie, nous appliquons la formule de Pauly (Pauly, 1980) qui permet d'estimer la valeur de M (2).
|
|
(2) |
Afin d'obtenir une valeur moyenne de la température (T) à laquelle vit le thiof, on utilise des données provenant de campagnes scientifiques où la température de fond a été relevée pour tous les coups de chaluts réalisés (campagnes du Laurent Amaro de 1970 à 1974). T est calculé comme étant la moyenne de la température de fond en prenant en compte tous les traits de chaluts où du thiof était présent. Le poids de chaque trait est pondéré par le nombre de thiof capturé par ce même trait.
- Capture en nombre d'individus
Pour l'analyse structurale, les captures (en tonnes) doivent être converties en nombre d'individus par classe de longueur. Les classes de longueur sont comptées par centimètre. A partir de l'échantillonnage de la structure de taille des captures, l'utilisation de la relation taille poids (Franqueville et Fréon, 1976) permet d'obtenir le nombre d'individus par classe.
|
|
(3) |
Ce travail est réalisé en pêche artisanale pour chaque type de pêche et chaque année afin de prendre en compte les différents diagrammes d'exploitation. Aussi, pour la pêche artisanale, le nombre d'individus capturés par classe de longueur est la somme des données des différents types de pêche. Pour la pêche industrielle, ne disposant pas de fréquence de taille, nous faisons l'hypothèse que le diagramme est le même que celui de l'ensemble de la pêche artisanale. Cette démarche constitue une approximation sans doute sans grandes conséquences sur les résultats finaux compte tenu des captures de la pêche industrielle qui sont faibles face à la pêche artisanale (moyenne de 17 pour cent sur la période 1989-1999).
- Conversion taille/âge
L'évaluation de stocks par l'approche structurale peut-être réalisée en taille ou en âge. Cette seconde approche est de loin préférable dans la mesure où elle permet de s'affranchir de l'hypothèse d'équilibre liée à l'analyse en taille. C'est donc cette analyse en âge qui est utilisée ici. Pour cela, une décomposition polymodale des captures est effectuée par la méthode du maximum de vraisemblance (Hasselblad, 1966) à l'aide du logiciel NORMSEP (Boer, 1986). Cet ajustement est réalisé sous contrainte, c'est ça dire en fixant, pour chaque âge, la longueur moyenne et l'écart type de taille (Gascuel, 1994). Lors d'un premier traitement, les tailles moyennes sont fixées conformément au modèle de croissance; le logiciel trouve par itération un ajustement, en estimant pour chaque mode un écartype de taille. Pour chaque année de notre série, on obtient une valeur de l'écartype par groupe d'âge. Ces données sont utilisées pour ajuster un modèle des écartypes (Gascuel, 1994); l'ajustement est réalisé par les moindres carrés.
|
|
(4) |
avec
L(T) est la longueur donnée par la courbe de croissance à un âge T.
Lors d'un deuxième traitement des données, on utilise le modèle des écartypes pour fixer à chaque groupe d'âge l'écartype correspondant. Ainsi, les mêmes valeurs sont utilisées pour toutes les années. Ce travail permet d'estimer un tableau des captures par groupe d'âge.
- Analyse des cohortes
L'analyse des cohortes (ou VPA) est réalisée sous le logiciel Excel selon une procédure de calcul en mode inverse. On initialise la VPA en fixant dans un premier temps la mortalité par pêche (F) du dernier groupe d'âge pour chaque année et la mortalité par pêche de chaque groupe d'âge pour la dernière année de pêche. A partir de l'équation (5), dérivant de l'équation de survie, les effectifs terminaux corresponds sont estimés. L'équation de Pope (6) permet ensuite de calculer l'ensemble des effectifs. Enfin, le calcul des mortalités par pêche se fait grâce à l'équation de survie (7).
|
|
(5) |
|
|
(6) |
|
|
(7) |
Les mortalités F terminales sont estimées par une procédure de stabilisation, en s'appuyant sur 2 hypothèses:
la mortalité F de dernier âge est égale à la moyenne des âges précédents. (âges x à y)
la mortalité F de dernière année est égale à la moyenne des 3 années précédentes multipliée par un coefficient «a» proche de 1.
Différents scénarios de l'évolution et donc de F de la dernière année sont ainsi testés en fonction du coefficient a (entre 1 et 1.2). Les résultats directs de l'analyse des cohortes sont les valeurs du recrutement, de la biomasse totale et féconde pour chaque année. La série de biomasse obtenue est comparée à celle calculée par le modèle global (Laurans et al., 2001) et à des séries d'indices d'abondance calculés dans différents travaux.
La mortalité par pêche de la dernière année est ensuite utilisée pour les diagnostics de rendement, de biomasse totale et féconde par recrue. La sensibilité de ces diagnostics à la valeur du paramètre a est analysée.
