![]()
![]()
![]()
The hydrography of the survey area was observed in sections across the shelf, extending just beyond the edge. The surface layer temperature was continuously recorded along the cruise track by a sea-surface thermograph.
The temperature distribution as observed in the surface layer during the cruises in May-, Sept- and Dec-81 and Feb to Mar- 82, is shown respectively in Figures 12–15. These temperature charts agree well with the general seasonal fluctuations of the hydrographic regime in the area as described in Troadec and Garcia (1980).
During the cruise in May-81 the area north of Cape Verde was still characterized by northern winter. Temperatures below 20°C are indicative of upwelling in this area. The frontal zone to the south of the upwelling area was observed south of Cape Verde, but was not very well defined. The temperature distribution between Cape Verde and Cape Verga was rather complex, possibly in connection with the transition period between northern winter and summer and the associated northward shift of the front between tropical surface water and upwelled water.
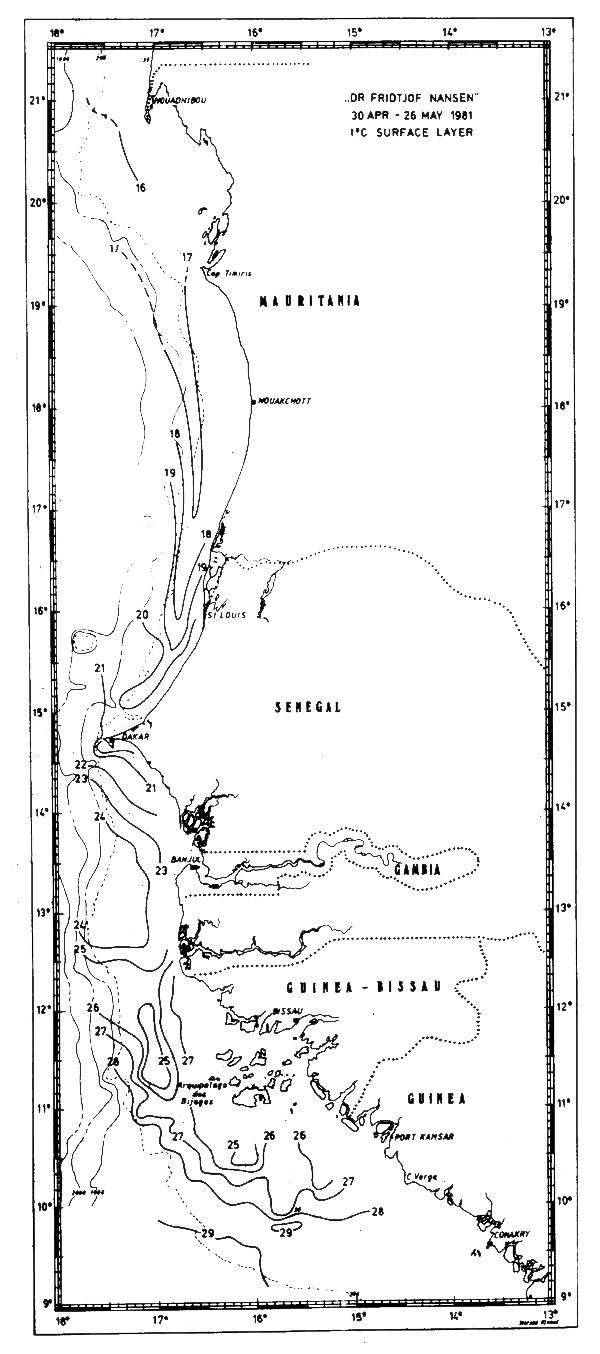
Fig. 12 Temperature in surface layer, Cape Blanc to Cape Verga, Apr 30 – May 26, 1981.
In September this front was observed off Cape Timiris where the temperatures in the surface layer increased from about 21°C to above 28°C (Fig. 13a). South of the front the surface layer had tropical characteristics and there was no evidence of upwelling.
In December the shelf area off Mauritania and Senegal (Fig. 14) was characterized by homogeneous surface temperatures close to 24°C. Indications of upwelling were observed only in a small area near Cape Blanc where temperatures were below 21°C. South of this area there was no evidence of upwelling. According to the general seasonal trends the frontal zone is located off the coast of Guinea in this season. In agreement with this no clear front was observed in the survey area.
In Feb-Mar-82 the characteristics of northern winter were dominant and upwelling obviously occurred along the coast north of about 13°N. The surface temperatures were decreasing towards the coast with inshore temperatures generally below 17°C. Just south of Cape Verde there was an area with temperatures below 15°C (Fig. 15a) and north of Cape Verde extensive areas with temperatures below 16°C were found along the coasts of Mauritania, Western Sahara and Morocco (Figs 15b-d).
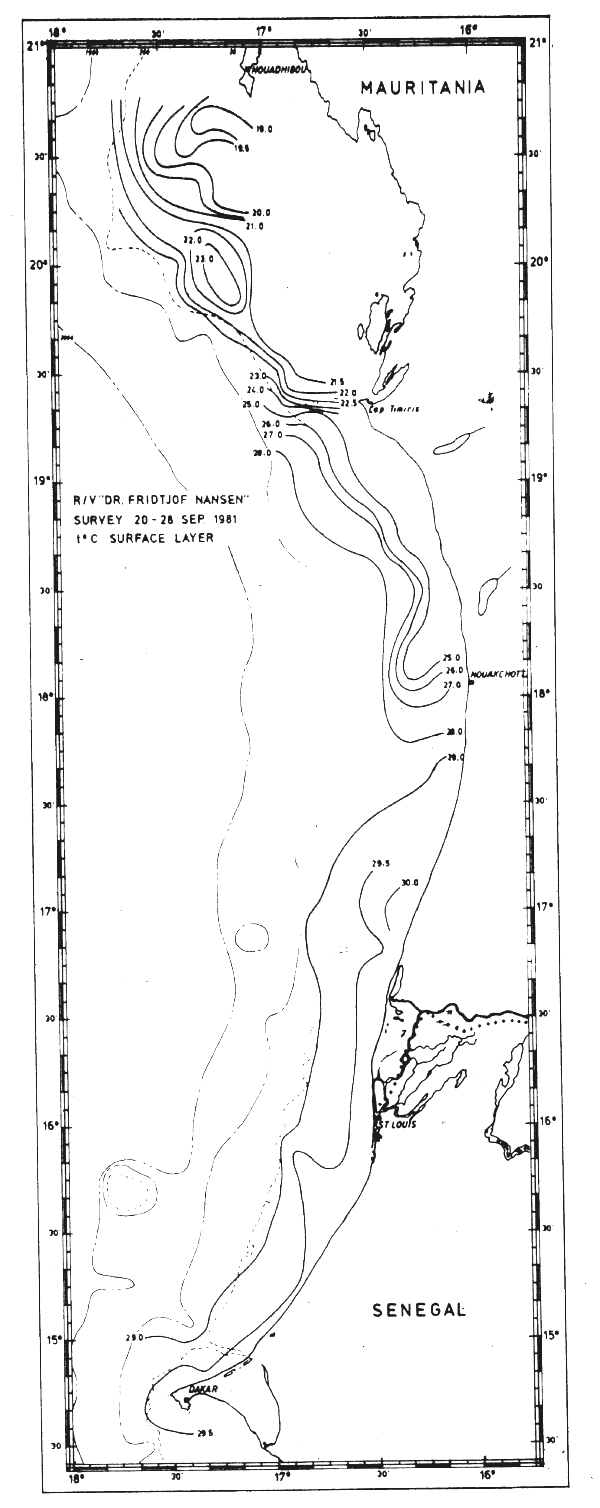
Fig. 13a Temperature in surface layer, Cape Blanc to Cape Verde, Sept 20–28, 1981.
South of the Bissagos Islands tropical surface waters with temperatures above 24°C covered most of the shelf area. The tongue of isotherms in the frontal zone south of the upwelling area was possibly associated with a southward drift of the upwelled waters and a northward current off the edge (Mittelstaedt, 1982). In May-81 a similar pattern had been observed just north of Cape Verde (Fig. 12) probably indicating that the upwelling area was dwindling at this time.
The sections covering the Cape Blanc - Cape Verde area in May-81 are shown in Fig. 16. In general the isolines tilted upwards towards the shore. In the two northernmost sections there was evidence of upwelling, but in the sections further south this structure could rather be ascribed to the southward current. South of Cape Verde (Fig. 17) a well defined thermocline was found with no indication of upwelling.
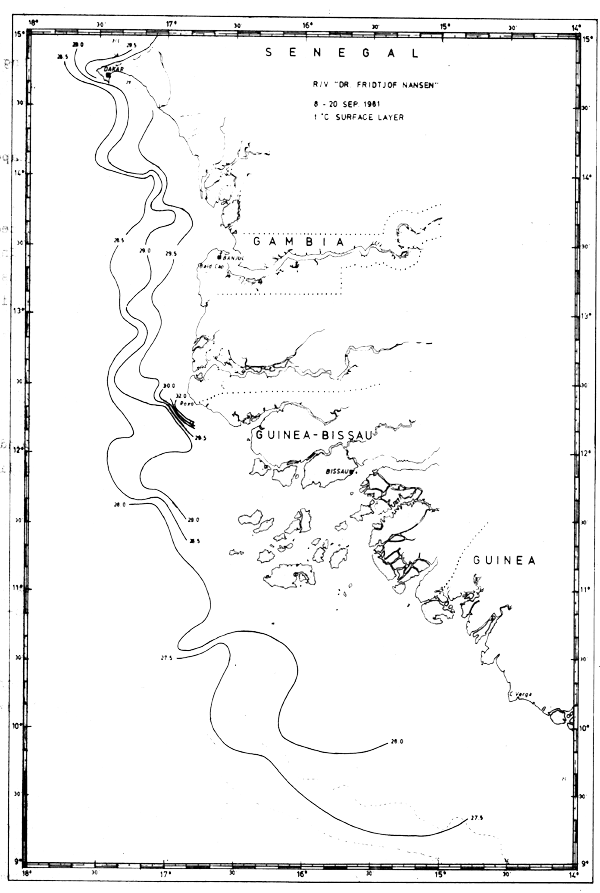
Fig. 13b Temperature in surface layer, Cape Verde to Cape Verga, Sept 8–20, 1981.
In Sept-81 the section off Cape Blanc revealed clear indications of upwelling (Fig. 18). However, further south a well-defined thermocline between 25 and 50 m depth indicated that no upwelling occurred. The sections worked south of Cape Verde are shown in Fig. 19.
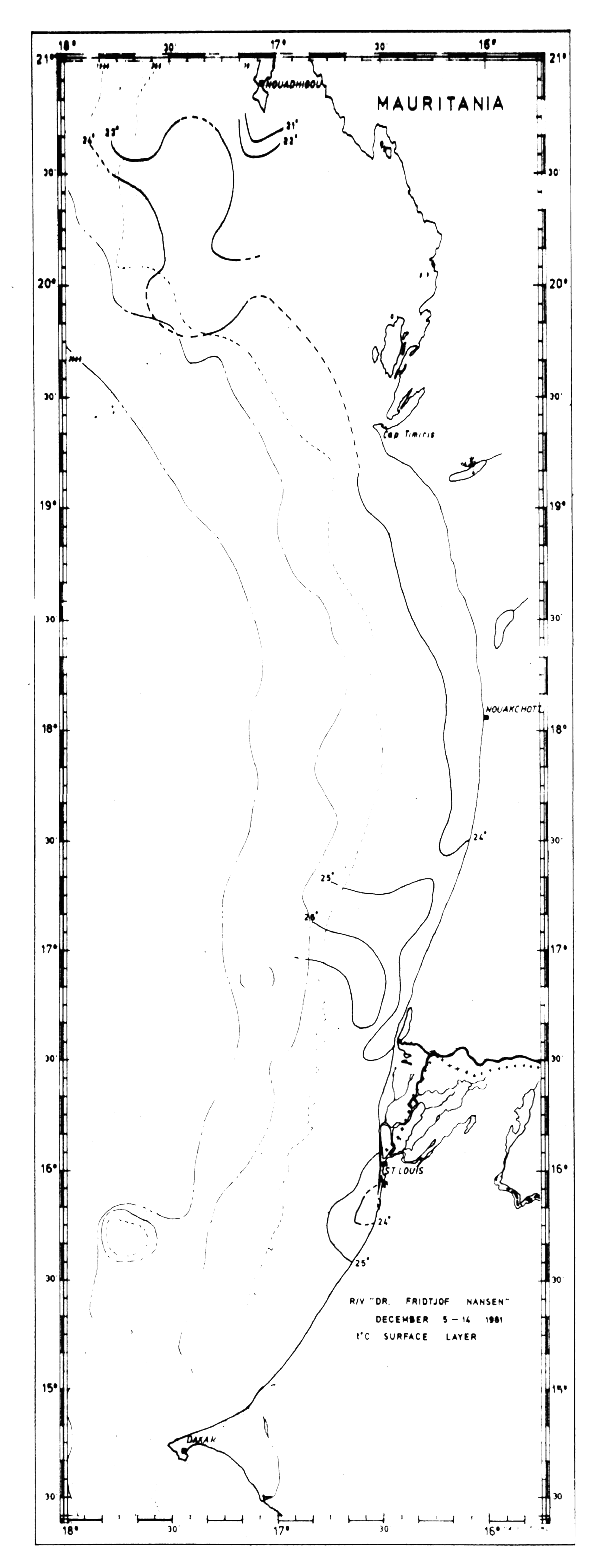
Fig. 14 Temperature in surface layer, Cape Blanc to Cape Verde, Dec 5–14, 1981.
The four sections worked between Cape Blanc and Cape Verde in Dec-81 are shown in Fig. 20. Moderate upwelling is possibly indicated in the two northern sections and the thermocline was observed at somewhat greater depth than in May and Sept-81.
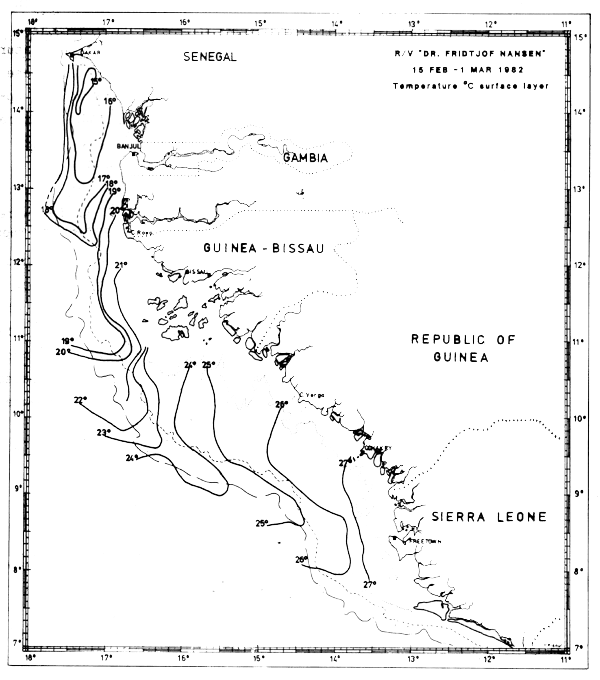
Fig. 15a Temperature in surface layer, Freetown to Cape Verde, Feb 15 - Mar 1, 1982.
Four sections worked between Cape Blanc and Cape Verde in Mar-82 are shown in Fig. 21. The distribution of temperature, salinity and density in these sections clearly showed an upwelling situation. The temperatures in near surface layers were between 18° and 15°C, decreasing towards the shore. The associated salinities were between 35.6 and 36.0°/oo. These characteristics indicate that South Atlantic central water was an important component of the upwelling water masses. T-S relationships for the various sections (not shown in figures) revealed a northward decrease in the admixture of South Atlantic central water and a correspondingly increased admixture of North Atlantic central water.
