![]()
![]()
![]()
D. G. NIKLES
D. GARTH NIKLES es el funcionario encargado de la Sección de Genética Arbórea de la Estación de Investigaciones Forestales de Beerwah, Queensland (Australia). Este trabajo se publica con autorización del Conservador del Departamento Forestal de Brisbane (Australia).
Se ha llevado ya a cabo la selección de las especies y las poblaciones geográficas más valiosas de plantas de cultivo y animales domésticos, y las prácticas actuales de mejora genética afectan sobre todo a la mejora de variedades conocidas. Por el contrario, la explotación y reproducción de bosques comerciales afecta principalmente a especies silvestres, con sólo una utilización reciente y secundaria de estirpes seleccionadas para alto rendimiento.
La selección genética para mejorar el rendimiento de los árboles forestales puede efectuarse en el plano de la especie, la variedad, la procedencia y el árbol.
Esta selección puede ser efectiva porque las especies, variedades o procedencias indígenas y naturalizadas no explotan siempre a fondo el ambiente ordenado de una región. Por lo general están adaptadas para sobrevivir a las condiciones extremas de clima y suelo locales más que para producir los rendimientos máximos de los productos que necesita el hombre. Además, puede que no produzcan el tipo deseado de madera. Donde las especies no son satisfactorias por éstas u otras razones, puede a menudo mejorarse el rendimiento mediante la introducción, ensayo y selección adecuadas de las poblaciones de árboles exóticos mejor adaptadas y más productivas.
La selección de individuos para aumentar el rendimiento puede explotar la variación entre árboles pertenecientes a una misma población. Está más indicada cuando se ha determinado la mejor procedencia (Dietrichson, 1969) y cuando se dispone localmente de una gran población para la selección.
La hibridación interespecífica y la inter-racial se han utilizado para mejorar el rendimiento de los árboles forestales y representan métodos prometedores de mejora genética en ciertas circunstancias.
Como base para el estudio de la mejora genética para crecimiento y rendimiento en este trabajo, se describen los componentes del rendimiento de un bosque comercial. Se presentan ejemplos de las ganancias que pueden conseguirse con la selección y la hibridación. También se estudian los problemas que se plantean en la preparación de programas eficientes de mejora genética para alto rendimiento.
La producción total de materia seca de un sistema vegetal constituye el rendimiento biológico, pero el hombre sólo aprovecha una parte de éste, el llamado rendimiento económico (Niciporovic, 1954; Ovington, 1965, 1968). La relación dinámica entre el rendimiento biológico (Ybiol) y el rendimiento económico (Yecon) de un cultivo en un ambiente específico puede expresarse como Ybiol x HI = Yecon, donde HI es el índice de aprovechamiento de Donald (1962). La mejora para aumentar el Yecon se hace posible si la variación heredable se presenta en Ybiol, o en HI, o en ambos factores. Debe admitirse la posibilidad de una interacción significativa de los componentes genéticos y ambientales de Yecon.
El rendimiento biológico de un bosque comercial depende de la capacidad inherente de crecimiento del material genético que se utiliza y del ambiente en que se cultiva. Ejemplos de las características del cultivo que pueden afectar a la producción orgánica total son la superficie y exposición foliar, la eficiencia fotosintética, el índice de crecimiento relativo, el ritmo del crecimiento, el hábito de radicación, el potencial de altura definitiva, el estado sanitario y la respuesta a la densidad vegetal (Kozlowski, 1962; Black, 1966; Wareing, 1966; Dietrichson, 1969).
El índice de aprovechamiento, en sentido literal, es el índice del volumen o peso comercializable de madera que puede extraerse del bosque en relación con el que el mismo produce. Este concepto general ignora, sin embargo, el hecho de que sólo una parte de la madera cosechada se recupera en forma de productos utilizables. El término específico índice de aprovechamiento neto (HIneto), es más útil a los fines de este trabajo. Se le define como el índice del volumen de material recuperable después de la elaboración en relación con el producido en el bosque (rendimiento biológico).
El índice de aprovechamiento neto que se obtiene al explotar un bosque depende en gran parte del aprovechamiento de la madera disponible y de la efectividad de la recolección y elaboración. En muchos casos, pueden lograrse considerables mejoras del rendimiento económico prestando mayor atención a esos aspectos (Ovington, 1968). Pero el índice de aprovechamiento neto está también influido por las características morfológicas de los árboles, por el efecto que éstas tienen en el tipo de aprovechamiento, la facilidad de recolección y la recuperación de productos.
Cabe esperar índices de aprovechamiento neto variables entre poblaciones y árboles que difieran en la rectitud del fuste, ángulo de ramificación, conicidad del tronco, calidad de la madera y distribución del fotosintato entre fuste, ramas, corteza, hojas, conos y raíces. Por ejemplo, las variedades de abundante ramificación y derechas de Pinus caribaea Mor. de Cuba (Slee y Nikles, 1968) y de Pinus pinaster Aiton de Córcega (Hopkins, 1960), deben tener índices de aprovechamiento neto mayores que los de otras variedades y razas de las especies respectivas.
Interacción de especies de Pinus - estación en Beerwah, Queensland (Australia), reflejada en el rendimiento económico superior de P. caribaea Mor., en comparación con P. taeda L. en suelos residuales de tierras rojas, y en el rendimiento mucho menor de P. caribaea con relación a P. taeda en suelos húmicos.
FOTO: NIKLES
El producto Ybiol x HIneto puede denominarse rendimiento económico neto o rendimiento recuperable. El rendimiento económico neto de un bosque comercial se define como el volumen, peso o valor de los productos que pueden obtenerse de la madera producida por unidad de superficie del bosque durante un turno. Los componentes biológicos del rendimiento económico neto se enumeran en el Cuadro 1 y constituyen los criterios generales que podrían utilizarse en la selección directa para aumento del rendimiento.
CUADRO 1. - COMPONENTES BIOLÓGICOS DEL RENDIMIENTO ECONÓMICO NETO
|
Componente del rendimiento |
Tipo de rendimiento |
|||
|
Arboles |
Rodales |
|||
|
Volumen |
Peso |
Volumen |
Peso |
|
|
Altura comercializable |
Sí |
Sí |
Sí |
Sí |
|
Area basimétrica sin corteza |
Sí |
Sí |
Sí |
Sí |
|
Coeficiente mórfico |
Sí |
Sí |
Sí |
Sí |
|
Coeficiente de calidad del fuste |
Sí |
Sí |
Sí |
Sí |
|
Coeficiente de calidad de la madera |
Sí |
Sí |
Sí |
Sí |
|
Densidad básica |
No |
Sí |
No |
Sí |
|
Coeficiente de densidad del rodal |
No |
No |
Sí |
Sí |
IMPORTANCIA DE LA SELECCIÓN DE ESPECIES
En muchos países, la producción de algunas especies exóticas es muy superior a la de la mayoría de los árboles forestales indígenas. Por ejemplo, Wood (1962) ha señalado que en el Reino Unido la especie introducida Pseudotsuga taxifolia (Poir.) Britt. rinde en 50 años un volumen aproximadamente doble al de las plantaciones de la indígena Quercus robur L. en 100 años en estaciones de la misma calidad. Hay otros muchos ejemplos de la utilización ventajosa de especies exóticas en Europa (Edwards, 1963), Sudamérica (Barrett, 1969), Africa (Iyamabo, 1969), Australia y Nueva Zelandia.
Muchas veces pueden implantarse con éxito en una región forestal varias especies indígenas e introducidas, pero puede haber entre ellas grandes diferencias en rendimiento biológico, índice de aprovechamiento neto y demanda de sus productos. Ello ocurre con más frecuencia cuando dentro de la región existen considerables variaciones ambientales, que originan importantes interacciones entre especies y estaciones. Por ejemplo, en las tierras bajas costeras de Queensland sudoriental, en Australia, se están estableciendo plantaciones de especies de Pinus. Pueden reconocerse varios tipos de suelos en los cuales tres especies satisfactorias han dado respuestas diferentes al cabo de 15 años (Cuadro 2 y Figura 4).
CUADRO 2. - CLASIFICACIÓN DE LOS INCREMENTOS MEDIOS ANUALES DE VOLUMEN DE LAS ESPECIES DE Pinus CULTIVADAS DURANTE 15 AÑOS EN TRES TIPOS DE SUELOS EN QUEENSLAND SUDORIENTAL
|
Especie |
Suelos residuales de tierras rojas |
Suelos podsólicos lateríticos |
Suelos húmicos |
|
P. caribaea Mor |
1 |
1 |
3 |
|
P. elliottii Engelm |
2 |
2 |
2 |
|
P. taeda L |
3 |
2 |
1 |
|
Diferencia entre las categorías 1 y 2 (por ciento) |
32 |
38 |
40 |
FUENTE: Datos facilitados por D. Jermyn, Estación de Investigaciones Forestales, Beerwah, Queensland (Australia).