Résultats
- Les captures
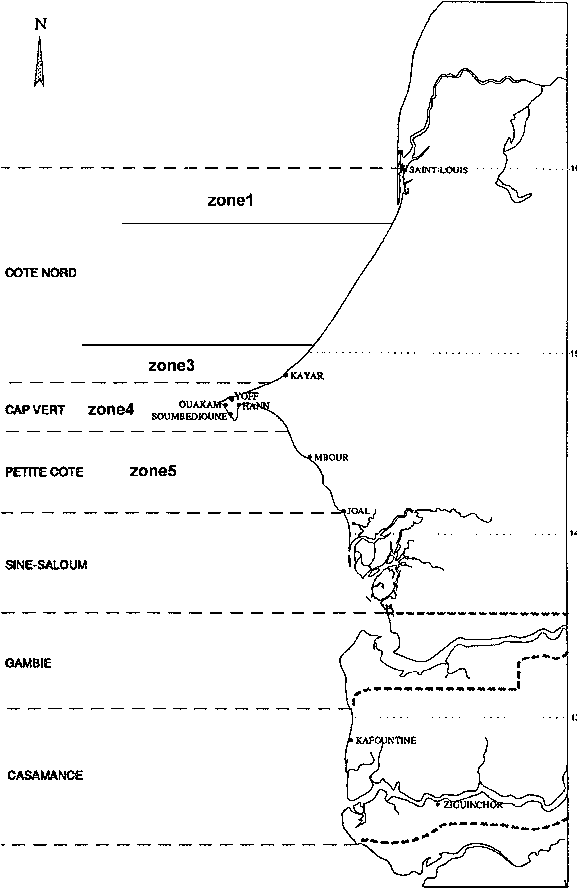
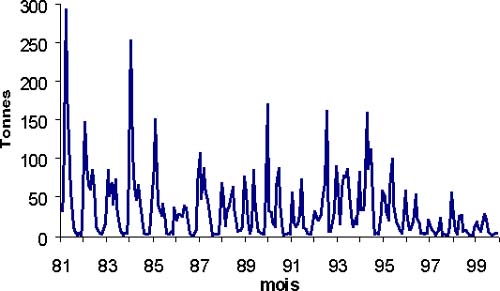
Depuis l'année 1981 (figure 2), la tendance principale des captures est la baisse des débarquements depuis 1986. En 1998, les plus faibles débarquements jamais atteints sont notés. La période de 1984 à 1987 est caractérisée par des captures très fortes. L'effort de pêche et les CPUE sur cette période sont pour certains types de pêche plus élevés. Ferraris (1994) précise que cette augmentation est du aux captures des zones 4 et 5 (figure 1).
Il faut noter que la part de la pêche industrielle dans les débarquements est faible (au maximum 27 pour cent en 1990). Cela est du en grande partie au biotope que fréquente le thiof; ce sont souvent des zones peu chalutables. C'est pourquoi cette espèce se capture principalement grâce à des engins munis d'hameçons.
Figure 2: Captures de thiof au Sénégal, part relative de la pêche artisanale (PA) et industrielle (PI).
- Courbe de croissance
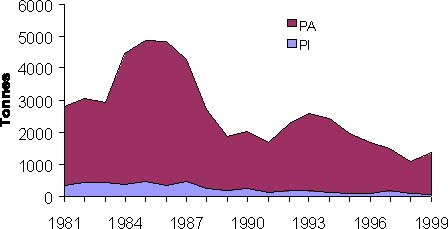
Les données de fréquence de taille sont présentées en annexe. Avec un Lmax de 115 cm (Cury et Worms, 1982; Bou-Ain et 1983a), la relation (1) implique un Linf de 109.25 cm. En utilisant la méthode de Shepherd, on obtient un K de 0,12 an-1. Les valeurs de ces paramètres sont utilisées pour calculer to, soit une valeur de -0.46 en prenant comme âge conventionnel 1 pour le premier mois de janvier que vit l'animal, avec à cet âge là une longueur de 6 cm (figure 3).
Figure 3: Courbe de croissance du thiof, estimée par la méthode de Shepherd. Cette figure ne montre qu'une partie des données trimestrielles utilisées.
- Mortalité naturelle
A partir des données des campagnes du Laurent Amaro, la température moyenne pondérée à laquelle se trouve les thiofs est d'environ 17,5 °C. Cela confirme bien le fait que le thiof est une espèce d'affinité saharienne (Domain,1980). L'utilisation de cette valeur et des paramètres de la courbe de croissance permet d'obtenir une estimation de M. Soit, d'après (4): 0,25 an-1. Cette valeur est utilisée dans la VPA. Elle est proche de celle utilisée par Cury et Worms (1982) pour les jeunes âges.
- Limites du stock
Champagnat et Domain (1978) font l'hypothèse que le stock de thiof est affecté d'une vaste migration nord-sud le long des côtes sénégalaises. En saison froide, les thiofs migreraient vers le sud, inversement en saison chaude. Cury et Worms (1982) font l'hypothèse de l'existence de deux stocks, l'un au nord du Cap-Vert qui réalise la migration jusqu'à la fosse de Kayar et l'un au sud étant sédentaire. Pour ces auteurs, la fosse de Kayar est l'équivalent d'une barrière entre ces deux stocks, avec possibilité d'échange. La migration est quant à elle, fortement liée à la présence d'un upwelling qui s'installe de novembre à juin sous l'effet des alizés (Champagnat et Domain, 1978; Domain, 1980; Cury et Worms, 1982; Cury et Roy, 1988).
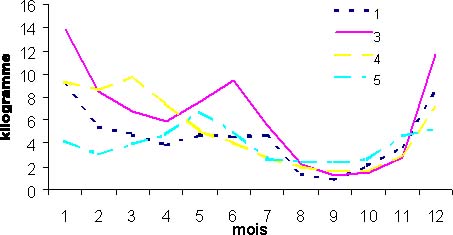
Le suivi des cpue de différents types de pêche artisanale par zone géographique montre des variations saisonnières similaires. C'est le cas pour le métier pirogue ligne moteur de la pêche artisanale (figure 4). Quelles que soient les zones, on observe la même diminution des cpue durant la saison chaude, période d'août à octobre. Contrairement à ce qu'avance Cury et Worms (1982), on peut donc dire que le thiof en saison froide se trouve à la côte au sud de la presqu'île du Cap-Vert. En saison froide, le pic de CPUE par zone semble être décalé dans le temps. Il semble progresser vers le sud au fur et à mesure que la saison froide s'installe, Kayar au mois de janvier, ensuite la presqu'île du Cap-Vert en mars, puis la grande côte en mai et finalement un deuxième pic en juin au niveau de Kayar. Champagnat et Domain (1978) indiquent un rythme de migration qui explique ces pics de CPUE décalés.
Figure 4: CPUE mensuelles par zone moyennées sur la période 1981-1994 pour le métier ligne moteur. (1:zone de Saint Louis, 3:zone de la grande côte, 4:zone du Cap-Vert, 5:zone de la petite côte).
Quelque soit la zone que l'on considère, il existe une forte saisonnalité dans les captures de thiof (figure 5). Ainsi, les captures de thiof sont les plus basses en saison chaude. La migration que l'on connaît se manifeste de la même façon pour toutes les zones.