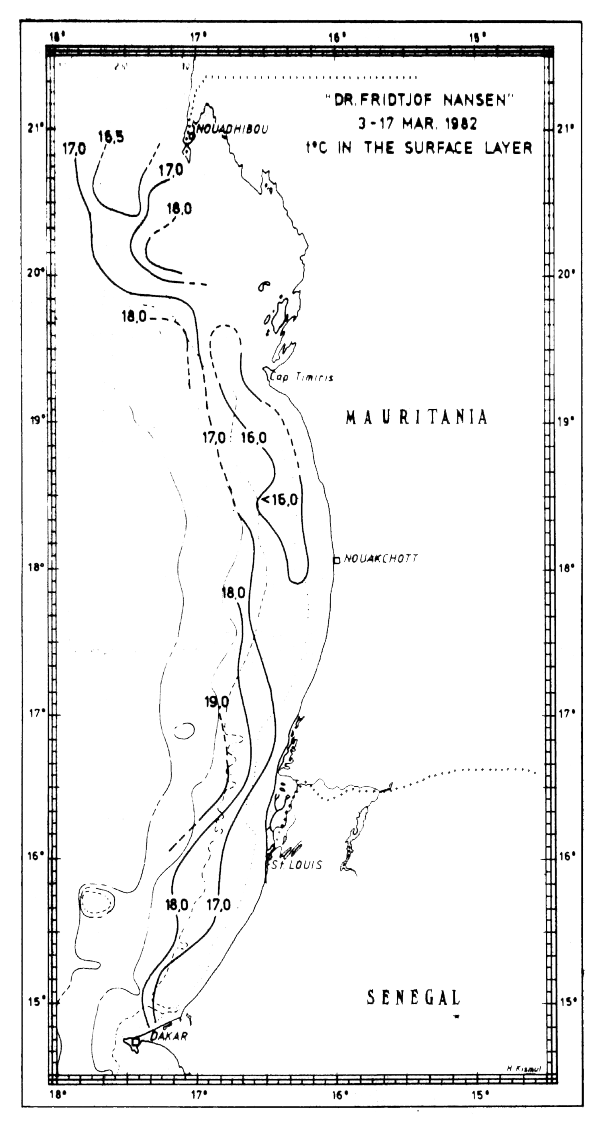
Fig. 15b Temperature in surface layer, Freetown to Cape Verde, Mar 3–17, 1982.
Sections VIII and IX, respectively to the north and south of Bissagos Islands, were worked in February 1982. They did not give any clear indications of upwelling. The density structure was suggestive of a southward flow near the edge and a northward flow of warmer, less saline waters of tropical origin over the shelf.
The surface layer in sections X and XI showed tropical characteristics with temperatures above 25°C and salinities below 35.5°/oo. There were no indications of upwelling.
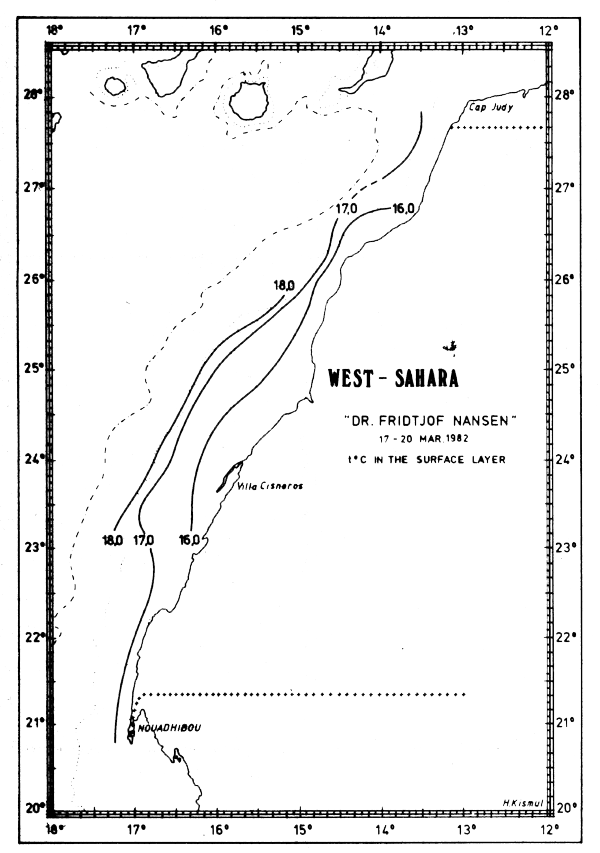
Fig. 15c Temperature in surface layer, Cape Blanc to Cape Juby, Mar 17–20, 1982.
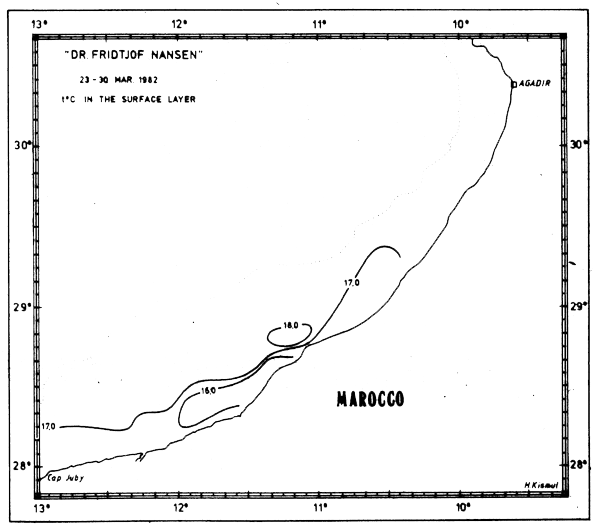
Fig. 15d Temperature in surface layer, Cape Juby to Agadir, Mar 23–30, 1982.
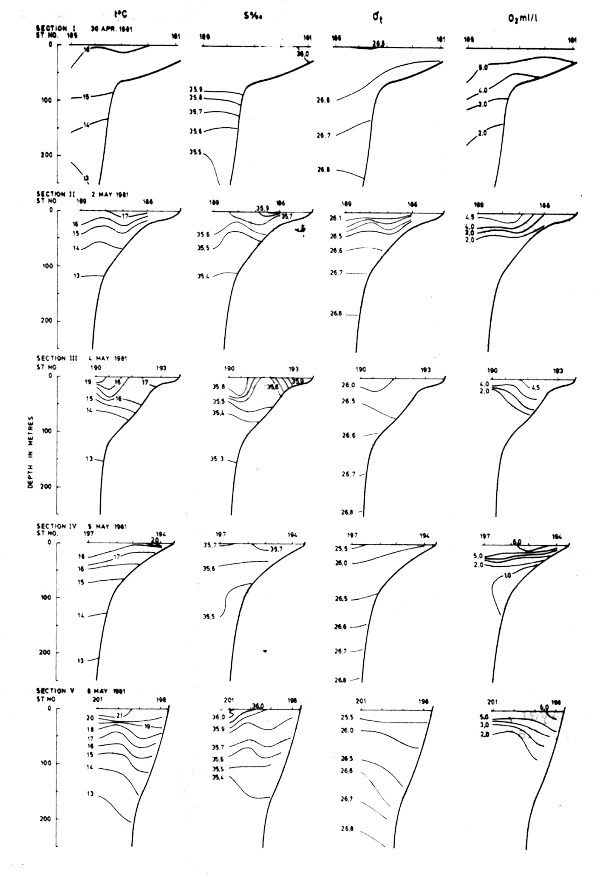
Fig. 16 Hydrographic sections in the area Cape Blanc to Cape Verde, Apr 30 - May 8, 1981
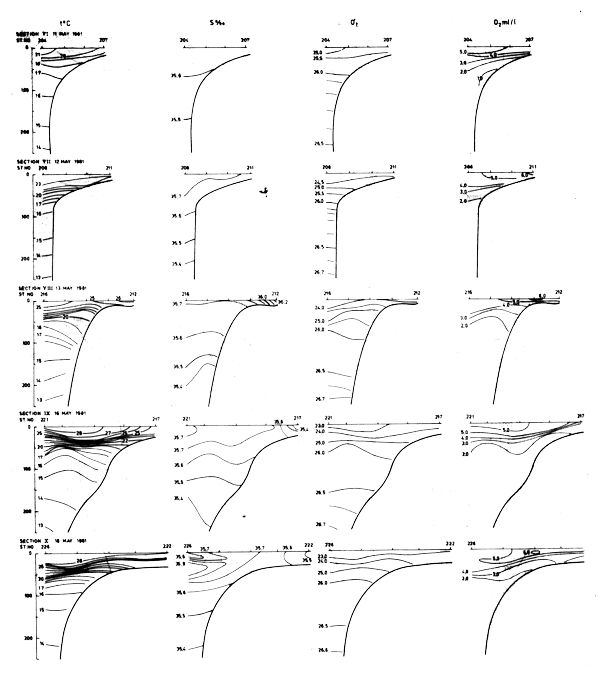
Fig. 17 Hydrographic sections in the area Cape Verde to Cape Verga, May 11–18, 1981
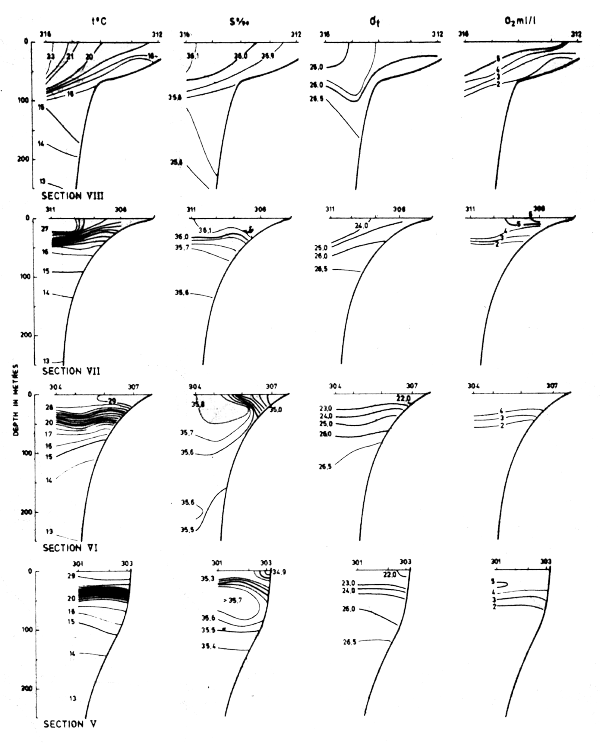
fig. 18 Hydrographic sections in the area Cape Blanc to Cape Verde, Sept 20–28, 1981.

Fig. 19 Hydrographic sections in the area Cape Verde to Cape Verga, Sept 8–20, 1981.
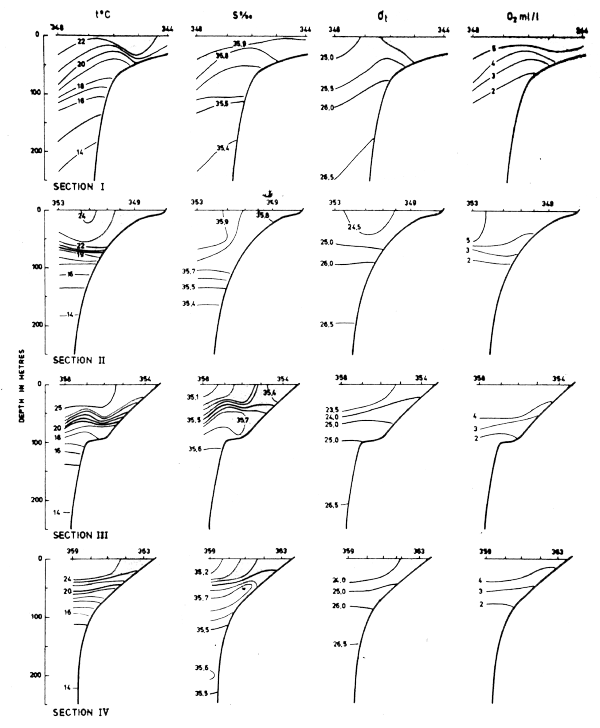
Fig. 20 Hydrographic sections in the area Cape Blanc to Cape Verde, Dec 5–14, 1981.
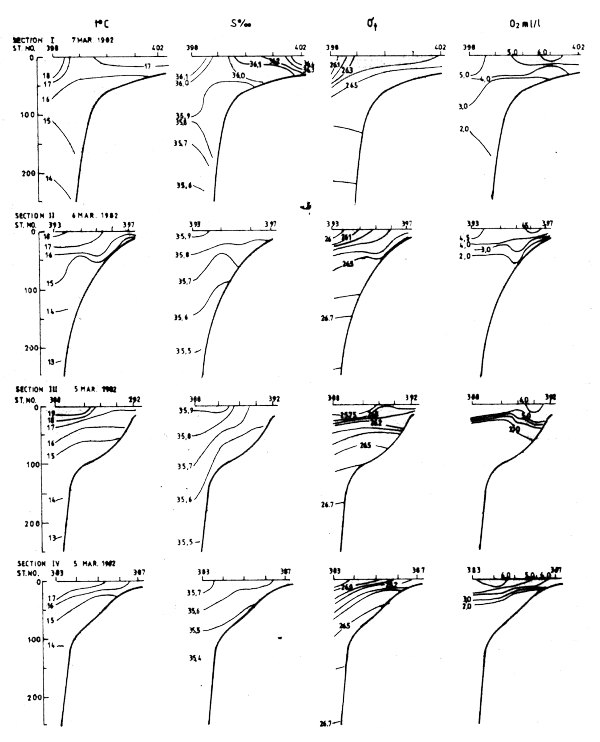
Fig. 21 Hydrographic sections in the area Cape Blanc to Cape Verde, Mar 3–17, 1982.
The density structure in the various sections and the temperature distribution at the surface are in general agreement with the model presented by Mittelstaedt, 1982. When upwelling occurred north of about 13°N during the northern winter there was a southward current near the coast. Further offshore, just past the shelf edge there were indications of a northward countercurrent. In Feb-82 such indications were observed south to the Guinean coast (Fig. 22). Such a structure also possibly occurred north of Cape Verde in May-81 (Fig. 16).
The observations between Cape Blanc and Cape Verde indicated that in May-81 upwelling associated with the past northern winter still occurred south of Cape Blanc. In September there was no upwelling and in December there were indications of moderate upwelling only close to Cape Blanc. In February 1982 the upwelling system was fully developed.
The distribution of dissolved oxygen indicates that an oxycline generally coincides with the thermocline. Here the oxygen content decreases with depth to less than 2 ml/l below the thermocline, i.e. at depths with temperatures below about 18°C. Below the mixed surface layer there is a well-defined interrelation between temperature and oxygen content.
In Jun-81 sea-surface temperatures were observed along the coast between Cape Verga and the Ghana-Togo border. The temperatures were generally between 27°C and 30°C, but close to the Ghana coast, temperatures below 26°C indicated that the local seasonal upwelling had just started (Fig. 23).
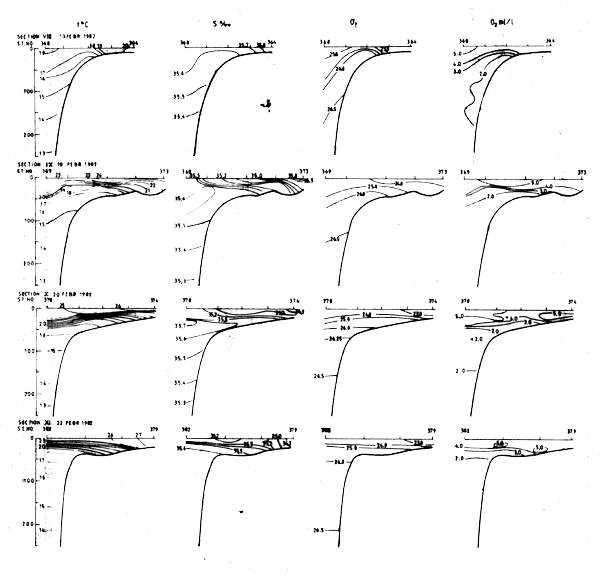
Fig. 22 Hydrographic section in the area Cape Verde to Freedown, Feb 8 - Mar 1, 1982.
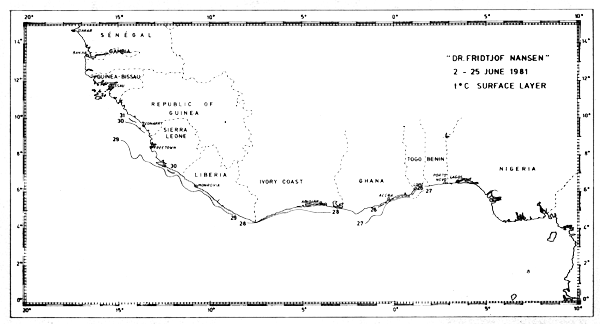
Fig. 23 Temperature in surface layer, Guinea to Ghana, June 2–25, 1981.