Jackson (1965) examina también otros ejemplos de las interacciones especie-estación. Señala que un medio efectivo de aumentar el rendimiento es la delineación apropiada de las estaciones principales y la asignación correcta de las especies a las mismas. En efecto, si una especie no se adapta bien a la estación de plantación, se presenta más pronto o más tarde el peligro de pérdidas económicas por causa de una tensión ambiental gradual o repentina.
Este concepto ideal de seleccionar las especies mejor adaptadas y más valiosas para cada una de las estaciones principales puede plantear problemas tales como la elección de las especies, la delineación de la estación, el establecimiento, la ordenación y recolección de bloques discontinuos de diferentes especies, y la comercialización de distintos tipos de productos. Por otro lado, la utilización de varias especies en una región puede constituir una salvaguardia valiosa contra las plagas y las enfermedades y contra los cambios en las necesidades de productos en el futuro.
IMPORTANCIA DE LA SELECCÍON DE PROCEDENCIAS
Los estudios genecológicos de muchas especies vegetales han mostrado que, como resultado de la selección natural y de otros fenómenos, se desarrollan poblaciones locales características (Cooper, 1963; Grant, 1963; Heslop-Harrison, 1964; y Langlet, 1967). Son muchas las pruebas que demuestran que las poblaciones geográficas genéticamente diferentes de la mayoría de las especies arbóreas de amplia difusión varían grandemente en rendimiento económico en las distintas estaciones de ensayo (Squillace y Silen, 1962; Langlet, 1963; Lines, 1965; Wells y Wakeley, 1966; Marsh, 1969; Wells, 1969).
En un estudio de la literatura forestal que trata de las interacciones entre el origen de las semillas y el ambiente, Squillace (1969) ha señalado la aparición frecuente de efectos importantes debidos a la localidad, la estación, el fotoperíodo y el tratamiento cultural. Como ejemplo del efecto combinado de la procedencia y de la localidad de plantación en El rendimiento, se presenta en el Cuadro 3 la clasificación de varias poblaciones de P. pinaster según su incremento medio anual de volumen en diversas localidades de ensayo en Sudáfrica y Australia occidental. La raza portuguesa es preferida para la plantación por su rendimiento consecuentemente alto, pero la raza de Córcega, que se caracteriza por una forma de fuste y un hábito de ramificación superiores (índice de aprovechamiento mayor), puede ser valiosa para la hibridación inter-racial.
CUADRO 3. - CLASIFICACIÓN DE LOS INCREMENTOS MEDIOS ANUALES DE VOLUMEN DE PROCEDENCIAS DE Pinus pinaster AITON CULTIVADOS EN CUATRO LOCALIDADES DE SUDÁFRICA Y OTRAS CUATRO DE AUSTRALIA OCCIDENTAL
|
Lugar de procedencia |
Sudáfrica |
Australia occidental |
||||||
|
A |
B |
C |
D |
A |
B |
C |
D |
|
|
Portugal |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
|
Las Landas |
2 |
2 |
2 |
2 |
2 |
2 |
2 |
2 |
|
Esterel |
3 |
3 |
3 |
3 |
4 |
4 |
4 |
3 |
|
Córcega |
3 |
3 |
3 |
4 |
3 |
3 |
3 |
4 |
FUENTE: Para Sudáfrica, Marsh (1969); para Australia occidental, Hopkins (IUFRO).
Las ganancias efectivas en la producción volumétrica que se pueden obtener con la selección de procedencias se han calculado en un ensayo con árboles de 14 años de 14 procedencias de Pinus taeda L. en Beerwah, Queensland, Australia. Se registró una diferencia del 38 por ciento entre las poblaciones mejores y las segundas. La población mejor produjo un volumen doble al de la peor.
PROBLEMAS DE SELECCIÓN DE ESPECIES Y PROCEDENCIAS
Jacobs, (1964) y Zobel (1969) han señalado el peligro de la utilización de especies o procedencias exóticas cuando no se tiene la seguridad de que se adapten a los ambientes locales:
1. Pueden sucumbir a las condiciones extremas no habituales del ambiente.2. Su índice de crecimiento y salud pueden disminuir notablemente después de un buen comienzo, debido a defectos no sospechados de la estación; esta disminución puede no ponerse de manifiesto hasta el segundo turno.
3. Pueden ser atacadas por insectos o patógenos, especialmente si crecen en estaciones marginales.
4. El tipo y calidad de la madera producida pueden ser decepcionantes.
Un programa dinámico y multifásico de introducción y ensayo es ineludible si las especies locales no son satisfactorias. A veces, no es práctico hacer ensayos de especies y procedencias exóticas durante un turno completo antes de iniciar el establecimiento de plantaciones comerciales. Por ello, deben implantarse inicialmente muchas especies y varias procedencias de cada una; la mayoría de ellas serán rechazadas en las primeras o últimas fases de los ensayos. Algunas irán progresando gradualmente hasta alcanzar la condición de especies comerciales importantes.
Entre los problemas que plantea la introducción y ensayo de especies y procedencias se cuentan la elección inicial, los ensayos tempranos, las pruebas de campo y la cooperación efectiva.
La elección inicial de especies y procedencias para su introducción se orientará por medio de un conocimiento general de los tipos de madera requeridos, y de la ecología y las asociaciones vegetales de especies convenientes que crezcan en regiones donantes de clima semejante al de la región receptora. El concepto de la equiparación de los climas no debe aplicarse con excesiva rigidez, ni deben tampoco ignorarse todas las especies de importancia secundaria de las regiones donantes. Varios autores han estudiado los detalles de la elección y muestreo de especies para introducción (Wright, 1962; Callaham, 1964; Jacobs, 1964; Lines, 1965, 1967a, 1967b; Morandini, 1967; Iyamabo, 1969).
Los estudios a corto plazo de la variación de las características de adaptividad en las poblaciones silvestres y de las progenies cultivadas en un vivero, invernadero o fitotrón pueden ayudar a reducir el material destinado a los estudios de campo y a la elección de las estaciones de ensayo (Roche, 1968). Entre las características que afectan al crecimiento y al rendimiento susceptibles de ser objeto de esos estudios se cuentan el ritmo del crecimiento (Roche, 1968; Dietrichson, 1969), la resistencia a la helada y la sequía (Day y Peace, 1934; van Buijtenen, 1966a; Bolotin, 1969) y la resistencia a las enfermedades y los insectos (Gerhold y otros, 1966).
El ensayo de campo multifásico, con repetición en el tiempo y en el espacio, es esencial para acomodar grandes números de especies o procedencias y para el muestreo adecuado de ambientes. Existen muy diversas opiniones en cuanto al tipo y tamaño convenientes de la parcela de ensayo, los tratamientos culturales, el diseño experimental, la duración del ensayo y los criterios de evaluación. Las decisiones al respecto deben adoptarse de acuerdo con los objetivos del programa. Como orientación en la planificación de ensayos de campo, véase Callaham, 1964; Leuchars, 1965; Lines, 1967a, 1967b; Morandini, 1967, e Iyamabo, 1969.
La necesidad de una cooperación internacional en los estudios genecológicos y de rendimiento de los árboles forestales ha sido recalcada muchas veces, pero queda aún mucho por hacer (FAO, 1969; Hagman, 1969; Larsen, 1969; Sarvas, 1969). Esta cooperación debe tener por objetivo facilitar colecciones de semillas adecuadas, asegurar la conservación de los conjuntos de genes de muestra, determinar las estructuras de la variación y sus causas, y formular normas de orientación para la transferencia de semillas en condiciones ventajosas. Se han realizado trabajos ejemplares sobre las normas relativas al movimiento de semillas en relación con Pinus, sylvestris L. en Suecia (Langlet, 1963), con varias coníferas exóticas en el Reino Unido (Lines, 1965), P. caribaea en Queensland (Slee y Nikles, 1968) y P. taeda en los Estados Unidos (Wells, 1969). El análisis de la variación por medio de la regresión múltiple, tal como lo describen Morgenstern y Roche (1969), constituye un nuevo método prometedor para la delineación de zonas de semillas.