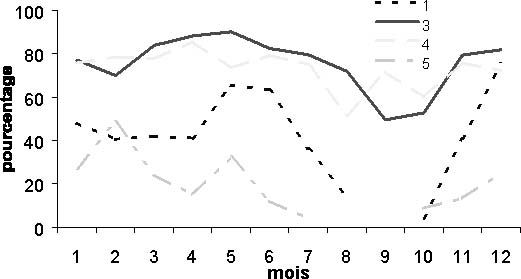
Pour toutes les zones, la proportion d'individus d'une taille supérieure à 60 cm diminue en saison chaude (figures 6a et 6b). Ainsi, pour le métier ligne moteur en zone 5 (petite côte) et 1 (St Louis), aucun thiof supérieur à 60 cm n'est capturé en saison chaude (figure 6b). Ces individus sont donc peu présents sur ces zones de pêche. Pour ce même métier, le poids moyen des individus pêchés est toujours inférieur en saison chaude (tableau 1). Aussi, le phénomène de disparition des gros thiofs (supérieur à 60 cm) en saison chaude ne semble pas être atténué au sud de la presqu'île de Cap-Vert comme l'avance Cury et Worms (1982).
Figure 5: Captures mensuelles par zone et par mois sur la période 1981-1999. A: St Louis
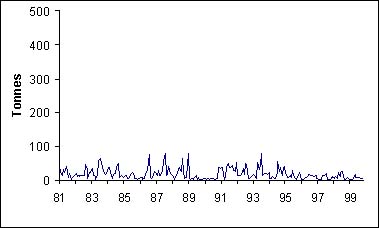
Figure 5: Captures mensuelles par zone et par mois sur la période 1981-1999. B: Kayar
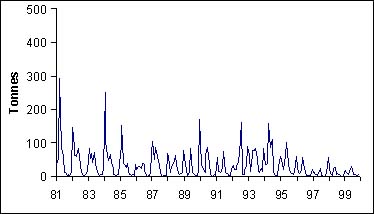
Figure 5: Captures mensuelles par zone et par mois sur la période 1981-1999. C: presqu'île Cap-Vert
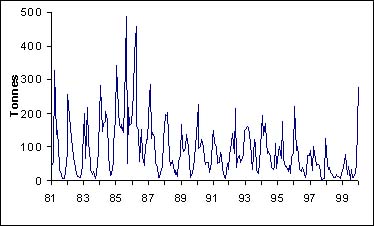
Figure 5: Captures mensuelles par zone et par mois sur la période 1981-1999. D: Thiès Sud.
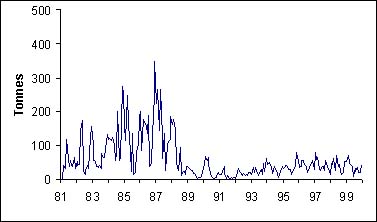
Figure 6: Pourcentage mensuel de thiofs d'une taille supérieure à 60 cm sur la période 1989-1999. a: tout métier confondu;
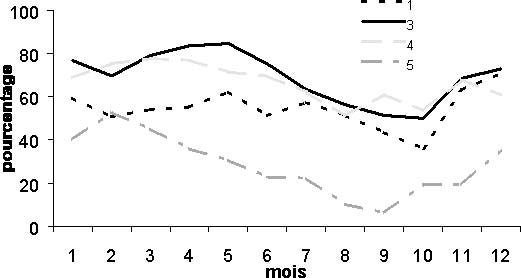
Figure 6: Pourcentage mensuel de thiofs d'une taille supérieure à 60 cm sur la période 1989-1999. b:pour le métier ligne moteur.
Les CPUE au niveau de Kayar (figure 7a) montre une forte saisonnalité. Elles augmentent en début de saison froide (novembre à janvier) puis diminuent avant de réaugmenter en fin de saison froide, cette tendance générale est plus ou moins marquée selon les années. La saison chaude est marquée par les CPUE les plus faibles de l'année. Durant la saison froide, contrairement à ce qu'avance Cury et Worms (1982), il est difficile de dire que la population de thiof se stabilise devant kayar au vu de la diminution de CPUE que l'on observe vers mars avril. Les débarquements de thiof à Kayar montrent cette même saisonnalité (figure 6b). Ferraris (1994) indique que les deux pics de débarquements correspondent aux deux passages d'individus migrateurs, le premier étant la migration vers le sud, le second la remontée vers la Mauritanie. La recapture d'un poisson au niveau du cap Roxo (Champagnat et Domain, 1978) qui a été marqué deux mois plus tôt en décembre à Kayar confirme cette migration vers le sud en saison froide et la possibilité de passage de la fosse de Kayar.
Figure 7: CPUE mensuelle de thiof à Kayar par le métier ligne moteur.
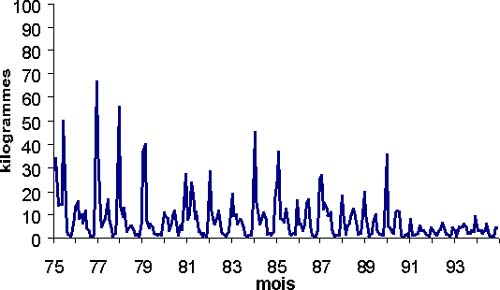
Figure 7: Capture mensuelle de thiof à Kayar par le métier ligne moteur.
Dans le présent travail, nous ne considérons pas deux stocks de thiof comme Cury et Worms, mais un seul stock qui opère une migration le long des côtes sénégalaises comme l'on fait Champagnat et Domain (1978). La limite nord serait proche du Cap Blanc en Mauritanie, quant à la limite sud, elle serait autour du Cap Roxo. Sous cette limite, très peu de thiofs sont capturés (Domain, com pers). Les tendances dans les pêcheries de chaque zone sont très proches et ne permettent pas de dire que les poissons capturés au sud de la presqu'île du Cap-Vert appartiennent à un stock différent de ceux capturés au nord.