Figures 24–32 show the fish distributions from all surveys in three density levels: scattered, slightly gathered and dense. In mean absolute densities these levels correspond roughly to 14–140 tonnes/nm2, 140–280 tonnes/nm2 and greater than 280 tonnes/nm2.
The dominant species in the different locations are identified by trawling and marked off in the figures by a species code. The estimates of fish density and abundance for the various countries and surveys are given in Table 3, and illustrated in Fig. 33. The procedure for the estimations is given in Annex I. It will be seen from Table 3 that the grand totals include estimates for the shallow uninvestigated parts of the shelf. These estimates are based on the general assumption that the density per unit area in the shallows is equal to that of the shelf areas surveyed.
| Pelagic 1 Clupeids and anchov | Pelagic 2 Carangids | Balistes Trigger fish | Other fish | Total | Mean density in surveyed area tonnes/nm2 | Estimate for shallow unsurveyed area | Estimated Grand Total | |
|---|---|---|---|---|---|---|---|---|
| West Sahara, Dec-81 | x | x | x | 430 | 34 | 190 | 620 | |
| Mar-82 | x | x | x | 1360 | 115 | 730 | 2090 | |
| Mauritania, May-81 | 60 | 370 | - | 30 | 460 | 65 | 230 | 690 |
| Sep-81 | 100 | 190 | 2 | 70 | 360 | 52 | 200 | 560 |
| Dec-81 | 15 | 470 | 0 | 40 | 525 | 72 | 240 | 765 |
| Dec-81, replica survey | - | 370 | 0 | 50 | 420 | 58 | 200 | 620 |
| Mar-82 | 50 | 470 | 0 | 260 | 780 | 110 | 400 | 1180 |
| Mar-82, replica survey | 40 | 170 | 0 | 450 | 660 | 94 | 350 | 1010 |
| Senegambia, May-81 | 210 | 570 | 70 | 60 | 910 | 134 | 390 | 1300 |
| Sep-81 | 380 | 90 | 140 | 60 | 670 | 88 | 180 | 850 |
| Feb-82 | 40 | 90 | 40 | 430 | 600 | 94 | 310 | 910 |
| Guinea Bissau, May-81 | 90 | 40 | 380 | 140 | 650 | 107 | 620 | 1270 |
| Sep-81 | 20 | 20 | 350 | 80 | 470 | 73 | 400 | 870 |
| Feb-82 | 70 | 400 | 590 | 130 | 1190 | 165 | 780 | 1970 |
| Guinea, May-June-81 | 70 | 30 | 510 | 30 | 640 | 62 | 210 | 850 |
| Feb-82 | 30 | 10 | 720 | 10 | 770 | 90 | 470 | 1240 |
| Sierra Leone, June-81 | 80 | 120 | 80 | 20 | 300 | 52 | 120 | 420 |
| Feb-82 | 15 | 5 | 10 | 40 | 70 | 27 | (100) | 170 |
| Total Mauritania - Sierra Leone May-June-81 | 510 | 1130 | 1040 | 280 | 2960 | 82 | 1570 | 4530 |
| Feb-Mar-82 | 205 | 975 | 1360 | 870 | 3410 | 107 | 2060 | 5470 |
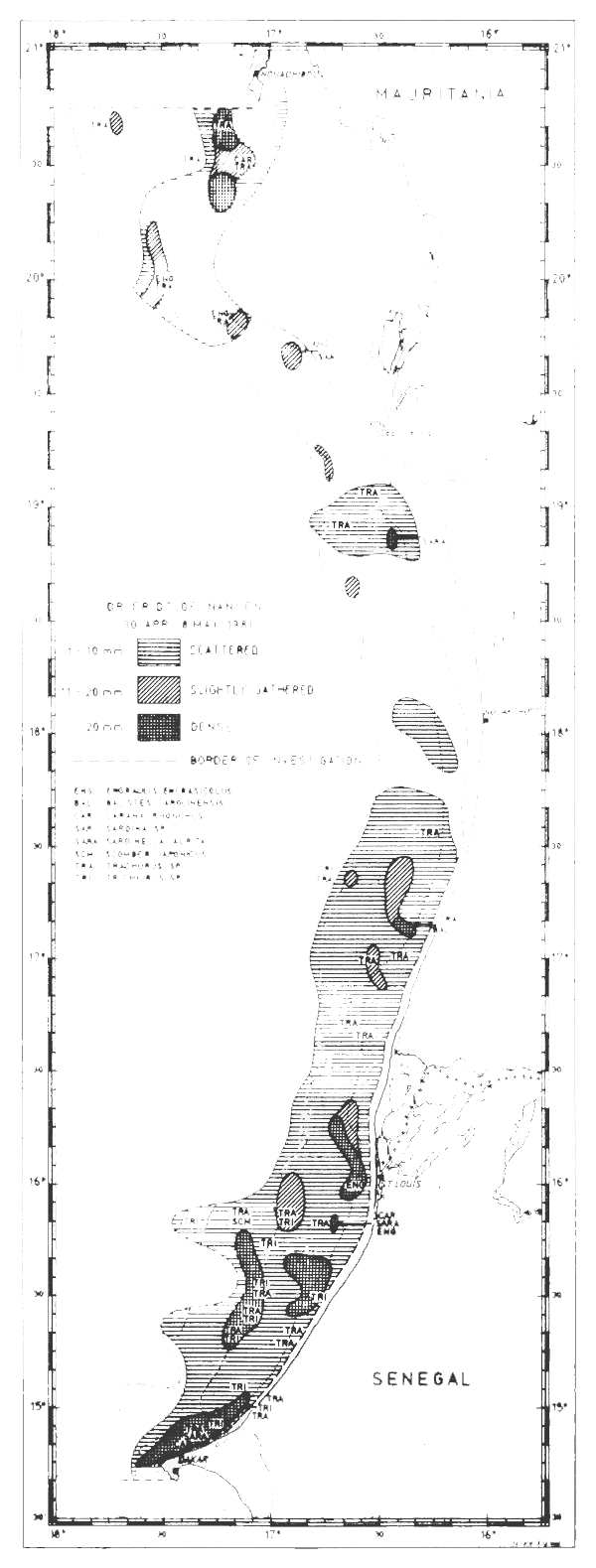
Fig. 24 Fish distributions, Cape Blanc to Cape Verde, Apr 30 May 8, 1981.
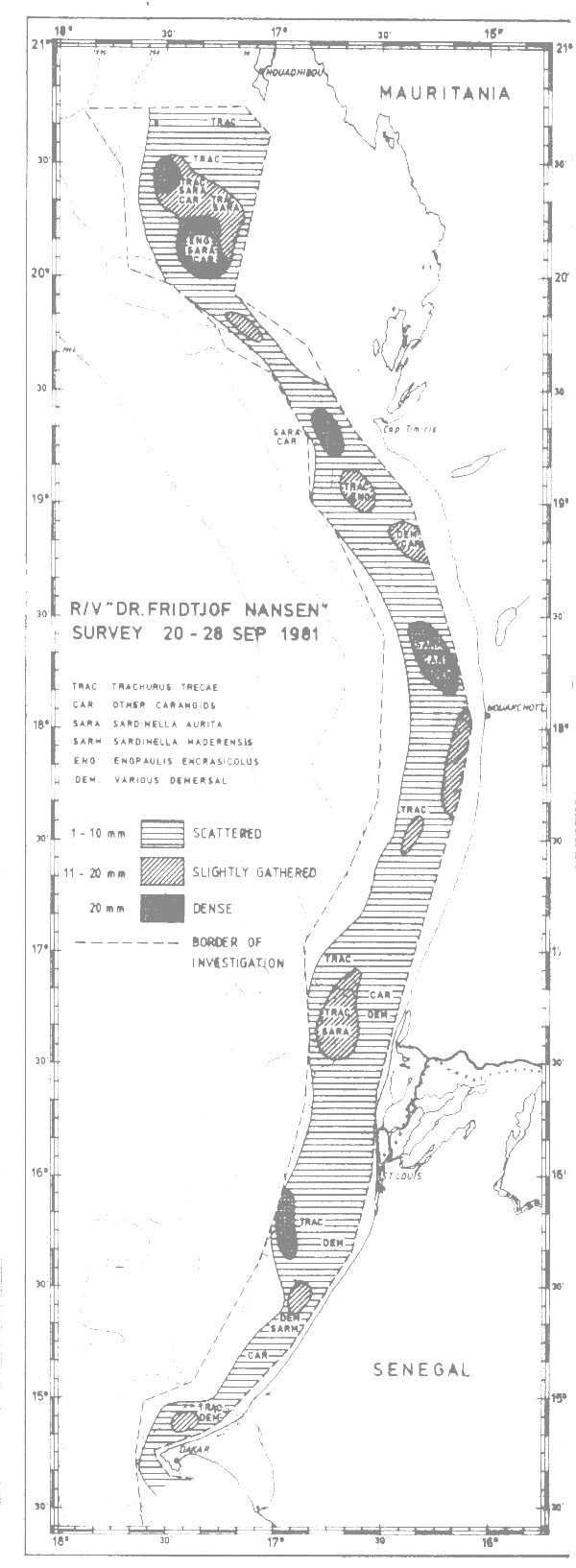
Fig. 25 Fish distributions, Cape Blanc to Cape Verde, Sept 20–28, 1981.
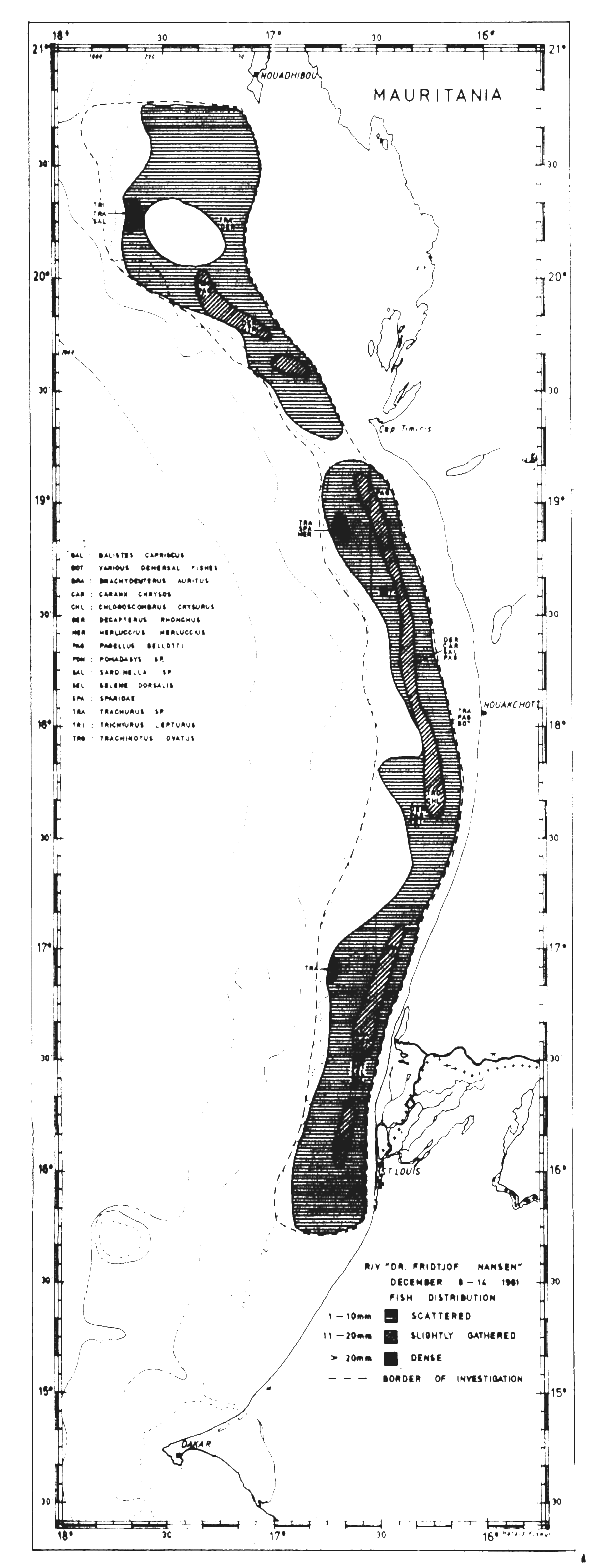
Fig. 26 Fish distributions, Cape Blanc to Cape Verde, Dec 8–14, 1981.
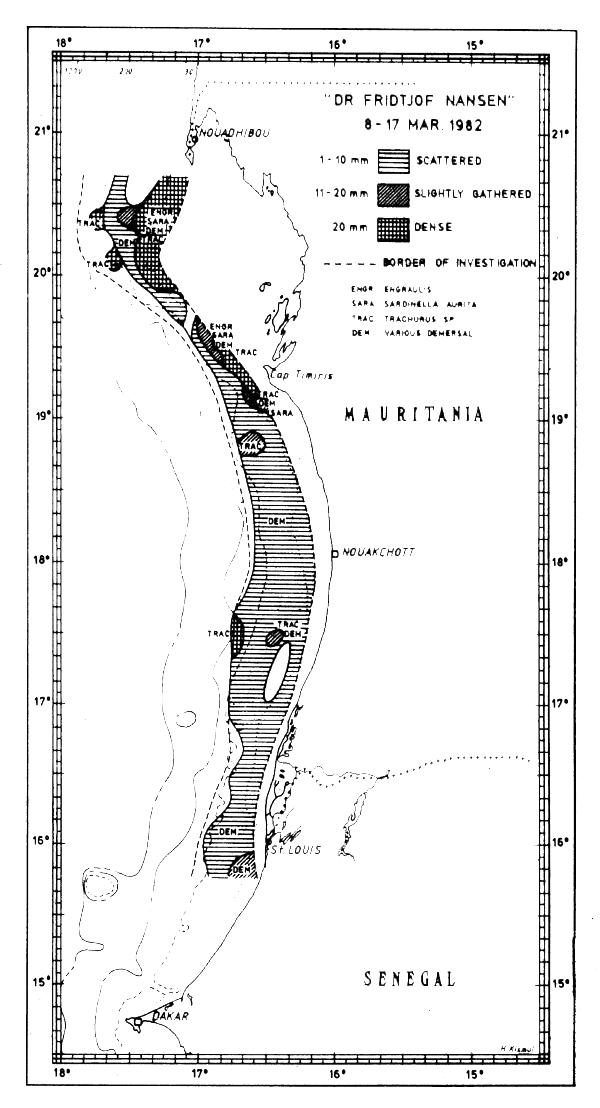
Fig. 27 Fish distributions, Mauritania, Mar 8–17, 1982.
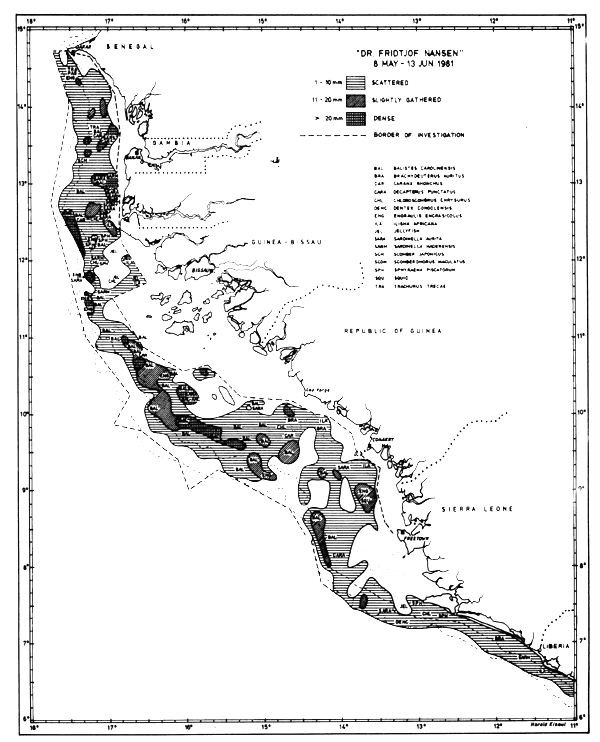
Fig. 28 Fish distributions, Cape Verde to Sierra Leone, May 8 - June 13, 1981.