Determinando y utilizando las razas mejor adaptadas y de alto rendimiento para cada una de las estaciones principales, se pueden evitar errores costosos y establecer la base esencial para conseguir una ganancia genética máxima por la mejora de la población.
GANANCIAS MEDIANTE LA SELECCIÓN INDIVIDUAL
En esta sección se estudiarán diversos métodos de utilización de individuos selectos para la mejora del rendimiento económico neto en una población base. Todos estos métodos se basan en la variación hereditaria en rendimiento biológico e índice de aprovechamiento neto entre los distintos árboles.
Se han demostrado grandes diferencias de rendimiento económico en edades jóvenes entre líneas de progenies (Ehrenberg, 1966; Eldridge, 1966; Webb y Barber, 1966; Orr-Ewing, 1967; van Buijtenen, 1968; Nikles y Smith, 1969; Snyder, 1969; Zobel y otros, 1969, Cuadro 4) y entre clones (Fielding y Brown, 1961; Burdon, 1968; Pawsey, 1968; Nienstaedt y King, 1969). Ello significa que la selección individual puede ser eficaz para mejorar el rendimiento. Se han predicho ganancias genéticas importantes en el caso de varios métodos empleando valores conocidos o supuestos de los parámetros de población y diversas intensidades de selección (Namkoong y otros, 1966; Shelbourne, 1968, 1969; Snyder, 1969).
La ganancia realizada puede estimarse comparando la progenie de los árboles selectos con una muestra aleatoria de la población de base. Se han realizado ganancias sustanciales en algunos ensayos de progenies (Cuadro 4). Los detalles de los ensayos con individuos jóvenes tienen importancia no sólo porque frecuentemente permiten predecir lo que ocurrirá, sino también porque el rápido crecimiento temprano puede ser útil per se, por ejemplo, para la pronta supresión de las malas hierbas, aun cuando ésta no es sostenida.
El procedimiento más simple de mejora es el de la selección masiva, es decir, el uso de semillas polinizadas por el viento de fenotipos superiores. Puede proporcionar ganancias apreciables si las heredabilidades son altas, pero, si son bajas, la selección basada en los resultados del ensayo de progenies es mucho más efectiva que la selección fenotípica simple (véase Cuadro 4: A1-4 y B1-4). Cuando el apareamiento se limita a progenitores selectos, lo que equivale a las condiciones de un huerto de semillas totalmente aislado, se obtienen ganancias mayores (compárese Cuadro 4: A2, A4 y C3, C5). Después de aplicar una nueva fase de selección, en la que sólo se utilizan los progenitores que tienen una gran aptitud combinativa general (equivalente a la depuración en un huerto de semillas), se obtiene una ganancia mayor (compárese Cuadro 4: C1, C2, C3 y D1, D2, D3). Otro pequeño aumento de la ganancia, en comparación con el tiempo y el trabajo que se necesitarían para obtenerlo, fue posible con P. elliottii escogiendo para la producción el mejor 10 por ciento de los cruzamientos monohíbridos (compárese Cuadro 4: D3 y E1). Estos resultados coinciden en general con las predicciones de Shelbourne (1969) en el caso de Pinus radiata Don.
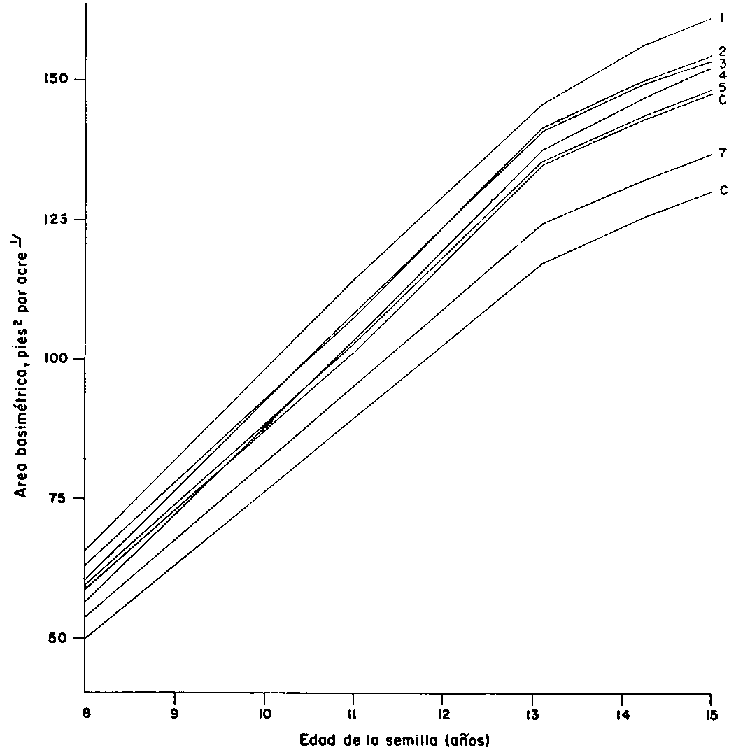
Las pruebas más claras de la obtención de ganancias en el rendimiento por la selección individual proceden de algunos de los ensayos de progenies más antiguas (compárese Cuadro 4: A5, C3 y C6). Los experimentos afectaban a parcelas separadas, repetidas, y de hileras múltiples de material comercial y progenies de árboles progenitores escogidos por fenotipo superior. Las comparaciones se referían al volumen o a la producción en área basimétrica (Cuadro 4) y a la densidad básica de la madera (Pawsey, 1968; Nikles y Smith, 1969). Las últimas mediciones se realizaron a los 12,5 a 15 años. En esa época, la cubierta de copas llevaba ya cerrada la mitad, aproximadamente, de los períodos de ensayo. Ello significa que casi toda 1 madera se produjo en condiciones competitivas dentro de las progenies. La edad de los árboles era más de un tercio de los turnos probables de las dos especies, P. radiata y P. elliottii. Esto sugiere que los resultados pueden ser indicaciones seguras de los rendimientos relativos al acercarse la época del turno. Las curvas de crecimiento del área basimétrica de varias progenies de fratrias de árboles y material comercial selectos de P. elliottii (Figura 5) y las fotografías (Figura 6) indican que el material escogido ha establecido una clara superioridad. Además, la densidad básica de la madera de las progenies selectas y de las comerciales era igual (Pawsey, 1968; Nikles y Smith, 1969). Esto quiere decir que el rendimiento económico bruto, expresado tanto en volumen como en peso de madera en seco, ha aumentado gracias a la selección.
1 1 pie cuadrado por ave = 0,2 m² por hectárea.
Las familias de árboles selectos de P. elliottii y P. radiata en los ensayos de progenies referidos anteriormente muestran también una mejora considerable en la derechura del fuste (Nikles, 1966; Brown, 1969, Pawsey, 1969). Esta mejora de la calidad del fuste dará por resultado un índice de aprovechamiento neto más elevado para la mayoría de los usos finales. De aquí se concluye que cabe esperar un aumento del rendimiento económico neto de un 30 por ciento, aproximadamente, a la edad de 15 años a partir del establecimiento y la depuración subsiguiente de algunos huertos de semillas de P. elliottii y P. radiata en Australia. Los programas de mejora genética de este tipo son sumamente valiosos, ya que los estudios recientes muestran que, para justificar los gastos, se necesitan ganancias del orden de sólo 5 por ciento en la época de la cosecha (Davis, 1969; Shepherd y Slee, 1969).