- Capture par âge
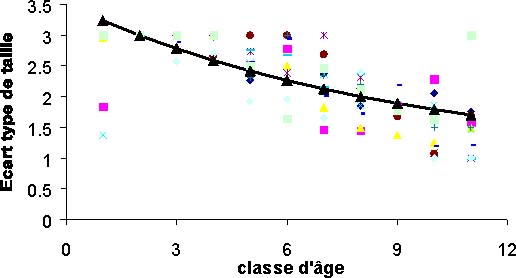
Les captures numériques par classe de longueur sont estimées pour chacune des années. A partir de ces données, une première décomposition polymodale permet d'obtenir des écartypes de taille par groupe d'âge (figure 8). Ces valeurs montrent clairement une diminution de l'écartype avec l'âge. Le modèle d'écartype ajusté est linéaire avec une pente positive.
|
|
(8) |
Figure 8: Modèle des écartypes et données sur lesquelles il a été ajusté.
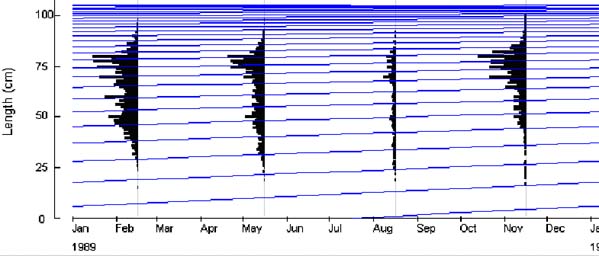
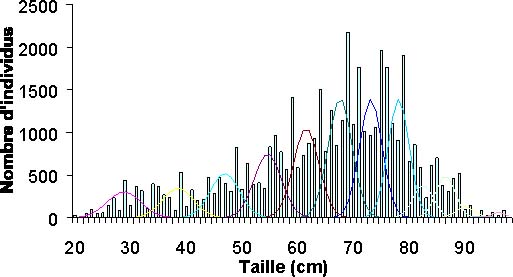
Un deuxième ajustement sous Normsep en tenant compte du précédent modèle, donne les décompositions polymodales des captures (figure 9) pour chacune des années, de 1989 à 1999. La comparaison des histogrammes des différentes années et les données de captures par groupe d'âge permet de voir comment évolue la pression de pêche. Une augmentation des captures dans les groupes d'âge jeune, notamment 1 et 2 (tableau 2), est visible.
Figure 9: Décomposition polymodale des captures e l'année 1991. Chaque mode correspond à un groupe d'âge.
- Analyse des cohortes
Figure 10: Résultats de l'analyse des cohortes pour le stock de thiof du Sénégal. Pour chaque graphique, les trois courbes présentées correspondent à des valeurs différentes du paramètre a. A: mortalité naturelle moyenne
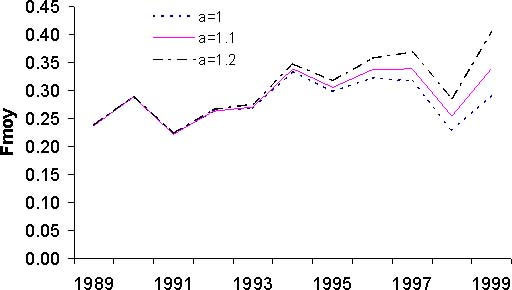
Figure 10: Résultats de l'analyse des cohortes pour le stock de thiof du Sénégal. Pour chaque graphique, les trois courbes présentées correspondent à des valeurs différentes du paramètre a. B: recrutement
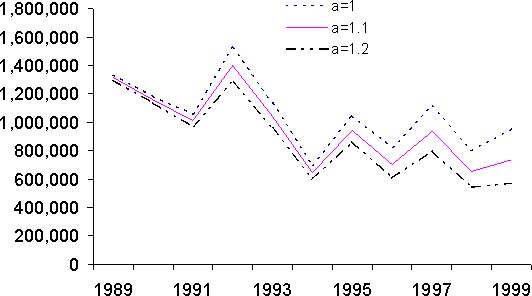
Figure 10: Résultats de l'analyse des cohortes pour le stock de thiof du Sénégal. Pour chaque graphique, les trois courbes présentées correspondent à des valeurs différentes du paramètre a. C: biomasse totale
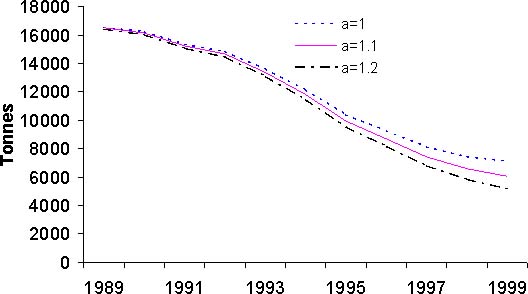
Figure 10: Résultats de l'analyse des cohortes pour le stock de thiof du Sénégal. Pour chaque graphique, les trois courbes présentées correspondent à des valeurs différentes du paramètre a. D: biomasse féconde
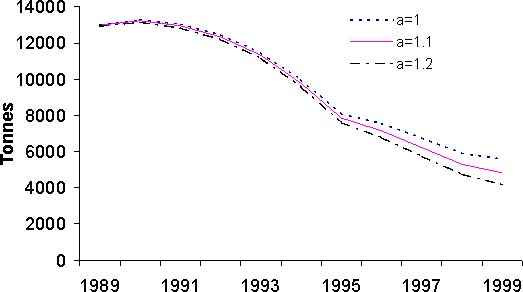
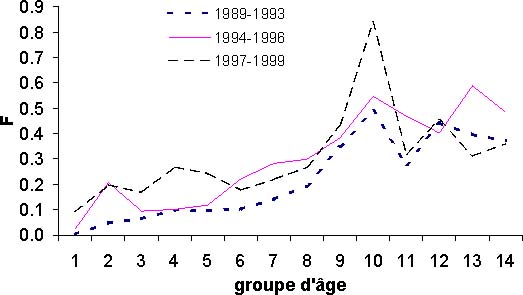
Les résultats issus de l'analyse des cohortes permettent de dresser un premier bilan de l'état du stock de thiof. La propriété de convergence de la VPA implique que les valeurs de mortalité par pêche sont très proches pour les années anciennes et cela quels que soient les paramètres d'initialisation de la VPA (figure 10A). Pour les années 1989 à 1992, les mortalités par pêche fluctuent mais une certaine stabilité est notée. Ensuite, la mortalité par pêche augmente fortement jusqu'en 1997. Suit une baisse importante puis remontée en 1999. Globalement sur la période 1993 à 1999, la tendance est à la hausse. Selon la valeur du paramètre a, on n'observe que peu de différences en 1999 et 1998. Pour le recrutement, selon la valeur du paramètre a, le diagnostic est assez semblable avec une tendance générale à la baisse (figure 10B). Avec une valeur de 1,2, cette tendance à la baisse est de l'ordre de 50 pour cent sur l'ensemble de la période. Il n'en est pas de même avec une valeur de 1; là le recrutement baisse de 1989 à 1994, mais montre une certaine stabilité depuis, voir même une légère hausse. Pour la biomasse féconde (BF) et la biomasse totale (B) du stock (figures 10C et 10D), les tendances sur la période sont identiques et les diagnostics ne varient pas selon la valeur du paramètre a. Il y a une forte diminution de ces deux indices sur la période d'étude, en moyenne par 2. Cette valeur nous indique que l'exploitation de ce stock est intense. Si l'on s'intéresse à l'évolution du diagramme d'exploitation (figure 11), on s'aperçoit que celui-ci a évolué au cours des années. En effet, la mortalité par pêche des jeunes groupes d'âges et des plus vieux a fortement augmenté. Cette modification du diagramme d'exploitation explique en partie la baisse de biomasse que l'on observe.