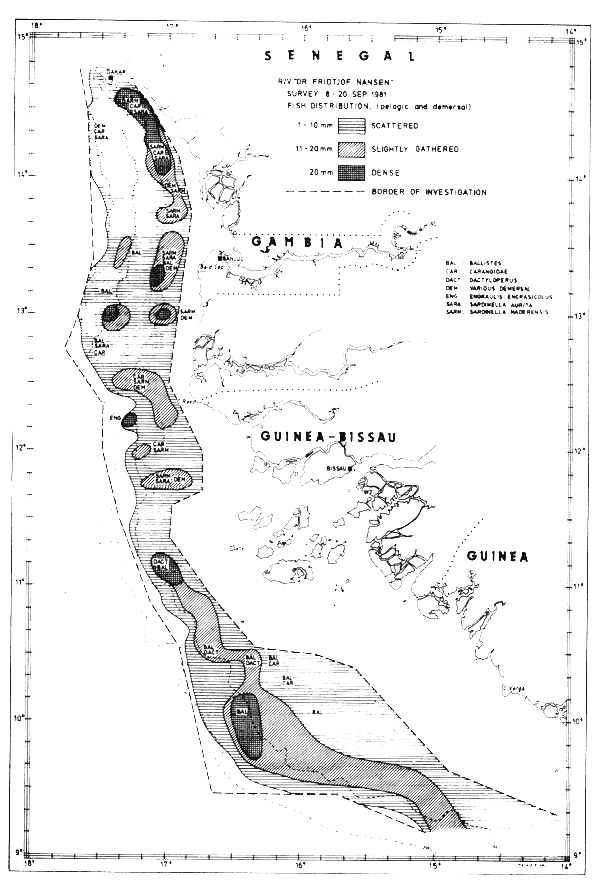
Fig. 29 Fish distributions, Cape Verde to Cape Verga, Sept 8–20, 1981.
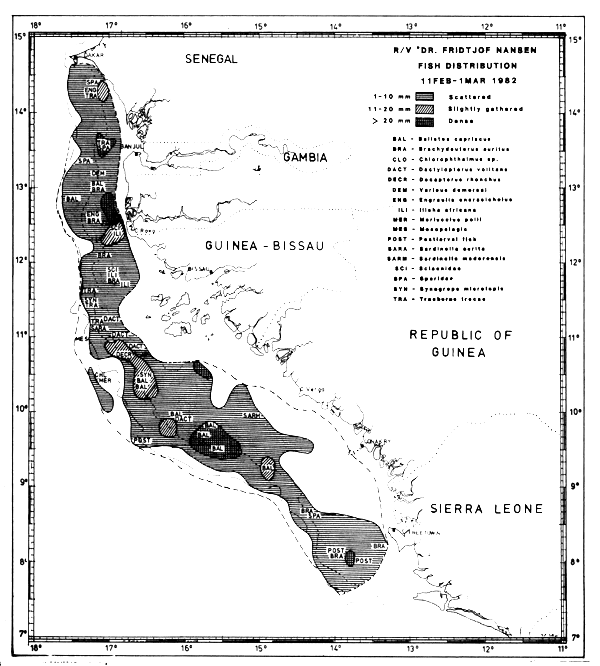
Fig. 30 Fish distributions, Cape Verde to Freedown, Feb 11 - Mar 1, 1982.
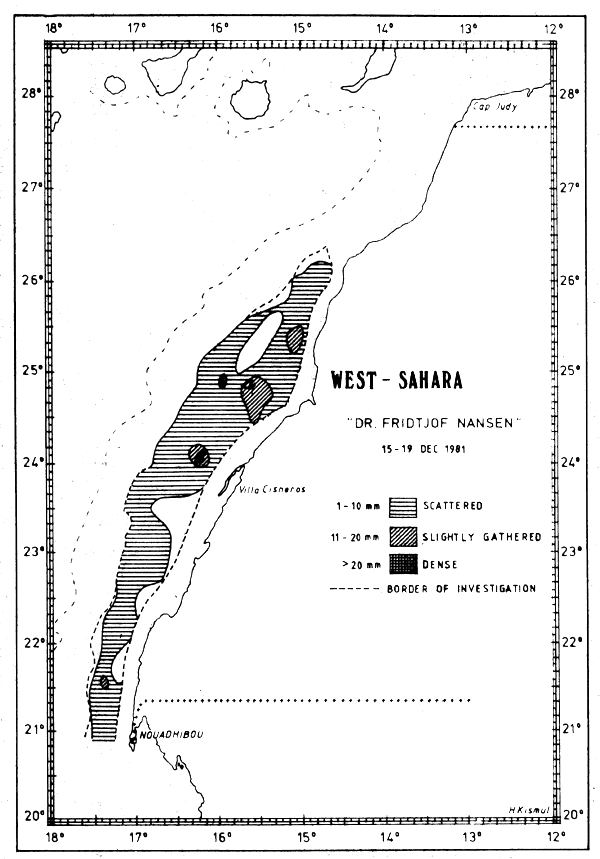
Fig. 31 Fish distributions, Cape Bojador to Cape Blanc, Dec 15–19, 1981.
The region Cape Blanc - Sherbro Island (Mauritania - Sierra Leone) has been fully surveyed twice during the programme, in May-Jun-81 and in Feb-Mar-1982. The total acoustic estimates are about 4.5 and 5.5 million tonnes respectively (Table 3). The distribution of fish groups was as follows (per cent):
| Clupeids and anchovies | Carangids | Trigger fish | Other fish | Total | |
|---|---|---|---|---|---|
| May-Jun-81 | 17 | 38 | 35 | 9 | 99 |
| Feb-Mar-82 | 6 | 29 | 40 | 25 | 100 |
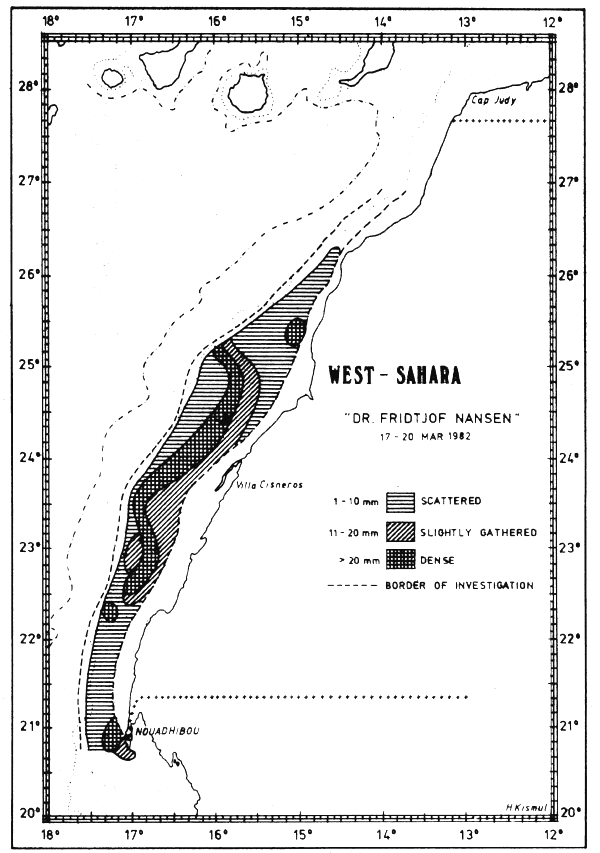
Fig. 32 Fish distributions, Cape Juby to Cape Blanc, Mar 17–20, 1982.
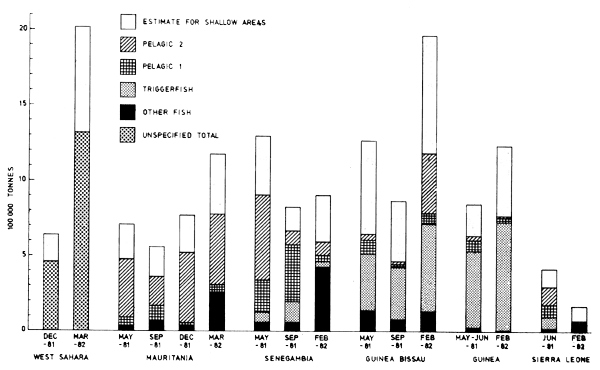
Fig. 33 Graphic illustration of the abundance estimates, in the regions from Western Sahara to Sierra Leone.
Only the triggerfish seems to be completely surveyed during both coverages as the area of investigation encompasses the distribution area of the species. Portions of the clupeid and carangid stocks are distributed north of the survey area as well, and large parts of the ‘other fish’ group seem to have had an offshore migration between the two coverages.
The two coverages off Western Sahara (Dec-81 and Mar-82) give total estimates of 0.6 and 2.1 million tonnes respectively. During the first survey the fish were located close to the shore, making only parts of it accessible to the vessel. During the second survey the center of distribution was more seaward and the fish community was thus more fully surveyed.
There does not seem to be any major ecological barriers in the whole shelf region from Sherbro Island to Cape Juby (at N 20°) except the seasonally fluctuating cold-water front, and in terms of fish communities the region can reasonably be treated as a single ecological unit.
This whole region was fully covered only once, in Feb-Mar-82. In this period many of the temporary shore-living species seem to have had a more seaward distribution than during the earlier surveys, and the total biomass was thus probably more fully covered by the vessel's sampling track. This therefore represents the most comprehensive survey. If one assumes the same density for the shallow areas as for the surveyed parts of the shelf, a total estimate for the region would be in the order of 7.5 million tonnes. Of this 1.4 million tonnes is allocated to the triggerfish. Separate estimates for the other fish groups are more uncertain as we do not know the species compositions off Western Sahara and in the shallow areas. From other investigations in the area we assume that the majority of the fish off Western Sahara belong to the horsemackerels (Trachurus trachurus, T. trecae and T. picturatus), the sardinellas (Sardinella aurita), the sardines (Sardina pilchardus) and mackerels (Scomber japonicus and S. scombrus). We therefore tentatively suggest that 75% of the fish off Western Sahara are small pelagics. The large shallow unsurveyed areas in the south, especially in Guinea and Guinea Bissau, probably hold considerable resources, and although the portion of pelagic fishes in this community is unknown, it is probably high. In order to obtain some rough assessments we assume the share of small pelagics as 50%, i.e. 1 million tonnes.
On the basis of the assumptions listed above and the biomass estimates in Table 3, we then arrive at a total estimate of small pelagic fish of 3.7 million tonnes for the region between Cape Juby and Sherbro Island. The resources of other fish are roughly set to 2.5 million tonnes. The regional estimates with their main subestimates and assumptions are summarized in Table 4.
| Abundance million tonnes rounded figures | Remarks | |
|---|---|---|
| Trigger fish | 1.4 | |
| Small pelagics: | ||
| Cape Blanc - Sierra Leone, surveyed area | 1.2 | |
| Cape Blanc - Sierra Leone, unsurveyed area* | 1.0 | Assuming (50%) small pelagic in the area |
| West Sahara, surveyed area | 1.0 | Assuming (75% )of total as small pelagics |
| West Sahara, unsurveyed area* | 0.5 | Assuming(75% )small pelagic in the area |
| Total small pelagics, whole region | 3.7 | |
| Other fish: | ||
| Cape Blanc - Sierra Leone, surveyed area | 0.9 | |
| Cape Blanc - Sierra Leone, unsurveyed area* | 1.0 | Assuming (50%) small pelagic in the area |
| West Sahara, surveyed area | 0.4 | Assuming (25%) of total as other fish |
| West Sahara, unsurveyed area* | 0.2 | Assuming (25%) other fish in the area |
| Total other fish, whole region | 2.5 | |
| Grand total estimate, whole region | 7.6 | |
* The unsurveyed areas are assumed to hold the same densities of fish as the surveyed areas.
One can reasonably question the high figures for the southern unsurveyed areas, and until further data are obtained from the area, more conservative assessments could be preferable. By halving the assumed densities in the area, the pelagic and other fish estimates for the whole region would be lowered to 3.2 and 2 million tonnes respectively.
The “Other Fish” group consists of dominating elements of seabreams (Sparidae), grunts (Brachydeuterus auritus, Pomadasys sp.), croakers (Sciaenidae), groupers (Serranidae), hairtails (Trichiurus lepturus), lizardfishes (Synodontidae) and barracudas (Sphyraenidae).
Tables 5 to 7 present catch distributions by family and bottom depth strata. The tables include all stations in the region from Cape Blanc to Sierra Leone, and the three coverages are treated separately. Further tables with the data sorted into subregions are given in Annex VI. The family codes used in the tables are explained in the beginning of the annex.
| % incidence in tot. no. of hauls | Mean c. of c. >1kg | % of total catch | MEAN CATCH OF ALL HAULS REFERRED TO BOTTOM DEPTH STRATA | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GEAR: BOTTOM TRAWL | |||||||||||||
| FAMILY | CATCH GROUPS (kg/h) | ||||||||||||
| 1–9 | 10–49 | 50–199 | 200–499 | >500 | <20m | 20–50m | 50–100m | >100m | >200m | ||||
| BAL | 1 | 0 | 3 | 5 | 14 | 48 | 1338.2 | 46 | 23.6 | 1254.9 | 67.2 | .0 | .0 |
| POD | 8 | 7 | 6 | 6 | 3 | 50 | 533.7 | 19 | 348.5 | 323.1 | 3.9 | .0 | .0 |
| CAP | 9 | 10 | 7 | 10 | 3 | 65 | 191.0 | 9 | 76.2 | 109.9 | 214.8 | 420.0 | .0 |
| JEL | 1 | 3 | 7 | 6 | 2 | 32 | 395.9 | 9 | 314.5 | 57.0 | 13.7 | .0 | .0 |
| SPA | 4 | 9 | 13 | 4 | 2 | 53 | 117.0 | 4 | 49.3 | 49.0 | 118.7 | 79.8 | .0 |
| DAC | 4 | 6 | 2 | 2 | 1 | 25 | 109.2 | 2 | 2.3 | 53.0 | .6 | .0 | .0 |
| SCI | 4 | 5 | 4 | 2 | 1 | 27 | 122.7 | 2 | 97.7 | 5.9 | 2.0 | 5.6 | .0 |
| ARD | 3 | 5 | 2 | 0 | 0 | 17 | 39.1 | 1 | 20.8 | .6 | .0 | .0 | .0 |
| CLU | 8 | 9 | 6 | 0 | 0 | 38 | 40.2 | 1 | 46.0 | 1.6 | 4.3 | .0 | .0 |
| ENG | 2 | 1 | 0 | 0 | 1 | 7 | 159.3 | 1 | 34.2 | .0 | 1.8 | .0 | .0 |
| PLN | 7 | 6 | 3 | 1 | 0 | 28 | 58.1 | 1 | 48.5 | 3.8 | .0 | .0 | .0 |
| SER | 3 | 6 | 1 | 1 | 0 | 18 | 41.3 | 1 | .2 | 5.4 | 26.1 | .0 | .0 |
| SPH | 4 | 5 | 2 | 1 | 0 | 20 | 45.2 | 1 | 17.8 | 7.2 | .4 | .0 | .0 |
| SQU | 8 | 9 | 4 | 1 | 0 | 37 | 38.2 | 1 | 4.7 | 18.0 | 19.6 | .0 | .0 |
| TRI | 2 | 8 | 0 | 1 | 0 | 18 | 52.5 | 1 | 29.2 | 1.7 | .0 | .0 | .0 |
| Other fish | 35.7 | 2 | |||||||||||
| MEAN CATCH OF ALL HAULS | 1410.5 | 1154.8 | 1918.7 | 499.0 | 792.0 | .0 | |||||||
| TOTAL NUMBER OF HAULS | 60 | 18 | 30 | 11 | 1 | 0 | |||||||
| % incidence in tot. no. of hauls | Mean c. of c. >1kg | %of total catch | MEAN CATCH OF ALL HAULS REFERRED TO BOTTOM DEPTH STRATA | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GEAR:PELAGIC TRAWL | |||||||||||||
| FAMILY | CATCH GROUPS (kg/h) | ||||||||||||
| 1–9 | 10–49 | 50–199 | 200–499 | >500 | <20m | 20–50m | 50–100m | >100m | >200m | ||||
| BAL | 7 | 6 | 3 | 1 | 4 | 24 | 1181.1 | 31 | 3.2 | 706.8 | 1.9 | .1 | .0 |
| ENG | 1 | 3 | 4 | 1 | 6 | 17 | 996.7 | 18 | 8.7 | 171.5 | 295.3 | 72.7 | 129.6 |
| CAR | 19 | 12 | 8 | 8 | 2 | 55 | 198.6 | 12 | 6.2 | 191.4 | 74.3 | 48.8 | 30.0 |
| JEL | 5 | 4 | 4 | 1 | 3 | 19 | 529.0 | 11 | .6 | 126.8 | 165.2 | 23.4 | 25.1 |
| CAP | 0 | 0 | 0 | 0 | 1 | 1 | 7520.0 | 9 | .0 | .0 | .0 | 300.8 | .0 |
| CLU | 8 | 12 | 8 | 2 | 1 | 35 | 110.4 | 4 | 311.2 | 42.9 | 15.1 | .1 | .0 |
| CRU | 3 | 3 | 4 | 0 | 2 | 14 | 171.6 | 3 | .0 | .0 | 33.3 | 50.4 | 12.6 |
| POD | 6 | 3 | 3 | 3 | 1 | 18 | 159.1 | 3 | 23.8 | 68.7 | .9 | .0 | .0 |
| SPA | 2 | 0 | 1 | 0 | 1 | 5 | 657.0 | 3 | .3 | 74.8 | .0 | .3 | .0 |
| MYC | 3 | 6 | 3 | 1 | 0 | 15 | 51.3 | 1 | .0 | .0 | .0 | 26.7 | 43.3 |
| SCM | 12 | 6 | 2 | 0 | 0 | 23 | 18.2 | 1 | 5.2 | 2.8 | 3.9 | 5.9 | 9.8 |
| SPH | 8 | 6 | 5 | 0 | 0 | 21 | 44.1 | 1 | 4.9 | 18.3 | 7.2 | .1 | .0 |
| TRI | 10 | 8 | 4 | 1 | 0 | 26 | 37.2 | 1 | .2 | 1.7 | 3.4 | 28.5 | 17.6 |
| Other fish | 21.9 | 2 | |||||||||||
| MEAN CATCH OF ALL HAULS | 913.8 | 388.8 | 1423.5 | 619.7 | 587.5 | 302.2 | |||||||
| TOTAL NUMBER OF HAULS | 89 | 5 | 35 | 24 | 25 | 14 | |||||||
Of the total catch in the pelagic trawl the families Balistilae (triggerfish), Carangidae (horsemackerel, scad), Clupeidae (sardinella) and Engraulidae (anchovy) constituted jointly 60%, 90% and 81% in the three surveys (Tables 4–6). Of the bottom trawl catch the families Carangidae, Balistidae, Pomadasyidae (grunts) and Sparidae (seabreams) comprised 78%, 69% and 59% in the three coverages. The family Dactylopteridae (flying gurnards)
made notable contributions to the total catches in September, but occurrences were restricted to a few stations with high catches (Table 6). Apogonidae (cardinal fishes) and Caproidae (boarfishes) made similar contributions, appearing in high numbers at single stations. It should be borne in mind that the tables also include the amount of jellyfish in the catches, and the above percentages would be increased if these were excluded.