CUADRO 4. - GANANCIAS GENÉTICAS EN EL RENDIMIENTO OBTENIDAS CON MÉTODOS DE MEJORA DE INTENSIDAD PROGRESIVA
|
Método de selección y apareamiento, referencia |
Especie tipo del rodal 1 |
Edad esperada a partir de semilla (años) |
Número de progenitores: familias |
Tipo de parcela ² |
Porcentaje de ganancias |
||
|
Altura |
Diámetro |
Volumen |
|||||
|
A. Selección fenotípica, polinización por el viento; no se especifica la intensidad de la selección |
|||||||
|
1. Barber (1964) |
P. elliottii (P) |
8 |
11:11 |
M |
7,6 |
4,9 |
- |
|
2. Nikles (1966; 1969) |
P. elliottii (P) |
10 |
19:19 |
M |
- |
- |
10,7 |
|
3. Woessner (1965). |
P. taeda (W) |
6 |
31:31 |
M |
3,6 |
5,9 |
13,0 |
|
4. Nikles (1969) |
Araucaria cunninghamii (P) |
6,5 |
20:20 |
M |
13,0 |
- |
- |
|
5. Brown (1969) |
P. radiata (P) |
2,5 |
23:23 |
M |
- |
³10,0 |
- |
|
B. Selección de ensayo de progenies, polinización por el viento; no se especifica la intensidad de la selección fenotípica |
|||||||
|
1. Barber (1964) |
P. elliottii (P) |
8 |
4:4 |
M |
12,4 |
8,9 |
- |
|
2. Nikles (1969) |
P. elliottii (P) |
10 |
8:8 |
M |
- |
- |
20,0 |
|
3. Woessner (1965) |
P. taeda (W) |
6 |
14:14 |
M |
6,2 |
10,3 |
- |
|
4. Nikles (1969) |
Araucaria cunninghamii (P) |
6,5 |
7:7 |
M |
22 |
- |
- |
|
C. Selección fenotípica,, policruzamiento o monocruzamiento parcial no se especifica la intensidad de la selección |
|||||||
|
1. Kraus (1968). |
P. elliottii (W) |
6 |
18:18 |
M |
4,6 |
- |
- |
|
|
P. elliottii (W) |
6 |
19:19 |
M |
10,1 |
- |
- |
|
2. Goddard (1968) |
P. elliottii (W) |
6 |
9:9 |
L |
- |
- |
439,4 |
|
|
P. elliottii (W) |
6 |
9:9 |
L |
- |
- |
438,0 |
|
|
P. elliottii (W) |
6 |
15:15 |
L |
4,3 |
6,1 |
411,8 |
|
3. Nikles (1969) |
P. elliottii (P) |
11,2 |
9:8 |
M |
- |
- |
17,9 |
|
|
P. elliottii (P) |
13,5 |
6:8 |
M |
- |
- |
22,3 |
|
|
P. elliottii (P) |
15 |
10:12 |
M |
- |
7,9 |
20,1 |
|
4. Thulin (1969) |
P. radiata (P) |
11 |
11:15 |
L |
- |
- |
14,0 |
|
5. Nikles (1969) |
Araucaria cunninghamii (P) |
6,5 |
9:12 |
M |
20,0 |
- |
- |
|
6. Pawsey (1968) |
P. radiata (P) |
12,5 |
21:21 |
M, L |
- |
- |
22,2 |
|
D. Selección de ensayo de progenies, policruzamiento o monocruzamiento parcial; 30 a 50 por ciento de familias conservadas |
|||||||
|
1. Kraus (1968) |
P. elliottii (W) |
6 |
9:9 |
M |
58,5 |
- |
- |
|
2. Goddard (1968) |
P. elliottii (W) |
6 |
8:8 |
L |
- |
- |
4,521,3 |
|
3. Nikles (1969) |
P. elliottii (P) |
11-15 |
9:9 |
M |
- |
- |
27, 5 |
|
E. Selección de ensayo de progenies, de las mejores fratrias; 10 por ciento de familias conservadas |
|||||||
|
1. Nikles (1969). |
P. elliottii (P) |
11-15 |
5:3 |
M |
- |
- |
32,5 |
1 Especie Pinus salvo indicación en contrario; árboles progenitores elegidos en plantaciones (P) o rodales silvestres (W). - ² M = parcela de hileras múltiples; L= parcela lineal. ³ Area basimétrica. - Ganancia expresada en peso en seco del fuste. - Ganancias mínimas computadas por no limitarse los apareamientos ea progenitores selectos.
Una progenie de fratria de 15 años (figura 6.1) de dos fenotipos selectos de Pinus elliottii Engelm., que es una de las varias familias que producen árboles más rectos y un volumen por hectárea superior en un 30 por ciento al del material comercial (figura 6.2) en condiciones comparables, en Beerwah, Queensland (Australia).


PROBLEMAS QUE PLANTEA LA MEJORA DEL RENDIMIENTO DENTRO DE UNA POBLACIÓN
Selección de individuos para aumentar el rendimiento
El rendimiento económico neto de un bosque determinado constituye evidentemente una característica muy compleja en la que entran muchos componentes (Cuadro 1). A menudo, cuando se inicia un programa de mejora genética, se sabe muy poco acerca de los valores económicos relativos, los grados de control genético y las interrelaciones de los factores que influyen en el rendimiento. La elección de los criterios de selección y la asignación de un peso relativo a los mismos debe, por tanto, ser al principio subjetiva. Por lo general, se prepara al comienzo del programa un plan sistemático para ayudar a la selección de fenotipos superiores (Haley, 1960; Mergen, 1962; Campbell, 1964; Forshell, 1964; Pederick, 1968).
Para identificar los genotipos de alto rendimiento, se necesitan medios más eficaces que la selección fenotípica. Con frecuencia, se han establecido ensayos de progenies para comparar los valores de mejora de los progenitores, pero no es probable que este único objetivo pueda justificar su costo elevado (Libby y otros, 1969; Toda, 1969). La selección dentro de los ensayos de progenies puede efectuarse con exactitud cada vez mayor combinando los datos sobre los rendimientos de individuos, familias y progenitores (Falconer, 1960; Namkoong, 1966). Esta selección combinada tendrá un uso extendido en los trabajos de mejora del árbol cuando se cuente con ensayos de progenie adecuados.
Los índices de selección son especialmente útiles cuando hay que mejorar simultáneamente varias características complejas e interconexas. La construcción de índices de características múltiples para la clasificación de un árbol por su propio rendimiento ha sido descrita por Northcott (1965), van Buijtenen (1966b), e Illy (1969). Para ser útiles, los índices deben ser fidedignos, es decir, basarse en estimaciones precisas de los parámetros genéticos, ambientales y económicos, pero esto exige la realización de experimentos amplios y costosos (Stonecypher, 1966, 1969a). Otro método es el sugerido por Namkoong (1969) para el caso frecuente en que no se dispone de los datos necesarios.
Integración de los experimentos de evaluación, selección y estimación
La selección recurrente de las múltiples características que afectan al rendimiento exige el apareamiento de muchos progenitores selectos, ya que se necesitan muchas progenies para evaluar a los progenitores y sus familias, para medir la ganancia genética realizada, para escoger árboles superiores para el ciclo siguiente de mejora, para calcular los parámetros de población, y por último para producir una cosecha ordinaria. Deben emplearse sistemas de apareamiento y diseños experimentales que sean eficaces para todos estos objetivos con objeto de ahorrar esfuerzo (Stern y Hattemer, 1964; Schutz y Cockerham, 1966; Libby, 1969, Toda, 1969).
Se han publicado descripciones de diversos diseños de apareamiento y experimentales que se utilizan para la mejora de la población arbórea (Johnsson, 1964; Libby 1964, 1968, 1969; Braaten, 1965; Goddard, 1965; Burley y otros, 1966; Stonecypher, 1966; Roberds y otros, 1967; Bingham, 1968). Los sistemas de apareamiento varían en complejidad y en costo desde la sencilla y barata polinización por el viento a las complicadas y costosas estructuras dialelas para las polinizaciones controladas. Los diseños presentan grandes diferencias en cuanto a su utilidad para los diversos objetivos que se han mencionado. Libby y otros (1969) hacen un breve examen de los diseños de apareamiento y Stonecypher (1967) describe los aspectos que deben tomarse en consideración para la realización de ensayos de campo. La elección de los diseños de apareamiento y experimentales debe hacerse con sumo detenimiento, tomando en consideración los recursos y objetivos del programa de mejora y la biología de la especie.
Nunca se insistirá bastante en la importancia del empleo de un plan sistemático de apareamiento y de diseños de campo bien ejecutados en un programa de mejora. Ambos son esenciales para lograr una mejora genética aceptable.
Importancia del conocimiento de los parámetros de población
Los parámetros de población de importancia fundamental para el genetista son las variancias y covariancias genéticas, ambientales y de interacción de las características específicas (Gardner, 1963; Dudley y Moll, 1969). Si bien se han realizado grandes progresos en la mejora de los árboles forestales sin contar con una información precisa sobre los parámetros de población, su conocimiento aumenta la eficacia y facilita todos los aspectos de la planificación y ejecución de programas de mejora de árboles. Ya se ha citado la necesidad de contar con estimaciones adecuadas de los parámetros genéticos y económicos para la construcción de índices de selección fidedignos. La importancia de los efectos recíprocos entre genotipos y ambientes y la utilización de los parámetros de población para escoger entre los diferentes métodos de mejora genética se tratarán en las secciones siguientes.