Figure 11: Diagramme d'exploitation moyen de la pêcherie sur différente période.
- Diagnostic
Figure 12: En fonction de la valeur du paramètre a, courbes de rendements par recrue
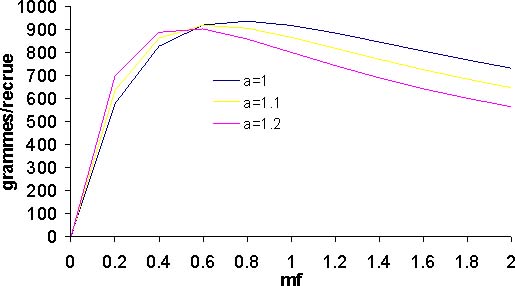
Figure 12: En fonction de la valeur du paramètre a, courbes de biomasse féconde par recrue
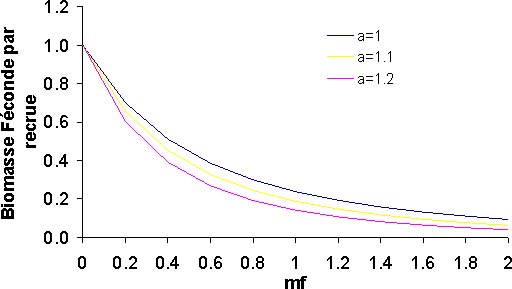
Le diagnostic de rendement par recrue est le même quelle que soit la valeur du paramètre a; le thiof est en état de surexploitation (figure 12a). En effet, l'effort actuel est supérieur à celui de maximisation de 1,3 à 1,7. La situation la plus préoccupante correspond à une valeur de a égale à 1,2. Dans le même temps, le rapport biomasse féconde sur recrutement est dans les trois cas supérieurs au seuil empirique de 10 pour cent (figure 12b), souvent considéré comme étant le seuil d'exploitation de recrutement. Néanmoins la situation est préoccupante, avec une valeur de a égale à 1,2, on se trouve avec un rapport de 14 pour cent dans la situation actuelle. Pour les autres valeurs de a, le ratio s'établi à 19 et 24 pour cent. La situation réelle, doit se trouver entre les valeurs extrêmes pour le rapport Y/R et BF/R. La situation du stock de thiof est préoccupante.
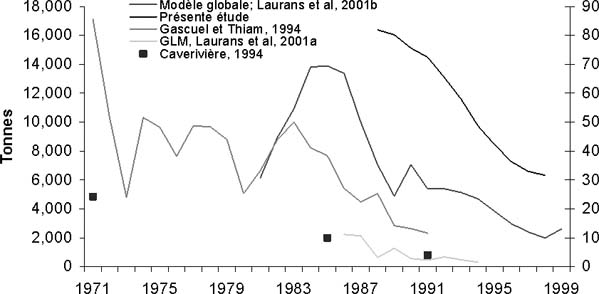
Figure 13: Evolution de l'abondance du thiof au Sénégal, d'après les différents indices disponibles (indices relatifs, en valeur arbitraire pour Gascuel et Thiam 1994, pour Laurans et al., 2001a et pour Caverivière 1994; biomasse du stock en tonne pour Laurans et al. 2001b (Modèle globale) et pour la présente étude).
Discussion
Cette tendance que l'on observe par approche structurale avait déjà été observée par approche globale (figure 13). Ces deux approches concluent sur les mêmes évolutions concernant la situation du stock (tableau 3). Néanmoins, le diagnostic semble plus pessimiste par l'approche globale.