| % incidence in tot. no. of hauls | Mean c. of c. >1kg | % of total catch | MEAN CATCH OF ALL HAULS REFERRED TO BOTTOM DEPTH STRATA | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GEAR:BOTTOM TRAWL | ||||||||||||
| FAMILY | CATCH GROUPS (kg/h) | |||||||||||
| 1–9 | 10–49 | 50–199 | 200–499 | >500 | <20m | 20–50m | 50–100m | >100m | ||||
| CAR | 5 | 5 | 7 | 5 | 6 | 76 | 631.7 | 29 | 281.7 | 166.2 | 2099.2 | 221.9 |
| SPA | 4 | 3 | 3 | 6 | 6 | 60 | 638.0 | 23 | 527.5 | 97.3 | 734.2 | 496.8 |
| DAC | 0 | 0 | 0 | 0 | 2 | 5 | 3104.4 | 10 | .0 | 355.6 | 175.0 | .0 |
| POD | 1 | 4 | 6 | 4 | 2 | 46 | 321.1 | 9 | 173.9 | 189.9 | .0 | .0 |
| PAL | 0 | 4 | 5 | 4 | 4 | 46 | 278.5 | 8 | 67.3 | 226.6 | 65.0 | .0 |
| CLU | 1 | 10 | 4 | 1 | 2 | 49 | 193.4 | 6 | 46.4 | 183.5 | 6.7 | .0 |
| ENC | 0 | 0 | 0 | 0 | 1 | 3 | 2000.0 | 3 | .0 | 133.3 | .0 | .0 |
| SCI | 2 | 1 | 0 | 0 | 2 | 14 | 401.1 | 3 | 133.5 | .2 | .0 | .0 |
| PLN | 2 | 2 | 2 | 0 | 1 | 19 | 130.9 | 2 | 58.0 | 3.1 | .0 | .0 |
| ARD | 3 | 0 | 2 | 1 | 0 | 16 | 105.1 | 1 | 42.0 | .0 | .0 | .0 |
| MUL | 1 | 3 | 3 | 1 | 0 | 22 | 92.6 | 1 | 35.0 | 14.3 | .0 | .0 |
| SHA | 2 | 2 | 1 | 1 | 0 | 16 | 67.4 | 1 | 3.3 | 23.6 | .0 | .0 |
| SHR | 2 | 0 | 0 | 0 | 1 | 8 | 248.0 | 1 | 49.6 | .0 | .0 | .0 |
| SPH | 1 | 3 | 3 | 0 | 0 | 19 | 46.3 | 1 | 10.9 | 10.7 | .0 | .0 |
| SQU | 4 | 6 | 3 | 0 | 0 | 35 | 36.4 | 1 | 7.1 | 17.6 | 18.2 | 5.7 |
| TRI | 0 | 2 | 4 | 0 | 0 | 16 | 69.9 | 1 | 17.2 | 10.8 | .0 | .0 |
| Other fish | 36.0 | 2 | ||||||||||
| MEAN OF TOTAL CATCHES | 1664.8 | 1493.9 | 1469.0 | 3123.0 | 769.5 | |||||||
| NUMBER OF HAULS | 37 TOTAL | 15 | 15 | 5 | 2 | |||||||
| % incidence in tot. no. of hauls | Mean c. of c. >1kg | % of total catch | MEAN CATCH OF ALL HAULS REFERRED TO BOTTOM DEPTH STRATA | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GEAR:PELAGIC TRAWL | |||||||||||||
| FAMILY | CATCH GROUPS (kg/h) | ||||||||||||
| 1–9 | 10–49 | 50–199 | 200–499 | >500 | <20m | 20–50m | 50–100m | >100m | |||||
| BAL | 2 | 1 | 2 | 1 | 3 | 23 | 1956.2 | 42 | .3 | 58.6 | 666.5 | .0 | |
| ENG | 0 | 0 | 1 | 1 | 1 | 8 | 4200.3 | 30 | .0 | 28.5 | 14.5 | 4000.0 | |
| CAR | 8 | 6 | 3 | 5 | 1 | 59 | 267.1 | 15 | 20.3 | 85.1 | 458.0 | .0 | |
| DAC | 2 | 0 | 0 | 1 | 2 | 13 | 476.6 | 6 | .0 | .2 | 238.0 | .0 | |
| CLU | 5 | 7 | 6 | 1 | 0 | 49 | 59.7 | 3 | 60.7 | 30.4 | 4.0 | .0 | |
| ARD | 2 | 0 | 0 | 1 | 0 | 8 | 157.9 | 1 | .2 | .1 | 47.0 | .0 | |
| POD | 1 | 0 | 1 | 1 | 0 | 8 | 123.2 | 1 | 37.0 | .0 | .0 | .0 | |
| SCM | 3 | 6 | 0 | 1 | 0 | 26 | 39.3 | 1 | 3.4 | 6.9 | 24.9 | .0 | |
| SHA | 0 | 1 | 2 | 1 | 0 | 10 | 111.3 | 1 | 36.2 | 1.5 | 6.0 | .0 | |
| TRI | 2 | 4 | 1 | 0 | 0 | 18 | 38.7 | 1 | .0 | 15.8 | 1.8 | .0 | |
| Other fish | 4.1 | 0 | |||||||||||
| MEAN OF TOTAL CATCHES | 1076.5 | 162.2 | 231.3 | 2463.2 | 4009.3 | ||||||||
| NUMBER OF HAULS | 39 TOTAL | 10 | 16 | 10 | 3 | ||||||||
To conclude, Tables 5–7 show with the cautions mentioned, that the biomass on the shelf from Mauritania to Sierra Leone is made up of some few families. Further analysis at the species level should reveal that these families are made up of some few species. These species will be dealt with below (see also Annex V).
| % incidence in tot. no. of hauls | Mean c. of c. >1kg | % of total catch | MEAN CATCH OF ALL HAULS REFERRED TO BOTTOM DEPTH STRATA | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GEAR: BOTTOM TRAWL | ||||||||||||
| FAMILY | CATCH GROUPS (kg/h) | |||||||||||
| 1–9 | 10–49 | 50–199 | 200–499 | >500 | <20m | 20–50m | 50–100m | >100m | ||||
| CAR | 10 | 4 | 4 | 2 | 4 | 71 | 171.2 | 18 | 2.5 | 88.6 | 199.7 | 382.0 |
| PAL | 3 | 1 | 0 | 0 | 1 | 15 | 725.1 | 16 | .0 | 241.7 | .0 | .0 |
| SPA | 3 | 8 | 5 | 0 | 3 | 56 | 164.2 | 14 | .0 | 103.9 | 138.3 | 88.3 |
| POD | 4 | 5 | 3 | 4 | 1 | 50 | 147.7 | 11 | 48.0 | 128.1 | 25.5 | .0 |
| APC | 0 | 0 | 0 | 0 | 1 | 3 | 2040.0 | 9 | .0 | .0 | 204.0 | .0 |
| SCI | 0 | 4 | 5 | 1 | 1 | 32 | 173.2 | 8 | 165.1 | 20.9 | 43.6 | .0 |
| JEL | 2 | 1 | 5 | 1 | 1 | 29 | 170.0 | 7 | 155.8 | 40.5 | .2 | .0 |
| TRI | 3 | 5 | 3 | 1 | 0 | 35 | 53.7 | 3 | 9.1 | 32.0 | 10.1 | .0 |
| ENG | 2 | 1 | 0 | 1 | 0 | 12 | 100.4 | 2 | .5 | 26.6 | .0 | .0 |
| SQU | 10 | 6 | 3 | 0 | 0 | 56 | 19.6 | 2 | 6.0 | 17.7 | 5.2 | 6.3 |
| ARD | 1 | 3 | 2 | 0 | 0 | 18 | 37.9 | 1 | 23.3 | 3.0 | 2.0 | .0 |
| CEN | 1 | 0 | 1 | 0 | 0 | 6 | 62.2 | 1 | .0 | .0 | .4 | 60.4 |
| CLU | 2 | 2 | 2 | 0 | 0 | 18 | 24.6 | 1 | 18.7 | 1.0 | .2 | .0 |
| KER | 2 | 0 | 1 | 0 | 0 | 9 | 35.9 | 1 | .0 | 6.8 | .0 | 2.9 |
| PLN | 2 | 1 | 0 | 1 | 0 | 12 | 73.5 | 1 | 42.0 | .0 | .0 | .0 |
| RAY | 4 | 2 | 1 | 0 | 0 | 21 | 16.9 | 1 | 1.7 | .3 | 10.2 | .0 |
| SER | 1 | 4 | 1 | 0 | 0 | 18 | 33.1 | 1 | .0 | 13.1 | .3 | .0 |
| SHR | 6 | 1 | 2 | 0 | 0 | 27 | 19.5 | 1 | 11.1 | .6 | 8.7 | .7 |
| TRG | 0 | 1 | 3 | 0 | 0 | 12 | 68.3 | 1 | .0 | 5.4 | 19.2 | .0 |
| ZEI | 4 | 4 | 0 | 0 | 0 | 24 | 19.2 | 1 | .0 | 1.3 | 9.3 | 20.2 |
| Other fish | 25.0 | 3 | ||||||||||
| MEAN CATCH OF ALL HAULS | 679.4 | 497.0 | 749.8 | 716.3 | 605.7 | |||||||
| TOTAL NUMBER OF HAULS | 34 | 7 | 15 | 10 | 2 | |||||||
| % incidence in tot. no. of hauls | Mean c. of c. >1kg | % of total catch | MEAN CATCH OF ALL HAULS REFERRED TO BOTTOM DEPTH STRATA | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GEAR: PELAGIC TRAWL | ||||||||||||
| FAMILY | CATCH GROUPS (kg/h) | |||||||||||
| 1–9 | 10–49 | 50–199 | 200–499 | >500 | <20m | 20–50m | 50–100m | >100m | ||||
| BAL | 0 | 3 | 2 | 1 | 4 | 23 | 1904.8 | 70 | .O | 389.1 | 1040.0 | 86.6 |
| POD | 6 | 1 | 1 | 1 | 1 | 23 | 318.4 | 12 | 4.8 | 21.8 | 273.0 | .0 |
| CLU | 2 | 2 | 0 | 4 | 0 | 18 | 158.6 | 5 | 91.9 | 24.8 | 40.6 | .0 |
| CAR | 7 | 5 | 0 | 0 | 1 | 30 | 62.6 | 3 | 4.6 | 1.3 | 75.8 | 1.0 |
| DAC | 1 | 1 | 1 | 1 | 0 | 9 | 175.4 | 3 | .4 | 35.0 | .0 | .0 |
| ENG | 0 | 2 | 3 | 1 | 0 | 14 | 120.0 | 3 | 88.3 | 18.3 | .0 | .0 |
| JEL | 1 | 4 | 3 | 0 | 0 | 18 | 43.4 | 1 | .0 | 9.0 | .3 | 16.5 |
| KYC | 0 | 0 | 1 | 0 | 0 | 2 | 156.8 | 1 | .0 | .0 | .0 | 15.7 |
| SQU | 6 | 1 | 2 | 0 | 0 | 21 | 22.8 | 1 | 2.1 | 8.1 | .6 | 2.8 |
| TRI | 3 | 3 | 1 | 0 | 0 | 16 | 39.1 | 1 | .0 | 9.5 | 5.8 | 2.6 |
| Other fish | 9.6 | 1 | ||||||||||
| MEAN CATCH OF ALL HAULS | 616.9 | 238.1 | 524.1 | 1438.4 | 132.4 | |||||||
| TOTAL NUMBER OF HAULS | 44 | 4 | 20 | 10 | 10 | |||||||
As already mentioned Table 3 includes biomass estimates for the various countries in the region. These estimates have obvious limitations as most of the dominant pelagic species migrate seasonally along the coast and form stocks shared between countries.
An exception is the triggerfish (Balistes capriscus) with its predominance in Guinea Bissau and Guinea, and only fringes of the distribution reaching into Senegambian and Sierra Leone waters. The estimates are from 350 to 590 thousand tonnes for Guinea Bissau and 510–720 thousand tonnes for Guinea. From two surveys it seems as if the abundance of triggerfish is somewhat higher in Guinea compared with Guinea Bissau, with respective means of 615 and 485 thousand tonnes.
The estimated fish densities (Table 3) can cautiously be taken as rough indications of the productivity in the area when the migration pattern and fishing mortality are taken into account. The estimated densities expressed as the means from all surveys are as follows:
| West Sahara | (2 coverages) | 75 tonnes/nm2 |
| Mauritania | (6 coverages) | 75 tonnes/nm2 |
| Senegambia | (3 coverages) | 105 tonnes/nm2 |
| Guinea Bissau | (3 coverages) | 115 tonnes/nm2 |
| Guinea | (2 coverages) | 75 tonnes/nm2 |
| Sierra Leone | (2 coverages) | 40 tonnes/nm2 |
It is reasonable to increase the first estimate for Western Sahara somewhat as the fish were then located close to the shore and are hence probably underestimated. A mean density in the order of 100 tonnes/nm2 is more reasonable, and in accordance with the second survey in the area.