Hattemer (1963) resumió las estimaciones publicadas sobre la heredabilidad en cuanto a rendimiento y otras características, pero los métodos de estimación de muchas de las razones fueron objeto de críticas por parte de Namkoong y otros (1966). Estas estimaciones y otras complementarias, basadas de nuevo principalmente en árboles jóvenes (Namkoong y otros, 1966; Stonecypher, 1966, 1969b; Stonecypher y Zobel, 1966; Squillace y otros, 1967; Eldridge, 1969; Shelbourne, 1969), sugieren que los valores de heredabilidad en cuanto a densidad de la madera, altura del árbol y diámetro pueden clasificarse por este mismo orden. La derechura del fuste tiene por lo general una heredabilidad más bien alta (Ehrenberg, 1969).
El conocimiento de la magnitud y el signo de las correlaciones genéticas entre las características que afectan al rendimiento tiene importancia para la selección. Por ejemplo, la existencia de fuertes relaciones negativas entre algunas características puede significar que es imposible la mejora simultánea de las mismas, aun cuando se utilicen índices de selección. Cuando se sabe que existen correlaciones positivas fuertes para un par de características, la selección puede concentrarse en la que sea más fácil de evaluar.
Interacción genotipo ambiente
La interacción genotipo-ambiente se refiere al diferente rendimiento de dos o más entidades genéticas cuando crecen en ambientes diferentes. Las interacciones pueden tener la amplitud suficiente para justificar la selección de estirpes distintas de árboles para ambientes específicos.
Squillace (1969) ha examinado la literatura relativa a las causas de las interacciones que afectan a especies, procedencias, progenies y clones. Los factores ambientales más importantes parecen ser la localidad, la estación y el tratamiento cultural. Entre las condiciones del cultivo que se sabe inducen interacciones importantes se cuentan la preparación de la estación, los abonos, el riego, la densidad de masa y la competencia (Donald, 1963; Giertych, 1969; Squillace, 1969). La clasificación selectiva a corto plazo de las líneas de progenies para una respuesta superior al tratamiento cultural puede no ser fidedigna, y se pueden necesitar amplios ensayos de campo (Goddard y Smith, 1969).
El medio más práctico de reducir al mínimo las pérdidas debidas a las interacciones crípticas es la utilización de una población genética de amplia base para la regeneración. Hasta que en un programa determinado de mejora genética no se conozcan las interacciones específicas genotipo-ambiente, es preferible realizar la selección en condiciones ambientales semejantes a aquellas en que se va a cultivar la progenie. Los planes de mejora deben prever el ensayo de progenies en varios ambientes que representen las principales estaciones, aplicando las prácticas de cultivo recomendadas para las plantaciones comerciales actuales y futuras.
Elección de un método de mejora genética
Un programa dinámico de mejora genética debe tener dos objetivos: la pronta producción de un material de plantación mejorado, y la obtención de nuevas ganancias en ciclos posteriores. Los diversos métodos disponibles para alcanzar esos objetivos difieren en la intensidad de la selección, el procedimiento de apareamiento, las ayudas a la selección y el tiempo transcurrido hasta que se producen semillas o estaquillas enraizadas mejoradas. Todos esos factores afectan a la ganancia anual esperada (Shelbourne, 1969). Como también varía el costo de los diferentes procedimientos de mejora genética, se necesita una base para elegir la estrategia que ofrezca los máximos beneficios en relación al costo en una situación determinada.
Si se cuenta con estimaciones satisfactorias de los parámetros de población y de los gastos de los diversos procedimientos, pueden predecirse las ganancias esperadas con el uso de diferentes procedimientos de mejora genética, y pueden compararse los métodos en cuanto a costo y beneficio (Namkoong y otros, 1966; Shelbourne, 1968). Shelbourne (1969) recomendó que para la mejora genética de árboles debe usarse el método más práctico que dé una mejora por ciclo próxima al máximo.
Al escoger un método de mejora, deben tenerse en cuenta la ganancia esperada, la biología de la especie, los recursos humanos y físicos disponibles, la escala del programa de repoblación forestal y los costos y beneficios, tanto de las fases iniciales como de las fases últimas del programa. La esperanza de obtener ganancias de bastante consideración con programas de mejora eficientes (Cuadro 4) justifica la realización de trabajos intensivos de mejora, pero, para orientar la elección del método de mejora en una serie de situaciones, se precisan estudios críticos complementarios, como los de Davis (1967, 1969), de la economía de los múltiples procedimientos diferentes de mejora genética.
DEFINICIONES, OBJETIVOS Y EJEMPLOS
La hibridación de árboles forestales significa el cruzamiento de entidades genéticamente diferentes, tales como especies, variedades, procedencias o estirpes selectas. Es raro que implique el cruzamiento de líneas intracruzadas de especies de polinización cruzada, debido a dificultades de producción (Keiding, 1968; Snyder, 1968; Orr-Ewing, 1969). En el término general de «híbridos», se incluyen los descendientes de los híbridos F1 (los F2 y retrocruzamientos).
Dos objetivos generales de la hibridación de árboles forestales son la combinación de las características valiosas de dos o más poblaciones, y la obtención de heterosis o vigor híbrido (Duffield y Snyder, 1958).
La hibridación combinativa es especialmente útil en la mejora de la resistencia a las plagas y otras características múltiples y en el desarrollo de árboles destinados a estaciones a las cuales no se adapte adecuadamente una especie pura. Buenos ejemplos de ello son los híbridos Thuja plicata Donn. x T. standishii Carr. resistentes a la mancha foliar, y Populus alba L. x P. grandidentata Michx. bien formados y de fácil radicación (Duffield y Snyder, 1958). Recientemente se ha señalado la presencia de varios híbridos de Pinus, prometedores de combinaciones de un rendimiento superior con otras características deseables (Cuadro 5).
La heterosis se pone de manifiesto cuando el rendimiento medio de un híbrido excede al de sus progenitores. Depende de la diferencia en las frecuencias génicas entre las dos poblaciones cruzadas y del nivel de dominancia de los aleles en los loci que afectan a las características de que se trata (Falconer, 1960). El vigor híbrido se manifiesta a menudo en el cruzamiento Larix leptolepis Gord. x L. decidua Mill. (Wright, 1962; Rohmeder, 1963), en muchos híbridos de chopo (Wright, 1964) y en otras muchas combinaciones interespecíficas (Wright, 1962).
La hibridación inter-racial se ha intentado con menos frecuencia que los cruzamientos entre especies, con muestras alentadoras, pero no definitivas, de resultados beneficiosos (Nilsson, 1963; Orr-Ewing, 1966; Woessner, 1968). Pueden esperarse, sin embargo, ganancias por medio de la heterosis y de los efectos combinativos, sobre todo en las especies de amplia esfera de difusión que cubren una gran diversidad da habitat.
Algunas veces las progenies retrocruzadas son más prometedoras que los híbridos F1 debido a su buen rendimiento y a la mayor facilidad para la producción de semilla comercial (Libby, 1958; Slee, 1969). Cabe esperar que los híbridos de generación avanzada formen la base de futuras estirpes mejoradas de árboles forestales.
CUADRO 5. - EJEMPLOS DE HÍBRIDOS INTERESPECÍFICOS PROMETEDORES DE Pinus, SEÑALADOS DESPUÉS DE LAS PUBLICACIONES DE DUFFIELD Y SNYDER (1958) Y WRIGHT (1962)
|
Referencia |
Especie |
Características aportadas |
Observaciones |
|
Jewell (1966) |
P. taeda L. |
Indice de crecimiento rápido |
Prometedores para las estaciones de P. taeda con fuerte incidencia de enfermedades |
|
P. echinata Mill. |
Resistencia a Cronartium fusiforme |
||
|
Derr (1966) |
P. palustris Mill. |
Buena forma del fuste y resistencia a C. fusiforme |
Prometedores para las estaciones de pinos con gran incidencia de ambas enfermedades |
|
P. elliottii Engelm. |
Indice de crecimiento rápido y resistencia a Scirrhia acicola |
||
|
Griffin y Conkle (1967) |
P. attenuata Lemm. |
Resistencia a la sequía y a la helada |
El vigor híbrido se manifiesta en algunas estaciones |
|
P. radiata Don. |
Crecimiento rápido |
||
|
Hyun (1969) |
P. taeda L. |
Buena forma del fuste e índice de crecimiento rápido |
Son prometedores los retrocumientos a P. rigida y F2 producidos por la polinización natural de árboles: F1 |
|
P. rigida Mill. |
Resistencia a la helada |
||
|
Slee (1969) |
P. elliottii Engelm. |
Buena forma del fuste y adaptabilidad a las estaciones húmedas |
Los retrocruzamientos y las progenies F1 y F2 tienen un alto valor potencial |
|
P. caribaea Mor. |
Indice de crecimiento rápido |
Los híbridos F1, de 11 años, de Pinus elliottii Engelm. x P. caribaea Mor. son superiores en rendimiento económico neto a P. elliottii y P. caribaea en Beerwah, Queensland (Australia), sólo cuando se los cultiva en estaciones pantanosas.