Aucun travail similaire (approche structurale) n'a été réalisé depuis 1982. La présente analyse permet ainsi de faire un diagnostic sur la situation actuelle du stock de thiof. Ce diagnostic est à l'image des données que nous avons au Sénégal. Les données mauritaniennes de statistiques de pêche n'étant pas utilisables, les captures de ce pays ne sont pas prises en compte dans les captures totales. Néanmoins, la connaissance de la pêcherie mauritanienne montre que les modes de pêche sont proches de ceux qu'utilisent les Sénégalais. Aussi, le diagramme d'exploitation est supposé être très proche. Ainsi, si nous faisons une erreur dans notre évaluation elle est d'ordre quantitative et non qualitative, et par-là même le diagnostic auquel nous arrivons resterait le même, à savoir la surexploitation pour le stock de thiof aujourd'hui. A deux reprises, l'évaluation du stock de thiof a été conduite par une approche globale (Gascuel et Ménard, 1997; Laurans et al., 2001b). Ces études arrivent à la même conclusion, le stock est surexploité. Les deux types d'approches donnent des résultats très similaires (tableau 3). Les différences (B/Bv) s'expliquent par le fait que les indicateurs de l'approche structurale sont établis à recrutement (R) constant. Or le recrutement diminue, la vision pessimiste donnée par le modèle global est donc réaliste et confirmée par l'approche structurale. L'effort actuel est le double de celui de maximisation, la perte de capture par rapport au MSY est de l'ordre de 30 à 40 pour cent, quant à la baisse de biomasse, elle l'est d'un facteur de l'ordre de 80 à 90 pour cent. Le calcul de biomasse totale montre une tendance proche de celle obtenue à partir de l'approche globale (figure 13). Néanmoins, en valeur brute, l'estimation par la présente étude donne des valeurs deux fois plus élevées. Cette différence peut être imputable à la valeur de la capturabilité utilisée par l'approche globale (Laurans et al., 2001b). A partir des données des chalutiers dakarois, Gascuel et Thiam (1994) estiment des indices d'abondance par modélisation GLM d'espèces démersales. L'indice d'abondance pour le thiof montre entre 1985 et 1991 une forte diminution, avec une division par trois. Les données de campagnes scientifiques du Louis Sauger sont également analysées par modélisation GLM (Laurans et al., 2001). Ce dernier indice (figure 13) confirme la diminution et montre que celle-ci se poursuit après 1991. Ces travaux sont complémentaires et montrent la situation de surexploitation du stock de thiof depuis une décennie. Cury et Worms (1982) concluent que le stock de thiof qu'ils ont défini au nord de la presqu'île du Cap-Vert est sous-exploité. Après leurs travaux, l'effort de pêche et les captures ont fortement augmenté jusqu'en 1986, ensuite les captures n'ont fait que diminuer jusqu'en 1999 (figure 1) dans des proportions fortes.
Nous avons avancé des éléments pour définir un stock unique de thiof devant les côtes sénégalaises. Dans leurs travaux, Cury et Worms (1982) font l'hypothèse de deux stocks. Un des éléments qu'ils avançaient était la différence entre le pic de PUE du métier de la ligne moteur de Soumbédioune et de Kayar, ce dernier étant plus important. Selon eux, cette différence que nous n'avons pas retrouvé dans les données récentes, montrait qu'une grosse partie de la population migrante ne passe pas la presqu'île du Cap-Vert et qu'elle se stabilise devant Kayar (PUE élevé de janvier à juin). Néanmoins, cette différence de PUE entre Kayar et Soumbédioune peut également découler d'une différence de capturabilité entre les deux zones liées aux caractéristiques du plateau continental. Au niveau de la fosse de Kayar, la topographie du fond et sa structure peut entraîner une concentration des poissons et de fait la disponibilité peut être plus élevée par une accessibilité plus grande. En effet, localement la disponibilité d'un stock peut varier positivement ou négativement. Un autre élément qu'ils avancent, est que les thiofs en saison chaude sont plus gros au sud de la presqu'île du Cap-Vert; aussi ces derniers ne prendraient pas part à la migration vers le nord. Nos résultats (tableau 2 et figure 7) montre l'inverse. A partir des données qu'utilisent les deux auteurs, il est donc difficile d'envisager la séparation en deux stocks. Cette même remarque est avancée par Barry-Gérard (1990). Cet auteur indique que certaines affirmations de Cury et Worms et de Champagnat et Domain ne sont pas confortés par les données. Avant 1980, il est difficile de comparer les données entre le sud et le nord de la presqu'île du Cap-Vert, car les pêcheries du sud sont peu suivies. Aussi, cet auteur reprend-il une comparaison de cpue par zone sur le métier de pirogue ligne moteur sur une série plus longue. Ce travail permet de montrer qu'au sud du Cap-Vert, le thiof est surtout pêché à la côte en saison froide. Dème et Diadhou indiquent qu'en Casamance les captures les plus importantes sont réalisées en saison froide. En été, dans la même zone, ces mêmes poissons sont capturés plus profondément, c'est à dire plus au large, mais en plus faible quantité. Aussi, le thiof migrerait-il selon deux schémas, la majorité de gros individus effectueraient une migration nord-sud, néanmoins il existerait une migration côte large qui affecteraient des individus plus petits. Ce dernier type de migration est noté en Côte d'Ivoire (Caverivière, 1991), le déplacement vers la côte se fait quand la thermocline est moins profonde.
Les travaux sur la croissance du thiof sont peu nombreux. Ils ont été réalisés à partir de la lecture de pièces osseuses (rayons épineux) et d'écailles. Aucune validation n'a pu confirmer les résultats avancés par les différents auteurs. Au Sénégal, deux études donnent des résultats très différents. Les travaux de Cadenat (Brusle, 1985) à partir de lecture d'écailles, donnent une croissance deux fois inférieure aux travaux de Cury et Worms (1982) réalisés à partir des rayons épineux de la nageoire dorsale. Les résultats de Cadenat sont très proche des travaux effectués en Tunisie (Bou-Ain et al., 1983a). Les résultats du présent document donnent également une croissance qui est inférieure de deux fois à celle de Cury et Worms (1982). Le suivi de la croissance des géniteurs conditionnés en bassin lors de travaux sur la reproduction (De Monbrisson, 1994) indiquent une croissance qui ne peut être de l'ordre de grandeur de celle qu'avance Cury et Worms (1982), mais de celles que nous obtenons. Enfin, un travail plus spécifique sur la vitesse de croissance à des fins d'élevages (Hassin et al., 1997) dans des conditions optimales d'alimentation (à satiété), montre une croissance plus forte que celle que nous trouvons, proche des résultats de Cury et Worms (1982). Cependant les poissons se trouvaient dans des conditions où les dépenses énergétiques sont faibles, avec un apport de nourriture optimal et des températures comprises entre 23 et 26°C. En définitive, un travail sur la croissance de cette espèce avec validation des lectures d'âge s'avère important pour affiner le diagnostic sur l'état de stock.
La relation taille-poids que nous avons utilisée (3) a été établie à partir de poissons capturés au Sénégal. En Tunisie, un travail similaire (Brusle et Prunus, 1980) permet de montrer qu'au Sénégal les jeunes individus (autour de 30 cm) ont un poids plus faible. Inversement, le poids des individus de grande taille est plus élevé au Sénégal.