Mauritania seems to be the most exploited area and the unfished density is probably considerably higher, perhaps well above 100 tonnes/nm2. From this we may assume that the fish production along the shelf from Western Sahara to Guinea Bissau is fairly uniform and that density values in the order of 100–150 tonnes are representative for relatively unexploited areas.
Guinea seems to have a slightly lower density level with around 75 tonnes/nm2, and Sierra Leone, receiving insignificant benefits from the upwelling system, has only 40 tonnes/nm2 mean density. This is a level usually more representative of steady state tropical communities, as for instance large parts of the shelf in Gulf of Guinea (see also Part 4 and StrÆmme et al. 1983).
Marchal and Boely (1977) assessed the fish biomass between Cape Verga and Cape Bojador in 1974 at roughly 11.2 million tonnes. As mentioned earlier we have assessed the biomass in the same region to be in the order of 7.5 million tonnes for early 1982.
Although this could indicate that the total biomass has been reduced to 2/3 of its 1974 level, small differences in the methods of calibration for the acoustic equipment and in the conversion factors used may easily have affected the estimates to the same degree. Unfortunately the calibration methods for the electronic equipment and for target strength measurements on fish have hitherto not been internationally standarized. This seriously hampers the comparability of the acoustic estimates.
The area between Cape Blanc and Cape Roxo is the most intensively covered by acoustic surveys conducted mainly by the R/V Capricorne. Table 8 gives the estimates from the 1974 survey and from the three most recent surveys from which reports are available. For comparision the estimates from the R/V Dr. Fridtjof Nansen surveys are transformed to cover the same area and are included in the table.
| Project | Year | Season | Estimated abundance (1000 tonnes) | Reference | ||||
|---|---|---|---|---|---|---|---|---|
| Total | Clupeids and anchovies | Carangids | Balistes | Other | ||||
| CAP7408 | 1974 | Oct-Nov | 2380 | 683 | 1013 | - | 684 | Marchal and Boely 1977 |
| ECHOSAR 1 | 1980 | Feb | 1582 | n.s. | n.s. | n.s. | n.s. | Marchal, Gerlotto and Josse 1981 |
| ECHOSAR 2 | 1980 | Sep | 1005 | n.s. | n.s. | n.s. | n.s. | Marchal and Josse 1981 |
| ECHOSAR 3 | 1981 | May | 2420 | 625 | 658 | 103 | 1034 | Marchal and Josse 1982 |
| NANSEN 1 | 1981 | May | 1370 | 270 | 940 | 70 | 90 | Table 3 |
| NANSEN 2 | 1981 | Sep | 1030 | 480 | 280 | 140 | 130 | Table 3 |
| NANSEN 3 | 1982 | Feb-Mar | 1380 | 90 | 560 | 40 | 690 | Table 3 |
The most striking feature in the table is the relatively high total estimates from the ECHOSAR 3 survey. The results from the Fridtjof Nansen surveys seem to agree reasonably well with the total estimates from the ECHOSAR 1 and 2 surveys, especially if one allows for some variation due to seasonal fluctuations. The first Nansen survey was carried out almost synchronously with ECHOSAR 3, and unfortunately the results from the two surveys do not correspond, neither in total estimates nor in allocation to species groups. The discrepancies between the estimates on total fish abundance could be due to different target strength reference values, while the differences between the estimated species composition seem to point to methodological differences in the design and execution of the survey particularly with respect to the sampling systems used.
ECHOSAR 3 uses - 33.4 dB/kg as target strength for a 17 cm fish on the 120 kHz system, whereas during the Fridtjof Nansen surveys TS = -34.3 dB/kg has been applied on the 38 kHz system. Saetersdal et al. (1983) has pointed to a systematic difference in recorded target strengths when the 120 and 38 kHz systems are run simultaneously. A mean target strength value 3–4 dB lower should be expected on the 38 kHz system. To make the two vessels' TS values comparable we transform the Nansen TS-value from - 34.3 dB (ref. 38 kHz) to 31.3 – 30.3 dB (ref. 120 kHz). This systematic difference of 2–3 dB between the two surveys could account for 60–100% higher estimates from the system onboard Capricorne.
The ECHOSAR 2 survey of September 1980 formed part of a joint survey with R/V Cornide de Saavedra covering the shelf from Sierra Leone to Guinea Bissau and R/V Ibn Sina off Western Sahara heading north to Gibraltar. The shelf from Cape Blanc south to, and including, Sierra Leone was estimated to hold roughly 2 million tonnes of fish (FAO/CECAF 1981a). The corresponding figures from the two R/V Dr. Fridtjof Nansen coverages are roughly 3 and 3.4 million tonnes. Some of the difference could be due to seasonal migration of the fish stocks. Some overlapping occurred in south Senegal during the joint survey, but this would account for less than 5% overestimation. During the joint survey the approximately same shelf area was covered by the last Fridtjof Nansen survey as by the Capricorne and Cornide de Saavedra. The magnitude of the disparity between the estimates could indicate that systematic differences either in calibration or in survey methods also have occurred.
The general migration pattern and distribution of the dominating species has been described by other authors (FAO/CECAF 1979, Troadec and Garcia 1980, Garcia, 1982). In the following we present the distribution pattern found during three surveys with R/V “Dr. Fridtjof Nansen” (four surveys off Mauritania) for the species Sardinella aurita, Sardinella maderensis and Engraulis encrasicolus classified as the “Pelagic 1” group in the estimates and Trachurus trachurus, Trachurus trecae and Decapterus rhonchus classified as the “Pelagic 2” group.
Abundance estimates will also be presented for each species, but these must be considered very tentative as the survey methods and sampling gear used do not allow a reliable separation at the species level. Two species can be fairly easily separated from the group estimates, the anchovy (Engraulis encrasicolus) and the Atlantic horsemackerel (Trachurus trachurus), as these species are quite restricted both in distribution and abundance in the area from Cape Blanc to Sherbro Island.
The lack of information of species composition off Western Sahara constitutes a serious limitation of the data collected. Only estimates of total fish abundance are given, as no trawling could be carried out in the area. This reduces the reliability of estimates of stock level for some species and invalidates them for others. The total biomass off Western Sahara is estimated in the order of 2 million tonnes (Table 3) and represents 27% of the total abundance in the area between Cape Bojador and Sherbro Island.
During the preparation of this report it was not possible to find data quantifying the species composition off Western Sahara. Although the composition undoubtedly varies both seasonally and between years, some information can be extracted from the known distribution and migration pattern of the common species (op.cit.). Trachurus trachurus and Scomber scombrus seem to be the dominant species in the area. Other important species are Sardinella aurita, Sardina pilchardus and Trachurus trecae, but their centers of distribution are outside Western Sahara waters. Sardinella maderensis is also present, but of lesser importance.
Since the species composition off Western Sahara was unknown at the time of the surveys, the stock abundance estimates given below are restricted to resources south of Cape Blanc, and only tentative references are given for the stocks further north.
Figure 34 gives the distribution of the Pelagic 1 group during three surveys, and the estimated densities are given in three levels. The estimated biomasses by country can be summarized as follows (thousand tonnes and the country's share as percentage of total):
| May-Jun-81 | Sept-81 | Feb-Mar-82 | |
|---|---|---|---|
| Mauritania | 60 (12%) | 100 (20%) | 50 (24%) |
| Senegambia | 210 (41%) | 380 (76%) | 40 (20%) |
| Guinea Bissau | 90 (18%) | 20 (4%) | 70 (34%) |
| Guinea | 70 (14%) | no survey | 30 (15%) |
| Sierra Leone | 80 (16%) | no survey | 15 (7%) |
| Total | 510 (100%) | 500 (100%) | 205 (100%) |
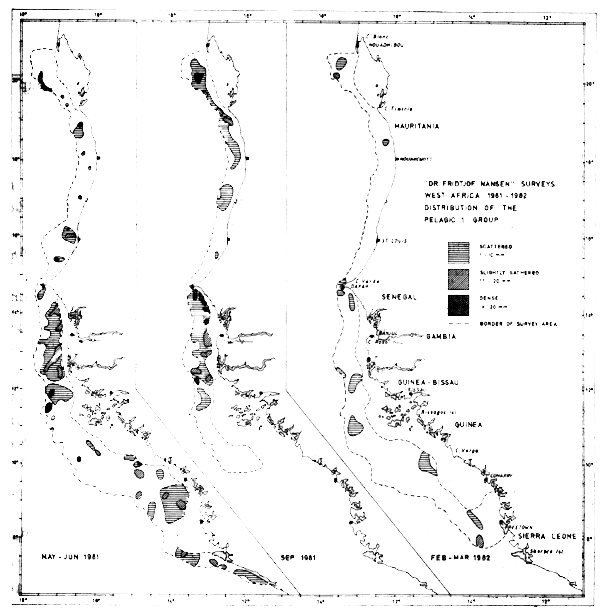
Fig. 34 Distribution of the Pelagic 1 group from three surveys between Cape Blanc and Sherbro Island, 1981–1982.
No corrections for the shallow unsurveyed areas have been included in these estimates. Due to its nearshore distribution the flat sardinella (Sardinella maderensis) is believed to be significantly underestimated. The population of round sardinella (S. aurita) is, on the other hand, assumed to be almost fully encompassed within the depth range surveyed, but the species has an large important stock component north of Cape Blanc, and it is uncertain whether a separate stock exists in this area or migrations into Mauritanian waters occur (Troadec & Garcia, 1980). The anchovy (Engraulis encrasicolus) is mostly found in some few dense patches along the coast and forms only a fraction of the estimated biomass of clupeoids. The given biomass estimates by country show that the main part of the Pelagic 1 group is located south of Mauritania during all three surveys.
There seems to be a permanent presence of sardinellas between Cape Blanc and Cape Timiris and between Cape Verde and the Bissagos Islands. Further south, between Cape Verga and Sherbro Island there are apparently also permanent concentrations of sardinellas, although this has only been found in two coverages. Between Cape Timiris and Cape Verde there are considerable concentrations of the Pelagic 1 group between June and September 1981 whereas the fish are practically absent in Mar-82. The shelf between the Bissagos Islands and Cape Verga seems to be of lesser importance for the distribution of the Pelagic 1 group.
Even if it is accepted that these 1981–82 assessments represent underestimates, they indicate that the stocks of sardinellas in this northern subtropical zone have declined from the mid-1970 level of about 1 million tonnes. The estimates from Feb–Mar-82 are striking, indicating that the sardinellas have been reduced to less than 50% of the level of the previous year. As this finding is from one survey only, it is possible that parts of the biomass have moved into the shallow waters not surveyed by the vessel. As mentioned, the flat sardinellas (S. maderensis) have their main distribution in the more shallow waters of the shelf, and it is possible that they have a seasonal migration perpendicular to the coast. This could explain part of the reduction in the estimates.
Another possibility is that portions of the sardinella stocks have migrated northwards off Western Sahara and there account for some of the increase in biomass in these waters during the February–March survey (Table 3). However this would be contrary to the seasonal migration pattern.
Comparisons with fishing data may show whether the decline is real or not.
Figure 36 and 37 later in this report show the distribution of S. aurita and S. maderensis based on the trawl catches only. It is also evident from these figures that the distributions of both species were drastically reduced in Feb–Mar-82, compared with the two earlier surveys.
In the following we will discuss results relevant to the species of the Pelagic 1 group.
Engraulis encrasicolus
Anchovies (Figure 35) were caught at scattered locations all over the survey area in May–Jun-81, while in Sept-81 and Feb– Mar-82 they were only found in three locations. The species seems to be permanently present in the Cape Blanc-Cape Timiris area and off Cape Roxo, having been found there during all three surveys. This is also the area where the best catches were obtained.
The total lengths measured from samples range from 4.0 to 14.5 cm (Annex III). No clear spawning season is indicated from the samples, as small specimens occur during all three surveys, but clear nursery areas for juvenile fish can be spotted off Cape Roxo and off Freetown, both in Feb-82. The mean length from all samples is 9 cm, a length that is also representative for the May–June and September samples. The mean for the February-March survey is 8 cm. No size-dependent distribution along the coast can be detected (Figure 35). The catches range from incidental specimens up to 12 tonnes/hour (south of Cape Roxo in Sept.-81).
Separate estimates of the abundance of anchovy are hindered by its frequent occurrence together with both sardines and horsemackerel. Sometimes, the anchovies formed very dense schools during daytime and could easily be caught in large quantities with midwater trawl, in contrast to the sardinellas and horsemackerels. Thus, the anchovies are clearly overrepresented in the tables of catch distribution.
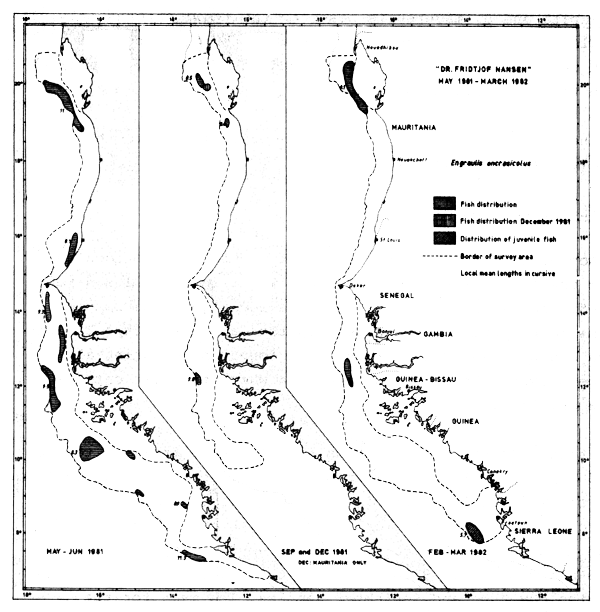
Fig. 35 Distribution of Engraulis encrasicolus from occurrence in trawl catches, Cape Blanc to Sherbro Island, 1981–1982.
In most locations, however, the catches were less than 100 kg/ hour. Catches exceeding 1 tonne/hour can all be allocated to around Cape Timiris and around Cape Roxo. Apart from these areas the anchovy is assumed to form only a small fraction of the Pelagic 1 group.
Separate acoustic estimates of dense concentrations of anchovy give figures of from 10–30 thousand tonnes. In May-81 these concentrations constituted 90 thousand tonnes of the Pelagic 1 group and in Sept-81 the corresponding figure was 45 thousand tonnes. In Feb–Mar-82 no really dense concentrations were located. As these concentrations were quite restricted in their distribution they could easily have been missed by the vessel's sampling track. There may also be seasonal variations in stock size. Taking into account the scattered occurrences of the anchovywe somewhat speculatively suggest the biomass of Engraulis encrasicolus to be in the order of 150 thousand tonnes, with perhaps an additional 50 thousand tonnes for the shallow unsurveyed areas.
Sardinella aurita
The round sardinella occurred along the coast in the whole survey area during the two first surveys (Figure 36). During the last survey, in Feb–Mar-82, the species was found only in some few trawl samples along the coast, and then mostly at catch rates of 2 kg/hour or less. Only off Bissagos Islands were some concentrations of significance located, and further southwards no recordings of Sardinella aurita were made. The species seems to be permanently present in the Cape Blanc-Cape Timiris area, and off Cape Roxo (Figure 36).
The total lengths measured from the samples range from 6 to 39.5 cm. Plots have been made to map the distribution of specimens of length < 10 cm in order to trace any restricted nursery areas. In May-June young fish were found in five locations from the Casamanche River Delta south to the Guinea Bissau-Guinea border. In September young fish were spotted only in the north, in the Cape Blanc-Cape Timiris area, while in Feb-Mar-82 no small sardinellas were caught.
Likewise, the distribution of fish of 30 cm length or more have been plotted in an attempt to trace any migration of the adult stock. The old fish were located between St. Louis and Gambia River in May–June, north between Cape Timiris and Nouakchott in September, and in Feb–Mar-82 at two widely separated locations, off Cape Timiris and off Bissagos Islands.