FOTO: NIKLES
GANANCIAS CONSEGUIDAS POR MEDIO DE LA HIBRIDACIÓN
Los híbridos F1 suelen dar muy buenos resultados en una gran diversidad de ambientes, debido probablemente a su composición genética muy heterocigótica (Allard y Bradshaw, 1964). Sin embargo, cuando se miden de modo apropiado las ganancias con relación al rendimiento de otras mejores especies puras disponibles para una estación determinada, se ve muchas veces que la superioridad de los híbridos se manifiesta sólo en los ambientes donde ninguno de los progenitores está bien adaptado para un alto rendimiento. Estos dos aspectos se demuestran por el hecho de que los híbridos F1 de 9 años de P. elliottii y P. caribaea crecen sumamente bien tanto en las estaciones pantanosas como en las calinosas en Beerwah, Queensland, pero su superioridad con respecto a las mejores especies progenitores en la estación pantanosa desfavorable es mucho mayor que en la estación calinosa donde prosperan ambas especies progenitores (Slee, 1969).
Son escasas las estimaciones cuantitativas fidedignas de las ganancias que pueden obtenerse por hibridación, bien porque los ensayos de híbridos no se regulen siempre de modo satisfactorio, o bien porque las progenies sean aún demasiado jóvenes para la evaluación. No obstante, existen claros indicios de que los híbridos F1 o sus descendientes pueden rendir ganancias apreciables en determinadas circunstancias. Por ejemplo, los híbridos F1 de 9 años de P. elliottii y P. caribaea descritos por Slee (1969) superan a P. caribaea en altura en 21 por ciento y en diámetro en 20 por ciento (y superan con un margen aún mayor a P. elliottii comparable), cuando crecen en una estación pantanosa. La ganancia probable en el rendimiento económico neto de los híbridos a la edad de 11 años se realzará por un índice de aprovechamiento neto mayor debido a la mejor calidad del fuste. Recientes resultados de estudios en árboles de 7 años (Nikles, 1969) indican que algunas progenies retrocruzadas habitantes en lugares pantanosos deben dar ganancias iguales al menos a las de los híbridos F1. Como los retrocruzamientos son más fáciles de producir que los F1, es más probable que se los pueda aprovechar comercialmente.
PROBLEMAS DE HIBRIDACIÓN
Hay diversos informes sobre variaciones en el rendimiento de los híbridos según el origen geográfico de las poblaciones progenitores y de los árboles apareados (Nilsson, 1963; Lotan, 1967; Hyun, 1969). La selección fenotípica entre las poblaciones que son objeto de hibridación y dentro de ellas puede ser efectiva en cuanto a las características de alta heredabilidad, pero es conveniente una selección y eliminación a fondo en lo que respecta a la aptitud combinativa de los progenitores para las características que presentan escasa variancia genética aditiva. La prueba del rendimiento y la asignación de híbridos a estaciones adecuadas son problemas que exigen observaciones prolongadas.
La producción en masa de árboles híbridos es sencilla en el caso de aquellos cuya propagación vegetativa puede efectuarse con facilidad. Sin embargo, por lo general, los problemas de la época de floración de los progenitores, la poca capacidad de producción de semillas y la escasa viabilidad de las semillas de los híbridos F1, contribuyen a que la producción en masa de semillas resulte muy cara. Aunque la semilla de retrocruzamientos y de híbridos F2 suele ser sumamente fértil y se produce más fácilmente por polinización natural, el rendimiento de estos descendientes híbridos debe ser puesto a prueba extensamente para demostrar su superioridad antes de que pueda justificarse su utilización comercial.
Como el material de plantación híbrido es casi siempre más caro que el de una especie pura, es necesario que los híbridos sean capaces de incremento del rendimiento económico bastante mayores que los que pueden obtenerse por mejora de la población. Ese material rinde el máximo beneficio cuando se puede producir en masa económicamente, cuando tiene un rendimiento superior al del progenitor que se quiere sustituir, y cuando es superior a cualquier otra especie pura. Debido a estos diversos requisitos de los híbridos, su utilización es bastante poco frecuente y sé los destina a usos especializados.
Cada vez se aprecian más las contribuciones independientes que pueden prestar a la mejora del rendimiento la genética vegetal, la preparación intensiva de la estación, la fertilización, el deshierbe, la atención al espaciamiento inicial y a los aclaramientos periódicos, la protección, las normas de aprovechamiento y la planificación de la explotación. Sin embargo, los mayores beneficios de las inversiones dedicadas a mejorar la cantidad y calidad del rendimiento económico de los bosques, deben proceder de un programa integrado con enfoque genético, ambiental y administrativo. Las mayores posibilidades de aumentar el índice de la producción forestal están en una combinación de esos diversos enfoques.
ALLARD, R. W. & BRADSHAW, A. D. 1964. Implications of genotype-environmental interactions in applied plant breeding. Crop Sci., 4: 503-508.
BARBER, J. C. 1964. Inherent variation among slash pine progenies at the Ida Cason Calloway Foundation. U.S. Forest Serv., Res. Pap. SE-10. 90 p.
BARRETT, W. H. G. 1969. Coníferas de rápido crecimiento para la República Argentina. Segunda Consulta Mundial - sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-2/2
BINGHAM, R. T. 1968. Breeding blister rust resistant western white pine. IV. Mixed pollen crosses for appraisal of general Combining ability. Silvae Genet., 17: 133-138.
BLACK, J. N. 1966. The utilisation of solar energy by forests. En Physiology in forestry, ed. por R.W.V. Palmer. Forestry, suplemento 1966, p. 98-109.
BOLOTIN, M. 1969. Selection for drought resistance in woody species. Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-6/3.
BRAATEN, M. O. 1965. The union of partial diallel mating designs and incomplete block environmental designs. Consolidated Univ. of North Carolina, Institute of Statistics. Mimeog. Series N° 432. 77 p.
BROWN, A. G. 1969. Comunicación personal. Canberra, Forest Research Institute.
BURDON, R. D. 1968. Preliminary findings from a clonal replication trial. New Zealand Forest Research Institute. Silvicultural Rept 98. 18 p.
BURLEY, J., BURROWS, P. M., ARMITAGE, F. B. & BARNES, R. D. 1966. Progeny test designs for Pinus patula in Rhodesia. Silvae Genet., 15(5-6): 166-173.
CALLAHAM, R. Z. 1964. Investigación de procedencias: estudio de la diversidad genética asociada a la geografía. Unasylva, 18(2-3): 40-50.
CAMPBELL, R. K. 1964. Plus tree selection. Weyerhauser Working Plan 15-2. Centralia, Washington.
COOPER, J. P. 1963. Species and population differences in climatic response. En Environmental control of plant growth, ed. por L. T. Evans. New York, Academic Press, p. 381-403.
DAVIS, L. S. 1967. Cost-return relationships of tree improvement programs. Proc. 9th South. Conf. Forest Tree Improvement, p. 20-26.
DAVIS, L. S. 1969. Economic models for program evaluation. Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-13/2.
DAY, W. R. & PEACE, T. R. 1934. The experimental production and the diagnosis of frost injury on forest trees. Oxford Forest Mem. 16. 60 p.
DERR, H. J. 1966. Longleaf x slash hybrids at age 7: survival, growth, and disease susceptibility. J. For., 64(4): 236-239.
DIETRICHSON, J. 1969. Growth rhythm and yield as related to provenance, progeny and environment. Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-2/3.
DONALD, C. M. 1962. In search of yield. J. Austr. Inst. agric. Sci., 28(3): 171-178.
DONALD, C. M. 1963. Competition among crop and pasture plants. Adv. Agron., 15: 1-118.
DUDLEY, J. W. & MOLL, R. H. 1969. Interpretation and use of estimates of heritability and genetic variances in plant breeding. Crop Sci., 9: 257-262.
DUFFIELD, J. W. & SNYDER, E. B. 1958. Benefits from hybridizing American forest trees. J. For., 56(11): 809-815.
EDWARDS, M. V. 1963. The use of exotic forest trees in increasing production with particular reference to north-western Europe. FAO/FORGEN 63-4/1.