L'ensemble de ce travail permet de voir les éléments qui doivent être à l'avenir affinés afin d'améliorer les données sur cette espèce et par suite le diagnostic sur l'état du stock. Une part de l'amélioration pourrait porter sur la mise en place d'un protocole d'échantillonnage de cette espèce pour la pêche industrielle. Ce type de démarche s'intégrerait dans une meilleure gestion de l'espèce mais aussi à un stade plus large dans celle de l'écosystème sénégalais.
Bibliographie
Albaret, J.J. 1994. Les poissons, biologie et peuplements. In Environnement et ressources aquatiques de Côte d'Ivoire. Tome II - Les milieux lagunaires. Edited by J.R. Durand, P. Dufour, D. Guiral, et S.G.F. Zabi. 1994 pp. 239-279.
Barry-Gérard, M. 1990. Le complexe fosse de Kayar-presqu'île du Cap-Vert constitue-t-il un obstacle aux migrations des poissons le long des côtes sénégalaises ? Doc. Sci. Centre. Rec. Océanogr. Dakar-Thiaroye, 119: 33.
Bou-Ain, A., Siau, Y. et Quignard, J.P. 1983. Les mérous des côtes sud-est de la Tunisie, 1. La pêche maritime 1262: 276-280.
Bou-Ain, A., Siau, Y. et Quignard, J.P. 1983. Les mérous des côtes sud-est de la Tunisie, 2. La pêche maritime 1263: 337-342.
Brusle, J. 1985. Exposé synoptique des données biologiques sur les mérous Epinephelus aeneus (Geoffroy Saint Hilaire, 1809) et Epinephelus guaza (Linnaeus, 1758) de l'Océan Atlantique et de la Méditerranée. FAO Synop. Pêches, (129):64p.
Brusle, J. et Brusle, S. 1975. Contribution à l'étude de la reproduction de deux espèces de mérous, Epinephelus aeneus Saint Hilaire, 1809 (Linné, 1758) et Epinephelus guaza des côtes de Tunisie. Revue de Travaux, Institut des Pêches Maritimes 39(3): 313-320.
Brusle, J. et Prunus, G. 1980. Relation taille-poids chez les mérous méditerranéens Epinephelus aeneus et Epinephelus guaza. Cybium 11: 15-21.
Cadenat, J. 1954. Notes d'ichthyologie ouest-africaine. VII. Biologie régime alimentaire. Bull.Inst.fr.Afr.Noire 16(2): 564-583.
Caverivière, A. 1994. Comparaison sur une période de 20 ans (1972-1992) des indices d'abondances obtenus sur le plateau continental sénégalais à partir des campagnes de chalutage de fond. In L'évaluation des ressources exploitables par la pêche artisanale sénégalaise, Tome 2. Edited by M. Barry-Gérard, T. Diouf, et A. Fonteneau. Colloques et Séminaires, ORSTOM ds. Paris, pp. 163-177.
Caverivière, A. 1994. Les peuplements ichtyologiques démersaux, écologie et biologie. In Environnement et ressources aquatiques de Côte d'Ivoire. Tome I- Le Milieu Marin. Edited by P. Le Loeuff, E. Marchal, et J.B. Amon Kothias. 1994 pp. 271-318.
Champagnat, C. et Domain, F. 1978. Migrations des poissons démersaux le long des côtes ouest-africaines de 10 à 24° de latitude Nord. Cah. O.R.S.T.O.M., sér. Océanogr., XVI: 239-261.
Cury, P. et Roy, C. 1988. Migration saisonnière du thiof (Epinephelus aeneus) au Sénégal: influence des upwellings sénégalais et mauritanien. Oceanologia Acta 11, n°1: 25-36.
Cury, P. et Worms, J. 1982. Pêche, biologie et dynamique du thiof (Epinephelus aeneus E.Geoffroy Saint Hilaire, 1817) sur les côtes sénégalaises. Centre de recherches océanographiques de Dakar-Tiaroye 86p.
De Monbrisson, D. 1994. Reproduction et élevage larvaire du mérou blanc. Rapp. DAA Unité Halieutique de l'ENSAR, Rennes, 46p.
Domain, F. 1980. Contribution à la connaissance de l'écologie des poissons démersaux du plateau continental sénégalo-mauritanien. Les ressources démersales dans le contexte général du golfe de Guinée. Thèse doct. d'Etat Univ. Paris VI et Mus. Hist. Nat. 342p.
Ezzat, A.A., Wadie, W.F., Mikail, M.Y., et Hashem, M.T. 1981. Age and growth of Epinephelus aeneus in the Egyptian Mediterraean waters. Bull. Inst. Oceanogr. Fish. 7(3): 395-406.
Ferraris, J. 1994. Prise par unité d'effort et indice d'abondance en pêche artisanale: Quelques éléments de réflexion à partir de la pêcherie sénégalaise. In L'évaluation des ressources exploitables par la pêche artisanale sénégalaise, Tome2. Edited by M. Barry-Gérard, T. Diouf et A. Fonteneau. pp. 105-120.
Ferraris, J., Samb, B., et Thiam, M. 1994. Les statistiques de pêche au CRODT. In L'évaluation des ressources exploitables par la pêche artisanale sénégalaise, Tome 2. Edited by M. Barry-Gérard, T. Diouf et A. Fonteneau. Colloques et Séminaires ORSTOM éds. Paris, pp. 73-93.
Fontana, A. 1979. Etude du stock démersal côtier congolais. Biologie et dynamique des principales espèces exploitées. Propositions d'aménagement de la pêcherie. Thèse doct. d'Etat Univ. Paris VI et Mus. Hist. Nat. 300p.
Franqueville, C. et Fréon, P. 1976. Relation poids-longueur des principales espèces de poissons marins au Sénégal. Doc. Sci. Centre. Rec. Océanogr. Dakar-Thiaroye, (24): 34p.
Gascuel, D. 1994. Une méthode simple d'ajustement des clés taille/âge: application aux captures d'albacores (Thunnus albacares) de l'Atlantique Est. Can. J. Fish. Aquat. Sci. 51: 723-733.