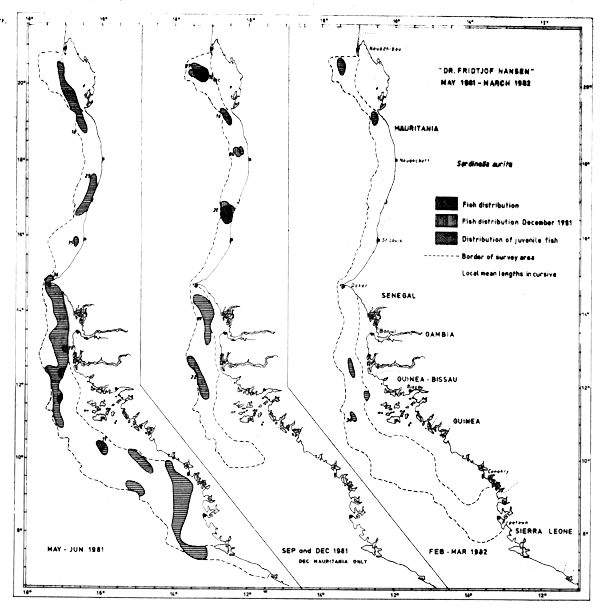
Fig. 36 Distribution of Sardinella aurita from occurrence in trawl catches, Cape Blanc to Sherbro Island, 1981–1982.
If one assumes that the old fish off Cape Timiris in Feb-Mar-82 belong to a Saharan stock at its southern part of the distribution, the findings both for young and old fish correspond well with the general migration and spawning pattern already known and summarized by Garcia (1982).
The catch figures (see Annex V) do not reflect the density and abundance of sardinellas, as the trawling speed is clearly below optimum for the rather fast swimming adult sardinellas. Only two catches exceeded 1 tonne/hour, both off Mauritania in September.
Generally the catchability of the trawl gears is highly species dependent, and the distribution of species in the catch may thus not reflect the composition of species in situ. Since the two sardinella species are quite similar both in shape and size, it seems reasonable to assume that they do not differ substantially in catchability. We have therefore used the catch records to calculate these two species' relative abundance in the various countries. Weighted indices of abundance are produced by counting all stations where the species are present and giving each station a weight in accordance with the size of the catch and the following scale: <1 kg/hour = 0, 1–10 kg/hour = 1, 10–100 kg/hour = 2, 0.1–1 tonne/hour = 3, >1 tonne/hour = 4. This scale was chosen in an attempt to take account of the irregular distribution of pelagic fish. The species' relative importance is calculated in percent. The results from these calculations are given in Table 9, and are applied to the acoustic estimates to give tentative and very rough figures of the abundance of the species.
| Country and survey: | Sardinella aurita | Sardinella maderensis | ||||
|---|---|---|---|---|---|---|
| O | W | % | O | W | % | |
| Mauritania, May -81 | 6 | 8 | 89 | 1 | 1 | 11 |
| Sept -81 | 11 | 22 | 73 | 5 | 8 | 27 |
| Dec -81 | 7 | 7 | 47 | 6 | 8 | 53 |
| Mar -82 | 4 | 3 | 75 | 2 | 1 | 25 |
| Senegambia, May -81 | 22 | 24 | 49 | 15 | 25 | 51 |
| Sept -81 | 10 | 19 | 35 | 21 | 35 | 65 |
| Feb/Mar -82 | 1 | 0 | 0 | 2 | 3 | 100 |
| Guinea Bissau, May/June -81 | 8 | 6 | 35 | 7 | 11 | 65 |
| Sept -81 | 1 | 2 | 33 | 2 | 4 | 67 |
| Feb -82 | 3 | 5 | 71 | 3 | 2 | 29 |
| Guinea, June -81 | 9 | 9 | 100 | 2 | 0 | - |
| Feb -82 | - | - | - | 1 | 1 | 100 |
| Sierra Leone, June -81 | 5 | 6 | 38 | 6 | 10 | 62 |
| Feb -82 | - | - | - | 3 | 6 | 100 |
By subtracting the estimates of anchovy from the ones for the Pelagic 1 group, and applying the figures on the relative importance of the two Sardinella species one arrives at the following very rough figures of the abundance of Sardinella aurita (thousand tonnes):
| May-Jun-81 | Sept-81 | Dec-81 | Feb-Mar-82 | |
|---|---|---|---|---|
| Mauritania | 20 | 55 | 4 | 40 |
| Senegambia | 105 | 130 | n.s. | - |
| Guinea Bissau | 15 | 5 | n.s. | 50 |
| Guinea | 70 | n.s. | n.s. | - |
| Sierra Leone | 30 | n.s. | n.s. | - |
| Total | 240 | (180) | 90 |
These estimates are believed to encompass most of the Sardinella aurita within the region surveyed, but leave open possible resources north of Mauritania.
Sardinella maderensis
The distribution of the flat sardinella by its occurrence in the trawl catches is shown in Figure 37. Three areas appeared to be of special importance during all coverages; the Cape Blanc-Cape Timiris area, the Cape Verde-Bissagos Islands area, and around Sherbro Island. Between Cape Timiris and Cape Verde the fish was found in several locations in September, while it was virtually absent from there in the May-June and FebruaryMarch surveys.
Juvenile fish of lengths 10 cm or less were localized off Casamance (south Senegal) and southward from Sherbro Island in May. In September juveniles were found between Cape Blanc and Cape Timiris and off St. Louis, i.e. only in Mauritania, and in Feb-Mar-82 they were caught only in the southern part of the area, off Freetown.
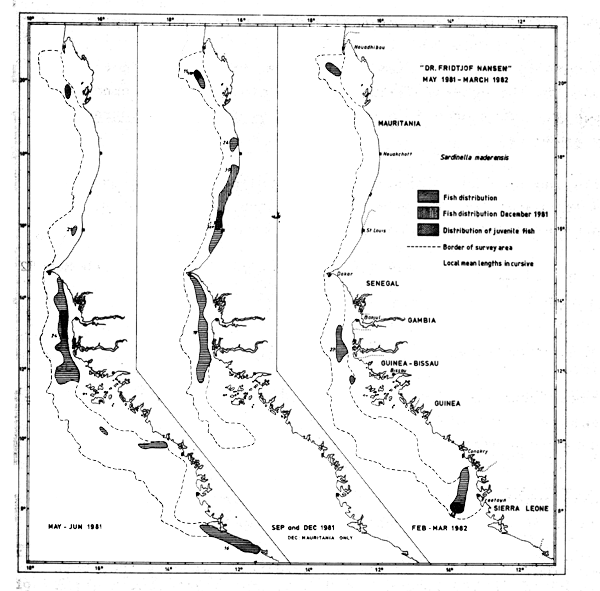
Fig. 37 Distribution of Sardinella maderensis from occurrence in trawl catches, Cape Blanc to Sherbro Island, 1981–1982.
Old fish with lengths of 30 cm or more were found in May-June only between Cape Verde and Bissagos Islands, in September only at three locations in Mauritania, and not at all during the February-March survey.
According to previous reports the species forms three substocks in the area, and no wide range migration is known (FAO/CECAF, 1979, Troadec & Garcia, 1980, Garcia, 1982). During the “Dr. Fridtjof Nansen” surveys the length frequency of S. maderensis in the important shallow areas is virtually unknown, and it is therefore hard to deduce any migration pattern from the samples.
The nursery area localized off Freetown in Feb-82 is not included in the general features on the biology of the species (op.cit.). This ground is shared with S. aurita and Engraulis encrasicolus.
Employing the same procedure as for Sardinella aurita one gets the following very rough estimates of the abundance of S. maderensis (thousand tonnes):
| May-Jun-81 | Sept-81 | Dec-81 | Feb-Mar-82 | |
|---|---|---|---|---|
| Mauritania | 2 | 20 | 4 | 10 |
| Senegambia | 100 | 230 | n.s. | 40 |
| Guinea Bissau | 30 | 15 | n.s. | 20 |
| Guinea | - | n.s. | n.s. | 30 |
| Sierra Leone | 50 | n.s. | n.s. | 15 |
| Total | (180) | (265) | 115 |
These figures are believed to represent serious underestimates because of the species' quite shallow distribution.
Comparing the totals for the two sardinella species they both seem to be in the order of 200 thousand tonnes during the two first surveys, but reduced to 100 thousand tonnes in the last survey. If one allows that 1/3 of the S. maderensis stock were in the shallow waters one gets estimates for the sum of the two species in the range of 500 thousand tonnes during summer/autumn 1981, with a possible reduction to 250 thousand tonnes in early 1982.
This group comprises the four species Trachurus trachurus, T. trecae, Decapterus rhonchus and D. punctatus, and Figure 38 gives the groups' distribution and estimated density in three levels during the three main surveys. Extracted from Table 3 the estimated biomasses by countries are (thousand tonnes):
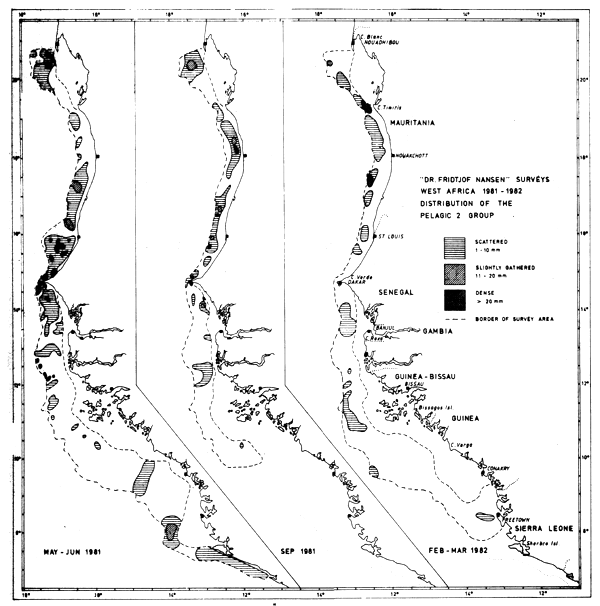
Fig. 38 Distribution of the Pelagic 2 group from three surveys between Cape Blanc and Sherbro Island, 1981–1982.
| May-Jun-81 | Sept-81 | Dec-81 | Feb-Mar-82 | |
|---|---|---|---|---|
| Mauritania | 370 | 190 | 470 | 350 |
| Senegambia | 570 | 90 | n.s. | 90 |
| Guinea Bissau | 40 | 20 | n.s. | 400 |
| Guinea | 30 | n.s. | n.s. | 10 |
| Sierra Leone | 120 | n.s. | n.s. | 5 |
| Total | 1130 | 300 | n.s. | 975 |
All the species concerned here may also have parts of their distribution in the shallow areas not surveyed, but we have not attempted to give any estimate for these areas, except the ones given in connection with total fish abundance (see 2.1). Decapterus punctatus is perhaps the most severely underestimated as good registrations and catches were obtained close to the near-shore limit of the survey area at several locations. The other three species have generally their highest abundance over the deep part of the shelf (Boely and Freon, 1980).
The total stock size of these species, perhaps with the exception of Decapterus rhonchus, are believed to be underestimated as they are also distributed outside the region surveyed. On the basis of the general distribution and migration pattern (Garcia, 1982) we assume that Trachurus trachurus had most of its biomass north of Cape Blanc during all surveys, and T. trecae probably also had a significant number in this area, especially during the September survey. Decapterus punctatus made up most of the Pelagic 2 group in Guinea and Sierra Leone during the May-June survey and the species had probably then been covered to its southern limit (off Liberia). In Feb-82 however, the southern survey limit was outside Freetown, and it is possible that D. punctatus is distributed further southwards during this period.
North of Guinea the estimates given are almost entirely comprised of the three species Trachurus trachurus, T. trecae and Decapterus rhonchus, commonly known as horsemackerels. The total estimates for these fluctuates from about 1 million tonnes in May–June to 300 thousand tonnes in September to about 750 thousand tonnes in Feb-Mar-82. The low values in September can generally be explained by a northward distribution during this period, with large portions of the biomass north of the survey area.
However, one should note that even during the February-March survey still close to 40% of the biomass of horsemackerels was located in Mauritanian waters which may indicate that considerable quantities of carangids are distributed still further north. This is a possibility supported by the high estimates of total fish biomass found off Western Sahara in March 1982. The 250 thousand tonnes reduction between the May-June and the February-March survey thus does not necessarily indicate any actual reduction in the stocks, but the possibility should be kept in mind.
By the same methods as described for the Sardinella species, we have made attempts to split the estimates on horsemackerels to the species level, using the catch records and assuming that the catchability is roughly the same for all three species. Table 10 gives indices of presence and relative abundance on the basis of the catch data, and very tentative abundance estimates are given below where the Pelagic 2 group are discussed species by species.
| Country and survey: | Trachurus trachurus | Trachurus trecae | Decapterus rhonchus | Decapterus punctatus | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| O | W | % | O | W | % | O | W | % | O | W | % | |
| Mauritania, May -81 | 8 | 6 | 12 | 18 | 38 | 74 | 3 | 7 | 14 | - | - | - |
| Sept -81 | - | - | - | 11 | 26 | 48 | 13 | 28 | 52 | - | - | - |
| Dec -81 | 4 | 8 | 13 | 8 | 20 | 33 | 16 | 32 | 54 | - | - | - |
| Mar -82 | 15 | 18 | 37 | 17 | 21 | 43 | 7 | 10 | 20 | - | - | - |
| Senegambia, May -81 | 3 | 2 | 2 | 24 | 45 | 56 | 18 | 24 | 42 | - | - | - |
| Sept -81 | - | - | - | 2 | 7 | 25 | 9 | 21 | 75 | 2 | 0 | 0 |
| Feb/Mar -82 | 1 | 0 | 0 | 13 | 23 | 100 | 1 | 0 | 0 | - | - | - |
| Guinea Bissau, | ||||||||||||
| May/June -81 | - | - | - | 5 | 6 | 35 | 8 | 11 | 65 | - | - | - |
| Sept -81 | - | - | - | - | - | - | - | - | - | - | - | - |
| Feb -82 | - | - | - | 5 | 12 | 80 | 2 | 3 | 20 | - | - | - |
| Guinea, June -81 | - | - | - | 3 | 3 | 50 | 3 | 3 | 50 | |||
| Feb -82 | - | - | - | 1 | 0 | 0 | - | - | - | - | - | - |
| Sierra Leone, | ||||||||||||
| June -81 | - | - | - | - | - | - | - | - | - | 4 | 7 | 100 |
| Feb -82 | - | - | - | - | - | - | 1 | 0 | 0 | 2 | 0 | 0 |
Trachurus trachurus
The information gathered on Trachurus trachurus is inadequate as the surveys have only covered the southern part of the distribution area. In May, the species was found south to Cape Verde, in September it was absent from the survey area, and in Dec-81 and Mar-82 it occurred in various areas along the coast down to St. Louis (Fig. 39).
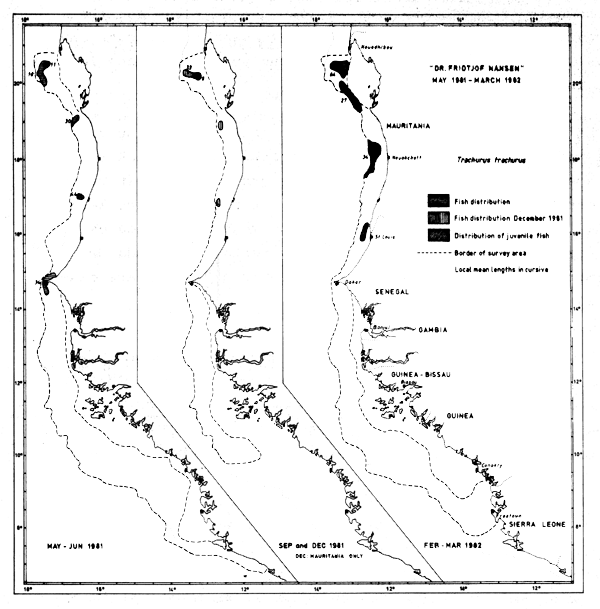
Fig. 39 Distribution of Trachurus trachurus from occurrence in trawl catches, Cape Blanc to Sherbro Island, 1981–1982.
The old fish of lengths 30 cm or more do not form outstanding aggregations or any clearly restricted distributions, and the group is always present in the southernmost samples of the species.
Juveniles of lengths < 10 cm were present south to St. Louis in May-81 and Mar-82, while they were only found a little south of Cape Blanc in Dec-81. Particularly samples from Mar-82 indicate a major (post-) spawning period by their distribution and size of juveniles. No juveniles were found in Senegalese waters.
The catches were never quantitatively substantial. The fish were caught on 33 occasions and on 21 of these the catch was less than 10 kg/hour. 7 catches exceeded 50 kg/hour, and only one was more than 200 kg/hour.