EHRENBERG, C. E. 1966. Parent-progeny relationships in scots pine (Pinus sylvestris L.). Stud. Forest. Suec., 40:1-52.
EHRENBERG, C. E. 1969. Breeding for stem quality. Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-3/1.
ELDRIDGE, K. G. 1966. Genetic improvement of Eucalyptus regnans by selection of parent trees. Appita, 19(6): 133-138.
ELDRIDGE, K. G. 1969. Variation in Eucalyptus regnans. Tesis, Dept. of Forestry, Austr. Nat. Univ., Canberra. (Inédito)
FALCONER, D. S. 1960. Introduction to quantitative genetics. Edinburgh, Oliver and Boyd.
FAO. 1969. Informe de la primera reunión del Cuadro de Expertos de la FAO en Recursos Genéticos Forestales Roma, 1968. Roma. FO: FGR/1/Report.
FIELDING, J. M. & BROWN, A. G. 1961. Tree-to-tree variations in the health and some effects of superphosphate on the growth and development of Monterey pine on a low-quality site. Australia, Commonwealth Forestry and Timb. Bur. Leaflet N° 79. 19 p.
FORSHELL, W. P. 1964. La genética en la práctica forestal en Suecia. Unasylva, 18(2-3): 119-127.
GARDNER, C. O. 1963. Estimates of genetic parameters in cross-fertilising plante and the implications in plant breeding. En Statistical genetics and plant breeding. National Research Council, Publication N° 982, p. 225-252.
GERHOLD, H. D., SCHREINER, E. J., DERMOTT, R. C. & WINIESKI, J. A. eds. 1966. Breeding pest-resistant trees. London, Pergamon Press.
GIERTYGH, M. M. 1969. Growth as related to nutrition and competition. Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-2/4.
GODDARD, R. E. 1965. Manual of progeny test procedures recommended for use in University of Florida Cooperative Forest Genetics Research Program. Part 1. Gainesville, Fla. 15 p. (Mimeografiado)
GODDARD, R. E. 1968. Comunicación personal. Gainesville, Fla., Univ. of Florida.
GODDARD, R. E. & SMITH, W. H. 1969. Progeny testing for intensive management. Proc. 10th South. Conf. Forest Tree Improvement, p. 76-83.
GRANT, V. 1963. The origin of adaptations. New York, Columbia University Press.
GRIFFIN, J. R. & CONKLE, M. T. 1967. Early performance of knobcone x Monterey pine hybrids - on marginal timber sites. U. S. Forest Serv. Res. Note PSW-156. 10 p.
HAGMAN, M. 1969. Research programmes. Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-8/1.
HALEY, C. 1960. Progress in the application of tree breeding in our planted forests. Austr. Forest., 24(1): 8-18.
HATTEMER, H. H. 1963. Estimates of heritability published in forest tree breeding research. FAO/FORGEN 63-2a/3.
HESLOP-HARRISON, J. 1964. Forty years of genecology. Adv. ecolog. Res., 2: 159-240.
HOPKINS, E. R. 1960. Variation in the growth rate and quality of Pinus pinaster Ait., in Western Australia. West Austr. Forests Dept. Bull. 67. 33 p.
HYUN, S. K. 1969. The growth performance of pitch-loblolly hybrid pine produced by different geographic races of loblolly pine in their early age. Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-7/7.
ILLY, G. 1969. Les indices de sélection, exemple du pin maritime. Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-7/3.
IYAMABO, D. 1969. Growth and species introduction in African Savannah. Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-2/6.
JACKSON, D. S. 1965. Species siting: climate, soil, and productivity. N.Z.J. For., 10(1): 90-102.
JACOBS, M. R. 1964. The use of exotic forest trees. Austr. For., 28(3): 150-156.
JEWELL, F. F. 1966. Inheritance of rust resistance in southern pines. En Breeding pest-resistant trees, p. 107-109. London, Pergamon Press.
JOHNSSON, H. 1964. Forest tree breeding by selection. Silvae Genet., 13(1-2): 41-48.
KEIDING, H. 1968. Preliminary investigations of inbreeding and outcrossing in larch. Silvae Genet., 17: 159-164.
KOZLOWSKI, T. T., 1962. ed. Tree growth. New York, Ronald Press.
KRAUS, J. 1968. Comunicación personal. Macon, Ga., Southeast Forest Expt Station.
LANGLET, O. 1963. Practical results and current problems in provenance research in Sweden. FAO/FORGEN 63-3/1.
LANGLET, O. 1967. Regional intra-specific variousness. Proc. 14th IUFRO Congress, 3: 435-458.
LARSEN, E. V. Seed procurement for research. Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-8/8.
LEUCHARS, D. 1965. The planning and practice of trials of exotic species. East African agric. for J., 31(1): 83-90.
LIBBY, W. J. 1958. The backcross hybrid Jeffrey x (Jeffrey x Coulter) pine. J. For., 56(11): 840-842.
LIBBY, W. J. 1964. Clonal selection, and an alternative seed orchard scheme. Silvae Genet., 13(1-2): 32-40.
LIBBY, W. J. 1968. Mating designs for second-generation selection in forest trees. Abstracts of presented papers, 11th annual meeting of the Western Forest Genetics Association, Corvallis, Ore., 1968.
LIBBY, W. J. 1969. Some possibilities of the clone in forest genetics research. Genetics Lectures. Vol. 1: 121-136. Genetics Institute, Oregon State Univ., Corvallis, Oregon.
LIBBY, W. J., STETTLER, R. F. & SEITZ, F. W. 1969. Forest Genetics and forest tree breeding. Ann. Rev. Genetics, 3.
LINES, R. 1965. Provenance and the supply of forest tree seed. Forestry, 59(1): 7-15.
LINES, R. 1967a. The planning and conduct of provenance experiments. Simposio Mundial de la FAO sobre Bosques Artificiales y su Importancia Industrial. FAO-FO/MMF: 67-4b/4.
LINES, R. comp. 1967b. Standardisation of methods for provenance research and testing. Proc. 14th IUFRO Congress, 3: 672-713.
LOTAN, J. E. 1967. Early performance of Pinus conforta x banksiana hybrids. U.S. Forest. Serv., Res. Note. INT-57. 8 p.
MARSH, E. K. 1969. Selecting adapted races of introduced species. Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-10/4.
MERGEN, F. 1962. Selection of superior forest trees. En Tree growth. ed. by T. T. Kozolwski. New York, Ronald Press, p. 327-344.
MORANDINI, R. 1967. Planning of species and provenance trials. Simposio Mundial de la FAO sobre Bosques Artificiales y su Importancia Industrial FAO-FO-/MMF: 67-4b/1.
MORGENSTERN, E. K. & ROCHE, L. 1969. Using concepts of selection to delimit seed zones. Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-2/16.
NAMKOONG, G. 1966. Family indices for seed-orchard selection. U.S. Forest Serv. Res. Pap. NC-6. 6 p.
NAMKOONG, G. 1969. Problems of multiple-trait breeding. Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-7/4.
NAMKOONG, G., SNYDER, E. B. & STONECYPHER, R. W. 1966. Heritability and gain concepts for evaluating breeding systems such as seedling orchards. Silvae Genet., 15(3): 76-84.
NICIPOROVIC, A. A. 1954. Fifteenth Timirjazev Lecture, U.S.S.R. Acad. Sci. (Resumen por Black, J. N. y Watson D. J. en Field Crop. Abstr., 13: 169, 1960).
NIENSTAEDT, H. & KING, J. P. 1969. Breeding for delayed bud-break in Picea glauca (Moench) Voss: potential frost avoidance and growth gains. Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-2/5.
NIKLES, D. G. 1966. Progeny tests of slash pine (Pinus elliottii Engelm.) in Queensland, Australia. Proc. 8th South. Conf. Forest Tree Improvement, p. 112-120.
NIKLES, D. G. 1969. Informes sobre investigaciones. Archivos de la Forest Research Station, Beerwah, Queensland.
NIKLES, D. G. & SMITH, W. J. 1969. Increased volume and dry-weight yield and improved stem straightness obtained through selection within a population of Pinus elliottii Engelm. Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-2/20.
NILSSON, B. 1963. Intraspecific hybridisation and heterosis within Picea abies. FAO-FORGEN 63-2b/6.
NORTHCOTT, P. L. 1965. The comparison of tree qualities. For. Chron., 41(2): 215-221.
ORR-EWING, A. L. 1966. Inter-and intraspecific crosses in Doug las fir, Pseudotsuga menziessii (Mirb.) Franco. Silvae Genet., 15(4): 121-126.