Gascuel, D. et Ménard, F. 1997. Assessment of a multispecies fishery in Senegal, using production models and diversity indices. Aquat. Living.Resour 10: 281-288.
Gascuel, D. et Thiam, M. 1994. Evolution de l'abondance des ressources démersales sénégalaises. In L'évaluation des ressources exploitables par la pêche artisanale sénégalaise, Tome 2. Edited by M. Barry-Gérard, T. Diouf, et A. Fonteneau. Colloques et Séminaires, ORSTOM ds. Paris, pp. 191-213.
Hasselblad, V. 1966. Estimation of parameters for mixture of normal distributions. Technometrics 8: 431-444.
Hassin, S., de Mondrisson, D., Hanin, Y., Elizur, A., Zohar, et Popper, D.M. 1997. Domestication of the white grouper, Epinephelus aeneus, Growth and Reproduction. Aquaculture 156: 305-316.
Kébé, M. 1994. Principales mutations de la pêche artisanale maritime sénégalaise. In L'évaluation des ressources exploitables par la pêche artisanale sénégalaise, Tome 2. Edited by M. Barry-Gérard, T. Diouf, et A. Fonteneau. Colloques et Séminaires ORSTOM éds. Paris, pp. 43-58.
Laurans, M., Barry, M. et Gascuel, D. 2001. Diagnostics de cinq stocks sénégalais par l'approche globale (Galeoides decadactylus, Pagellus bellottii, Pseudupeneus prayensis, Pagrus caeruleostictus, Epinephelus aeneus). SIAP, Analyses, Doc. Tec. 2: 26-35.
Laurans, M., Mohammed-Fall, K.O., et Gascuel, D. 2001. Rapport de la réunion «Analyses monospécifiques» Nouadhibou, 3 au 7 juin 2000. SIAP, Analyses, Doc. Tec.1.
Pauly, D. 1980. On the interrelationships between natura mortality, growth parameters, and mean environmental temperature in 175 fish stocks. J. Cons. int. Explor. Mer, 39(2): 175-192.
Pauly, D. et Moreau, J. 1997. Méthodes d'évaluation des ressources halieutiques.
Shepherd, J.G. 1987. A weakly parametric method for estimating growth parameters from length composition data. In Length-based methods in fisheries research. Edited by D. Pauly and G.R. Morgan. ICLARM Conf. Proc13., pp. 113-119.
ANNEXE
Tableau 1: Poids moyen en gramme des thiofs capturés par zone selon la saison.
|
|
Saison froide |
Saison chaude |
|
Zone 1 |
3 472 |
933 |
|
Zone 3 |
4 939 |
3 471 |
|
Zone 4 |
3 996 |
1 964 |
|
Zone 5 |
2 021 |
994 |
Tableau 2: Capture en nombre d'individu par groupe d'âge.
|
|
Année |
||||||||||
|
Groupe d'âge |
1989 |
1990 |
1991 |
1992 |
1993 |
1994 |
1995 |
1996 |
1997 |
1998 |
1999 |
|
1 |
1 543 |
843 |
2 043 |
2 333 |
4 050 |
3 521 |
7 739 |
8 757 |
82 246 |
24 812 |
34 591 |
|
2 |
39 438 |
26 149 |
30 284 |
80 228 |
23 375 |
72 924 |
71 952 |
213 813 |
99 125 |
120 121 |
103 820 |
|
3 |
49 933 |
26 519 |
25 942 |
109 473 |
52 082 |
35 151 |
4 6281 |
56 773 |
62 426 |
60 903 |
81 223 |
|
4 |
86 551 |
65 769 |
42 085 |
60 712 |
95 024 |
40 249 |
57 064 |
66 283 |
60 372 |
88 240 |
55 792 |
|
5 |
50 276 |
57 844 |
55 101 |
48 889 |
72 367 |
68 610 |
30 311 |
60 203 |
49 344 |
44 448 |
34 725 |
|
6 |
67 492 |
81 128 |
70 703 |
54 452 |
53 628 |
58 963 |
73 394 |
61 891 |
62 992 |
44 376 |
23 162 |
|
7 |
64 006 |
56 135 |
72 244 |
87 705 |
87 610 |
109 318 |
45 759 |
34 762 |
31 585 |
19 999 |
68 118 |
|
8 |
75 798 |
79 289 |
66 464 |
75 580 |
81 754 |
109 592 |
77 840 |
27 161 |
57 419 |
36 493 |
32 521 |
|
9 |
62 979 |
76 735 |
42 426 |
69 212 |
31 057 |
49 843 |
46 133 |
74 349 |
29 956 |
21 144 |
19 662 |
|
10 |
21 823 |
24 364 |
13 333 |
25 966 |
18 339 |
16 349 |
16 877 |
11 299 |
22 836 |
5 000 |
10 000 |
|
11 |
14 396 |
13 213 |
19 078 |
13 992 |
10 414 |
29 149 |
21 100 |
11 872 |
17 000 |
4 200 |
8 500 |
|
12 |
3 743 |
7 525 |
2 783 |
5 150 |
2 343 |
5 923 |
7 951 |
5 508 |
6 000 |
1 792 |
4 872 |
|
13 |
2 808 |
5 644 |
2 087 |
2 059 |
1 757 |
4 442 |
5 963 |
4 131 |
4 500 |
1 344 |
1 587 |
|
14 |
1 872 |
3 762 |
1 391 |
1 373 |
1 171 |
2 962 |
3 975 |
2 754 |
3 000 |
896 |
1 058 |
|
15 |
936 |
1 881 |
696 |
686 |
586 |
1 481 |
1 988 |
1 377 |
1 500 |
448 |
529 |
Tableau 3: Comparaison des résultats obtenus par l'approche structurale et globale pour le stock de thiof du Sénégal.
|
|
mf MSY |
Y/MSY |
B/Bv |
|
Approche globale |
0.47/0.6 |
0.56/0.7 |
0.08/0.16 |
|
Approche analytique |
0.52/0.77 |
0.8/0.9 |
0.19/0.28* |
* B/R relative (ne prends pas en compte la baisse de R).
![]()
![]()
![]()