By the very rough methods already described we have estimated the biomass by country as follows (thousand tonnes):
| May-81 | Sept-81 | Dec-81 | Feb-Mar-82 | |
|---|---|---|---|---|
| Mauritania | 45 | - | 60 | 170 |
| Senegal (north) | 10 | - | no survey | 0 |
| Total | 60 | - | 60 | 170 |
Trachurus trecae
The distribution of Trachurus trecae, by its presence in the trawl catches, is shown in Figure 40. In May-81 and Feb-82 the species was located south to the Bissagos Islands while in Sept-81 the southern limit was around St. Louis. In December of the same year only Mauritania was surveyed and the fish were distributed at least as far south as in September.
The old fish of lengths > 30 cm were located around the Bissagos Islands in Feb-82 and formed probably the head of the southward migration. In May-81 the corresponding southern limit was Cape Verde and, in accordance with the general migration picture, the fish were probably moving north. In September a few old fish were located near Nouakchott and the majority thus probably being distributed north of Cape Blanc.
Juveniles of lengths less than 10 cm were located between St. Louis and Bissagos Islands in May, between Nouachott and Cape Blanc in September and between Cape Timiris and Cape Roxo in Feb-Mar-82.
The species was caught at 106 stations during the entire programme. 41% of the catches were less than 10 kg/hour, and 32% of the hauls exceeded 200 kg/hour. Catches exceeding 1 t/hr were obtained only on 6 occasions, but the catch figures are in no way representative of a commercial fishery as the trawling speed and the gear size was clearly below optimum for this fast swimming pelagic species.
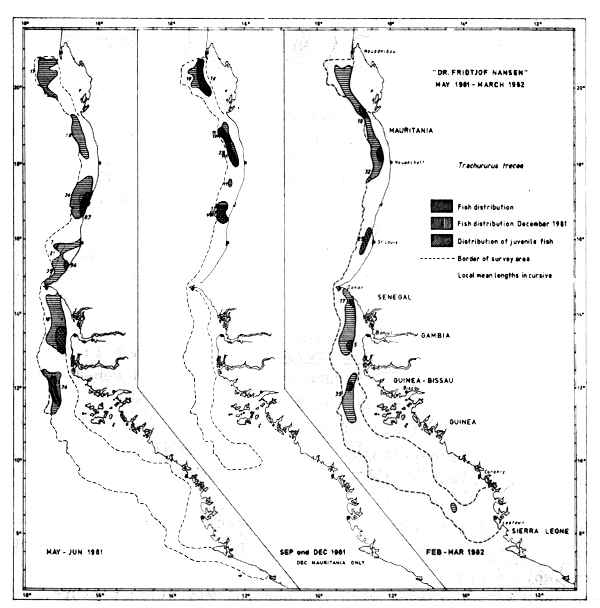
Fig. 40 Distribution of Trachurus trecae from occurrence in trawl catches, Cape Blanc to Sherbro Island, 1981–1982.
The abundance of Trachurus trecae are roughly estimated as follows (thousand tonnes):
| May-Jun-81 | Sept-81 | Dec-81 | Feb-Mar-82 | |
|---|---|---|---|---|
| Mauritania | 275 | 90 | 150 | 200 |
| Senegambia | 320 | 20 | n.s. | 90 |
| Guinea Bissau | 15 | - | n.s. | 150 |
| Guinea | - | n.s. | n.s. | - |
| Sierra Leone | - | n.s. | n.s. | - |
| Total | 610 | 110 | 440 |
The total estimates vary from about 600 thousand to 100 thousand tonnes during the programme. The major reduction in the estimates from May-June to September can largely be ascribed to migration north of Cape Blanc.
One should also keep in mind that even in February-March nearly 50% of the total biomass of the carangids is allocated to Mauritania, although the general migration picture states the species to be in their southern part of the migration cycle. This makes it possible that T. trecae are still distributed further northwards, off Western Sahara. The high total fish abundance off Western Sahara at this time may support this. Due to the uncertainties of species composition off Western Sahara the reduction of 170 thousand tonnes in T. trecae from spring -81 to early -82 does not give us a firm reason to believe in an actual reduction of the stock.
Decapterus rhonchus
The yellow horsemackerel appears on the Arguin Bank year round, but apart from this the distribution of the species on basis of the trawl catches (Figure 41) suggests a seasonal migration along the coast. In May-Jun-81 the species was found mostly south of St. Louis, while in September and December same year a widespread distribution occurred northwards up to Cape Blanc. In September the southern limit of the distribution was off the Gambia, while in Feb-Mar-82 the fish were found as far south as the Bissagos Islands. Here big fish of mean length 36 cm occurred together with Trachurus trecae and Sardinella aurita, possibly forming the frontier of a southward migration. We find it remarkable that Decapterus rhonchus was not located in Senegal at this time, and that the main part of the biomass was still in Mauritania.
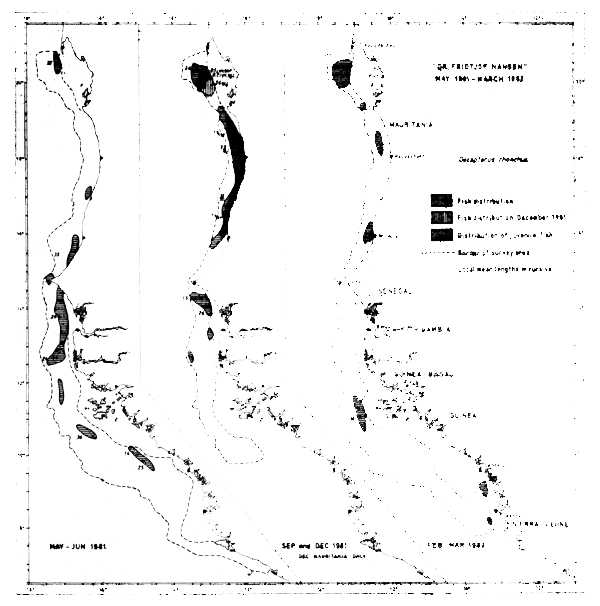
Fig. 41 Distribution of Decapterus rhoncus from occurrence in trawl catches, Cape Blanc to Sherbro Island, 1981–1982.
The yellow horsemackerel was also located south of the Bissagos Islands in May-81 and in Feb-82. This fish probably forms an independent southern population already identified by Soviet scientists (FAO/CECAF, 1979).
Juveniles of 10 cm length or less were located between Cape Verde and Cape Blanc in September and on the Arguin Bank in December. In May-Jun-81 and Feb-Mar-82 no juveniles were caught north of Sierra Leone, although in the February-March survey juveniles were located off Freetown along with other species. This seems to be an important nursery ground for the southern area of the investigation.
The species was caught on 86 occasions during the surveys, but in 37% of the hauls the catch was less than 10 kg/hour. Six hauls or 7% exceeded 500 kg/hour. The highest catch was 9800 kg/ hour, off St. Louis in Sept-81.
Stressing more the order of magnitude than absolute abundances, the following biomass estimates are given for Decapterus rhonchus (thousand tonnes):
| May-Jun-81 | Sept-81 | Dec-81 | Feb-Mar-82 | |
|---|---|---|---|---|
| Mauritania | 50 | 100 | 250 | 100 |
| Senegambia | 240 | 70 | n.s. | - |
| Guinea Bissau | 25 | 20 | n.s. | 40 |
| Guinea | 15 | n.s. | n.s. | 10 |
| Sierra Leone | - | n.s. | n.s. | - |
| Total | 330 | 190 | 150 |
Only small fractions of the stock are generally assumed to be distributed north of Cape Blanc (Troadec & Garcia, 1980). The surveys with “Dr. Fridtjof Nansen” are therefore believed to have encompassed most of the stock, at least of the important northern population. If, as we believe, great parts of the stock did not seasonally move into the very shallow waters unsurveyed by the vessel, the given biomass estimates indicate a reduction of the population in Feb-Mar-82 to near half of the level of May the previous year.
Decapterus punctatus
In Jun-81 this species constituted the majority of the carangids found in Sierra Leone, and a portion of the biomass in the shallow waters of Guinea. In Feb-82, the last survey carried out in the area, the species was identified by trawl only in two locations in Sierra Leone. During this coverage major aggregates of fish close to the bottom were located in shallow waters just south of the Bissagos Islands. Because of very coarse bottom conditions it was not possible to trawl on the registrations, but the characteristics of the traces are clearly that of one of the pelagic species, most likely D. punctatus.
Based on the few data and the methods already described, we have estimated the biomass of the species as follows: (thousand tonnes)
| Jun-81 | Feb-82 | |
|---|---|---|
| Guinea Bissau | - | 210 |
| Guinea | 15 | - |
| Sierra Leone | 120 | 5 |
The southern limit of the latest survey was off Freetown, so it is possible that part of the population is distributed further south. The species occurred mainly in quite shallow waters and there formed dense schools detected by the sonar. As the species is assumed to have a large fraction of the population in shallow waters, the above given estimates do in no way indicate the size of the stock, but show only the fraction available for off-shore fishing.
The catches were never substantial. Of 25 catches 17 were less than 10 kg/hour and the best were two catches just under 200 kg/ hour. Purse-seining the schools in the shallow waters would probably give considerably higher catches.
Extracted from Table 3 the biomass estimates on the “Other Fish” group are as follows (thousand tonnes):
| May–Jun-81 | Sept-81 | Feb–Mar-82 | |
|---|---|---|---|
| Mauritania | 30 | 70 | 260 |
| Senegambia | 60 | 60 | 430 |
| Guinea Bissau | 140 | 80 | 125 |
| Guinea | 30 | n.s. | 10 |
| Sierra Leone | 20 | n.s. | 40 |
| Total | 280 | 210 | 865 |
The estimates from the May–June and the September survey seem to be in fair agreement, while the February–March estimates are substantially higher in Mauritania and especially in Senegambia. During the last survey about 50% of the biomass of “Other Fish” was found in dense aggregations in shallow waters off Mauritania and Senegambia, while the two first coverages were characterized by a much more even distribution. The patches consisted mostly of Pomadasyidae (mainly Brachydeuterus auritus), Sciaenidae, Sparidae and Trichiuridae (Trichiurus lepturus). This considerable increase in biomass suggests that a migration had taken place before the last survey from the shallows into the shallow waters covered by the vessel. Such a migration perpendicular to the coast could be a reaction to the cold and oxygen-depleted water replacing the nearshore waters during upwelling.
The surveys carried out with “Dr. Fridtjof Nansen” off Western Africa are essentially acoustic both in purpose and design. Bottom trawling was then carried out only when fish were somewhat aggregated and formed notable contributions to the echointegrator, contributions that needed to be identified. Schooling species or other species with an aggregated distribution pattern are consequently oversampled in relation to species with a more solitary behaviour and dispersed distribution. This should be kept in mind when making use of the trawl data for biological studies.
Several families made notable contributions to the catches, namely the seabreams (Sparidae), the grunts (Pomadasyidae), hairtails (Trichiuridae), croakers (Sciaenidae), catfish (Ariidae) and threadfins (Polynemidae). Of the families Carangidae and Clupeidae species other than the ones dealt with in the foregoing chapter made important contributions to the catches. To be mentioned are Alectis alexandrinus, Caranx chrysos, C. senegalus, Chloroscombrus chrysurus and Selene dorsalis among the carangids and Ilisha africana among the clupeoids. For further information on these and other species we refer to the Annexes V and IV where catch data and length distributions are presented by species. Included in Annex VII are also maps of all fishing stations for each coverage. The occurrence of a certain species can then be illustrated by marking the stations where the species was caught. By preparing the data this way we hope to have facilitated further specialized study.
Of the many species belonging to the “Other Fish” group, we present distribution charts for two species commonly occurring in the catches; the bigeye grunt (Brachydeuterus auritus) and the hairtail (Trichiurus lepturus). The triggerfish (Balistes capriscus) was treated separately during all surveys and the findings are covered in a separate chapter in this report.
Brachydeuterus auritus
The distribution of the bigeye grunt during the surveys is shown in Figure 42. The species is found along the entire region from Cape Blanc to Sherbro Island, but the distribution pattern seems to be seasonally dependent. In the winter (FebMar-82) and spring (May-Jun-81) the species seems to be mainly restricted to south of Mauritania while in September and December it is widespread in Mauritanian waters up to Cape Timiris. Especially in Dec-81 the fish was spread abundantly between Cape Timiris and St. Louis. The findings coincide with the general migration picture already known (Garcia 1982).
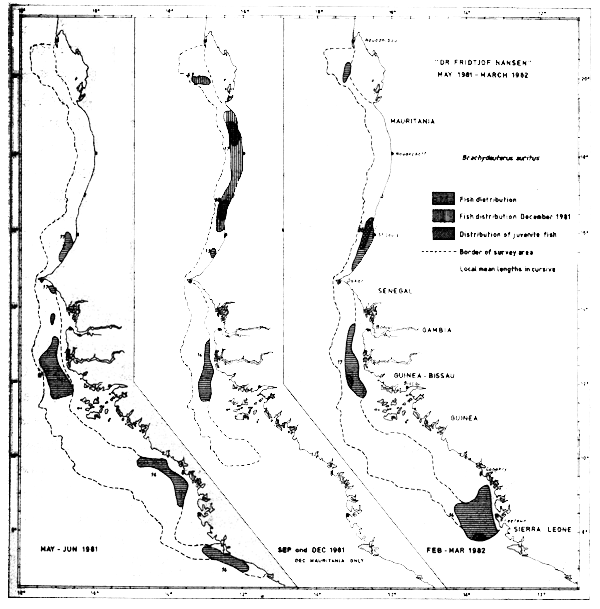
Fig. 42 Distribution of Brachydeuterus auritus from occurrence in trawl catches, Cape Blanc to Sherbro Island, 1981–1982.
Juveniles were caught in Feb-Mar-82 off St. Louis, Cape Roxo and Freetown, and in December between Cape Timiris and St. Louis (southern limit of the December survey).
The species was found in 93 hauls during surveys in the region and of these 8 hauls exceeded 1 tonne/hour. The big catches, except one, were all caught between Cape Verde and Nouachott (between 15°N and 17°N specifically).
Trichiurus lepturus
The distribution of the largehead hairtail by its occurrence in the trawl catches is shown in Figure 43. In May-81 the species seems to form two major distribution clusters in the region north of the Bissagos Islands, one off Senegal, north of Cape Verde and one in the waters off Cape Roxo. In September the species was found mostly north of Cape Verde and up to Cape Blanc, while in Feb-Mar-82 the fish was again concentrated in North Senegal and off Cape Roxo, and also in a cluster on the Arguin Bank (Cape Blanc area). The hairtails were not located between Bissagos Islands and Freetown during the last survey.
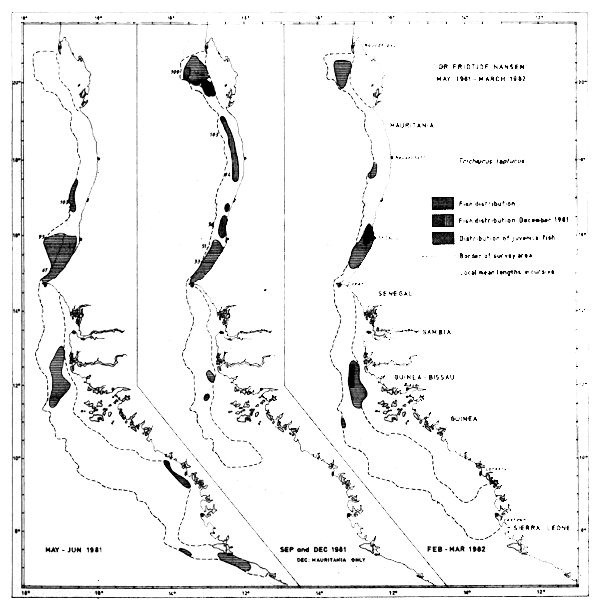
Fig. 43 Distribution of Trichiurus lepturus from occurrence in trawl catches, Cape Blanc to Sherbro Island, 1981–1982.
Juveniles were caught in Feb-Mar-82 off St. Louis and off Cape Roxo and in December on the Arguin Bank.
The species was caught in 90 hauls, generally in small quantities and mixed with other fish. The two best catches were around 300–400 kg/hour and only seven other hauls exceeded 100 kg/hour.
![]()
![]()
![]()