ORR-EWING, A. L. 1967. A progeny test of Douglas-fir to demonstrate the importance of selection in forest practice, Brit. Columbia Forest Serv., Res. Note 43, 23 p.
ORR-EWING, A. L. 1969. Inbreeding to the S- generation in Douglas-fir. Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-8/6.
OVINGTON, J. D. 1965. Organic production turnover and mineral cycling in woodlands. Biol. Rev., 40: 295-336.
OVINGTON, J. D. 1968. Considerations of forest productivity. Austr. For., 32(4): 191-204.
PAWSEY, C. K. 1968. Comunicación personal. Mount Burr, S. Austr., Forest Research Institute.
PEDERICK, L. A. 1968. Selection criteria - methods and records. Proc. Austr. Forest Res. Working Group N° 1, Canberra. Item 6.
ROBERDS, J. H., ZOBEL, B. J. & KELLISON R. C. 1967. Progeny testing in the N. C. State University-Industry tree improvement program. Proc. 14th IUFRO Congress, 3: 387-394.
ROCHE, D. 1968. The value of short term studies in Provenance research. Commonw. for Rev., 47(1): 14-26.
ROHMEDER, E. 1963. Experiments on forest tree hybrids in Bavaria from 1936 to 1962. FAO/FORGEN 63-2b/1.
SARVAS, R. 1969. Genetical adaptation of forest trees to the heat factor of the climate. Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-2/15.
SCHUTZ, W. M. & COCKERHAM, C. C. 1966. The effect of field blocking on gain from selection. Biometrics, 22(4): 843-863.
SHELBOURNE, C. J. A. 1968. Tree breeding methods. New Zealand Forest Research Institute. Tree Impr. Rept. 39. 31 p.
SHELBOURNE, C. J. A. 1969. Predicted genetic improvement from different breeding methods. Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-8/16.
SHEPHERD, K. R. & SLEE, M. U. 1969. Tree breeding programmes and silvicultural practice in Australian man-made forests. Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-9/9.
SLEE, M. U. 1969. Height and girth growth of slash - Caribbean hybrids in southern Queensland. Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-2/10.
SLEE, M. U. & NIKLES, D. G. 1968. Variability of Pinus caribaea Morelet in young Queensland plantations. Ninth Brit. Commonw. Forestry Conf. Documento presentado por el Dept of Forestry, Brisbane. 50 p.
SNYDER, E. B. 1968. Seed yield and nursery performance of selfpollinated slash pines. For. Sci., 14: 68-74.
SNYDER, E. B. 1969. Parental selection versus half-sib family selection of longleaf pine. Proc. 10th South. Conf. Forest Tree Improvement, p. 84-88.
SQUILLACE, A. E. 1969. Field experiences of the kinds and sizes of genotype-environment interaction. Documento presentado a un Working Group on Quantitative Genetics, Workshop on Forest Tree Breeding, IUFRO, Section 22. 23 p. (Mimeografiado)
SQUILLACE, A. E. & SILEN, R. R. 1962. Racial variation in ponderosa pine. For. Sci. Monog., 2: 1-27.
SQUILLACE, A. E., BINGHAM, R. T., NAMKOONG, G. & ROBINSON, H. F. 1967. Heritability of juvenile growth rate and expected gain from selection in western white pine. Silvae Genet., 16(1): 1-6.
STERN, K. & HATTEMER, H. H. 1964. Problems involved in some models of selection in forest tree breeding. Silvae Genet., 13(1-2): 27-31.
STONECYPHER, R. W. 1966. The loblolly pine heritability study. Internat. Paper Co. Southlands Exp. Forest. Tech. Bull. 5. 128 p.
STONECYPHER, R. W. 1967. Field testing and data handling in forest genetics research. Proc. 9th South. Conf. Forest Tree Improvement, p. 112-120.
STONECYPHER, R. W. 1969a. The whole tree concept. Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-7/1.
STONECYPHER, R. W. 1969b. Comunicación personal. Bainbridge, Georgia, Internat. Paper Co.
STONECYPHER, R. W. & ZOBEL, B. J. 1966. Inheritance of specific gravity in five-year-old seedlings of loblolly pine. Tappi, 49(7): 303-305.
THULIN, I. J. 1969. Breeding of Pinus radiata through seed improvement and clonal afforestation. Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-9/5.
TODA, R. 1969. Progeny evaluation problems - Cryptomeria. Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol. FAO-FO-FTB-69-7/5.
VAN BUIJTENEN, J. P. 1966a. Comunicación personal. College Station, Texas. Texas Forest Service.
VAN BUIJTENEN, J. P. 1966b. Testing loblolly pines (Pinus taeda) for drought resistance. Texas Forest Service. Tech. Rept 13. 15 p.
VAN BUIJTENEN, J. P. 1968. Sixteenth progress report of the Co-operative Forest Tree Improvement Program. Texas Forest Service. Circular 112. 11 p.
WAREING, P. F. 1966. The physiologists approach to tree growth. En Physiology in forestry, ed. by R.W.V. Palmer. Forestry, suplemento 1966, p. 7-18.
WEBB, C. D. & BARBER, J. C. 1966. Selection in slash pine brings marked improvement in diameter and height growth plus rust resistance. Proc. 8th South. Conf. Forest Tree Improvement, p. 67-72.
WELLS, O. O. 1969. Results of the south-wide pine seed source study through 1968-69. Proc. 10th South. Conf. forest Tree Improvement, p. 117-129.
WELLS, O. O. & WAKELEY, P. C. 1966. Geographic variation in survival, growth, and fusiform-rust infection of planted loblolly pine. For. Sci. Monog., 11: 1-40.
WOESSNER, R. A. 1965. Growth, form and disease resistance in four-year-old control and five-year-old open-pollinated progeny of loblolly pine selected for use in seed orchards. Supplement. North Carolina State Univ., School of Forestry. 34 p.
WOESSNER, R. A. 1968. A juvenile assessment of wide crosses of loblolly pine select trees indigenous to different geographical areas. North Carolina State Univ., School of Forest Resources. (Tesis).
WOOD, R. S. 1962. Some influences on practice in the espacement, thinning and pruning of coniferous plantations. Eighth Brit. Commonwealth Forest. Conf. Documento presentado por la Forestry Comm., Londres. 8 p.
WRIGHT, J. W. 1964. Mejoramiento genético de árboles forestales. Roma. FAO: Estudios de Silvicultura y Productos Forestales N° 16.
WRIGHT, J. W. 1964. Hibridación entre especies y razas. Unasylva, 18(2-3): 30-39.
ZOBEL, B. J. 1969. Best adapted seed sources. FAO-North Carolina State Forest Tree Improvement Training Centre, p. 60-62. School of Forest Resources, N.C. State Univ.
ZOBEL, B. J., KELLISON, R. & MATTHIAS, MARTHA. 1969. Genetic improvement in forest trees - growth rate and wood characteristics in young loblolly pine. Proc. 10th South. Conf. Forest Tree Improvement, p. 59-75.
|
REPERTORIO MUNDIAL DE GENETISTAS FORESTALES Y CRIADORES DE ARBOLES Una de las actividades relacionadas con la Segunda Consulta Mundial sobre Genética Forestal y Mejora del Arbol consistió en la preparación de un Repertorio Mundial de Genetistas Forestales y Criadores de Arboles puesto al día. En 1960 y 1962 la Sociedad de Forestales Americanos había publicado en el Journal of Forestry listas de genetistas forestales y criadores de árboles que eran muy utilizadas por organismos internacionales tales como la FAO y la IUFRO, institutos de investigación del mundo entero e investigadores. Hacia 1967 se hacía sentir claramente la necesidad de listas corregidas y aumentadas y por tal motivo el Comité de Mejora del Arbol de dicha Sociedad comenzó a preparar un repertorio nuevo, de cuya redacción se encargó el Dr. Hans Nienstaedt, del Instituto de Genética Forestal, Star Route 2, Rhinelander, Wisconsin 54501, miembro del Comité de Mejora del Arbol. A principios de 1968 se enviaron por correo más de 1 400 cuestionarios y en la Consulta de Wáshington se puso a disposición de los interesados el proyecto del nuevo repertorio para que lo revisaran. De resultas de ello se recibieron muchas correcciones y más de 300 nuevos nombres. La versión final contiene 1 134 nombres en orden alfabético, por países, con la dirección y las materias que les interesan en particular y en general, a lo cual se agrega: un índice alfabético de nombres, uno de materias y otro de géneros. Se espera publicar este Repertorio como suplemento de las actas de la Consulta. |
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}